Геобиология

Геобиология — это область научных исследований , изучающая взаимодействие между физической Землей и биосферой . Это относительно молодая область, и ее границы подвижны. Существует значительное совпадение с областями экологии , эволюционной биологии , микробиологии , палеонтологии и особенно почвоведения и биогеохимии . Геобиология применяет принципы и методы биологии, геологии и почвоведения для изучения древней истории совместной эволюции жизни и Земли, а также роли жизни в современном мире. [2] Геобиологические исследования, как правило, сосредоточены на микроорганизмах и на роли жизни в изменении химической и физической среды педосферы , которая существует на пересечении литосферы , атмосферы , гидросферы и /или криосферы . Она отличается от биогеохимии тем, что основное внимание уделяется процессам и организмам в пространстве и времени, а не глобальным химическим циклам.
Геобиологические исследования синтезируют геологические данные с современными биологическими исследованиями. Он посвящен процессам – как организмы влияют на Землю и наоборот – а также истории – как Земля и жизнь изменились вместе. Многие исследования основаны на поисках фундаментального понимания, но геобиология также может быть применена, как в случае с микробами, которые очищают разливы нефти . [3]
Геобиология использует молекулярную биологию , микробиологию окружающей среды , органическую геохимию и геологические данные для исследования эволюционной взаимосвязи жизни и Земли. Он пытается понять, как изменилась Земля с момента зарождения жизни и какой она могла быть на этом пути. Некоторые определения геобиологии даже раздвигают границы этих временных рамок – к пониманию происхождения жизни и той роли, которую люди играли и будут продолжать играть в формировании Земли в антропоцене . [3]
История [ править ]

Термин «геобиология» был введен Лоренсом Баасом Бекингом в 1934 году. По его словам, геобиология «является попыткой описать отношения между организмами и Землей», поскольку «организм является частью Земли, и его судьба переплетена с судьбой Земли». Земля." Определение геобиологии, данное Баасом Бекингом, родилось из желания объединить биологию окружающей среды с лабораторной биологией. То, как он это практиковал, тесно связано с современной микробной экологией окружающей среды , хотя его определение остается применимым ко всей геобиологии. В своей книге «Геобиология» Басс Бекинг заявил, что у него нет намерения изобретать новую область исследований. [4] На понимание геобиологии Баасом Бекингом сильно повлияли его предшественники, в том числе Мартинус Бейеринк , его учитель из Голландской школы микробиологии. Среди других был Владимир Вернадский , который в своей книге «Биосфера» 1926 года утверждал, что жизнь меняет поверхностную среду Земли. [5] и Сергей Виноградский, известный открытием литотрофных бактерий. [6]
Первой лабораторией, официально посвященной изучению геобиологии, была геобиологическая лаборатория Бааса Бекинга в Австралии, открывшая свои двери в 1965 году. [4] Однако потребовалось еще около 40 лет, чтобы геобиология стала прочно укоренившейся научной дисциплиной, отчасти благодаря достижениям в области геохимии и генетики , которые позволили ученым начать синтезировать исследования жизни и планеты.
х годах Альфред Трейбс обнаружил хлорофиллоподобные в порфирины В 1930 - нефти , подтвердив ее биологическое происхождение. [7] тем самым основав органическую геохимию и установив понятие биомаркеров , важнейшего аспекта геобиологии. Но прошло несколько десятилетий, прежде чем появились инструменты, позволяющие начать всерьез искать химические признаки жизни в горных породах. В 1970-х и 80-х годах такие ученые, как Джеффри Эглингтон и Роджер Саммонс, начали находить липидные биомаркеры в породах, используя такое оборудование, как GCMS . [8]
Что касается биологии, то в 1977 году Карл Везе и Джордж Фокс опубликовали филогению жизни на Земле, включая новый домен — археи . [9] А в 1990-е годы стали возможными исследования в области генетики и геномики , расширившие сферу изучения взаимодействия жизни и планеты.
Сегодня геобиология имеет свои собственные журналы, такие как Geobiology , основанный в 2003 году. [10] и биогеонауки , основанные в 2004 году, [11] а также признание на крупных научных конференциях. у него была собственная Гордонская исследовательская конференция . В 2011 году [12] издан ряд учебников по геобиологии, [3] [13] и многие университеты по всему миру предлагают программы на получение степени по геобиологии (см. Внешние ссылки).
Основные геобиологические события
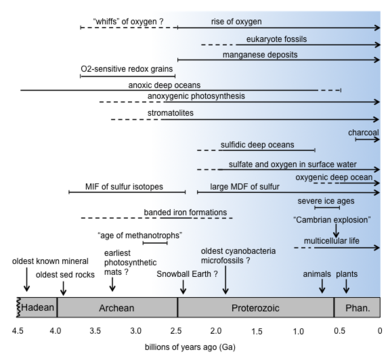
Возможно, самым глубоким геобиологическим событием является введение кислорода в атмосферу фотосинтезирующими бактериями . Это насыщение кислородом первичной атмосферы Земли (так называемая кислородная катастрофа или Великое событие насыщения кислородом ) и насыщение кислородом океанов изменили поверхностные биогеохимические циклы и типы организмов, для которых был выбран эволюционный отбор.
Следующим важным изменением стало появление многоклеточности . Присутствие кислорода позволило эукариотам , а позже и многоклеточной жизни развиваться.
Более антропоцентрические геобиологические события включают в себя происхождение животных и формирование наземной растительной жизни, что повлияло на континентальную эрозию и круговорот питательных веществ и, вероятно, изменило типы наблюдаемых рек, что позволило организовать русло того, что раньше было преимущественно разветвленными реками.
Более тонкие геобиологические события включают роль термитов в переворачивании отложений, коралловых рифов в отложении карбоната кальция и прибоя волн, губок в поглощении растворенного морского кремнезема, роли динозавров в прорыве речных дамб и содействии наводнениям, а также роли навоза крупных млекопитающих в распределяя питательные вещества. [15] [16]
Важные понятия [ править ]
Геобиология основана на нескольких основных концепциях, которые объединяют изучение Земли и жизни. Хотя многие аспекты изучения прошлых и настоящих взаимодействий между жизнью и Землей остаются неясными, несколько важных идей и концепций составляют основу знаний в геобиологии, которые служат платформой для постановки исследовательских вопросов, включая эволюцию жизни и планеты и совместная эволюция обоих, генетика - как с исторической, так и с функциональной точки зрения, метаболическое разнообразие всей жизни, седиментологическое сохранение прошлой жизни и происхождение жизни.
