Симбиогенез
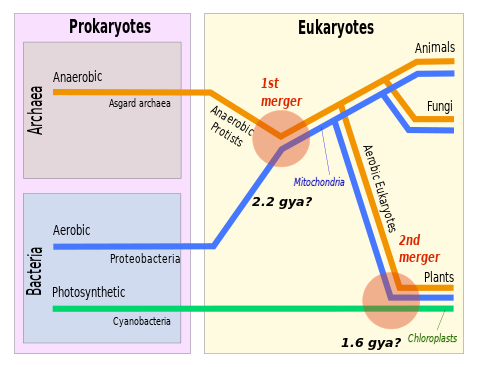
Симбиогенез ( эндосимбиотическая теория , или серийная эндосимбиотическая теория [2] ) — ведущая эволюционная теория происхождения эукариотических клеток от прокариотических организмов. [3] Теория утверждает, что митохондрии , пластиды , такие как хлоропласты , и, возможно, другие органеллы эукариотических клеток произошли от ранее свободноживущих прокариот (более тесно связанных с бактериями , чем с археями ), взятых друг в друга в эндосимбиозе . Митохондрии, по-видимому, филогенетически связаны с бактериями Rickettsiales , тогда как хлоропласты, как полагают, связаны с цианобактериями .
Идея о том, что хлоропласты изначально были независимыми организмами, которые вступали в симбиотические отношения с другими одноклеточными организмами, восходит к 19 веку, когда ее поддержали такие исследователи, как Андреас Шимпер . Эндосимбиотическая теория была сформулирована в 1905 и 1910 годах русским ботаником Константином Мерешковским , а также развита и подтверждена микробиологическими данными Линн Маргулис в 1967 году.
Среди многих доказательств, подтверждающих симбиогенез, есть то, что митохондрии и пластиды содержат свои собственные хромосомы и размножаются путем деления на две части , параллельно, но отдельно от полового размножения остальной части клетки; что хромосомы некоторых митохондрий и пластид представляют собой одиночные кольцевые молекулы ДНК, подобные кольцевым хромосомам бактерий; что транспортные белки, называемые поринами , обнаружены во внешних мембранах митохондрий и хлоропластов, а также в мембранах бактериальных клеток; и что кардиолипин обнаруживается только во внутренней митохондриальной мембране и мембранах бактериальных клеток.
История
[ редактировать ]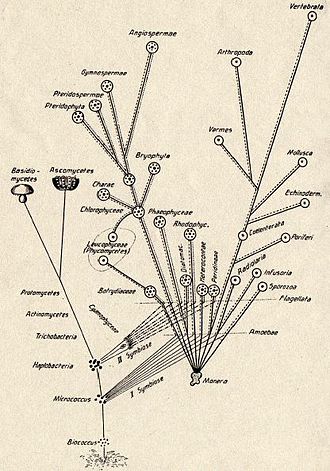
Русский греческого ботаник Константин Мерешковский впервые изложил теорию симбиогенеза (от «Природа и σύν syn «вместе», βίος bios «жизнь» и γένεσις генезис «происхождение, рождение») в своей работе 1905 года происхождение хроматофоров в растении» . королевстве , а затем развил ее в своей книге 1910 года «Теория двух плазм как основа симбиогенеза: новое исследование происхождения организмов» . [5] [6] [7] Мерешковский предположил, что сложные формы жизни возникли в результате двух эпизодов симбиогенеза: объединения симбиотических бактерий с последующим образованием ядер и хлоропластов . [4] Мерешковский знал о работах ботаника Андреаса Шимпера . В 1883 году Шимпер заметил, что деление хлоропластов зеленых растений очень похоже на деление свободноживущих цианобактерий . Шимпер предварительно предположил (в сноске), что зеленые растения возникли в результате симбиотического союза двух организмов. [8] В 1918 году французский учёный Поль Жюль Портье опубликовал книгу «Симбиоты» , в которой утверждал, что митохондрии возникли в результате процесса симбиоза. [9] [10] Иван Валлин отстаивал идею эндосимбиотического происхождения митохондрий в 1920-х годах. [11] [12] Русский ботаник Борис Козо-Полянский стал первым, кто объяснил теорию с точки зрения дарвиновской эволюции . [13] В своей книге « Новый принцип биологии» 1924 года. Очерк теории симбиогенеза , [14] он писал: «Теория симбиогенеза — это теория отбора, основанная на явлении симбиоза». [15]
Эти теории не получили распространения до тех пор, пока не были проведены более детальные электронно-микроскопические сравнения цианобактерий и хлоропластов, например, Хансом Рисом в 1961 и 1962 годах. [16] [17] В сочетании с открытием того, что пластиды и митохондрии содержат собственную ДНК, [18] привели к возрождению идеи симбиогенеза в 1960-х годах. Линн Маргулис выдвинула и обосновала эту теорию микробиологическими данными в статье 1967 года « О происхождении митозирующих клеток». [19] В своей работе 1981 года «Симбиоз в клеточной эволюции» она утверждала, что эукариотические клетки возникли как сообщества взаимодействующих объектов, включая эндосимбиотические спирохеты , которые развились в эукариотические жгутики и реснички . Эта последняя идея не получила большого признания, поскольку жгутики лишены ДНК и не имеют ультраструктурного сходства с бактериями или архей (см. Также: Эволюция жгутиков и прокариотического цитоскелета ). По мнению Маргулиса и Дориона Сагана , [20] «Жизнь захватила земной шар не в результате боя, а посредством взаимодействия» (т.е. путем сотрудничества). Кристиан де Дюв предположил, что пероксисомы , возможно, были первыми эндосимбионтами, позволявшими клеткам противостоять растущему количеству свободного молекулярного кислорода в атмосфере Земли. Однако сейчас оказывается, что пероксисомы могут образовываться de novo , что противоречит идее о их симбиотическом происхождении. [21] В настоящее время широкое признание получила фундаментальная теория симбиогенеза как происхождения митохондрий и хлоропластов. [3]
От эндосимбионтов к органеллам
[ редактировать ]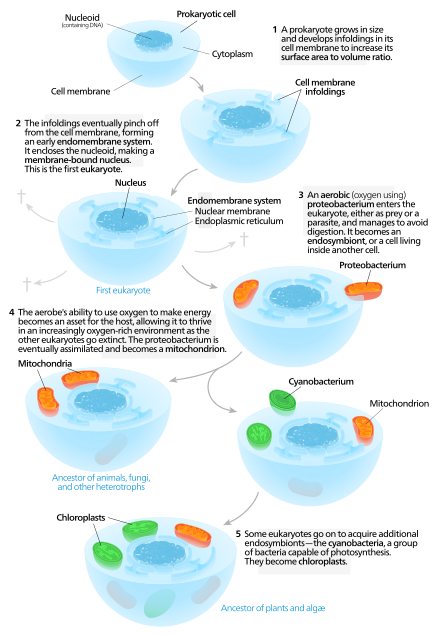
Биологи обычно отличают органеллы от эндосимбионтов – целых организмов, живущих внутри других организмов – по уменьшенному размеру генома . [23] Когда эндосимбионт эволюционирует в органеллу, большая часть его генов переносится в геном клетки-хозяина . [24] Поэтому клетка-хозяин и органелла должны разработать механизм транспорта, который позволит вернуть белковые продукты, необходимые органелле, но теперь производимые клеткой. [25]
Свободноживущие предки
[ редактировать ]Раньше считалось, что альфапротеобактерии являются свободноживущими организмами, наиболее тесно связанными с митохондриями. [25] Более поздние исследования показали, что митохондрии наиболее тесно связаны с бактериями Pelagibacterales , в частности с бактериями клады SAR11. [26] [27]
Азотфиксирующие нитчатые цианобактерии — свободноживущие организмы, наиболее близкие к пластидам. [25] [28] [29]
И цианобактерии, и альфапротеобактерии имеют большой (>6 МБ ) геном, кодирующий тысячи белков. [25] Пластиды и митохондрии демонстрируют резкое уменьшение размера генома по сравнению с их бактериальными родственниками. [25] Геномы хлоропластов фотосинтезирующих организмов обычно имеют размер 120–200 т.п.н. [30] кодирующий 20–200 белков [25] а митохондриальные геномы у человека имеют размер примерно 16 т.п.н. и кодируют 37 генов, 13 из которых являются белками. [31] используя пример пресноводного амебоида Однако, Paulinella chromatophora , который содержит хроматофоры, которые, как выяснилось, произошли от цианобактерий, Килинг и Арчибальд утверждают, что это не единственный возможный критерий; клетки другой заключается в том, что клетка-хозяин взяла на себя контроль над регуляцией деления бывшего эндосимбионта, тем самым синхронизируя его с собственным делением . [23] Новак и ее коллеги секвенировали ген хроматофора (1,02 Мб) и обнаружили, что только 867 белков кодируются этими фотосинтезирующими клетками. Сравнение с ближайшими к ним свободноживущими цианобактериями рода Synechococcus (с размером генома 3 Мб и 3300 генами) показало, что хроматофоры претерпели резкое сокращение генома. Хроматофоры содержали гены, отвечающие за фотосинтез , но не имели генов, которые могли бы выполнять другие биосинтетические функции; это наблюдение предполагает, что эти эндосимбиотические клетки сильно зависят от своих хозяев в плане механизмов выживания и роста. Таким образом, было обнаружено, что эти хроматофоры нефункциональны для специфичных для органелл целей по сравнению с митохондриями и пластидами. Это различие могло способствовать ранней эволюции фотосинтетических органелл. [32]
