Водоросли
| Водоросли Неофициальный термин для разнообразной группы фотосинтезирующих эукариот. Временной диапазон: | |
|---|---|
 | |
| Разнообразие водорослей, растущих на морском дне на мелководье | |
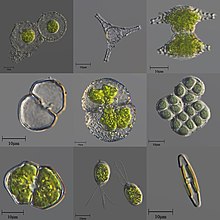 | |
| Разнообразие микроскопических одноклеточных и колониальных пресноводных водорослей. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Группы включены | |
| |
| Обычно исключается | |
| |
Водоросли ( Великобритания : / ˈ æ l ɡ iː / AL -ghee , США : / ˈ æ l dʒ iː / AL -jee ; [3] сг. : alga / ˈ æ l ɡ ə / AL -gə — любые представители большой и разнообразной группы фотосинтезирующих эукариотических ) организмов. Название является неофициальным термином для полифилетической группы, включающей виды из нескольких различных клад . Включенные организмы варьируются от одноклеточных микроводорослей , таких как хлорелла , прототека и диатомовые водоросли , до многоклеточных форм, таких как гигантская бурая водоросль , большая бурая водоросль , которая может вырасти до 50 метров (160 футов) в длину. Большинство из них обитают в воде и лишены многих отдельных типов клеток и тканей, таких как устьица , ксилема и флоэма , которые встречаются у наземных растений . Самые крупные и сложные морские водоросли называются морскими водорослями , а наиболее сложными пресноводными формами являются Charophyta , подразделение зеленых водорослей , в которое входят, например, спирогира и костянки . Водоросли, переносимые водой, — это планктон , в частности фитопланктон .
Водоросли составляют полифилетическую группу. [4] поскольку они не имеют общего предка, и хотя их пластиды, по-видимому, имеют единое происхождение - от цианобактерий, [5] они были приобретены разными способами. Зеленые водоросли являются примерами водорослей, у которых есть первичные хлоропласты, полученные из эндосимбиотических цианобактерий. Диатомеи и бурые водоросли являются примерами водорослей со вторичными хлоропластами, происходящими от эндосимбиотических красных водорослей . [6] Водоросли демонстрируют широкий спектр репродуктивных стратегий: от простого бесполого деления клеток до сложных форм полового размножения . [7]
У водорослей отсутствуют различные структуры, характерные для наземных растений , такие как филлиды (листоподобные структуры) мохообразных , ризоиды несосудистых растений , а также корни , листья и другие органы, обнаруженные у трахеофитов ( сосудистых растений ). Большинство из них фототрофны , хотя некоторые и миксотрофны , получая энергию как за счет фотосинтеза, так и за счет поглощения органического углерода путем осмотрофии , мизотрофии или фаготрофии . Некоторые одноклеточные виды зеленых водорослей , многие золотистые водоросли , эвглениды , динофлагелляты и другие водоросли стали гетеротрофами (также называемыми бесцветными или апохлоротическими водорослями), иногда паразитическими, полностью полагающимися на внешние источники энергии и имеющими ограниченный или отсутствующий фотосинтетический аппарат. [8] [9] [10] Некоторые другие гетеротрофные организмы, такие как apicomplexans , также произошли от клеток, предки которых обладали пластидами , но традиционно не считаются водорослями. Водоросли имеют фотосинтетический аппарат, в конечном итоге полученный от цианобактерий , которые производят кислород в качестве побочного продукта фотосинтеза, в отличие от других фотосинтезирующих бактерий, таких как пурпурные и зеленые серные бактерии . Ископаемые нитчатые водоросли из бассейна Виндхья датируются периодом от 1,6 до 1,7 миллиардов лет назад. [11]
Из-за широкого спектра типов водорослей они находят все большее применение в различных промышленных и традиционных целях в человеческом обществе. Традиционные методы выращивания морских водорослей существуют уже тысячи лет и имеют сильные традиции в Восточной Азии пищевых культурах . Более современные применения альгакультуры расширяют пищевые традиции для других целей, включая корм для скота, использование водорослей для биовосстановления или борьбы с загрязнением, преобразование солнечного света в топливо из водорослей или другие химические вещества, используемые в промышленных процессах, а также в медицинских и научных целях. Обзор 2020 года показал, что такое применение водорослей может сыграть важную роль в связывании углерода , чтобы смягчить последствия изменения климата , одновременно обеспечивая прибыльную продукцию с добавленной стоимостью для мировой экономики. [12]
и изучение Этимология
Единственное число «водоросль» — это латинское слово, означающее «водоросли», и оно сохраняет это значение в английском языке. [13] Этимология неясна . Хотя некоторые предполагают, что это слово связано с латинским algēre , «быть холодным». [14] неизвестна причина, по которой водоросли связывают с температурой. Более вероятный источник — alliga , «связывание, переплетение». [15]