Коэволюция жизни и Земли [ править ]
Основная концепция геобиологии заключается в том, что жизнь меняется со временем в ходе эволюции . Теория эволюции постулирует, что уникальные популяции организмов или видов возникли в результате генетических модификаций предковой популяции, которые передавались по наследству в результате дрейфа и естественного отбора . [17]
Наряду со стандартной биологической эволюцией жизнь и планета развиваются совместно. Поскольку лучшими адаптациями являются те, которые соответствуют экологической нише , в которой живет организм, физические и химические характеристики окружающей среды стимулируют эволюцию жизни путем естественного отбора, но может быть и обратное: с каждым появлением эволюции окружающая среда изменения.
Классическим примером коэволюции является эволюция кислород производящих фотосинтезирующих цианобактерий, атмосферу Земли , которые насыщали кислородом архейскую . Предки цианобактерий начали использовать воду в качестве источника электронов для использования энергии Солнца и вытеснения кислорода до или во время раннего палеопротерозоя . В это время, примерно 2,4–2,1 миллиарда лет назад, [18] геологические данные свидетельствуют о том, что уровень содержания кислорода в атмосфере начал повышаться во время так называемого Великого события оксигенации (GOE) . [19] [20] Неясно, как долго цианобактерии осуществляли кислородный фотосинтез до GOE. Некоторые данные свидетельствуют о том, что существовали геохимические «буферы» или поглотители, подавляющие подъем кислорода, например, в результате вулканизма. [21] хотя цианобактерии, возможно, производили его и до ПХ. [22] Другие данные указывают на то, что рост оксигенного фотосинтеза совпал с GOE. [23]

Присутствие кислорода на Земле с момента его первого производства цианобактериями до GOE и по сегодняшний день радикально повлияло на ход эволюции жизни и планеты. [19] Это могло спровоцировать образование окисленных минералов. [24] и исчезновение окисляемых минералов, таких как пирит, из русел древних рек. [25] Присутствие образований полосчатого железа (BIF) было интерпретировано как ключ к повышению содержания кислорода, поскольку небольшие количества кислорода могли вступать в реакцию с восстановленным двухвалентным железом (Fe(II)) в океанах, что приводило к отложению отложений, содержащих Оксид Fe(III) в таких местах, как Западная Австралия. [26] Однако любая окислительная среда, в том числе предоставляемая микробами, такими как фотоавтотроф, окисляющий железо Rhodopseudomonas palustris , [27] может вызвать образование оксида железа и, следовательно, отложение BIF. [28] [29] [30] Другие механизмы включают окисление ультрафиолетовым светом . [31] Действительно, BIF происходят на больших участках истории Земли и могут не коррелировать только с одним событием. [30]
Другие изменения, коррелирующие с повышением содержания кислорода, включают появление ржаво-красных древних палеопочв . [19] различное изотопное фракционирование таких элементов, как сера , [32] глобальные оледенения и явления «Земля-снежок» , [33] возможно, вызвано окислением метана кислородом, не говоря уже о пересмотре типов организмов и метаболизма на Земле. В то время как организмы до появления кислорода, вероятно, были отравлены газообразным кислородом, как и многие анаэробы сегодня. [34] те, кто разработал способы использования способности кислорода принимать электроны и давать энергию, были готовы к процветанию и колонизации аэробной среды.

Земля изменилась [ править ]
Земля не осталась прежней с момента своего планетарного образования 4,5 миллиарда лет назад. [35] [36] Континенты формировались, распадались и сталкивались, создавая новые возможности и препятствия для распространения жизни. Изменилось окислительно-восстановительное состояние атмосферы и океанов, о чем свидетельствуют изотопные данные. Колеблющиеся количества неорганических соединений, таких как углекислый газ , азот , метан и кислород, были обусловлены развитием новых биологических метаболизмов жизни для производства этих химических веществ и стимулировали эволюцию новых метаболизмов для использования этих химических веществ. Земля приобрела магнитное поле около 3,4 млрд лет назад [37] которая претерпела серию геомагнитных инверсий порядка миллионов лет. [38] Температура поверхности постоянно колеблется, падая во время оледенений и событий «Земля-снежок» из-за обратной связи между льдом и альбедо . [39] поднимается и тает из-за вулканического выделения газов и стабилизируется из-за воздействия силикатного выветривания . [40]
И Земля не единственная, кто изменился – светимость Солнца со временем увеличилась. Поскольку в горных породах зафиксирована история относительно постоянных температур с момента зарождения Земли, должно быть, было больше парниковых газов , чтобы поддерживать температуру на высоком уровне в архейском периоде, когда Солнце было моложе и слабее. [41] Все эти основные различия в окружающей среде Земли накладывали совершенно разные ограничения на эволюцию жизни на протяжении всей истории нашей планеты. Более того, всегда происходят более тонкие изменения в среде обитания жизни, формирующие организмы и следы, которые мы наблюдаем сегодня и в летописях горных пород.
функцию и кодируют геобиологическую историю Гены
Генетический код является ключом к наблюдению за историей эволюции и пониманию возможностей организмов. Гены являются основной единицей наследственности и функций и, как таковые, являются основной единицей эволюции и средством метаболизма . [42]
историю эволюции предсказывает Филогения

Филогения берет генетические последовательности живых организмов и сравнивает их друг с другом, чтобы выявить эволюционные связи, подобно тому, как генеалогическое древо показывает, как люди связаны со своими дальними родственниками. [43] Это позволяет нам расшифровать современные отношения и сделать вывод о том, как эволюция происходила в прошлом.