Утрата генетической автономии, то есть потеря многих генов эндосимбионтов, произошла на очень ранних этапах эволюции. [33] Принимая во внимание весь исходный геном эндосимбионта, существует три основных возможных судьбы генов в ходе эволюции. Во-первых, это потеря функционально избыточных генов. [33] при котором гены, уже представленные в ядре, в конечном итоге теряются. Второй – это перенос генов в ядро, а третий – то, что гены остаются в органелле, которая когда-то была организмом. [25] [33] [34] [35] [36] Утрата автономии и интеграция эндосимбионта с хозяином в первую очередь может быть связана с переносом ядерных генов. [36] Поскольку геномы органелл в ходе эволюции значительно сократились, ядерные гены расширились и стали более сложными. [25] В результате многие пластидные и митохондриальные процессы управляются кодируемыми в ядре генными продуктами. [25] Кроме того, многие ядерные гены, происходящие от эндосимбионтов, приобрели новые функции, не связанные с их органеллами. [25] [36]
Механизмы переноса генов
[ редактировать ]Механизмы переноса генов до конца не известны; однако существует множество гипотез, объясняющих это явление. Возможные механизмы включают гипотезу комплементарной ДНК (кДНК) и гипотезу объемного потока. [25] [34]
Гипотеза кДНК предполагает использование информационной РНК (мРНК) для транспортировки генов из органелл в ядро, где они преобразуются в кДНК и включаются в геном. [25] [34] Гипотеза кДНК основана на исследованиях геномов цветковых растений. Кодирующие белок РНК в митохондриях подвергаются сплайсингу и редактированию с использованием специфичных для органелл сайтов сплайсинга и редактирования. Однако ядерные копии некоторых митохондриальных генов не содержат специфичных для органелл сайтов сплайсинга, что указывает на процессируемый промежуточный продукт мРНК. Гипотеза кДНК с тех пор была пересмотрена, поскольку отредактированные митохондриальные кДНК вряд ли будут рекомбинировать с ядерным геномом и с большей вероятностью будут рекомбинировать со своим нативным митохондриальным геномом. Если отредактированная митохондриальная последовательность рекомбинирует с митохондриальным геномом, сайты сплайсинга митохондрий больше не будут существовать в митохондриальном геноме. Поэтому при любом последующем переносе ядерных генов также будут отсутствовать сайты сплайсинга митохондрий. [25]
Гипотеза объемного потока является альтернативой гипотезе кДНК, утверждающей, что механизмом переноса генов является ускользающая ДНК, а не мРНК. [25] [34] Согласно этой гипотезе, нарушения в органеллах, включая аутофагию (нормальное разрушение клеток), гаметогенез (образование гамет) и клеточный стресс, высвобождают ДНК, которая импортируется в ядро и включается в ядерную ДНК с помощью негомологичного соединения концов (восстановление двунитевых разрывов). [34] Например, на начальных стадиях эндосимбиоза из-за отсутствия переноса основных генов клетка-хозяин практически не имела контроля над эндосимбионтом. Эндосимбионт претерпел клеточное деление независимо от клетки-хозяина, в результате чего внутри клетки-хозяина появилось множество «копий» эндосимбионта. Некоторые из эндосимбионтов лизировались (взрывались), и высокие уровни ДНК были включены в ядро. Считается, что аналогичный механизм имеет место в растениях табака, которые демонстрируют высокую скорость переноса генов и чьи клетки содержат множество хлоропластов. [33] Кроме того, гипотеза объемного потока также подтверждается наличием неслучайных кластеров генов органелл, что предполагает одновременное движение нескольких генов. [34]
Форд Дулитл предположил, что (каким бы ни был механизм) перенос генов ведет себя как храповик, приводя к однонаправленному переносу генов из органеллы в ядерный геном. [37] Когда генетический материал органеллы включается в ядерный геном, либо органелла, либо ядерная копия гена могут быть потеряны из популяции. Если копия органеллы потеряна и зафиксирована или потеряна в результате генетического дрейфа, ген успешно переносится в ядро. Если ядерная копия потеряна, горизонтальный перенос генов может произойти снова, и клетка может «снова попытаться» осуществить успешный перенос генов в ядро. [37] Ожидается, что таким образом гены из органеллы будут накапливаться в ядерном геноме в течение эволюционного времени. [37]
Эндосимбиоз протомитохондрий
[ редактировать ]Эндосимбиотическая теория происхождения митохондрий предполагает, что протоэукариот поглотил протомитохондрию, и этот эндосимбионт стал органеллой, что стало важным шагом в эукариогенезе , создании эукариотов. [38]
Митохондрии
[ редактировать ]
Митохондрии — это органеллы, которые синтезируют переносящую энергию молекулу АТФ для клетки путем метаболизма на основе углерода макромолекул . [39] Наличие ДНК в митохондриях и белков, полученных из мтДНК , позволяет предположить, что эта органелла могла быть прокариотом до ее интеграции в протоэукариот . [40] Митохондрии считаются органеллами, а не эндосимбионтами, поскольку митохондрии и клетки-хозяева имеют общие части своего генома , одновременно делятся и обеспечивают друг друга средствами для производства энергии. [40] Было высказано предположение, что эндомембранная система и ядерная мембрана произошли от протомитохондрий . [41] [42] [43]
Ядерная мембрана
[ редактировать ]Наличие ядра является одним из основных различий между эукариотами и прокариотами . [44] Некоторые консервативные ядерные белки у эукариот и прокариот позволяют предположить, что у этих двух типов был общий предок. [45] Другая теория нуклеации заключается в том, что ранние белки ядерной мембраны заставляли клеточную мембрану сворачиваться и образовывать сферу с порами, подобную ядерной оболочке . [46] Можно было бы ожидать, что эндосимбиоз как способ формирования ядерной мембраны будет использовать меньше энергии, чем если бы клетка развивала метаболический процесс для сворачивания клеточной мембраны для этой цели. [42] Переваривание поглощенных клеток без производящих энергию митохондрий было бы сложной задачей для клетки-хозяина. [41] С этой точки зрения мембраносвязанные пузырьки или везикулы, покидающие протомитохондрии, могли образовывать ядерную оболочку. [41]
Процесс симбиогенеза, посредством которого ранняя эукариотическая клетка интегрировала протомитохондрию , вероятно, включал защиту генома хозяина от архейного высвобождения активных форм кислорода . Они могли образоваться в ходе окислительного фосфорилирования и производства АТФ протомитохондриями. Ядерная мембрана , возможно, возникла как адаптивная инновация для защиты от повреждений ДНК ядерного генома , вызванных активными формами кислорода. [47] Существенный перенос генов из предкового протомитохондриального генома в ядерный геном, вероятно, произошел во время ранней эволюции эукариот. [48] Большая защита ядерного генома от активных форм кислорода, обеспечиваемая ядерной мембраной, может объяснить адаптивное преимущество этого переноса генов.