Древнегреческое . слово «водоросли» было φῦκος ( phŷkos ), что могло означать либо морские водоросли (вероятно, красные водоросли), либо красный краситель, полученный из них Латинизация fucus означала прежде всего косметические румяна. Этимология неясна, но сильным кандидатом уже давно является какое-то слово, связанное с библейским פוך ( пук ), «краска» (если не само это слово), косметические тени для век , используемые древними египтянами и другими жителями восточного региона. Средиземноморье. Это может быть любой цвет: черный, красный, зеленый или синий. [16]
Изучение водорослей чаще всего называют фикологией (от греческого phykos «водоросли»); термин альгология выходит из употребления. [17]
Классификации [ править ]
Одно из определений водорослей состоит в том, что они «имеют хлорофилл в качестве основного фотосинтетического пигмента и лишены стерильного покрытия вокруг своих репродуктивных клеток ». [18] С другой стороны, бесцветные прототеки под Chlorophyta лишены хлорофилла. Хотя цианобактерии часто называют «сине-зелеными водорослями», большинство авторов исключают все прокариоты , включая цианобактерии, из определения водорослей. [4] [19]
Водоросли содержат хлоропласты , по строению сходные с цианобактериями. Хлоропласты содержат кольцевую ДНК, подобную ДНК цианобактерий, и интерпретируются как редуцированные эндосимбиотические цианобактерии . Однако точное происхождение хлоропластов у разных линий водорослей различно, что отражает их приобретение в ходе разных эндосимбиотических событий. В таблице ниже описан состав трех основных групп водорослей. Их родословные отношения показаны на рисунке в правом верхнем углу. Многие из этих групп содержат представителей, которые больше не являются фотосинтезирующими. Некоторые сохраняют пластиды, но не хлоропласты, а другие полностью утрачивают пластиды. [20]
Филогения на основе пластид [21] не нуклеоцитоплазматическая генеалогия:
| |||||||||||||
| Принадлежность к супергруппе | Члены | Эндосимбионт | Краткое содержание |
|---|---|---|---|
| Примоплантае / Археопластида | Цианобактерии | Эти водоросли имеют «первичные» хлоропласты , то есть хлоропласты окружены двумя мембранами и, вероятно, развились в результате одного эндосимбиотического события. В хлоропластах красных водорослей имеются хлорофиллы а и с (часто) и фикобилины , а у зеленых водорослей — хлоропласты с хлорофиллом а и б без фикобилинов. Наземные растения пигментированы так же, как зеленые водоросли, и, вероятно, развились из них, таким образом, Chlorophyta является родственным таксоном растений; иногда Chlorophyta, Charophyta и наземные растения группируются вместе как Viridiplantae . | |
| Экскавата и Ризария | Зеленые водоросли | Эти группы имеют зеленые хлоропласты, содержащие хлорофиллы a и b . [22] Их хлоропласты окружены четырьмя и тремя мембранами соответственно и, вероятно, остались от проглоченных зеленых водорослей. Хлорарахниофиты , принадлежащие к типу Cercozoa , содержат небольшой нуклеоморф , являющийся остатком водоросли ядра . Эвглениды , принадлежащие к типу Euglenozoa , живут преимущественно в пресной воде и имеют хлоропласты только с тремя мембранами. Эндосимбиотические зеленые водоросли могли быть приобретены посредством мизоцитоза, а не фагоцитоза . [23] (Другая группа эндосимбионтов зеленых водорослей - это род динофлагеллят Lepidodinium , который заменил свой первоначальный эндосимбионт красного водорослевого происхождения на эндосимбионт зеленого водорослевого происхождения. Присутствует нуклеоморф, и геном хозяина все еще содержит несколько генов красных водорослей, приобретенных в результате эндосимбиотического переноса генов. Также геном эвгленид и хлорарахниофитов содержит гены очевидного происхождения от красных водорослей. [24] [25] [26] | |
| Хальвария и Хакробия | Красные водоросли | Эти группы имеют хлоропласты, содержащие хлорофиллы а и с и фикобилины. Форма может варьироваться; они могут быть дискоидными, пластинчатыми, сетчатыми, чашевидными, спиралевидными или лентовидными. У них есть один или несколько пиреноидов для сохранения белка и крахмала. Последний тип хлорофилла не известен ни у каких прокариот или первичных хлоропластов, но генетическое сходство с красными водорослями предполагает его родство. [27] В первых трех из этих групп ( Chromista ) хлоропласты имеют четыре мембраны, сохраняющие нуклеоморф у криптомонад , и они, вероятно, имеют общего пигментированного предка, хотя другие данные ставят под сомнение, являются ли гетероконты , Haptophyta и криптомонады на самом деле более тесно связаны друг с другом, чем с другими группами. [28] [29] Типичный хлоропласт динофлагеллят имеет три мембраны, но внутри группы существует значительное разнообразие хлоропластов, и, по-видимому, произошел ряд эндосимбиотических событий. [5] Apicomplexa , которые не являются фотосинтезирующими, но, по-видимому , , группа близкородственных паразитов, также имеет пластиды, называемые апикопластами имеют общее происхождение с динофлагеллятными хлоропластами. [5] |

Линней в «Видах растений» (1753 г.), [30] Отправной точкой современной ботанической номенклатуры признано 14 родов водорослей, из которых к водорослям в настоящее время относят только четыре. [31] В «Системе природы роды Volvox и Corallina , а также вид Acetabularia (как Madrepora » Линней описал среди животных ).