Филогения может дать некоторое представление об истории в сочетании с немного большей информацией. Каждое различие в ДНК указывает на расхождение между одним видом и другим. [43] Это расхождение, будь то в результате дрейфа или естественного отбора, отражает некоторый промежуток времени. [43] Сравнение последовательностей ДНК само по себе дает запись истории эволюции с произвольной мерой филогенетического расстояния, «датирующего» последнего общего предка. Однако, если доступна информация о скорости генетических мутаций или присутствуют геологические маркеры для калибровки эволюционного расхождения (т. е. окаменелости ), у нас есть временная шкала эволюции. [44] Отсюда, имея представление о других одновременных изменениях в жизни и окружающей среде, мы можем начать рассуждать о том, почему могли быть выбраны определенные эволюционные пути. [45]
Гены метаболизм кодируют
Молекулярная биология позволяет ученым понять функцию гена с помощью микробного культивирования и мутагенеза . Поиск подобных генов в других организмах, а также в метагеномных и метатранскриптомных данных позволяет нам понять, какие процессы могут быть актуальными и важными в данной экосистеме, обеспечивая понимание биогеохимических циклов в этой среде.
Например, интересной проблемой геобиологии является роль организмов в глобальном круговороте метана . Генетика показала, что ген метанмонооксигеназы ( pmo ) используется для окисления метана и присутствует у всех аэробных метан-окислителей или метанотрофов . [46] Присутствие последовательностей ДНК гена pmo в окружающей среде можно использовать в качестве показателя метанотрофии. [47] [48] Более обобщающим инструментом является ген 16S рибосомальной РНК , обнаруженный у бактерий и архей. Этот ген развивается очень медленно с течением времени и обычно не передается горизонтально , поэтому его часто используют для различения различных таксономических единиц организмов в окружающей среде. [9] [49] Таким образом, гены являются ключом к метаболизму и идентичности организма. Генетика позволяет нам спрашивать: «Кто там?» и «что они делают?» Этот подход называется метагеномикой . [49]

разнообразие влияет на Метаболическое окружающую среду
Жизнь использует химические реакции для выработки энергии, биосинтеза и устранения отходов. [52] Разные организмы используют совершенно разные метаболические подходы для удовлетворения этих основных потребностей. [53] В то время как животные, такие как мы, ограничены аэробным дыханием , другие организмы могут «дышать» сульфатом (SO42-), нитратом (NO3-), трехвалентным железом (Fe(III)) и ураном (U(VI)) или жить за счет энергия брожения . [53] Некоторые организмы, например растения, являются автотрофами , то есть могут улавливать углекислый газ для биосинтеза. Растения являются фотоавтотрофами , поскольку они используют энергию света для фиксации углерода. Микроорганизмы используют оксигенную и аноксигенную фотоавтотрофию, а также хемоавтотрофию . Микробные сообщества могут координировать свои действия в синтрофном метаболизме, чтобы изменить кинетику реакций в свою пользу. Многие организмы могут осуществлять множественный метаболизм для достижения одной и той же конечной цели; их называют миксотрофами . [53]
Биотический метаболизм напрямую связан с глобальным круговоротом элементов и соединений на Земле. Геохимическая среда питает жизнь, которая затем производит различные молекулы, которые попадают во внешнюю среду. (Это имеет прямое отношение к биогеохимии .) Кроме того, биохимические реакции катализируются ферментами, которые иногда предпочитают один изотоп другим. Например, кислородный фотосинтез катализируется RuBisCO , который предпочитает углерод-12 углероду-13, что приводит к фракционированию изотопов углерода в породах. [54]

историю Осадочные рассказывают породы
Осадочные породы сохраняют остатки истории жизни на Земле в виде окаменелостей , биомаркеров , изотопов и других следов. Летопись породы далека от совершенства, а сохранение биосигнатур — редкое явление. Понимание того, какие факторы определяют степень сохранности и значение того, что сохраняется, является важным компонентом распутывания древней истории совместной эволюции жизни и Земли. [8] Летопись осадочных пород позволяет ученым наблюдать изменения в жизни и составе Земли с течением времени, а иногда даже датировать крупные изменения, такие как события вымирания.
Некоторые классические примеры геобиологии в осадочной летописи включают строматолиты и образования полосчатого железа. Роль жизни в происхождении того и другого является широко обсуждаемой темой. [19]
Жизнь – это, по сути, химия [ править ]
Первая жизнь возникла в результате абиотических химических реакций . Когда это произошло, как это произошло и даже на какой планете это произошло, неизвестно. Однако жизнь следует правилам и возникла из безжизненной химии и физики . Это ограничено такими принципами, как термодинамика . Это важная концепция в этой области, поскольку она представляет собой воплощение взаимосвязанности, если не одинаковости, жизни и Земли. [55]
Хотя попытки понять, как и когда возникла жизнь, часто делегируются области астробиологии , они также актуальны и для геобиологии. [56] Первые крупные шаги к пониманию того, «как» были сделаны благодаря эксперименту Миллера-Юри , когда аминокислоты образовались из смоделированного « первичного супа ». Другая теория состоит в том, что жизнь зародилась в системе, очень похожей на гидротермальные жерла в срединно-океанических центрах распространения . В синтезе Фишера-Тропша различные углеводороды в условиях, подобных жерлам, образуются . Другие идеи включают в себя гипотезу «мира РНК» , которая постулирует, что первой биологической молекулой была РНК , и идею о том, что жизнь зародилась где-то в Солнечной системе и была принесена на Землю, возможно, через метеорит . [55]
Методология [ править ]

Хотя геобиология представляет собой разнообразную и разнообразную область, охватывающую идеи и методы из широкого спектра дисциплин, существует ряд важных методов, которые являются ключевыми для изучения взаимодействия жизни и Земли, которые освещены здесь. [3]
- Лабораторное культивирование микробов используется для характеристики метаболизма и образа жизни представляющих интерес организмов.
- генов Секвенирование позволяет ученым изучать взаимоотношения между современными организмами с помощью филогенетики.
- Экспериментальные генетические манипуляции или мутагенез используются для определения функции генов в живых организмах.
- Микроскопия используется для визуализации микробного мира. Работа с микроскопом варьируется от наблюдения за окружающей средой до количественных исследований с помощью зондов ДНК и визуализации границы раздела микроб-минерал в высоком разрешении с помощью электронного микроскопа (ЭМ).
- Изотопные индикаторы можно использовать для отслеживания биохимических реакций и понимания микробного метаболизма.
- Естественное содержание изотопов в горных породах можно измерить, чтобы определить фракционирование изотопов , соответствующее биологическому происхождению.
- Детальная характеристика окружающей среды важна для понимания того, что в среде обитания может стимулировать эволюцию жизни и, в свою очередь, как жизнь может изменить эту нишу. Оно включает, помимо прочего, температуру, свет, pH, соленость, концентрацию определенных молекул, таких как кислород, и биологическое сообщество.