Эндомембранная система
[ редактировать ]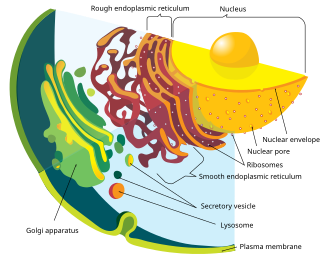
Современные эукариотические клетки используют эндомембранную систему для транспортировки продуктов и отходов внутрь, внутри и из клеток. Мембрана ядерной оболочки и эндомембранных пузырьков состоят из сходных мембранных белков. [49] Эти везикулы также имеют общие мембранные белки с органеллой, из которой они произошли или к которой направляются. [50] Это предполагает, что то, что сформировало ядерную мембрану, также сформировало эндомембранную систему. У прокариот нет сложной внутренней мембранной сети, как у эукариот, но они могут производить внеклеточные везикулы из своей внешней мембраны. [41] После того, как ранний прокариот был поглощен протоэукариотом, прокариот продолжал бы производить пузырьки, которые накапливались внутри клетки. [41] Взаимодействие внутренних компонентов везикул могло привести к тому, что эндоплазматическая сеть и аппарат Гольджи стали частями эндомембранной системы. [41]
Цитоплазма
[ редактировать ]Гипотеза синтрофии, предложенная Лопесом-Гарсией и Морейрой примерно в 2000 году, предполагала, что эукариоты возникли в результате объединения метаболических способностей архей, ферментирующей дельтапротеобактерии и метанотрофной альфапротеобактерии, которая стала митохондрией. В 2020 году та же команда обновила свое предложение по синтрофии, включив в него архей Асгарда, производящего водород, с дельтапротеобактерией, окисляющей серу. Третий организм, альфапротеобактерия, способная дышать как аэробно, так и анаэробно и окислять серу, развилась в митохондрии; возможно, он также был способен к фотосинтезу. [51]
Дата
[ редактировать ]Вопрос о том, когда произошел переход от прокариотической формы к эукариотической и когда на Земле появились первые эукариоты коронной группы , остается нерешенным. Самые старые известные окаменелости тела, которые можно с уверенностью отнести к эукариотам, - это акантоморфные акритархи из формации Деонар 1,631 г. в Индии. [52] Эти окаменелости до сих пор можно идентифицировать как производные постъядерных эукариот со сложным, генерирующим морфологию цитоскелетом , поддерживаемым митохондриями. [53] Эти ископаемые свидетельства указывают на то, что эндосимбиотическое приобретение альфапротеобактерий должно было произойти до 1,6 Гя. Молекулярные часы также использовались для оценки последнего общего предка эукариот, однако эти методы имеют большую внутреннюю неопределенность и дают широкий диапазон дат. Разумные результаты включают оценку c. 1,8 Гя. [54] Оценка 2,3 Гя [55] также кажется разумным и имеет дополнительную привлекательность, поскольку совпадает с одним из наиболее выраженных биогеохимических возмущений в истории Земли — Великим событием оксигенации в раннем палеопротерозое . Заметное увеличение концентрации кислорода в атмосфере в то время было предложено как одна из причин эукариогенеза, вызывающая эволюцию митохондрий, детоксицирующих кислород. [56] С другой стороны, Великое событие окисления могло быть следствием эукариогенеза и его влияния на экспорт и захоронение органического углерода. [57]
Органелларные геномы
[ редактировать ]Пластомы и митогеномы
[ редактировать ]
Некоторые гены эндосимбионтов остаются в органеллах. Пластиды и митохондрии сохраняют гены, кодирующие рРНК, тРНК, белки, участвующие в окислительно-восстановительных реакциях, а также белки, необходимые для транскрипции, трансляции и репликации. Существует множество гипотез, объясняющих, почему органеллы сохраняют небольшую часть своего генома; однако ни одна гипотеза не применима ко всем организмам, и эта тема до сих пор остается весьма спорной. Гипотеза гидрофобности утверждает, что высокогидрофобные ( ненавидящие воду) белки (такие как мембраносвязанные белки, участвующие в окислительно-восстановительных реакциях) нелегко транспортируются через цитозоль, и поэтому эти белки должны кодироваться в соответствующих органеллах. Гипотеза несоответствия кода утверждает, что ограничение на передачу обусловлено различиями в генетических кодах и редактировании РНК между органеллой и ядром. Гипотеза окислительно-восстановительного контроля утверждает, что гены, кодирующие белки окислительно-восстановительной реакции, сохраняются, чтобы эффективно сочетать потребность в восстановлении и синтезе этих белков. Например, если один из фотосистемы теряется из пластиды, промежуточные переносчики электронов могут потерять или получить слишком много электронов, что сигнализирует о необходимости восстановления фотосистемы. Задержка во времени, связанная с передачей сигнала ядру и транспортировкой цитозольного белка к органелле, приводит к образованию повреждающих активных форм кислорода . Последняя гипотеза утверждает, что сборка мембранных белков, особенно тех, которые участвуют в окислительно-восстановительных реакциях, требует скоординированного синтеза и сборки субъединиц; однако координацию трансляции и транспорта белков сложнее контролировать в цитоплазме. [25] [30] [33] [58]
Нефотосинтезирующие пластидные геномы
[ редактировать ]Большинство генов в митохондриях и пластидах связаны с экспрессией (транскрипцией, трансляцией и репликацией) генов, кодирующих белки, участвующие либо в фотосинтезе (в пластидах), либо в клеточном дыхании (в митохондриях). Можно предсказать, что потеря фотосинтеза или клеточного дыхания приведет к полной потере пластидного или митохондриального генома соответственно. [25] [30] [33] Хотя существуют многочисленные примеры потомков митохондрий ( митосом и гидрогеносом ), которые утратили весь свой органеллярный геном, [50] нефотосинтезирующие пластиды имеют тенденцию сохранять небольшой геном. Существуют две основные гипотезы, объясняющие это явление: [33] [59]
Гипотеза эссенциальной тРНК отмечает, что не было документально подтверждено функционального переноса генов из пластиды в ядро генов, кодирующих продукты РНК (тРНК и рРНК). В результате пластиды должны создавать свои собственные функциональные РНК или импортировать ядерные аналоги. Однако гены, кодирующие тРНК-Glu и тРНК-fmet, кажутся незаменимыми. Пластида отвечает за биосинтез гема , для которого в качестве молекулы-предшественника требуется кодируемая пластидой тРНК-Glu (из гена trnE). Как и другие гены, кодирующие РНК, trnE не может быть перенесен в ядро. Кроме того, маловероятно, что trnE может быть заменен цитозольной тРНК-Glu, поскольку trnE высоко консервативен; Изменения одного основания в trnE привели к потере синтеза гема. Ген тРНК- формилметионина (тРНК-fmet) также кодируется в пластидном геноме и необходим для инициации трансляции как в пластидах, так и в митохондриях. Пластида необходима для продолжения экспрессии гена тРНК-fmet до тех пор, пока митохондрия транслирует белки. [33]
Гипотеза ограниченного окна предлагает более общее объяснение сохранения генов в нефотосинтезирующих пластидах. [59] Согласно этой гипотезе, гены передаются в ядро после нарушения органелл. [34] Нарушения были обычным явлением на ранних стадиях эндосимбиоза, однако, как только клетка-хозяин получила контроль над делением органелл, эукариоты могли эволюционировать и иметь только одну пластиду на клетку. Наличие только одной пластиды серьезно ограничивает перенос генов. [33] поскольку лизис одной пластиды, скорее всего, приведет к гибели клеток. [33] [59] В соответствии с этой гипотезой, организмы с несколькими пластидами демонстрируют 80-кратное увеличение переноса генов из пластиды в ядро по сравнению с организмами с одиночными пластидами. [59]
Доказательство
[ редактировать ]Существует множество доказательств того, что митохондрии и пластиды, включая хлоропласты, произошли от бактерий. [60] [61] [62] [63] [64]
- Новые митохондрии и пластиды образуются только посредством бинарного деления — формы деления клеток, используемой бактериями и архей. [65]
- Если из клетки удалить митохондрии или хлоропласты, у клетки не будет возможности создавать новые. [66] У некоторых водорослей , таких как эвглена , пластиды могут быть разрушены определенными химическими веществами или длительным отсутствием света, не влияя иным образом на клетку: пластиды не регенерируют.