В 1768 году Самуэль Готлиб Гмелин (1744–1774) опубликовал Historia Fucorum — первую работу, посвященную морским водорослям, и первую книгу по морской биологии, в которой использовалась новая тогда биномиальная номенклатура Линнея. Он включал тщательно продуманные иллюстрации морских водорослей на сложенных листьях. [32] [33]
WH Харви (1811–1866) и Ламуру (1813) [34] были первыми, кто разделил макроскопические водоросли на четыре отдела в зависимости от их пигментации. Это первое использование биохимического критерия в систематике растений. Четыре отдела Харви: красные водоросли (Rhodospermae), бурые водоросли (Melanospermae), зеленые водоросли (Chlorospermae) и диатомовые. [35] [36]
В это время микроскопические водоросли были обнаружены и о них сообщила другая группа работников (например, О. Ф. Мюллер и Эренберг ), изучавшая инфузории (микроскопические организмы). В отличие от макроводорослей , которые явно рассматривались как растения, микроводоросли часто считались животными, поскольку они часто подвижны. [34] Даже неподвижные (кокковидные) микроводоросли иногда рассматривались просто как стадии жизненного цикла растений, макроводорослей или животных. [37] [38]
Хотя он использовался как таксономическая категория в некоторых додарвиновских классификациях, например, Линнея (1753), [39] Жюссье (1789 г.), [40] Ламуру (1813 г.), Харви (1836 г.), Хоранинов (1843 г.), Агассис (1859 г.), Уилсон и Кассен (1864 г.), [39] в дальнейших классификациях «водоросли» рассматриваются как искусственная полифилетическая группа. [41]
На протяжении XX века большинство классификаций рассматривали следующие группы как подразделения или классы водорослей: цианофиты , родофиты , хризофиты , ксантофиты , бацилляриофиты , феофиты , пиррофиты ( криптофиты и динофиты ), эвгленофиты и хлорофиты . Позднее было открыто множество новых групп (например, Bolidophyceae ), а от более старых групп выделились другие: харофиты и глаукофиты (от хлорофитов), многие гетероконтофиты (например, синурофиты от хризофитов или эустигматофиты от ксантофитов), гаптофиты (от хризофитов), гаптофиты (от хризофитов), и хлорарахниофиты (из ксантофитов). [ нужна ссылка ]
С отказом от дихотомической классификации растений и животных большинство групп водорослей (иногда все) были включены в состав Protista , позднее также от него отказались в пользу Eukaryota . Однако, как наследие более старой схемы жизни растений, некоторые группы, которые также рассматривались как простейшие в прошлом , все еще имеют дублирующиеся классификации (см. Амбирегнальные простейшие ). [ нужна ссылка ]
Некоторые паразитические водоросли (например, зеленые водоросли Prototheca и Helicosporidium , паразиты многоклеточных животных, или Cephaleuros , паразиты растений) первоначально были отнесены к грибам , споровикам или протистанам incertae sedis , [42] в то время как у других (например, зеленые водоросли Phyllosiphon и Rhodochytrium , паразиты растений, или красные водоросли Pterocladiophila и Gelidiocolax mammillatus , паразиты других красных водорослей, или динофлагелляты Oodinium , паразиты рыб) их связь с водорослями предполагалась рано. В других случаях некоторые группы первоначально характеризовались как паразитические водоросли (например, Chlorochytrium ), но позже были замечены как эндофитные водоросли. [43] Некоторые нитчатые бактерии (например, Beggiatoa ) первоначально рассматривались как водоросли. Более того, такие группы, как apicomplexans, также являются паразитами, происходящими от предков, имевших пластиды, но не включенными ни в одну группу, традиционно рассматриваемую как водоросли. [ нужна ссылка ]
Эволюция [ править ]
Водоросли полифилетичны , поэтому их происхождение не может быть прослежено до одного гипотетического общего предка . Считается, что они возникли в результате цианобактерий одноклеточным коккоидных фотосинтезирующих гетеротрофным фагоцитирования эукариотом ( протистом ). [44] образуя двухмембранные первичные пластиды . Считается, что такие симбиогенные события (первичный симбиогенез) произошли более 1,5 миллиардов лет назад в Калимский период , в начале «Скучного миллиарда» , но из-за такого большого временного разрыва трудно отследить ключевые события. [45] Первичный симбиогенез дал начало трем подразделениям архепластид Viridiplantae , а именно хищничества ( зеленые водоросли и более поздние растения ), Rhodophyta ( красные водоросли ) и Glaucophyta («серые водоросли»), чьи пластиды в дальнейшем распространились в другие линии протистов посредством эукариот-эукариотного , поглощения. и последующие эндосимбиозы (вторичный и третичный симбиогенез). [45] Этот процесс последовательного «захвата» и «порабощения» клеток объясняет разнообразие фотосинтезирующих эукариот. [44]
Недавние геномные и филогеномные подходы существенно прояснили эволюцию пластидного генома , горизонтальное перемещение генов эндосимбионта и «хозяина» к ядерному геному распространение пластид по эукариотическому древу жизни . [44]
Связь с наземными растениями [ править ]
Окаменелости изолированных спор позволяют предположить, что наземные растения могли существовать примерно 475 миллионов лет назад (млн лет назад) в период позднего кембрия / раннего ордовика . [46] [47] из сидячих мелководных пресноводных харофитовых водорослей, очень похожих на Чару , [48] которые, вероятно, выброшены на берег, когда уровень речной / озёрной воды падал в засушливые сезоны . [49] У этих харофитовых водорослей, вероятно, уже развились нитчатые слоевища и опоры , внешне напоминающие растений стебли и корни , и, вероятно, наблюдалась изоморфная смена поколений . Возможно, они появились около 850 млн лет назад. [50] и может даже достигать 1 Гя во время поздней фазы « Скучного миллиарда» . [51]
Морфология [ править ]

ряд морфологий Представлен водорослей, и часто встречается конвергенция признаков в несвязанных группах. Единственные группы, демонстрирующие трехмерные многоклеточные слоевища, — это красные и коричневые , а также некоторые хлорофиты . [52] Апикальный рост ограничен подмножествами этих групп: красными флоридеофитами , различными коричневыми и харофитами. [52] Форма харофитов сильно отличается от формы красных и коричневых, поскольку у них есть отчетливые узлы, разделенные междоузлиями «стеблями»; мутовки ветвей, напоминающие хвощи . В узлах встречаются [52] Концептуалы - еще одна полифилетическая черта; они появляются у кораллиновых водорослей и Hildenbrandiales , а также у коричневых. [52]
Большинство простейших водорослей представляют собой одноклеточные жгутиконосцы или амебоиды , но в нескольких группах независимо развились колониальные и неподвижные формы. Некоторые из наиболее распространенных организационных уровней, более одного из которых могут встречаться в жизненном цикле вида:
- Колониальный : небольшие, регулярные группы подвижных клеток.