- седиментология и стратиграфия Для изучения горных пород используются . Летопись горных пород хранит историю геобиологических процессов в отложениях, которую можно раскрыть, понимая процессы отложения , седиментации , уплотнения , диагенеза и деформации .
- Поиск и изучение окаменелостей, хотя их часто относят к отдельной области палеонтологии , играют важную роль в геобиологии, хотя масштабы окаменелостей обычно меньшие ( микропалеонтология ).
- Биохимический анализ биомаркеров , которые представляют собой окаменелые или современные молекулы, указывающие на присутствие определенной группы организмов или метаболизма, используется для ответа на вопросы о доказательствах жизни и метаболического разнообразия. [8]
- Палеомагнетизм — это исследование древнего магнитного поля планеты. Это важно для понимания магнитофоссилий , биоминерализации и глобальных изменений экосистем.
[ править ]
Как следует из названия, геобиология тесно связана со многими другими областями исследований и не имеет четко определенных границ или полного согласия относительно того, что именно они включают. Некоторые практикующие специалисты рассматривают его границы очень широко, охватывая многие старые и более устоявшиеся области, такие как биогеохимия, палеонтология и микробная экология. Другие придерживаются более узкой точки зрения, приписывая ее новым исследованиям, которые находятся между этими существующими областями, например, геомикробиологии. В следующий список входят как те, которые явно являются частью геобиологии, например, геомикробиология, так и те, которые разделяют научные интересы, но исторически не считались субдисциплиной геобиологии, например, палеонтология.
Астробиология [ править ]
Астробиология — это междисциплинарная область, которая использует сочетание геобиологических и планетарных данных для установления контекста для поиска жизни на других планетах . Происхождение жизни из неживой химии и геологии, или абиогенез , является основной темой астробиологии. Несмотря на то, что это фундаментально касается Земли и, следовательно, представляет большой геобиологический интерес, изучение происхождения жизни требует рассмотрения того, что требуется для жизни, что особенного на Земле, если вообще что-то есть, что могло измениться, чтобы позволить жизни расцвести, что представляет собой свидетельство существования жизни и даже то, что представляет собой саму жизнь. Это те же вопросы, которые ученые могут задать при поиске инопланетной жизни. Кроме того, астробиологи исследуют возможность жизни на основе других метаболизмов и элементов, выживаемость земных организмов на других планетах или космических кораблях, эволюцию планет и солнечной системы, а также космическую геохимию. [57]
Биогеохимия [ править ]
Биогеохимия — это системная наука, которая синтезирует изучение биологических, геологических и химических процессов для понимания реакций и состава природной среды. В первую очередь он касается глобальных элементарных циклов, таких как цикл азота и углерода. Отцом биогеохимии был Джеймс Лавлок , чья « гипотеза Геи » предположила, что биологические, химические и геологические системы Земли взаимодействуют, чтобы стабилизировать условия на Земле, поддерживающие жизнь. [58]
Геобиохимия [ править ]

Геобиохимия похожа на биогеохимию , но отличается тем, что уделяет особое внимание влиянию геологии на развитие биохимических процессов жизни, в отличие от роли жизни в земных циклах. Его основная цель — связать биологические изменения, включая эволюционные модификации генов и изменения в экспрессии генов и белков, с изменениями температуры, давления и состава геохимических процессов, чтобы понять, когда и как развивался метаболизм. Геобиохимия основана на представлении о том, что жизнь — это планетарный ответ, поскольку метаболический катализ позволяет высвободить энергию, захваченную остывающей планетой. [59]
Экологическая микробиология
Микробиология — это широкая научная дисциплина, занимающаяся изучением той жизни, которую лучше всего рассматривать под микроскопом. Она охватывает несколько областей, имеющих непосредственное отношение к геобиологии, и все инструменты микробиологии относятся к геобиологии. Экологическая микробиология особенно переплетена с геобиологией, поскольку она направлена на понимание реальных организмов и процессов, которые имеют отношение к природе, в отличие от традиционного лабораторного подхода к микробиологии. Микробная экология аналогична, но, как правило, больше фокусируется на лабораторных исследованиях и отношениях между организмами внутри сообщества, а также внутри экосистемы их химической и геологической физической среды. Оба метода основаны на таких методах, как сбор образцов из различных сред, метагеномика , секвенирование ДНК и статистика .
и Геомикробиология геохимия микробная

Геомикробиология традиционно изучает взаимодействие микробов и минералов . Хотя микробная геохимия обычно полагается на инструменты микробиологии, она использует геологические и химические методы, чтобы подойти к той же теме с точки зрения горных пород. Геомикробиология и микробная геохимия (ГМГ) — относительно новая междисциплинарная область, которая в более широком смысле изучает взаимоотношения между микробами, Землей и системами окружающей среды. Заявленная как часть геобиологии и геохимии, GMG стремится понять элементарные биогеохимические циклы и эволюцию жизни на Земле. В частности, он задает вопросы о том, где живут микробы, их локальное и глобальное распространение, их структурная и функциональная биохимия, как они развивались, биоминерализация, а также их потенциал сохранения и присутствие в летописи горных пород. Во многих отношениях GMG кажется эквивалентом геобиологии, но отличается по объему: геобиология фокусируется на роли всей жизни, тогда как GMG является строго микробной. Тем не менее, именно эти мельчайшие существа доминировали в истории жизни, интегрированной с течением времени, и, похоже, имели самые далеко идущие последствия. [60]
Молекулярная геомикробиология
Молекулярная геомикробиология использует механистический подход к пониманию биологических процессов, имеющих геологическое значение. Он может находиться на уровне ДНК, белка, липидов или любого метаболита . Одним из примеров исследований в области молекулярной геомикробиологии является изучение того, как недавно образовавшиеся лавовые поля колонизируются микробами. Университет Хельсинки в настоящее время проводит исследования, чтобы определить, какие конкретные микробные признаки необходимы для успешной первоначальной колонизации и как волны микробной сукцессии могут превратить вулканическую породу в плодородную почву. [61]
Органическая геохимия [ править ]
Органическая геохимия — это изучение органических молекул, которые появляются в летописи окаменелостей в осадочных породах. Исследования в этой области касаются молекулярных окаменелостей , которые часто являются липидными биомаркерами. Молекулы, такие как стерины и гопаноиды, мембранные липиды, обнаруженные у эукариот и бактерий соответственно, могут сохраняться в летописях горных пород на протяжении миллиардов лет. После смерти организма, из которого они произошли, и осаждения они подвергаются процессу, называемому диагенезом , при котором многие специфические функциональные группы липидов теряются, но углеводородный скелет остается нетронутым. Эти окаменелые липиды называются стеранами и гопанами соответственно. [62] Существуют также другие типы молекулярных ископаемых, такие как порфирины , открытие которых в нефти Альфредом Трейбсом фактически привело к открытию этой области. [8] Другие аспекты геохимии , которые также имеют отношение к геобиологии, включают изотопную геохимию, в которой ученые ищут фракционирование изотопов в горных породах, и химический анализ биоминералов , таких как магнетит или микробиологическое осаждение золота.