- Транспортные белки , называемые поринами , обнаружены во внешних мембранах митохондрий и хлоропластов, а также в мембранах бактериальных клеток. [67] [68] [69]
- Мембранный липид кардиолипин содержится исключительно во внутренней мембране митохондрий и мембранах бактериальных клеток. [70]
- Некоторые митохондрии и некоторые пластиды содержат одиночные кольцевые молекулы ДНК, сходные с ДНК бактерий как по размеру, так и по структуре. [71]
- Сравнение геномов предполагает тесную связь между митохондриями и альфапротеобактериями . [72]
- Сравнение геномов позволяет предположить тесную связь между пластидами и цианобактериями . [73]
- Многие гены в геномах митохондрий и хлоропластов утрачены или перенесены в ядро клетки-хозяина. Следовательно, хромосомы многих эукариот содержат гены, произошедшие из геномов митохондрий и пластид. [71]
- Митохондрии и пластиды содержат собственные рибосомы ; они больше похожи на таковые у бактерий (70S), чем у эукариот. [74]
- Белки, созданные митохондриями и хлоропластами, используют N-формилметионин в качестве инициирующей аминокислоты, как и белки, созданные бактериями, но не белки, созданные ядерными генами эукариот или архей. [75] [76]

Вторичный эндосимбиоз
[ редактировать ]Первичный эндосимбиоз предполагает поглощение клетки другим свободным живым организмом. Вторичный эндосимбиоз возникает, когда продукт первичного эндосимбиоза поглощается и удерживается другим свободноживущим эукариотом. Вторичный эндосимбиоз возникал несколько раз и дал начало чрезвычайно разнообразным группам водорослей и других эукариот. Некоторые организмы могут воспользоваться оппортунистическим преимуществом аналогичного процесса, когда они поглощают водоросль и используют продукты ее фотосинтеза, но как только объект-жертва умирает (или теряется), хозяин возвращается в состояние свободной жизни. Облигатные вторичные эндосимбионты становятся зависимыми от своих органелл и не могут выжить в их отсутствие. Вторичный эндосимбиоз с участием предковой красной водоросли и гетеротрофного эукариота привел к эволюции и диверсификации нескольких других фотосинтетических линий, включая Cryptophyta , Haptophyta , Stramenopiles (или Heterokontophyta) и Alveolata . [77]
Возможный вторичный эндосимбиоз наблюдался у гетеротрофного простейшего Hatena . Этот организм ведет себя как хищник, пока не проглотит зеленую водоросль , которая теряет жгутики и цитоскелет, но продолжает жить как симбионт. Тем временем Хатена , ставшая хозяином, переходит на фотосинтетическое питание, обретает способность двигаться к свету и теряет свой питательный аппарат. [78]
Несмотря на разнообразие организмов, содержащих пластиды, морфология, биохимия, геномная организация и молекулярная филогения пластидных РНК и белков предполагают единое происхождение всех существующих пластид, хотя эта теория до сих пор обсуждается. [79] [80]
Некоторые виды, включая Pediculus humanus (вши), имеют несколько хромосом в митохондриях. Это, а также филогенетика генов, закодированных в митохондриях, позволяют предположить, что митохондрии имели несколько предков, что они были приобретены в результате эндосимбиоза несколько раз, а не только один раз, и что имели место обширные слияния и перестановки генов на нескольких исходных митохондриальных хромосомах. [81]
Нитропласты
[ редактировать ]Одноклеточная морская водоросль Braarudosphaera bigelowii ( кокколитофор , являющаяся эукариотом) была обнаружена с цианобактерией в качестве эндосимбионта. Цианобактерия образует азотфиксирующую структуру, получившую название нитропласт . Он делится равномерно, когда клетка-хозяин подвергается митозу, и многие из его белков происходят из водоросли-хозяина, что означает, что эндосимбионт далеко продвинулся по пути становления органеллой. Цианобактерия получила название Candidatus Atelocyanobacterium thalassa и сокращенно UCYN-A. Водоросль — первый известный эукариот, обладающий способностью фиксировать азот. [82] [83]
См. также
[ редактировать ]- Angomonas deanei , простейшее, содержащее облигатный бактериальный симбионт.
- Hatena arenicola — вид, который, похоже, находится в процессе приобретения эндосимбионта.
- Водородная гипотеза , теория о том, что митохондрии были приобретены водородозависимыми архей, а их эндосимбионты являются факультативно анаэробными бактериями.
- Клептопластика — выделение пластид из проглоченных водорослей.
- Mixotricha paradoxa , которая сама по себе является симбионтом, содержит многочисленные эндосимбиотические бактерии.
- Parakaryon myojinensis , возможный результат независимого от эукариот эндосимбиоза
- Ева-паразит , фантастика об эндосимбиозе
- Strigomonas culicis , еще одно простейшее, имеющее облигатный бактериальный симбионт.
- Вирусный эукариогенез , гипотеза о том, что ядро клетки возникло в результате эндосимбиоза.
Ссылки
[ редактировать ]- ^ Латорре, А.; Дурбан, А.; Мойя, А.; Перето, Дж. (2011). «Роль симбиоза в эволюции эукариот» . В Гарго, М.; Лопес-Гарсиа, П.; Мартин, Х. (ред.). Происхождение и эволюция жизни: астробиологическая перспектива . Кембридж: Издательство Кембриджского университета. стр. 326–339. ISBN 978-0-521-76131-4 . Архивировано из оригинала 24 марта 2019 года . Проверено 27 августа 2017 г.
- ^ «Серийная эндосимбиотическая теория (СЭТ)» . flax.nzdl.org . Архивировано из оригинала 18 августа 2021 года . Проверено 8 марта 2021 г.
- ^ Jump up to: а б Корниш-Боуден, Атель (7 декабря 2017 г.). «Линн Маргулис и происхождение эукариотов» . Журнал теоретической биологии . Происхождение митозирующих клеток: 50-летие классической статьи Линн Саган (Маргулис). 434 : 1. Бибкод : 2017JThBi.434....1C . дои : 10.1016/j.jtbi.2017.09.027 . ПМИД 28992902 .
- ^ Jump up to: а б «Древо жизни Мерешковского» . Научный американец . Проверено 1 мая 2017 г.
