метанотроф
Метанотрофы (иногда называемые метанофилами ) — это прокариоты , которые метаболизируют метан как источник углерода и химической энергии . Это бактерии или археи , они могут расти аэробно или анаэробно и для выживания им требуются одноуглеродные соединения .
Метанотрофы особенно распространены в средах, где производится метан, или вблизи них, хотя некоторые метанотрофы могут окислять атмосферный метан . Их среда обитания включает водно-болотные угодья, почвы, болота, рисовые поля, свалки, водные системы (озера, океаны, ручьи) и многое другое. Они представляют особый интерес для исследователей, изучающих глобальное потепление , поскольку играют значительную роль в глобальном балансе метана, уменьшая количество метана, выбрасываемого в атмосферу. [1] [2]
Метанотрофия — это особый случай метилотрофии , при котором используются одноуглеродные соединения, которые более восстановлены, чем углекислый газ. Однако некоторые метилотрофы также могут использовать многоуглеродные соединения; это отличает их от метанотрофов, которые обычно являются привередливыми окислителями метана и метанола. Единственными факультативными метанотрофами, выделенными на сегодняшний день, являются представители рода Mmethylocella silvestris . [3] [4] Метилокапса золотистая [5] и несколько штаммов Mmethylocystis . [6]
В функциональном отношении метанотрофы называются метанокисляющими бактериями. Однако к метанокисляющим бактериям относятся и другие организмы, которые не считаются единственными метанотрофами. По этой причине метанокисляющие бактерии были разделены на подгруппы: группы метанассимилирующих бактерий (МАБ), метанотрофы и автотрофные аммиакоокисляющие бактерии (ААОБ), соокисляющие метан. [2]
Классификация [ править ]
Метанотрофами могут быть как бактерии , так и археи . Какой вид метанотрофов присутствует, главным образом определяется наличием акцепторов электронов . Известны многие виды метанокисляющих бактерий (МОБ). Различия в способе фиксации формальдегида и строении мембран делят этих бактериальных метанотрофов на несколько групп. Среди метанотрофных архей выделяют несколько подгрупп.
Аэробика [ править ]
В аэробных условиях метанотрофы объединяют кислород и метан с образованием формальдегида , который затем включается в органические соединения по сериновому пути или по пути рибулозомонофосфата (RuMP), а также высвобождается углекислый газ. Метанотрофы типов I и X являются частью гаммапротеобактерий и используют путь RuMP для ассимиляции углерода. Метанотрофы типа II являются частью альфапротеобактерий и используют сериновый путь ассимиляции углерода. Для них также характерна система внутренних мембран, внутри которых происходит окисление метана . Метанотрофы у гаммапротеобактерий известны из семейства Mmethylococcaceae . [7] Метанотрофы из Alphaproteobacteria встречаются в семействах Mmethylocystaceae и Beijerinckiaceae .
Аэробные метанотрофы также известны из Mmethylacidiphilaceae (тип Verrucomicrobiota ). [8] В отличие от гаммапротеобактерий и альфапротеобактерий метанотрофы типа Verrucomicrobiota являются миксотрофами . [9] [10] В 2021 году был обнаружен бактериальный контейнер из типа Gemmatimonadota под названием « Candidatus Mmethylotropicum kingii», демонстрирующий аэробную метанотрофию, что позволяет предположить, что метанотрофия присутствует в четырех бактериальных типах. [11]
В некоторых случаях аэробное окисление метана может происходить в бескислородной среде. « Candidatus Mmethylomirabilis oxyfera » принадлежит к типу бактерий NC10 и может катализировать восстановление нитритов по «внутриаэробному» пути, при котором вырабатываемый внутри организма кислород используется для окисления метана. [12] [13] В озерах с чистой водой метанотрофы могут жить в бескислородной толще воды, но получают кислород от фотосинтезирующих организмов, который затем напрямую потребляют для окисления метана. [14]
Аэробные метанотрофные археи не известны.
Анаэробный [ править ]
В бескислородных условиях метанотрофы используют разные акцепторы электронов для окисления метана. Это может произойти в бескислородных средах обитания, таких как морские или озерные отложения , зоны минимума кислорода , бескислородные водные столбы, рисовые поля и почвы. Некоторые специфические метанотрофы могут восстанавливать нитраты, [15] нитрит, [16] железо, [17] сульфат, [18] или ионы марганца и соединяют их с окислением метана без синтрофного партнера. Исследования в морской среде показали, что метан может окисляться анаэробно консорциумами метанокисляющих архей и сульфатредуцирующих бактерий . [19] [20] Этот тип анаэробного окисления метана (АОМ) преимущественно происходит в бескислородных морских отложениях. Точный механизм до сих пор остается предметом споров, но наиболее широко распространенная теория заключается в том, что археи используют обратный путь метаногенеза для производства углекислого газа и другого, неизвестного промежуточного продукта, который затем используется сульфатредуцирующими бактериями для получения энергии за счет восстановления. сульфата до сероводорода и воды.