- Капсоид: отдельные неподвижные клетки, заключенные в слизь.
- Коккоид: отдельные неподвижные клетки с клеточными стенками.
- Пальмеллоид: неподвижные клетки, заключенные в слизь.
- Нитчатый: цепочка соединенных неподвижных клеток, иногда ветвящихся.
- Паренхиматозный: клетки, образующие слоевище с частичной дифференцировкой тканей.
В трех линиях достигнуты еще более высокие уровни организации с полной дифференциацией тканей. Это бурые водоросли, [53] — некоторые из которых могут достигать 50 м в длину ( ламинарии ) [54] — красные водоросли, [55] и зеленые водоросли. [56] Наиболее сложные формы встречаются среди харофитовых водорослей (см. Charales и Charophyta ), в линии, которая в конечном итоге привела к высшим наземным растениям. Инновацией, характеризующей эти неводорослевые растения, является наличие женских репродуктивных органов с защитными слоями клеток, которые защищают зиготу и развивающийся эмбрион. Поэтому наземные растения называются эмбриофитами .
Газоны [ править ]
Термин «водорослевой газон» широко используется, но не имеет четкого определения. Водорослевые газоны представляют собой толстые, похожие на ковры заросли морских водорослей, которые удерживают осадок и конкурируют с такими основными видами, как кораллы и водоросли , и обычно имеют высоту менее 15 см. Такой газон может состоять из одного или нескольких видов и обычно занимает площадь порядка квадратного метра или более. Перечислены некоторые общие характеристики: [57]
- Водоросли, образующие скопления, которые были описаны как газоны, включают диатомовые водоросли, цианобактерии, хлорофиты, феофиты и родофиты. Газоны часто состоят из множества видов в широком диапазоне пространственных масштабов, но часто сообщается о моноспецифичных газонах. [57]
- Морфологически газоны могут сильно различаться в географических масштабах и даже внутри видов в локальных масштабах, и их может быть трудно идентифицировать с точки зрения составляющих видов. [57]
- Дерн определяли как короткие водоросли, но это слово использовалось для описания диапазона высоты от менее 0,5 см до более 10 см. В некоторых регионах описания приближались к высоте, которую можно было бы назвать навесом (от 20 до 30 см). [57]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( февраль 2021 г. ) |
Физиология [ править ]
Многие водоросли, особенно виды Characeae , [58] послужили модельными экспериментальными организмами для понимания механизмов водопроницаемости мембран, осморегуляции , регуляции тургора , [ нужны разъяснения ] солеустойчивость , цитоплазматический поток и генерация потенциалов действия .
Фитогормоны содержатся не только в высших растениях, но и в водорослях. [59]
Симбиотические водоросли [ править ]
Некоторые виды водорослей образуют симбиотические отношения с другими организмами. В этих симбиозах водоросли снабжают организм хозяина фотосинтетами (органическими веществами), обеспечивая защиту клеток водорослей. Организм-хозяин частично или полностью получает энергию от водорослей. Примеры:
Лишайники [ править ]

лишайники определяет Международная ассоциация лихенологии как «ассоциацию гриба и фотосинтетического симбионта, приводящую к образованию стабильного вегетативного тела, имеющего специфическую структуру». [60] Грибы, или микобионты, происходят в основном из Ascomycota и в небольшом количестве из Basidiomycota . В природе они не встречаются отдельно от лишайников. Неизвестно, когда они начали сотрудничать. [61] Один микобионт ассоциируется с одним и тем же видом фикобионтов, реже с двумя, из зеленых водорослей, за исключением того, что в качестве альтернативы микобионт может ассоциироваться с одним видом цианобактерий (следовательно, более точным термином является «фотобионт»). Фотобионт может быть связан со многими различными микобионтами или жить независимо; соответственно, лишайники называются и классифицируются как виды грибов. [62] Ассоциация называется морфогенезом, поскольку лишайник обладает формой и возможностями, которыми не обладают только виды-симбионты (их можно выделить экспериментально). Фотобионт, возможно, запускает скрытые в микобионте гены. [63]
Трентеполия является примером распространенного во всем мире рода зеленых водорослей, который может расти сам по себе или подвергаться лихенизации. Таким образом, лишайники частично разделяют среду обитания и часто имеют сходный внешний вид со специализированными видами водорослей ( аэрофитами ), растущими на открытых поверхностях, таких как стволы деревьев и камни, и иногда обесцвечивающими их.