Палеонтология [ править ]
Возможно, самая старая из всех, палеонтология – это изучение окаменелостей. Он включает в себя открытие, раскопки, датировку и палеоэкологическое понимание любого типа окаменелостей, микробов или динозавров, следов или окаменелостей тел. Микропалеонтология особенно важна для геобиологии. Предполагаемые бактериальные микроокаменелости и древние строматолиты используются в качестве доказательства развития метаболизма, такого как кислородный фотосинтез. [63] Поиск молекулярных окаменелостей, таких как липидные биомаркеры, такие как стераны и гопаны, также сыграл важную роль в геобиологии и органической геохимии. [8] Соответствующие субдисциплины включают палеоэкологию и палеобиогеографию .
Биогеография [ править ]
Биогеография – это изучение географического распределения жизни во времени. Он может смотреть на нынешнее распределение организмов по континентам или между микронишами, или на распределение организмов во времени или в прошлом, что называется палеобиогеографией.
Эволюционная биология [ править ]
Эволюционная биология — это изучение эволюционных процессов, которые сформировали разнообразие жизни на Земле. Он включает в себя генетику , экологию, биогеографию и палеонтологию для анализа таких тем, как естественный отбор , изменчивость, адаптация , дивергенция, генетический дрейф и видообразование .
Экогидрология [ править ]
Экогидрология — это междисциплинарная область, изучающая взаимодействие воды и экосистем. Стабильные изотопы воды иногда используются в качестве индикаторов источников воды и путей потока между физической средой и биосферой. [64] [65]
См. также [ править ]
Ссылки [ править ]
- ^ «Жизнь в экстремальную жару — Йеллоустонский национальный парк (Служба национальных парков США)» . www.nps.gov . Проверено 4 июня 2016 г.
- ^ Дилек, Йылдирим; Харальд Фернес; Карлис Мюленбахс (2008). Связи между геологическими процессами, микробной деятельностью и эволюцией жизни . Спрингер. п. ISBN против 978-1-4020-8305-1 .
- ^ Jump up to: Перейти обратно: а б с д Нолл, Эндрю Х.; Кэнфилд, профессор Дон Э.; Конхаузер, Курт О. (30 марта 2012 г.). Основы геобиологии . Джон Уайли и сыновья. ISBN 9781118280881 .
- ^ Jump up to: Перейти обратно: а б Бекинг, LGM Баас (2015). Кэнфилд, Дональд Э. (ред.). Геобиология Бааса Бекинга . Джон Уайли и сыновья.
- ^ Вернадский, Владимир Иванович (1926). Биосфера .
- ^ Виноградский, Сергей (1887). «О серобактериях». Газета Бот (45): 489–610.
- ^ Трейбс, Альфред (19 сентября 1936 г.). «Производные хлорофилла и гемина в органических минералах». Прикладная химия . 49 (38): 682–686. Бибкод : 1936АнгЧ..49..682Т . дои : 10.1002/anie.19360493803 . ISSN 1521-3757 .
- ^ Jump up to: Перейти обратно: а б с д и Гейнс, С.М. (2008). Эхо жизни: что ископаемые молекулы рассказывают об истории Земли . Издательство Оксфордского университета, США.
- ^ Jump up to: Перейти обратно: а б Вёзе, Карл Р.; Фокс, Джордж Э. (1 ноября 1977 г.). «Филогенетическая структура прокариотической области: первичные царства» . Труды Национальной академии наук . 74 (11): 5088–5090. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ISSN 0027-8424 . ПМК 432104 . ПМИД 270744 .
- ^ «Геобиология». Геобиология . дои : 10.1111/(issn) 1472-4669 .
- ^ «Редакторы журнала Biogeosciences обсуждают успех журнала - ScienceWatch.com - Thomson Reuters» . archive.sciencewatch.com . Рейтер . Проверено 20 мая 2016 г.
- ^ «Гордонские исследовательские конференции - Подробности конференции - Геобиология» . www.grc.org . Проверено 20 мая 2016 г.
- ^ Честное слово, Генри Лутц; Ньюман, Дайан К.; Капплер, Андреас (15 октября 2015 г.). Геомикробиология Эрлиха, шестое издание . ЦРК Пресс. ISBN 9781466592414 .
- ^ Беккер, Андрей (01 января 2014 г.). «Великое событие оксигенации». В Амилсе, Рикардо; Гарго, Мюриэль; Кинтанилья, Хосе Серничаро; Кливс, Хендерсон Джеймс; Ирвин, Уильям М.; Пинти, Даниэле; Визо, Мишель (ред.). Энциклопедия астробиологии . Шпрингер Берлин Гейдельберг. стр. 1–9. дои : 10.1007/978-3-642-27833-4_1752-4 . ISBN 9783642278334 .
- ^ Даути, CE; Вольф, А.; Малхи, Ю. (2013). «Наследие вымирания плейстоценовой мегафауны на доступность питательных веществ в Амазонии» . Природа Геонауки . 6 (9): 761–764. Бибкод : 2013NatGe...6..761D . дои : 10.1038/ngeo1895 . [ постоянная мертвая ссылка ]
- ^ Баттерфилд, Нью-Джерси (2011). «Животные и изобретение фанерозойской системы Земли». Тенденции в экологии и эволюции . 26 (2): 81–87. дои : 10.1016/j.tree.2010.11.012 . ПМИД 21190752 .