Анаэробные метанотрофы не связаны с известными аэробными метанотрофами; Ближайшими культурными родственниками анаэробных метанотрофов являются метаногены из отряда Methanosarcinales . [21]
Особый вид [ править ]
Mmethylococcus capsulatus используется для производства кормов для животных из природного газа. [22]
В 2010 году была идентифицирована новая бактерия Candidatus Mmethylomirabilis oxyfera из типа NC10 , которая может сочетать анаэробное окисление метана с восстановлением нитрита без необходимости синтрофного партнера. [12] На основании исследований Эттвига и др., [12] Считается, что M. oxyfera окисляет метан анаэробно, используя кислород, образующийся внутри организма в результате дисмутации оксида азота на азот и газообразный кислород.
Таксономия [ править ]
Многие метанотрофные культуры были изолированы и официально охарактеризованы за последние 5 десятилетий, начиная с классического исследования Уиттенбери (Whittenbury et al., 1970). 18 родов культивируемых аэробных метанотрофных гаммапротеобактерий и 5 родов альфапротеобактерий В настоящее время известно , представленных ок. 60 различных видов. [23]
Окисление метана [ править ]
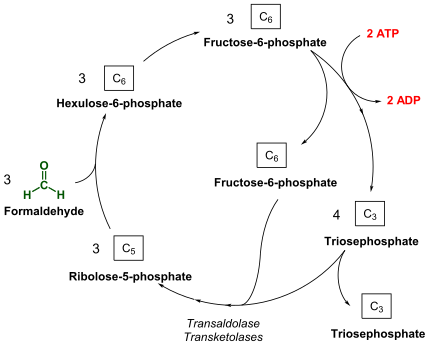

Метанотрофы окисляют метан, сначала инициируя восстановление дикислорода до H 2 O 2 и преобразование метана в CH 3 OH с помощью метанмонооксигеназ (ММО) . [24] Кроме того, из метанотрофов были выделены два типа ММО: растворимая метанмонооксигеназа (sMMO) и метанмонооксигеназа в виде частиц (pMMO) .
Клетки, содержащие pMMO, продемонстрировали более высокие способности к росту и более высокое сродство к метану, чем клетки, содержащие sMMO. [24] Предполагается, что ионы меди могут играть ключевую роль как в регуляции pMMO, так и в ферментативном катализе, тем самым ограничивая клетки pMMO средой, более богатой медью, чем клетки, продуцирующие sMMO. [25]
См. также [ править ]
Ссылки [ править ]
- ^ Оремланд Р.С. , Калбертсон К.В. (1992). «Значение метанокисляющих бактерий в балансе метана, выявленное с помощью специфического ингибитора». Природа . 356 (6368): 421–423. Бибкод : 1992Natur.356..421O . дои : 10.1038/356421a0 . S2CID 4234351 .
- ^ Jump up to: Перейти обратно: а б Холмс А.Дж., Рослев П., Макдональд И.Р., Иверсен Н., Хенриксен К., Мюррел Дж.К. (август 1999 г.). «Характеристика популяций метанотрофных бактерий в почвах, демонстрирующих поглощение метана из атмосферы» . Прикладная и экологическая микробиология . 65 (8): 3312–8. Бибкод : 1999ApEnM..65.3312H . дои : 10.1128/АЕМ.65.8.3312-3318.1999 . ПМК 91497 . ПМИД 10427012 .
- ^ Дедыш С.Н., Книф С., Данфилд П.Ф. (июль 2005 г.). «Виды метилоцелл факультативно метанотрофны» . Журнал бактериологии . 187 (13): 4665–70. дои : 10.1128/JB.187.13.4665-4670.2005 . ПМЦ 1151763 . ПМИД 15968078 .
- ^ Чен Ю., Кромби А., Рахман М.Т., Дедыш С.Н., Лисак В., Стотт М.Б. и др. (июль 2010 г.). «Полная последовательность генома аэробного факультативного метанотрофа Mmethylocella silvestris BL2» . Журнал бактериологии . 192 (14): 3840–1. дои : 10.1128/JB.00506-10 . ПМЦ 2897342 . ПМИД 20472789 .