Коралловые рифы [ править ]

Коралловые рифы накоплены из известковых экзоскелетов морских беспозвоночных отряда Scleractinia (каменистые кораллы ). Эти животные метаболизируют сахар и кислород, чтобы получить энергию для процессов построения клеток, включая секрецию экзоскелета, с водой и углекислым газом в качестве побочных продуктов. Динофлагелляты (водорослевые протисты) часто являются эндосимбионтами в клетках морских беспозвоночных, образующих кораллы, где они ускоряют метаболизм клеток-хозяев, генерируя сахар и кислород, сразу же доступные в процессе фотосинтеза с использованием падающего света и углекислого газа, вырабатываемого хозяином. Каменистым кораллам, образующим рифы ( герматипические кораллы необходимы эндосимбиотические водоросли рода Symbiodinium . ), для поддержания здорового состояния [64] Утрата симбиодиниума хозяином известна как обесцвечивание коралла — состояние, которое приводит к ухудшению состояния рифа.
Морские губки [ править ]
Эндосимбионтные зеленые водоросли обитают вблизи поверхности некоторых губок, например, хлебных губок ( Halichondria Panica ). Таким образом, водоросль защищена от хищников; губка снабжается кислородом и сахарами, на которые у некоторых видов может приходиться от 50 до 80% роста губки. [65]
Жизненный цикл [ править ]
Rhodophyta , Chlorophyta и Heterokontophyta , три основных отдела водорослей , имеют жизненные циклы, которые демонстрируют значительные различия и сложность. В общем, существует бесполая фаза, когда клетки морских водорослей диплоидны , половая фаза, когда клетки гаплоидны , за которым следует слияние мужских и женских гамет . Бесполое размножение позволяет эффективно увеличивать популяцию, но возможны меньшие вариации. Обычно при половом размножении одноклеточных и колониальных водорослей две специализированные, совместимые по половому признаку гаплоидные гаметы вступают в физический контакт и сливаются, образуя зиготу . Чтобы обеспечить успешное спаривание, развитие и выпуск гамет строго синхронизированы и регулируются; феромоны могут играть ключевую роль в этих процессах. [66] Половое размножение допускает больше вариаций и обеспечивает эффективное рекомбинационное восстановление повреждений ДНК во время мейоза , ключевого этапа полового цикла. [67] Однако половое размножение обходится дороже, чем бесполое. [68] Было показано, что мейоз происходит у многих различных видов водорослей. [69]
Числа [ править ]

Коллекция водорослей Национального гербария США (находится в Национальном музее естественной истории ) состоит примерно из 320 500 высушенных экземпляров, что хотя и не является исчерпывающим (исчерпывающей коллекции не существует), но дает представление о порядке численности водорослей. видов (это число остается неизвестным). [70] Оценки сильно различаются. Например, согласно одному стандартному учебнику, [71] Британских островах на Согласно отчету Руководящей группы Великобритании по биоразнообразию, насчитывается 20 000 видов водорослей. Другой контрольный список сообщает только о 5000 видах. Что касается разницы примерно в 15 000 видов, в тексте делается вывод: «Потребуется множество подробных полевых исследований, прежде чем можно будет дать надежную оценку общего числа видов…»
Также были сделаны региональные и групповые оценки:
- 5000–5500 видов красных водорослей во всем мире.
- «около 1300 в австралийских морях» [72]
- 400 видов морских водорослей западного побережья Южной Африки, [73] и 212 видов с побережья Квазулу-Натал. [74] Некоторые из них являются дубликатами, поскольку ареал простирается на оба побережья, а общее количество зарегистрированных видов, вероятно, составляет около 500 видов. Большинство из них занесены в Список морских водорослей Южной Африки . Сюда не входят фитопланктон и корковые кораллины.
- 669 морских видов из Калифорнии (США). [75]
- 642 в контрольном списке Британии и Ирландии [76]
и так далее, но, не имея какой-либо научной основы или надежных источников, эти цифры заслуживают не большего доверия, чем упомянутые выше британские. В большинстве оценок также не учитываются микроскопические водоросли, такие как фитопланктон.
По последним оценкам, во всем мире насчитывается 72 500 видов водорослей. [77]
Распространение [ править ]
Распространение видов водорослей достаточно хорошо изучено со времени зарождения фитогеографии в середине XIX века. [78] Водоросли распространяются главным образом путем распространения спор, распространению криптогамных растений спорами аналогично . Споры можно найти в самых разных средах: пресных и морских водах, воздухе, почве, а также в других организмах или на них. [78] Сможет ли спора вырасти во взрослый организм, зависит от вида и условий окружающей среды, в которых приземляется спора.
Споры пресноводных водорослей распространяются преимущественно проточной водой и ветром, а также живыми переносчиками. [78] Однако не все водоемы могут содержать все виды водорослей, поскольку химический состав некоторых водоемов ограничивает количество водорослей, которые могут выжить в них. [78] Морские споры часто распространяются океанскими течениями. Океанская вода представляет собой множество совершенно разных сред обитания в зависимости от температуры и наличия питательных веществ, в результате чего образуются фитогеографические зоны, регионы и провинции. [79]
В некоторой степени распространение водорослей подвержено флористическим нарушениям, вызванным географическими особенностями, такими как Антарктида , большие расстояния океана или общие массивы суши. Таким образом, можно идентифицировать виды, встречающиеся по местоположению, например, « тихоокеанские водоросли» или « водоросли Северного моря ». Когда они происходят вне своих местностей, обычно можно предположить какой-либо транспортный механизм, например корпуса кораблей. Например, Ulva reticulata и U. fasciata путешествовали с материка на Гавайи таким образом .