- ^ Мерешковский, Константин (15 сентября 1905 г.). «О природе и происхождении хроматофоров в царстве растений» . Биологический Centralblatt (на немецком языке). 25 (18): 593–604.
- ^ См.:
- Мерешковский Константин (15 апреля 1910 г.). «Теория двух типов плазмы как основа симбиогенеза, новое исследование происхождения организмов [часть 1 из 4]». Биологический Centralblatt (на немецком языке). 30 (8): 278–288.
- Мерешковский, Константин (1 мая 1910 г.). «Теория двух типов плазмы как основа симбиогенеза, новое исследование происхождения организмов [часть 2 из 4]». Биологический Centralblatt (на немецком языке). 30 (9): 289–303.
- Мерешковский, Константин (15 мая 1910 г.). «Теория двух типов плазмы как основа симбиогенеза, новое исследование происхождения организмов [часть 3 из 4]». Биологический Centralblatt (на немецком языке). 30 (10): 321–347.
- Мерешковский, Константин (1 июня 1910 г.). «Теория двух типов плазмы как основа симбиогенеза, новое исследование происхождения организмов [часть 4 из 4]». Биологический Centralblatt (на немецком языке). 30 (11): 353–367.
- ^ Мартин, Уильям Ф .; Реттгер, Мэйо; Клёсгес, Торстен; и др. «Современная эндосимбиотическая теория: включение в уравнение латерального переноса генов» (PDF) . Журнал эндоцитобиоза и клеточных исследований . 23 : 1–5. Архивировано из оригинала (PDF) 9 марта 2022 г. Проверено 20 июля 2015 г. (URL журнала: [1] Архивировано 9 марта 2022 г. в Wayback Machine )
- ^ См.:
- Шимпер, AFW (16 февраля 1883 г.). «О развитии хлорофилльных гранул и цветных тел [часть 1 из 4]». Ботаническая газета (на немецком языке). 41 (7): 105–114. Из стр. 105: «Тем временем профессор Шмитц сообщил мне, что... высшие растения будут вести себя таким же образом». (Между тем, проф. Шмитц сообщил мне, что у водорослей не происходит образования гранул хлорофилла из клеточной плазмы, а возникают они исключительно друг из друга путем деления. Споры получают от материнского растения гранулы хлорофилла, которые создают Это открытие у водорослей заставило проф. Шмитца предположить, что высшие растения будут вести себя аналогичным образом.) Со с. 106: «Мои исследования показали... образование развивающейся ткани из апикальной меристемы». (Мои исследования показали, что точки вегетации [т. е. точки вегетативного роста] всегда содержат дифференцированные хлорофилловые тельца или их бесцветные зачатки; что они возникают не путем создания из клеточной плазмы, а друг из друга путем деления, и что они создают все хлорофилльные тельца и крахмалообразующие [тельца] тканей, развивающихся из апикальной меристемы.) Из с. 112, сноска 2: «Sollte es sich definitiv bestätigen, … an eine Symbiose erinnern». (Если бы определенно было подтверждено, что пластиды в яйцеклетках не образуются заново, то их отношение к содержащему их организму в некоторой степени наводило бы на мысль о симбиозе.)
- Шимпер, AFW (23 февраля 1883 г.). «О развитии хлорофилловых гранул и цветных тел [часть 2 из 4]». Ботаническая газета (на немецком языке). 41 (8): 121–131.
- Шимпер, AFW (2 марта 1883 г.). «О развитии хлорофилловых гранул и цветных тел [часть 3 из 4]». Ботаническая газета (на немецком языке). 41 (9): 137–146.
- Шимпер, AFW (9 марта 1883 г.). «О развитии хлорофилловых гранул и цветных тел [часть 4 из 4]». Ботаническая газета (на немецком языке). 41 (10): 153–162.
- ^ Портье, Поль (1918). Les Symbiotes (на французском языке). Париж, Франция: Masson et Cie. 293. Со с. 293: «Эта модификация в связях ядерных и митохондриальных устройств может привести к результату двух механизмов.… Cette la parthénogénèse ». (Эта модификация во взаимоотношениях ядерной и митохондриальной систем могла быть результатом двух механизмов: (а) Происходит сочетание двух факторов: вклад новых симбионтов посредством сперматозоида и редукционного деления. Это и есть оплодотворение . (б) А. существует единственный фактор: редукционное деление: в этом случае яйцо содержит достаточно активных симбионтов. Это партеногенез .)
- ^ Лейн, Ник (2005). Власть, Секс, Самоубийство. Митохондрии и смысл жизни . Нью-Йорк: Издательство Оксфордского университета . п. 14 . ISBN 9780199205646 .
- ^ Валлин, Иван Э. (1923). «Проблема митохондрий». Американский натуралист . 57 (650): 255–61. дои : 10.1086/279919 . S2CID 85144224 .
- ^ Валлин, Иван Э. (1927). Симбионтизм и происхождение видов . Балтимор: Уильямс и Уилкинс . п. 117.
- ^ Маргулис, Линн (2011). «Симбиогенез. Новый принцип эволюции, переоткрытие Бориса Михайловича Козо-Полянского (1890–1957)» . Палеонтологический журнал . 44 (12): 1525–1539. дои : 10.1134/S0031030110120087 . S2CID 86279772 .
- ^ Kozo-Polyansky, Boris Mikhaylovich (1924). Новый принцип биологии. Очерк теории симбиогенеза [ A New Principle of Biology. Essay on the Theory of Symbiogenesis ] (in Russian). Moscow and Leningrad (St. Petersburg), Russia: Пучина (Puchina).
- Английский перевод: Козо-Полянский, Борис Михайлович (2010). Маргулис, Линн (ред.). Симбиогенез: новый принцип эволюции . Перевод Фета Виктора. Кембридж, Массачусетс: Издательство Гарвардского университета .
- Рассмотрено в: Никлас, Карл Дж. (2010). «Борис М. Козо-Полянский, Симбиогенез: новый принцип эволюции». Симбиоз . 52 (1): 49–50. Бибкод : 2010Symbi..52...49N . дои : 10.1007/s13199-010-0098-7 . S2CID 41635248 .
- ^ Корнинг, Питер А. (2010). Целостный дарвинизм: синергия, кибернетика и биоэкономика эволюции . Чикаго: Издательство Чикагского университета . п. 81. ИСБН 978-0-22611-633-4 .
- ^ Рис, Ганс; Плаут, Уолтер (июнь 1962 г.). «Ультраструктура ДНК-содержащих участков хлоропласта хламидомонады» . Журнал клеточной биологии . 13 (3): 383–91. дои : 10.1083/jcb.13.3.383 . ПМК 2106071 . ПМИД 14492436 .
- ^ Рис, Ганс; Сингх, Р.Н. (январь 1961 г.). «Электронно-микроскопические исследования сине-зеленых водорослей» . Журнал биофизической и биохимической цитологии . 9 (1): 63–80. дои : 10.1083/jcb.9.1.63 . ПМК 2224983 . ПМИД 13741827 .
- ^ Чулок, К.; Гиффорд, Э. (1959). «Включение тимидина в хлоропласты спирогиры ». Биохим. Биофиз. Рез. Коммун . 1 (3): 159–64. дои : 10.1016/0006-291X(59)90010-5 .
- ^ Саган, Линн (март 1967 г.). «О происхождении митозирующих клеток». Журнал теоретической биологии . 14 (3): 255–74. Бибкод : 1967JThBi..14..225S . дои : 10.1016/0022-5193(67)90079-3 . ПМИД 11541392 .
- ^ Маргулис, Линн ; Саган, Дорион (1997). Микрокосмос: четыре миллиарда лет микробной эволюции . Беркли, Лос-Анджелес, Лондон: Издательство Калифорнийского университета . п. 29. ISBN 0-520-21064-6 .