- ^ Данфилд П.Ф., Белова С.Е., Воробьев А.В., Корниш С.Л., Дедыш С.Н. (ноябрь 2010 г.). «Mmethylocapsa aurea sp. nov., факультативный метанотроф, обладающий метанмонооксигеназой в виде частиц, и исправленное описание рода Mmethylocapsa» . Международный журнал систематической и эволюционной микробиологии . 60 (Часть 11): 2659–2664. дои : 10.1099/ijs.0.020149-0 . ПМИД 20061505 .
- ^ Белова С.Е., Баани М., Сузина Н.Е., Боделье П.Л., Лизак В., Дедыш С.Н. (февраль 2011 г.). «Утилизация ацетата как стратегия выживания обитающих в торфе видов Mmethylocystis» . Отчеты по экологической микробиологии . 3 (1): 36–46. дои : 10.1111/j.1758-2229.2010.00180.x . ПМИД 23761229 .
- ^ Стейн Л.И., Рой Р., Данфилд П.Ф. (16 апреля 2012 г.). «Аэробная метанотрофия и нитрификация: процессы и связи». ЭЛС . Чичестер, Великобритания: John Wiley & Sons, Ltd., стр. a0022213. дои : 10.1002/9780470015902.a0022213 . ISBN 978-0-470-01617-6 . Проверено 17 января 2021 г.
- ^ Оп ден Кэмп Х.Дж., Ислам Т., Стотт М.Б., Харханги Х.Р., Хайнс А., Схаутен С. и др. (октябрь 2009 г.). «Экологические, геномные и таксономические перспективы метанотрофной веррукомикробии» . Отчеты по экологической микробиологии . 1 (5): 293–306. дои : 10.1111/j.1758-2229.2009.00022.x . hdl : 2066/75111 . ПМИД 23765882 .
- ^ Carere CR, Hards K, Houghton KM, Power JF, McDonald B, Collet C и др. (ноябрь 2017 г.). «Миксотрофия способствует расширению ниши веррукомикробных метанотрофов» . Журнал ISME . 11 (11): 2599–2610. дои : 10.1038/ismej.2017.112 . ПМК 5649168 . ПМИД 28777381 .
- ^ Шарп CE, Стотт МБ, Данфилд П.Ф. (2012). «Обнаружение автотрофных веррукомикробных метанотрофов в геотермальной среде с помощью зондирования стабильными изотопами» . Границы микробиологии . 3 : 303. дои : 10.3389/fmicb.2012.00303 . ПМЦ 3421453 . ПМИД 22912630 .
- ^ Бэй С.К., Донг X, Брэдли Дж.А., Люнг П.М., Гринтер Р., Джирапанджават Т. и др. (январь 2021 г.). «Микрогазовые окислители — широко распространенные и активные члены почвенных микробных сообществ» . Природная микробиология . 6 (2): 246–256. doi : 10.1038/s41564-020-00811-w . ПМИД 33398096 . S2CID 230663681 .
- ^ Jump up to: Перейти обратно: а б с Эттвиг К.Ф., Батлер М.К., Ле Паслье Д., Пеллетье Э., Манжено С., Кайперс М.М. и др. (март 2010 г.). «Анаэробное окисление метана нитритами оксигенными бактериями» (PDF) . Природа . 464 (7288): 543–8. Бибкод : 2010Natur.464..543E . дои : 10.1038/nature08883 . ПМИД 20336137 . S2CID 205220000 .
- ^ Жу Б., ван Дейк Г., Фриц С., Смолдерс А.Дж., Пол А., Джеттен М.С., Эттвиг К.Ф. (декабрь 2012 г.). «Анаэробное окисление метана в минеротрофном торфянике: обогащение нитритзависимыми метанокисляющими бактериями» . Прикладная и экологическая микробиология . 78 (24): 8657–65. Бибкод : 2012ApEnM..78.8657Z . дои : 10.1128/АЕМ.02102-12 . ПМЦ 3502929 . ПМИД 23042166 .
- ^ Милука Дж., Кирф М., Лу Л., Крупке А., Лам П., Литтманн С. и др. (сентябрь 2015 г.). «Окисление метана в сочетании с кислородным фотосинтезом в бескислородных водах» . Журнал ISME . 9 (9): 1991–2002. дои : 10.1038/ismej.2015.12 . ПМК 4542029 . ПМИД 25679533 .