Картирование возможно только для избранных видов: «есть много достоверных примеров ограниченного распространения». [80] Например, Clathromorphum — арктический род, и он не нанесен на карты далеко к югу от него. [81] Однако ученые считают общие данные недостаточными из-за «трудностей проведения подобных исследований». [82]
Экология [ править ]

Водоросли широко распространены в водоемах, распространены в наземной среде и встречаются в необычных средах, например, на снегу и льду . Морские водоросли растут в основном на мелководье, на глубине менее 100 м (330 футов); однако некоторые из них, такие как Navicula pennata, были зарегистрированы на глубине 360 м (1180 футов). [83] Тип водорослей Ancylonema nordenskioeldii был обнаружен в Гренландии в районах, известных как «Темная зона», что вызвало увеличение скорости таяния ледникового покрова. [84] Такие же водоросли были обнаружены в итальянских Альпах после того, как на некоторых участках ледника Презена появился розовый лед. [85]
Различные виды водорослей играют важную роль в водной экологии. Микроскопические формы, живущие во взвешенном состоянии в толще воды ( фитопланктон ), обеспечивают пищевую основу для большинства морских пищевых цепей . При очень высокой плотности ( цветение водорослей ) эти водоросли могут обесцветить воду и вытеснить, отравить или удушить другие формы жизни.
Водоросли можно использовать в качестве индикаторных организмов для мониторинга загрязнения различных водных систем. [86] Во многих случаях метаболизм водорослей чувствителен к различным загрязнителям. В связи с этим видовой состав популяций водорослей может меняться в присутствии химических загрязнителей. [86] Чтобы обнаружить эти изменения, можно сравнительно легко взять образцы водорослей из окружающей среды и хранить их в лабораториях. [86]
По месту обитания водоросли можно разделить на: водные ( планктонные , донные , морские , пресноводные , чечевичные , лотковые ), [87] наземный , воздушный (субаэральный), [88] литофитный , галофитный (или эвригалинный ), псаммонный , термофильный , криофильный , эпибионтный ( эпифитный , эпизойный ), эндосимбионтный ( эндофитный , эндозойский), паразитический , кальцифильный или лихеновый (фикобионт). [89]
Культурные ассоциации [ править ]
В классическом китайском языке слово 藻 используется как для обозначения «водорослей», так и (в скромной традиции имперских ученых ) для обозначения «литературного таланта». Третий остров на озере Куньмин рядом с Летним дворцом в Пекине известен как Цзаоцзянь Тан Дао (藻鑒堂島), что, таким образом, одновременно означает «Остров зала для наблюдения за водорослями» и «Остров зала для размышлений о литературном таланте». .
Культивирование [ править ]

Альгакультура — это форма аквакультуры, включающая выращивание видов водорослей. [90]
Большинство водорослей, которые намеренно культивируются, относятся к категории микроводорослей (также называемых фитопланктоном , микрофитами или планктонными водорослями ). Макроводоросли , широко известные как морские водоросли , также имеют множество коммерческих и промышленных применений, но из-за их размера и особых требований окружающей среды, в которой они должны расти, они не так легко поддаются культивированию (однако ситуация может измениться). с появлением новых культиваторов морских водорослей, которые по сути представляют собой очистители водорослей, использующие восходящие пузырьки воздуха в небольших контейнерах). [ нужна ссылка ]
Коммерческое и промышленное выращивание водорослей имеет множество применений, включая производство нутрицевтиков, таких как жирные кислоты омега-3 (в виде водорослевого масла). [91] [92] [93] или натуральные пищевые красители и красители , продукты питания , удобрения , биопластики , химическое сырье (сырье), богатые белком корма для животных/ аквакультуры , фармацевтические препараты и топливо из водорослей , [94] а также может использоваться в качестве средства контроля загрязнения и улавливания природного углерода . [95]
Мировое производство выращиваемых водных растений, в котором преобладают морские водоросли, выросло по объему с 13,5 миллионов тонн в 1995 году до чуть более 30 миллионов тонн в 2016 году. [96] Культивируемые микроводоросли уже вносят вклад в широкий спектр секторов развивающейся биоэкономики . [97] Исследования показывают, что существует большой потенциал и преимущества альгакультуры для развития будущей здоровой и устойчивой продовольственной системы . [98] [95]Выращивание морских водорослей [ править ]


Выращивание морских водорослей или выращивание ламинарии — это практика выращивания и сбора морских водорослей . урожая В простейшем случае фермеры собирают урожай с естественных грядок, тогда как в другом крайнем случае фермеры полностью контролируют жизненный цикл .