- ^ Габальдон, Тони; Снел, Беренд; Циммерен, Франк ван; и др. (23 марта 2006 г.). «Происхождение и эволюция пероксисомального протеома» . Биология Директ . 1 (1): 8. дои : 10.1186/1745-6150-1-8 . ПМЦ 1472686 . ПМИД 16556314 . (Предоставляет доказательства, которые противоречат эндосимбиотическому происхождению пероксисом и вместо этого предполагают, что они происходят эволюционно из эндоплазматической сети )
- ^ Пизани Д., Коттон Дж.А., Макинерни Дж.О. (август 2007 г.). «Супердеревья раскрывают химерическое происхождение геномов эукариот» . Молекулярная биология и эволюция . 24 (8): 1752–1760. дои : 10.1093/molbev/msm095 . ПМИД 17504772 .
- ^ Jump up to: а б Килинг, П.Дж.; Арчибальд, Дж. М. (апрель 2008 г.). «Эволюция органелл: что в названии?» . Современная биология . 18 (8): Р345-7. Бибкод : 2008CBio...18.R345K . дои : 10.1016/j.cub.2008.02.065 . ПМИД 18430636 . S2CID 11520942 .
- ^ Сиванен, Майкл; Кадо, Кларенс И. (30 января 2002 г.). Горизонтальный перенос генов . Академическая пресса . п. 405. ИСБН 978-0126801262 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Тиммис, Джереми Н.; Эйлифф, Майкл А.; Хуан, Чун Ю.; Мартин, Уильям (2004). «Эндосимбиотический перенос генов: геномы органелл создают эукариотические хромосомы». Обзоры природы Генетика . 5 (2): 123–135. дои : 10.1038/nrg1271 . ПМИД 14735123 . S2CID 2385111 .
- ^ «Митохондрии имеют общего предка с SAR11, морским микробом глобального значения» . ScienceDaily . 25 июля 2011 года . Проверено 26 июля 2011 г.
- ^ Трэш, Дж. Кэмерон; Бойд, Алекс; Хаггетт, Меган Дж.; и др. (14 июня 2011 г.). «Филогеномные доказательства общего предка митохондрий и клады SAR11» . Научные отчеты . 1 (1): 13. Бибкод : 2011НатСР...1Е..13Т . дои : 10.1038/srep00013 . ПМК 3216501 . ПМИД 22355532 .
- ^ Дойш, О.; Ландан, Г.; Реттгер, М.; и др. (14 февраля 2008 г.). «Гены цианобактериального происхождения в ядерных геномах растений указывают на предка пластид, образующих гетероцисты». Молекулярная биология и эволюция . 25 (4): 748–761. дои : 10.1093/molbev/msn022 . ПМИД 18222943 .
- ^ Очоа де Альда, Хесус АГ; Стивен, Росио; Диаго, Мария Лус; и др. (15 сентября 2014 г.). «Пластидный предок произошел от одной из основных линий цианобактерий» . Природные коммуникации . 5 (1): 4937. Бибкод : 2014NatCo...5.4937O . дои : 10.1038/ncomms5937 . ПМИД 25222494 .
- ^ Jump up to: а б с Лила Куманду, В.; Нисбет, Р. Эллен Р.; Барбрук, Адриан К.; и др. (май 2004 г.). «Хлоропласты динофлагеллят — куда делись все гены?». Тенденции в генетике . 20 (5): 261–267. дои : 10.1016/j.tig.2004.03.008 . ПМИД 15109781 .
- ^ Таанман, JW (февраль 1999 г.). «Митохондриальный геном: структура, транскрипция, трансляция и репликация» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1410 (2): 103–23. дои : 10.1016/S0005-2728(98)00161-3 . ПМИД 10076021 .
- ^ Новак, ЕС; Мелконян М.; Глокнер, Г. (март 2008 г.). «Последовательность хроматофорного генома Паулинеллы проливает свет на приобретение фотосинтеза эукариотами» . Современная биология . 18 (6): 410–8. Бибкод : 2008CBio...18..410N . дои : 10.1016/j.cub.2008.02.051 . ПМИД 18356055 . S2CID 15929741 .
- ^ Jump up to: а б с д и ж г час я дж Барбрук, Адриан К.; Хау, Кристофер Дж.; Пертон, Сол (февраль 2006 г.). «Почему пластидные геномы сохраняются у нефотосинтезирующих организмов?». Тенденции в науке о растениях . 11 (2): 101–8. doi : 10.1016/j.tplants.2005.12.004 . ПМИД 16406301 .
- ^ Jump up to: а б с д и ж г Лейстер, Д. (декабрь 2005 г.). «Происхождение, эволюция и генетические эффекты ядерных вставок ДНК органелл» . Тенденции в генетике . 21 (12): 655–63. дои : 10.1016/j.tig.2005.09.004 . hdl : 11858/00-001M-0000-0012-3B56-7 . ПМИД 16216380 .
- ^ Килинг, П.Дж. (октябрь 2004 г.). «Разнообразие и история эволюции пластид и их хозяев» . Американский журнал ботаники . 91 (10): 1481–93. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 .
- ^ Jump up to: а б с Арчибальд, Дж. М. (январь 2009 г.). «Загадка эволюции пластид» . Современная биология . 19 (2): Р81–Р88. Бибкод : 2009CBio...19..R81A . дои : 10.1016/j.cub.2008.11.067 . ПМИД 19174147 . S2CID 51989 .
- ^ Jump up to: а б с Форд Дулиттл, W (1 декабря 1998 г.). «Вы то, что вы едите: храповик для переноса генов может объяснить бактериальные гены в ядерных геномах эукариот» . Тенденции в генетике . 14 (8): 307–311. дои : 10.1016/S0168-9525(98)01494-2 . ISSN 0168-9525 . ПМИД 9724962 .
- ^ Зиморски, Верена; Ку, Чуан; Мартин, Уильям Ф; Гулд, Свен Б (2014). «Эндосимбиотическая теория происхождения органелл». Современное мнение в микробиологии . 22 : 38–48. дои : 10.1016/j.mib.2014.09.008 . ПМИД 25306530 .
- ^ «Митохондрии, клеточная энергия, АТФ-синтаза: изучайте науку в Scitable» . www.nature.com . Проверено 24 марта 2019 г.
- ^ Jump up to: а б Грубер, А. (январь 2019 г.). «Что в названии? Как отличить органеллы эндосимбиотического происхождения от эндосимбионтов» . Микробная клетка . 6 (2): 123–133. дои : 10.15698/mic2019.02.668 . ПМК 6364258 . ПМИД 30740457 .
- ^ Jump up to: а б с д и ж Гулд, Свен Б.; Гарг, Шрирам Г.; Мартин, Уильям Ф. (июль 2016 г.). «Секреция бактериальных пузырьков и эволюционное происхождение эндомембранной системы эукариот». Тенденции в микробиологии . 24 (7): 525–534. дои : 10.1016/j.tim.2016.03.005 . ПМИД 27040918 .
- ^ Jump up to: а б Мартин, Уильям Ф.; Гарг, Шрирам; Зиморски, Верена (сентябрь 2015 г.). «Эндосимбиотические теории происхождения эукариот» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1678): 20140330. doi : 10.1098/rstb.2014.0330 . ПМЦ 4571569 . ПМИД 26323761 .
- ^ Гаравис, Мигель; Гонсалес, Карлос; Вилласанте, Альфредо (июнь 2013 г.). «О происхождении эукариотической хромосомы: роль неканонических структур ДНК в эволюции теломер» . Геномная биология и эволюция . 5 (6): 1142–50. дои : 10.1093/gbe/evt079 . ПМЦ 3698924 . ПМИД 23699225 .