- ^ Харун М.Ф., Ху С., Ши Ю., Имелфорт М., Келлер Дж., Хугенхольц П. и др. (август 2013 г.). «Анаэробное окисление метана в сочетании с восстановлением нитратов у новой линии архей». Природа . 500 (7464): 567–70. Бибкод : 2013Natur.500..567H . дои : 10.1038/nature12375 . ПМИД 23892779 . S2CID 4368118 .
- ^ Эттвиг К.Ф., Батлер М.К., Ле Паслье Д., Пеллетье Э., Манжено С., Кайперс М.М. и др. (март 2010 г.). «Анаэробное окисление метана нитритами оксигенными бактериями» . Природа . 464 (7288): 543–8. Бибкод : 2010Natur.464..543E . дои : 10.1038/nature08883 . hdl : 2066/84284 . ПМИД 20336137 . S2CID 205220000 .
- ^ Эттвиг К.Ф., Чжу Б., Спет Д., Келтьенс Дж.Т., Джеттен М.С., Картал Б. (ноябрь 2016 г.). «Археи катализируют железозависимое анаэробное окисление метана» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (45): 12792–12796. Бибкод : 2016PNAS..11312792E . дои : 10.1073/pnas.1609534113 . ПМК 5111651 . ПМИД 27791118 .
- ^ Милука Дж., Фердельман Т.Г., Полерецкий Л., Францке Д., Вегенер Г., Шмид М. и др. (ноябрь 2012 г.). «Нульвалентная сера является ключевым промежуточным продуктом в окислении морского метана» . Природа . 491 (7425): 541–6. Бибкод : 2012Natur.491..541M . дои : 10.1038/nature11656 . ПМИД 23135396 . S2CID 32356495 .
- ^ Оффре П., Спанг А., Шлепер С. (8 сентября 2013 г.). «Археи в биогеохимических циклах». Ежегодный обзор микробиологии . 67 (1): 437–57. doi : 10.1146/annurev-micro-092412-155614 . ПМИД 23808334 .
- ^ Тауэр РК (июнь 2011 г.). «Анаэробное окисление метана сульфатом: об обратимости реакций, катализируемых ферментами, также участвующими в метаногенезе из СО2». Современное мнение в микробиологии . 14 (3): 292–9. дои : 10.1016/j.mib.2011.03.003 . ПМИД 21489863 .
- ^ Боэций А. , Равеншлаг К., Шуберт С.Дж., Рикерт Д., Виддель Ф., Гизеке А. и др. (октябрь 2000 г.). «Морской микробный консорциум, по-видимому, опосредующий анаэробное окисление метана» . Природа . 407 (6804): 623–6. Бибкод : 2000Natur.407..623B . дои : 10.1038/35036572 . ПМИД 11034209 . S2CID 205009562 .
- ^ Ле Пейдж М (19 ноября 2016 г.). «Пища, приготовленная из природного газа, скоро будет кормить сельскохозяйственных животных – и нас» . Новый учёный . Проверено 11 декабря 2016 г.
- ^ Ората Ф.Д., Мейер-Колтхофф Дж.П., Соважо Д., Стейн Л.Ю. (2018). «Филогеномный анализ гаммапротеобактериальных метанотрофов (отряда Mmethylococcales ) требует реклассификации членов на уровне рода и вида» . Границы микробиологии . 9 : 3162. дои : 10.3389/fmicb.2018.03162 . ПМК 6315193 . ПМИД 30631317 .
- ^ Jump up to: Перейти обратно: а б Хэнсон Р.С., Хэнсон Т.Э. (июнь 1996 г.). «Метанотрофные бактерии» . Микробиологические обзоры . 60 (2): 439–71. дои : 10.1128/MMBR.60.2.439-471.1996 . ПМК 239451 . ПМИД 8801441 .
- ^ Либерман Р.Л., Розенцвейг AC (2004). «Биологическое окисление метана: регуляция, биохимия и структура активного центра метанмонооксигеназы твердых частиц». Критические обзоры по биохимии и молекулярной биологии . 39 (3): 147–64. дои : 10.1080/10409230490475507 . ПМИД 15596549 . S2CID 21628195 .
Внешние ссылки [ править ]
- Анаэробное окисление метана
- Насекомое, питающееся метаном, обещает сократить выбросы парниковых газов Пресс-релиз, GNS Science, Новая Зеландия