Семью наиболее культивируемыми таксонами являются виды Eucheuma , Kappaphycus alvarezii , виды Gracilaria , Saccharina japonica , Undaria pinnatifida , виды Pyropia и Sargassum fusiforme . Eucheuma и K. alvarezii привлекательны каррагинаном ( желирующим агентом ); Gracilaria выращивается на агаре ; остальные съедаются после ограниченной обработки. [99] Морские водоросли отличаются от мангровых зарослей и морских трав , поскольку они являются фотосинтезирующими водорослевыми организмами. [100] и не цветут. [99]
Крупнейшими странами-производителями морских водорослей по состоянию на 2022 год являются Китай (58,62%) и Индонезия (28,6%); за ней следуют Южная Корея (5,09%) и Филиппины (4,19%). Другие известные производители включают Северную Корею (1,6%), Японию (1,15%), Малайзию (0,53%), Занзибар ( Танзания , 0,5%) и Чили (0,3%). [101] [102] Выращивание морских водорослей часто развивалось для улучшения экономических условий и снижения нагрузки на рыболовство. [103]
( ФАО Продовольственная и сельскохозяйственная организация ) сообщила, что мировое производство в 2019 году составило более 35 миллионов тонн. Северная Америка произвела около 23 000 тонн влажных морских водорослей. Аляска, Мэн, Франция и Норвегия более чем удвоили производство морских водорослей с 2018 года . По состоянию на 2019 год морские водоросли составляли 30% морской аквакультуры . [104]
Выращивание морских водорослей — это культура с отрицательным выбросом углерода и высоким потенциалом смягчения последствий изменения климата . [105] [106] Специальный доклад МГЭИК об океане и криосфере в условиях меняющегося климата рекомендует «дальнейшее исследовательское внимание» в качестве тактики смягчения последствий. [107] Всемирный фонд дикой природы , организации «Океан 2050» и The Nature Conservancy публично поддерживают расширение выращивания морских водорослей. [104]Биореакторы [ править ]

Биореактор водорослей используется для выращивания микро- или макроводорослей . Водоросли можно культивировать с целью производства биомассы (как в культиваторе морских водорослей ), очистки сточных вод , CO 2 фиксации или фильтрации аквариума/пруда с помощью очистителя водорослей . [108] Биореакторы из водорослей сильно различаются по конструкции и делятся на две категории: открытые реакторы и закрытые реакторы. Открытые реакторы подвергаются воздействию атмосферы, тогда как закрытые реакторы, также обычно называемые фотобиореакторами , в различной степени изолированы от атмосферы. В частности, биореакторы из водорослей можно использовать для производства топлива, такого как биодизель и биоэтанол , для производства корма для животных или для снижения содержания загрязняющих веществ, таких как NO x и CO 2 в топливе.
газы электростанций. По сути, этот тип биореактора основан на реакции фотосинтеза , которую осуществляют сами хлорофиллсодержащие водоросли с использованием растворенного углекислого газа и солнечного света. Углекислый газ диспергируется в реакторной жидкости, чтобы сделать его доступным для водорослей. Биореактор должен быть изготовлен из прозрачного материала.Использует [ править ]

Агар [ править ]
Агар , желеобразное вещество, полученное из красных водорослей, имеет ряд коммерческих применений. [109] Это хорошая среда для выращивания бактерий и грибов, поскольку большинство микроорганизмов не переваривают агар.
Альгинаты [ править ]
Альгиновая кислота , или альгинат, добывается из бурых водорослей . Его применение варьируется от желирующих веществ в пищевых продуктах до медицинских повязок. Альгиновая кислота также использовалась в области биотехнологии в качестве биосовместимой среды для инкапсуляции и иммобилизации клеток. Молекулярная кухня также использует это вещество из-за его желирующих свойств, благодаря чему оно становится средством доставки ароматизаторов.
от 100 000 до 170 000 влажных тонн Macrocystis собирают Ежегодно в Нью-Мексико для экстракции альгината и корма для морских ушек. [110] [111]
Источник энергии [ править ]
Чтобы быть конкурентоспособным и независимым от нестабильной поддержки со стороны (местной) политики в долгосрочной перспективе, биотопливо должно равняться или превосходить уровень затрат на ископаемое топливо. Здесь большие перспективы имеют виды топлива на основе водорослей. [112] [113] напрямую связано с возможностью производить больше биомассы на единицу площади в год, чем любая другая форма биомассы. По оценкам, точка безубыточности для биотоплива на основе водорослей наступит к 2025 году. [114]
Удобрения [ править ]

На протяжении веков морские водоросли использовались в качестве удобрения; Джордж Оуэн из Хенлли писал в 16 веке о заносных сорняках в Южном Уэльсе : [115]
Такую руду они часто собирают и складывают в большие кучи, где она разлагается и гниет и имеет сильный и отвратительный запах; когда они настолько гнилые, они выбрасывают землю, как и навоз, и из нее вырастает хорошая кукуруза, особенно ячмень ... После весенних приливов или больших морских буровых установок они привозят ее в мешках на спинах лошадей и везут те же три, четыре или пять миль и бросьте его на поле, которое гораздо лучше подходит для выращивания кукурузы и травы.
Сегодня водоросли используются людьми по-разному; например, в качестве удобрений , кондиционеров для почвы и корма для скота. [116] Водные и микроскопические виды культивируются в прозрачных резервуарах или прудах и либо собираются, либо используются для очистки сточных вод, перекачиваемых через пруды. альгакультура в больших масштабах является важным видом аквакультуры В некоторых местах . Maerl обычно используется в качестве кондиционера почвы.
Питание [ править ]

Естественно растущие морские водоросли являются важным источником пищи, особенно в Азии, поэтому некоторые называют их суперпродуктами . [117] Они содержат множество витаминов, в том числе: A, , B1 B2 , кальцием B6 , и ниацин а C , также богаты йодом , калием , железом магнием и , . [118] Кроме того, коммерчески культивируемые микроводоросли, включая водоросли и цианобактерии, продаются как пищевые добавки, такие как спирулина , [119] Хлорелла и добавка витамина С от Dunaliella с высоким содержанием бета-каротина .