- ^ Дарвин, К. (1859). О происхождении вида Лондон. Великобритания: Джон Мюррей .
- ^ Лайонс, Тимоти В.; Рейнхард, Кристофер Т.; Планавский, Ной Дж. (2014). «Повышение содержания кислорода в раннем океане и атмосфере Земли». Природа . 506 (7488): 307–315. Бибкод : 2014Natur.506..307L . дои : 10.1038/nature13068 . ПМИД 24553238 . S2CID 4443958 .
- ^ Jump up to: Перейти обратно: а б с д Фишер, Вудворд В.; Конопля, Джеймс; Джонсон, Йена Э. (28 июня 2013 г.). «Эволюция кислородного фотосинтеза» . Ежегодный обзор наук о Земле и планетах . 44 : 647–683. Бибкод : 2016AREPS..44..647F . doi : 10.1146/annurev-earth-060313-054810 . ISSN 0084-6597 .
- ^ Холланд, Генрих Д. (1 ноября 2002 г.). «Вулканические газы, черные курильщики и великое событие окисления». Geochimica et Cosmochimica Acta . 66 (21): 3811–3826. Бибкод : 2002GeCoA..66.3811H . дои : 10.1016/S0016-7037(02)00950-X .
- ^ Гайяр, Фабрис; Скайлет, Бруно; Арндт, Николас Т. (2011). «Атмосферная оксигенация, вызванная изменением давления вулканической дегазации» (PDF) . Природа . 478 (7368): 229–232. Бибкод : 2011Natur.478..229G . дои : 10.1038/nature10460 . ПМИД 21993759 . S2CID 205226436 .
- ^ Анбар, Ариэль Д.; Дуань, Юн; Лайонс, Тимоти В.; Арнольд, Гейл Л.; Кендалл, Брайан; Кризер, Роберт А.; Кауфман, Алан Дж.; Гордон, Гвинет В.; Скотт, Клинтон (28 сентября 2007 г.). «Запах кислорода перед великим событием окисления?». Наука . 317 (5846): 1903–1906. Бибкод : 2007Sci...317.1903A . дои : 10.1126/science.1140325 . ISSN 1095-9203 . ПМИД 17901330 . S2CID 25260892 .
- ^ Киршвинк, Джозеф Л.; Копп, Роберт Э (27 августа 2008 г.). «Палеопротерозойские ледяные дома и эволюция ферментов, опосредующих кислород: доводы в пользу позднего происхождения фотосистемы II» . Философские труды Королевского общества B: Биологические науки . 363 (1504): 2755–2765. дои : 10.1098/rstb.2008.0024 . ISSN 0962-8436 . ПМК 2606766 . ПМИД 18487128 .
- ^ Сверженский, Дмитрий А.; Ли, Намхи (01 февраля 2010 г.). «Великое событие окисления и диверсификация полезных ископаемых». Элементы . 6 (1): 31–36. дои : 10.2113/gselements.6.1.31 . ISSN 1811-5209 .
- ^ Джонсон, Йена Э.; Герфейда, Айя; Лэмб, Майкл П.; Фишер, Вудворд В. (2014). «Ограничения O 2 из палеопротерозойского обломочного пирита и уранинита». Бюллетень Геологического общества Америки . 126 (5–6): 813–830. Бибкод : 2014GSAB..126..813J . дои : 10.1130/b30949.1 .
- ^ Клауд, П. (1973). «Палеоэкологическое значение полосчатой железистой формации». Экономическая геология . 68 (7): 1135–1143. Бибкод : 1973EcGeo..68.1135C . дои : 10.2113/gsecongeo.68.7.1135 .
- ^ Цзяо, Юнцинь; Капплер, Андреас; Кроал, Лаура Р.; Ньюман, Дайан К. (1 августа 2005 г.). «Выделение и характеристика генетически поддающейся фотоавтотрофной Fe(II)-окисляющей бактерии, штамма Rhodopseudomonas palustris TIE-1» . Прикладная и экологическая микробиология . 71 (8): 4487–4496. Бибкод : 2005ApEnM..71.4487J . дои : 10.1128/АЕМ.71.8.4487-4496.2005 . ISSN 0099-2240 . ПМЦ 1183355 . PMID 16085840 .
- ^ Кроу, Шон А.; Джонс, КэрриЭйн; Кацев, Сергей; Маген, Седрик; О'Нил, Эндрю Х.; Штурм, Арне; Кэнфилд, Дональд Э.; Хаффнер, Г. Дуглас; Муччи, Альфонсо (14 октября 2008 г.). «Фотоферротрофы процветают в аналоге архейского океана» . Труды Национальной академии наук . 105 (41): 15938–15943. Бибкод : 2008PNAS..10515938C . дои : 10.1073/pnas.0805313105 . ISSN 0027-8424 . ПМК 2572968 . ПМИД 18838679 .
- ^ Капплер, Андреас; Паскеро, Клаудия; Конхаузер, Курт О.; Ньюман, Дайан К. (2005). «Отложение полосчатых железных образований аноксигенными фототрофными Fe(II)-окисляющими бактериями». Геология . 33 (11): 865. Бибкод : 2005Гео....33..865К . дои : 10.1130/g21658.1 .
- ^ Jump up to: Перейти обратно: а б Конхаузер, Курт О.; Хамаде, Тристан; Рэйсуэлл, Роб; Моррис, Ричард С.; Феррис, Ф. Грант; Саутэм, Гордон; Кэнфилд, Дональд Э. (1 декабря 2002 г.). «Могли ли бактерии образовать докембрийские полосчатые железные образования?». Геология . 30 (12): 1079–1082. Бибкод : 2002Geo....30.1079K . doi : 10.1130/0091-7613(2002)030<1079:CBHFTP>2.0.CO;2 . ISSN 0091-7613 .
- ^ Кэрнс-Смит, АГ (21 декабря 1978 г.). «Фотохимия докембрийского раствора, обратная сегрегация и полосчатые образования железа». Природа . 276 (5690): 807–808. Бибкод : 1978Natur.276..807C . дои : 10.1038/276807a0 . S2CID 4315423 .