- ^ «Типичные прокариотические (слева) и эукариотические (справа) клетки: изучайте науку в Scitable» . www.nature.com . Проверено 24 марта 2019 г.
- ^ Девос, Дэмиен П.; Греф, Ральф; Филд, Марк К. (июнь 2014 г.). «Эволюция ядра» . Современное мнение в области клеточной биологии . 28 (100): 8–15. дои : 10.1016/j.ceb.2014.01.004 . ПМК 4071446 . ПМИД 24508984 .
- ^ Уилсон, Кэтрин Л.; Доусон, Скотт К. (октябрь 2011 г.). «Эволюция: функциональная эволюция ядерной структуры» . Журнал клеточной биологии . 195 (2): 171–81. дои : 10.1083/jcb.201103171 . ПМК 3198171 . ПМИД 22006947 .
- ^ Бернштейн, Х.; Бернштейн, К. (2017). «Сексуальное общение архей, предшественник мейоза». В Вицани, Г. (ред.). Биокоммуникация архей . Международное издательство Спрингер. стр. 103–117. дои : 10.1007/978-3-319-65536-9 . ISBN 978-3-319-65535-2 . S2CID 26593032 .
- ^ Габальдон, Т.; Хуйнен, Массачусетс (август 2003 г.). «Реконструкция протомитохондриального метаболизма». Наука . 301 (5633): 609. doi : 10.1126/science.1085463 . ПМИД 12893934 . S2CID 28868747 .
- ^ Ляшкович Иван; Шахин, Виктор (август 2017 г.). «Функциональное значение общего эволюционного происхождения комплекса ядерных пор и систем управления эндомембраной». Семинары по клеточной биологии и биологии развития . 68 : 10–17. дои : 10.1016/j.semcdb.2017.04.006 . ПМИД 28473267 .
- ^ Jump up to: а б Хоу, Кристофер Дж. (май 2008 г.). «Клеточная эволюция: что находится в митохондрии?» . Современная биология . 18 (10): Р429–Р431. Бибкод : 2008CBio...18.R429H . дои : 10.1016/j.cub.2008.04.007 . ПМИД 18492476 . S2CID 15730462 .
- ^ Лопес-Гарсия, Purificación; Морейра, Дэвид (27 апреля 2020 г.). «Возвращение к гипотезе синтрофии происхождения эукариот» (PDF) . Природная микробиология . 5 (5): 655–667. дои : 10.1038/s41564-020-0710-4 . ISSN 2058-5276 . ПМИД 32341569 . S2CID 81678433 .
- ^ Прасад, Пиджай (август 2005 г.). «Микрокаменелости с органическими стенками из протерозойской супергруппы Виндхи в долине Сон, Мадхья-Прадеш, Индия» (PDF) . Палеоботаник . 54 .
- ^ Баттерфилд, Николас Дж. (26 ноября 2014 г.). «Ранняя эволюция эукариот» . Палеонтология . 58 (1): 5–17. дои : 10.1111/пала.12139 .
- ^ Парфри, Лаура Вегенер; Лар, Дэниел Дж.Г.; Нолл, Эндрю Х.; Кац, Лаура А. (август 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Хеджес, С. Блэр; Блэр, Джейми Э.; Вентури, Мария Л.; Шу, Джейсон Л. (январь 2004 г.). «Молекулярная временная шкала эволюции эукариот и возникновения сложной многоклеточной жизни» . Эволюционная биология BMC . 4 :2. дои : 10.1186/1471-2148-4-2 . ПМК 341452 . ПМИД 15005799 .
- ^ Гросс, Джефферсон; Бхаттачарья, Дебашиш (август 2010 г.). «Объединение происхождения пола и эукариот в развивающемся кислородном мире» . Биология Директ . 5:53 . дои : 10.1186/1745-6150-5-53 . ПМЦ 2933680 . ПМИД 20731852 .
- ^ Баттерфилд, Николас Дж. (1997). «Экология планктона и переход протерозой-фанерозой». Палеобиология . 23 (2): 247–262. Бибкод : 1997Pbio...23..247B . дои : 10.1017/S009483730001681X . S2CID 140642074 .
- ^ Яннакис, Константинос; Эроусмит, Сэмюэл Дж.; Ричардс, Люк; и др. (16 сентября 2022 г.). «Эволюционный вывод эукариот выявляет универсальные особенности, определяющие сохранение генов органелл» . Клеточные системы . 13 (11): 874–884.е5. дои : 10.1016/j.cels.2022.08.007 . hdl : 11250/3045694 . ПМИД 36115336 . S2CID 252337501 .
- ^ Jump up to: а б с д Лейн, Ник (2011). «Пластиды, геномы и вероятность переноса генов» . Геномная биология и эволюция . 3 : 372–374. дои : 10.1093/gbe/evr003 . ПМК 3101016 . ПМИД 21292628 .
- ^ Кимбалл, Дж. 2010. Страницы биологии Кимбалла . Архивировано 22 июня 2017 г. на Wayback Machine. Доступ 13 октября 2010 г. Онлайн-текст по биологии с открытым исходным кодом, написанный профессором Гарварда и автором учебника по общей биологии Джоном В. Кимбаллом.
- ^ Рис, Дж., Лиза А. Урри, Майкл Л. Кейн, Стивен А. Вассерман, Питер В. Минорски, Роберт Б. Джексон, 2010. Кэмпбелл Биология. 9-е издание Бенджамин Каммингс; 9-е изд. (7 октября 2010 г.)
- ^ Рэйвен, П.; Джонсон, Джордж; Мейсон, Кеннет; и др. (14 января 2010 г.). Биология (9-е изд.). МакГроу-Хилл.
- ^ Грей, М.В. (1992). «Возвращение к гипотезе эндосимбионта». Международный обзор цитологии . 141 : 233–357. дои : 10.1016/S0074-7696(08)62068-9 . ISBN 9780123645449 . ПМИД 1452433 .
- ^ Зиморский, В.; Ку, К.; Мартин, ВФ; Гулд, С.Б. (декабрь 2014 г.). «Эндосимбиотическая теория происхождения органелл». Современное мнение в микробиологии . 22 : 38–48. дои : 10.1016/j.mib.2014.09.008 . ПМИД 25306530 .
- ^ Марголин, Уильям (ноябрь 2005 г.). «ФцЗ и деление прокариотических клеток и органелл» . Обзоры природы. Молекулярно-клеточная биология . 6 (11): 862–71. дои : 10.1038/nrm1745 . ПМЦ 4757588 . ПМИД 16227976 .
- ^ Уайз, Роберт Р.; Хубер, Дж. Кеннет (2007). Строение и функции пластид . Берлин: Шпрингер. п. 104. ИСБН 9781402065705 .
- ^ Фишер, К.; Вебер, А.; Бринк, С.; и др. (октябрь 1994 г.). «Порины растений. Молекулярное клонирование и функциональная характеристика двух новых членов семейства поринов» . Журнал биологической химии . 269 (41): 25754–60. дои : 10.1016/S0021-9258(18)47312-7 . ПМИД 7523392 .
- ^ Зет, К.; Тейн, М. (октябрь 2010 г.). «Порины у прокариот и эукариот: общие темы и вариации». Биохимический журнал . 431 (1): 13–22. дои : 10.1042/BJ20100371 . ПМИД 20836765 . S2CID 22073622 .
- ^ Фэрман, Дж.В.; Нойнай, Н.; Бьюкенен, СК (август 2011 г.). «Структурная биология мембранных белков β-ствола: краткое изложение последних сообщений» . Современное мнение в области структурной биологии . 21 (4): 523–331. дои : 10.1016/j.sbi.2011.05.005 . ПМК 3164749 . ПМИД 21719274 .