- ^ Фаркуар, ноль; Бао, ноль; Тименс, ноль (4 августа 2000 г.). «Атмосферное влияние самого раннего цикла серы на Земле». Наука . 289 (5480): 756–759. Бибкод : 2000Sci...289..756F . дои : 10.1126/science.289.5480.756 . ISSN 1095-9203 . ПМИД 10926533 . S2CID 12287304 .
- ^ Копп, Роберт Э.; Киршвинк, Джозеф Л.; Хилберн, Исаак А.; Нэш, Коди З. (9 августа 2005 г.). «Палеопротерозойская Земля-снежок: климатическая катастрофа, вызванная эволюцией кислородного фотосинтеза» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (32): 11131–11136. Бибкод : 2005PNAS..10211131K . дои : 10.1073/pnas.0504878102 . ISSN 0027-8424 . ПМЦ 1183582 . ПМИД 16061801 .
- ^ Пан, Н.; Имлей, Дж.А. (01 марта 2001 г.). «Как кислород подавляет центральный метаболизм у облигатных анаэробов Bacteroides thetaiotaomicron» . Молекулярная микробиология . 39 (6): 1562–1571. дои : 10.1046/j.1365-2958.2001.02343.x . ISSN 0950-382X . ПМИД 11260473 .
- ^ Паттерсон, Клэр (1 октября 1956 г.). «Эпоха метеоритов и Земли». Geochimica et Cosmochimica Acta . 10 (4): 230–237. Бибкод : 1956GeCoA..10..230P . дои : 10.1016/0016-7037(56)90036-9 .
- ^ Уайльд, ЮАР; Вэлли, JW; Пек, штат Вашингтон; Грэм, CM (11 января 2001 г.). «Свидетельства обломочных цирконов о существовании континентальной коры и океанов на Земле 4,4 миллиарда лет назад». Природа . 409 (6817): 175–178. Бибкод : 2001Natur.409..175W . дои : 10.1038/35051550 . ISSN 0028-0836 . ПМИД 11196637 . S2CID 4319774 .
- ^ Тардуно, Джон А.; Блэкман, Эрик Г.; Мамаек, Эрик Э. (01 августа 2014 г.). «Обнаружение старейшего геодинамо и сопутствующей защиты от солнечного ветра: последствия для обитаемости». Физика Земли и недр планет . 233 : 68–87. arXiv : 1502.04410 . Бибкод : 2014PEPI..233...68T . дои : 10.1016/j.pepi.2014.05.007 . S2CID 118656570 .
- ^ Ларсон, Роджер Л.; Хильда, Томас У.К. (10 июня 1975 г.). «Пересмотренная временная шкала инверсий магнитного поля для раннего мела и поздней юры». Журнал геофизических исследований . 80 (17): 2586–2594. Бибкод : 1975JGR....80.2586L . дои : 10.1029/JB080i017p02586 . ISSN 2156-2202 .
- ^ Карри, Дж.А.; Шрамм, Дж.Л.; Эберт, Э.Э. (1 января 1996 г.). «Механизм климатической обратной связи по альбедо морского льда» . Обзор океанографической литературы . 2 (43). ISSN 0967-0653 .
- ^ Уокер, Джеймс К.Г.; Хейс, П.Б.; Кастинг, Дж. Ф. (20 октября 1981 г.). «Механизм отрицательной обратной связи для долгосрочной стабилизации температуры поверхности Земли». Журнал геофизических исследований: Океаны . 86 (С10): 9776–9782. Бибкод : 1981JGR....86.9776W . дои : 10.1029/JC086iC10p09776 . ISSN 2156-2202 .
- ^ Фойлнер, Георг (01 июня 2012 г.). «Проблема слабого молодого Солнца». Обзоры геофизики . 50 (2): РГ2006. arXiv : 1204.4449 . Бибкод : 2012RvGeo..50.2006F . дои : 10.1029/2011RG000375 . ISSN 1944-9208 . S2CID 119248267 .
- ^ «Что такое ген?» . Домашний справочник по генетике . Проверено 26 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б с Ней М. и Кумар С. (2000). Молекулярная эволюция и филогенетика . Издательство Оксфордского университета.
- ^ Смит, Эндрю Б.; Петерсон, Кевин Дж. (1 января 2002 г.). «Датирование времени происхождения основных клад: молекулярные часы и летопись окаменелостей». Ежегодный обзор наук о Земле и планетах . 30 (1): 65–88. Бибкод : 2002AREPS..30...65S . doi : 10.1146/annurev.earth.30.091201.140057 .
- ^ Добжанский, Феодосий (1 марта 1973). «Ничто в биологии не имеет смысла, кроме как в свете эволюции». Американский учитель биологии . 35 (3): 125–129. CiteSeerX 10.1.1.324.2891 . дои : 10.2307/4444260 . ISSN 0002-7685 . JSTOR 4444260 . S2CID 207358177 .
- ^ Хэнсон, РС; Хэнсон, TE (1 июня 1996 г.). «Метанотрофные бактерии» . Микробиологические обзоры . 60 (2): 439–471. дои : 10.1128/MMBR.60.2.439-471.1996 . ISSN 0146-0749 . ПМК 239451 . ПМИД 8801441 .
- ^ Холмс, Эндрю Дж.; Оуэнс, Ник Дж. П.; Мюррелл, Дж. Колин (1 января 1995 г.). «Обнаружение новых морских метанотрофов с использованием филогенетических и функциональных генных зондов после обогащения метаном» . Микробиология . 141 (8): 1947–1955. дои : 10.1099/13500872-141-8-1947 . ПМИД 7551057 .
- ^ Луескен, Франциска А.; Чжу, Баоли; Ален, Тео А. ван; Батлер, Маргарет К.; Диас, Марина Родригес; Сон, Понкын; Кэмп, Хууб Дж.М. Опубликовано; Джеттен, Майк С.М.; Эттвиг, Катарина Ф. (01 июня 2011 г.). «Праймеры pmoA для обнаружения анаэробных метанотрофов» . Прикладная и экологическая микробиология . 77 (11): 3877–3880. Бибкод : 2011ApEnM..77.3877L . дои : 10.1128/АЕМ.02960-10 . ISSN 0099-2240 . ПМК 3127593 . ПМИД 21460105 .
- ^ Jump up to: Перейти обратно: а б Ризенфельд, Кристиан С.; Шлосс, Патрик Д.; Хандельсман, Джо (1 января 2004 г.). «Метагеномика: геномный анализ микробных сообществ». Ежегодный обзор генетики . 38 : 525–552. CiteSeerX 10.1.1.526.482 . дои : 10.1146/annurev.genet.38.072902.091216 . ISSN 0066-4197 . ПМИД 15568985 .
- ^ Jump up to: Перейти обратно: а б Гротцингер, Джон П.; Нолл, Эндрю Х. (1999). «СТРОМАТОЛИТЫ В ДОКАМБРИЙСКИХ КАРБОНАТАХ: Эволюционные вехи или экологические ориентиры?». Ежегодный обзор наук о Земле и планетах . 27 (1): 313–358. Бибкод : 1999AREPS..27..313G . дои : 10.1146/annurev.earth.27.1.313 . ПМИД 11543060 .
- ^ Хофманн, HJ; Грей, К.; Хикман, А.Х.; Торп, Род-Айленд (1 августа 1999 г.). «Хофманн Х.Дж., Грей К., Хикман А.Х. и Торп Р.И. Происхождение конуформных строматолитов возрастом 3,45 млрд лет в группе Варравуна, Западная Австралия. Geol. Soc. Am. Bull. 111, 1256-1262» . Бюллетень Геологического общества Америки . 111 (8): 1256–1262. Бибкод : 1999GSAB..111.1256H . doi : 10.1130/0016-7606(1999)111<1256:OOGCSI>2.3.CO;2 . ISSN 0016-7606 .
- ^ Берг, Дж. М., Тимочко, Дж. Л., и Страйер, Л. (2002). Биохимия. 5-й.
- ^ Jump up to: Перейти обратно: а б с Л, Слончевски, Джоан; В., Фостер, Джон (01 октября 2013 г.). Микробиология: развивающаяся наука: третье международное студенческое издание . WW Нортон и компания. ISBN 9780393923216 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Стабильные изотопы и взаимоотношения растительного углерода и воды . Эльзевир. 2012-12-02. ISBN 9780080918013 .
- ^ Jump up to: Перейти обратно: а б Макколлом, Томас М. (1 января 2013 г.). «Миллер-Юри и не только: что мы узнали о реакциях органического синтеза пребиотиков за последние 60 лет?». Ежегодный обзор наук о Земле и планетах . 41 (1): 207–229. Бибкод : 2013AREPS..41..207M . doi : 10.1146/annurev-earth-040610-133457 .
- ^ Шарф, Калеб; Дева, Натаниэль; Кливс, Х. Джеймс; Аоно, Масаси; Обер-Като, Натанаэль; Айдыноглу, Арсев; Бараона, Ана; Бардж, Лаура М.; Беннер, Стивен А. (1 декабря 2015 г.). «Стратегия исследования происхождения жизни» . Астробиология . 15 (12): 1031–1042. Бибкод : 2015AsBio..15.1031S . дои : 10.1089/ast.2015.1113 . ISSN 1531-1074 . ПМЦ 4683543 . ПМИД 26684503 .
- ^ «Астробиология НАСА» . astrobiology.nasa.gov . Проверено 26 мая 2016 г.
- ^ Лавлок, Джеймс (28 сентября 2000 г.). Гайя: новый взгляд на жизнь на Земле . ОУП Оксфорд. ISBN 9780191606694 .
- ^ Шок, Эверетт Л.; Бойд, Эрик С. (1 декабря 2015 г.). «Принципы геобиохимии». Элементы . 11 (6): 395–401. дои : 10.2113/gselements.11.6.395 . ISSN 1811-5209 .
- ^ Друшель, Грегори К.; Капплер, Андреас (01 декабря 2015 г.). «Геомикробиология и микробная геохимия». Элементы . 11 (6): 389–394. дои : 10.2113/gselements.11.6.389 . hdl : 1805/10102 . ISSN 1811-5209 .
- ^ «Геомикробиология» . Университет Хельсинки . 27 января 2017 г. Проверено 9 февраля 2020 г.
- ^ Брокс, Йохен Дж.; Логан, Грэм А.; Бьюик, Роджер; Вызов, Роджер Э. (13 августа 1999 г.). «Архейские молекулярные окаменелости и раннее появление эукариотов». Наука . 285 (5430): 1033–1036. Бибкод : 1999Sci...285.1033B . CiteSeerX 10.1.1.516.9123 . дои : 10.1126/science.285.5430.1033 . ISSN 0036-8075 . ПМИД 10446042 .
- ^ Нолл, Эндрю Х. (22 марта 2015 г.). Жизнь на молодой планете: первые три миллиарда лет эволюции на Земле . Издательство Принстонского университета. ISBN 9781400866045 .
- ^ Эваристо, Хайвиме; Ясечко, Скотт; Макдоннелл, Джеффри Дж. (2015). «Глобальное отделение транспирации растений от грунтовых вод и речного стока». Природа . 525 (7567): 91–94. Бибкод : 2015Natur.525...91E . дои : 10.1038/nature14983 . ПМИД 26333467 . S2CID 4467297 .
- ^ Хорошо, Стивен П.; Никто, Дэвид; Боуэн, Габриэль (10 июля 2015 г.). «Гидрологическая связь ограничивает разделение глобальных потоков наземных вод» . Наука . 349 (6244): 175–177. Бибкод : 2015Sci...349..175G . дои : 10.1126/science.aaa5931 . ISSN 0036-8075 . ПМИД 26160944 .
Внешние ссылки [ править ]
- Геобиология, Научный журнал
- Геобиология в Калифорнийском технологическом институте
- Геобиология в Университете Южной Калифорнии. Архивировано 25 февраля 2020 г. в Wayback Machine.
- Геобиология в Массачусетском технологическом институте. Архивировано 9 ноября 2019 г. в Wayback Machine.
- Геобиология и история Земли в Гарварде
- Геобиология в Стэнфорде. Архивировано 22 июля 2016 г. в Wayback Machine.
- Геобиология в Калифорнийском университете в Лос-Анджелесе
- Agouron - Летние курсы по геобиологии, спонсируемые Университетом Южной Калифорнии, на острове Каталина
- Геобиология в Техасском университете A&M. Архивировано 22 ноября 2015 г. в Wayback Machine.
- Геологическое общество Америки - геобиология. Архивировано 9 ноября 2019 г. в Wayback Machine.