- ^ Милейковская Е.; Доуэн, В. (октябрь 2009 г.). «Мембранные домены кардиолипина у прокариот и эукариот» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1788 (10): 2084–91. дои : 10.1016/j.bbamem.2009.04.003 . ПМЦ 2757463 . ПМИД 19371718 .
- ^ Jump up to: а б Тиммис, Джереми; Эйлифф, Майкл; Хуан, Чун; Мартин, Уильям (февраль 2004 г.). «Эндосимбиотический перенос генов: геномы органелл создают эукариотические хромосомы». Обзоры природы. Генетика . 5 (2): 123–35. дои : 10.1038/nrg1271 . ПМИД 14735123 . S2CID 2385111 .
- ^ Муньос-Гомес, Серхио; Суско, Эдвард; Уильямсон, Келси; и др. (январь 2022 г.). «Анализ гетерогенных сайтов и ветвей расширенного набора данных свидетельствует о том, что митохондрии являются сестрами известных альфапротеобактерий». Экология и эволюция природы . 6 (3): 253–62. Бибкод : 2022NatEE...6..253M . дои : 10.1038/s41559-021-01638-2 . ПМИД 35027725 . S2CID 245958471 .
- ^ Даган, Таль; Реттгер, Мэйо; Стукен, Карина; и др. (2013). «Геномы стигонематальных цианобактерий (подраздел V) и эволюция оксигенного фотосинтеза от прокариот к пластидам» . Геномная биология и эволюция . 5 (1): 31–44. дои : 10.1093/gbe/evs117 . ПМК 3595030 . ПМИД 23221676 .
- ^ Мануэль, Андреа Л.; Киспе, Джоэл; Мэйфилд, Стивен П. (август 2007 г.). «Структура рибосомы хлоропласта: новые домены регуляции трансляции» . ПЛОС Биология . 5 (8): е209. дои : 10.1371/journal.pbio.0050209 . ЧВК 1939882 . ПМИД 17683199 .
- ^ Шварц, Джеймс Х.; Мейер, Ральф; Айзенштадт, Джером М.; Браверман, Джордж (май 1967 г.). «Участие N-формилметионина в инициации синтеза белка в бесклеточных экстрактах Euglena gracilis». Журнал молекулярной биологии . 25 (3): 571–4. дои : 10.1016/0022-2836(67)90210-0 . ПМИД 5340700 .
- ^ Смит, А.Е.; Маркер, К.А. (декабрь 1968 г.). «N-формилметионил-переносная РНК в митохондриях дрожжей и печени крыс». Журнал молекулярной биологии . 38 (2): 241–3. дои : 10.1016/0022-2836(68)90409-9 . ПМИД 5760639 .
- ^ Макфадден, солдат (2001). «Первичный и вторичный эндосимбиоз и происхождение пластид». Журнал психологии . 37 (6): 951–959. Бибкод : 2001JPcgy..37..951M . дои : 10.1046/j.1529-8817.2001.01126.x . S2CID 51945442 .
- ^ Окамото, Н.; Иноуе, И. (октябрь 2005 г.). «Идет вторичный симбиоз?» . Наука . 310 (5746): 287. doi : 10.1126/science.1116125 . ПМИД 16224014 . S2CID 22081618 .
- ^ Макфадден, солдат; ван Доорен, Г.Г. (июль 2004 г.). «Эволюция: геном красных водорослей подтверждает общее происхождение всех пластид» . Современная биология . 14 (13): Р514-6. Бибкод : 2004CBio...14.R514M . дои : 10.1016/j.cub.2004.06.041 . ПМИД 15242632 . S2CID 18131616 .
- ^ Гулд, Свен Б.; Уоллер, Росс Ф.; Макфадден, Джеффри И. (2008). «Пластидная эволюция». Ежегодный обзор биологии растений . 59 (1): 491–517. doi : 10.1146/annurev.arplant.59.032607.092915 . ПМИД 18315522 . S2CID 30458113 .
- ^ Георгиадес, К.; Рауль, Д. (октябрь 2011 г.). «Корневище Reclinomonas americana, Homo sapiens, Pediculus humanus и митохондрий Saccharomyces cerevisiae» . Биология Директ . 6:55 . дои : 10.1186/1745-6150-6-55 . ПМК 3214132 . ПМИД 22014084 .
- ^ Массана, Рамон (12 апреля 2024 г.). «Нитропласт: азотфиксирующая органелла» . Наука . 384 (6692): 160–161. Бибкод : 2024Sci...384..160M . дои : 10.1126/science.ado8571 . HDL : 10261/354070 . ISSN 0036-8075 . ПМИД 38603513 . Архивировано из оригинала 15 апреля 2024 года . Проверено 15 апреля 2024 г.
- ^ Коул, Тайлер Х.; Локонте, Валентина; Турк-Кубо, Кендра А.; Ванслембрук, Бике; Мак, Винг Кван Эстер; Ченг, Шуньянь; Экман, Аксель; Чен, Цзянь-Хуа; Хагино, Кёко; Такано, Ёшихито; Нисимура, Томохиро; Адачи, Масао; Ле Гро, Марк; Ларабелл, Кэролайн; Зер, Джонатан П. (12 апреля 2024 г.). «Азотфиксирующая органелла морской водоросли». Наука . 384 (6692): 217–222. Бибкод : 2024Sci...384..217C . дои : 10.1126/science.adk1075 . ISSN 0036-8075 . ПМИД 38603509 .
Дальнейшее чтение
[ редактировать ]- Альбертс, Брюс (2002). Молекулярная биология клетки . Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . (Общий учебник)
- Бринкман, Фиона СЛ ; Бланшар, Джеффри Л.; Черкасов Артем; и др. (август 2002 г.). «Доказательства того, что растительные гены у видов Chlamydia отражают наследственное родство между Chlamydiaceae, цианобактериями и хлоропластами» . Геномные исследования . 12 (8): 1159–67. дои : 10.1101/гр.341802 . ЧВК 186644 . ПМИД 12176923 .
- Коэн, МЫ; Гарднер, Р.С. (1959). «Вирусная теория и эндосимбиоз» (PDF) . Архивировано из оригинала (PDF) 15 июля 2011 г. Проверено 26 августа 2009 г. (Обсуждается теория происхождения эукариотических клеток путем включения митохондрий и хлоропластов в анаэробные клетки с акцентом на бактериально-фаговые и предполагаемые вирусные взаимодействия митохондрий и хлоропластов.)
- Джарвис, П. (апрель 2001 г.). «Внутриклеточная передача сигналов: хлоропласты разговаривают!» . Современная биология . 11 (8): Р307-10. Бибкод : 2001CBio...11.R307J . дои : 10.1016/S0960-9822(01)00171-3 . ПМИД 11369220 . S2CID 11753648 . (Приводятся доказательства того, что белки, кодируемые хлоропластами, влияют на транскрипцию ядерных генов, в отличие от более хорошо документированных случаев, когда белки, кодируемые ядром, влияют на митохондрии или хлоропласты.)
- Бланшар, Дж.Л.; Линч, М. (июль 2000 г.). «Органелларные гены: почему они попадают в ядро?». Тенденции в генетике . 16 (7): 315–20. дои : 10.1016/S0168-9525(00)02053-9 . ПМИД 10858662 . (Обсуждаются теории о том, как гены митохондрий и хлоропластов передаются в ядро, а также какие этапы должен пройти ген, чтобы завершить этот процесс.)
- Окамото, Н.; Иноуе, И. (октябрь 2005 г.). «Идет вторичный симбиоз?». Наука . 310 (5746): 287. doi : 10.1126/science.1116125 . ПМИД 16224014 . S2CID 22081618 .
- Понимание научной группы. «Клетки внутри клеток: экстраординарное утверждение с экстраординарными доказательствами» (PDF) . Калифорнийский университет в Беркли . Проверено 16 февраля 2014 г.