Водоросли являются национальным продуктом питания многих народов: Китай потребляет более 70 видов, в том числе жирную чой , цианобактерию, считающуюся овощем; Япония — более 20 видов, таких как нори и аонори ; [120] Ирландия, милая ; Чили , кочаюйо . [121] Лавер используется для приготовления лаврового хлеба в Уэльсе , где он известен как бара лоур . В Корее . лавра делают соус из зеленого Он также используется вдоль западного побережья Северной Америки от Калифорнии до Британской Колумбии , на Гавайях и маори Новой Зеландии . Морской салат и бэддерлок являются ингредиентами салатов в Шотландии , Ирландии, Гренландии и Исландии . Водоросли считаются потенциальным решением мировой проблемы голода. [122] [123] [124]
В кухне используются две популярные формы водорослей:
- Хлорелла : Эта форма водорослей встречается в пресной воде и содержит фотосинтетические пигменты в хлоропластах . Он богат железом , цинком , магнием , витамином В2 и жирными кислотами Омега-3 .
Кроме того, он содержит все девять незаменимых аминокислот, которые организм не производит самостоятельно. [125]
Масла некоторых водорослей имеют высокий уровень ненасыщенных жирных кислот . Например, Parietochromis incisa отличается высоким содержанием арахидоновой кислоты , где она достигает 47% пула триглицеридов. [126] Некоторые виды водорослей, предпочитаемые вегетарианством и веганством, содержат длинноцепочечные незаменимые жирные кислоты омега-3 , докозагексаеновую кислоту (DHA) и эйкозапентаеновую кислоту (EPA). Рыбий жир содержит жирные кислоты омега-3, но первоначальным источником являются водоросли (в частности, микроводоросли), которые поедаются морскими обитателями, такими как копеподы , и передаются по пищевой цепи. [127] В последние годы водоросли стали популярным источником жирных кислот омега-3 для вегетарианцев, которые не могут получить длинноцепочечные ЭПК и ДГК из других вегетарианских источников, таких как льняное масло , которое содержит только короткоцепочечную альфа-линоленовую кислоту (АЛК). .
Контроль загрязнения [ править ]
- Сточные воды можно очищать водорослями. [128] сокращение использования большого количества токсичных химикатов, которые в противном случае были бы необходимы.
- Водоросли можно использовать для улавливания удобрений из стоков с ферм. При последующем сборе обогащенные водоросли можно использовать в качестве удобрения.
- Аквариумы и пруды можно фильтровать с помощью водорослей, которые поглощают питательные вещества из воды с помощью устройства, называемого очистителем водорослей , также известным как очиститель газона из водорослей. [129] [130]
Ученые Службы сельскохозяйственных исследований обнаружили, что 60–90% стоков азота и 70–100% стоков фосфора можно улавливать из навозных стоков с помощью горизонтального скруббера из водорослей, также называемого скруббером из водорослей (ATS). Ученые разработали ATS, который состоит из неглубоких 100-футовых желобов из нейлоновой сетки, где могут образовываться колонии водорослей, и изучали его эффективность в течение трех лет. Они обнаружили, что водоросли можно легко использовать для уменьшения стока питательных веществ с сельскохозяйственных полей и повышения качества воды, поступающей в реки, ручьи и океаны. Исследователи собрали и высушили богатые питательными веществами водоросли из САР и изучили их потенциал в качестве органического удобрения. Они обнаружили, что рассада огурцов и кукурузы растет при использовании органических удобрений САР так же хорошо, как и при использовании коммерческих удобрений. [131] Скрубберы из водорослей, использующие версии с восходящим потоком пузырьков или версиями с вертикальным водопадом, теперь также используются для фильтрации аквариумов и прудов.
Полимеры [ править ]
Из водорослей можно создавать различные полимеры, которые могут быть особенно полезны при создании биопластиков. К ним относятся гибридные пластики, пластики на основе целлюлозы, полимолочная кислота и биополиэтилен. [132] Несколько компаний начали коммерческое производство полимеров из водорослей, в том числе для использования в шлепанцах. [133] и в досках для серфинга. [134]
Биоремедиация [ править ]
водоросль Stichococcus bacillaris Было замечено, что колонизирует силиконовые смолы, используемые на археологических раскопках; биоразложение синтетического вещества. [135]
Пигменты [ править ]
Натуральные пигменты ( каротиноиды и хлорофиллы ), вырабатываемые водорослями, могут использоваться в качестве альтернативы химическим красителям и красителям. [136] Присутствие некоторых отдельных пигментов водорослей вместе с определенными соотношениями концентраций пигментов специфично для таксона: анализ их концентраций с помощью различных аналитических методов, в частности, высокоэффективной жидкостной хроматографии , может, таким образом, дать глубокое понимание таксономического состава и относительной численности природных водорослей. Популяции водорослей в пробах морской воды. [137] [138]
Стабилизирующие вещества [ править ]
Каррагинан, получаемый из красной водоросли Chondrus Crispus , используется в качестве стабилизатора в молочных продуктах.
Дополнительные изображения [ править ]
 Водорослевой пузырь
Водорослевой пузырь

См. также [ править ]
- AlgaeBase
- ВодорослиПАРК
- Эвтрофикация
- Железные удобрения
- Маримо водоросли
- Микробиотопливо
- Микрофит
- Фотобиореактор
- Фикотехнология
- Растение
- Анатоксин – анатоксин
Ссылки [ править ]
- ^ Баттерфилд, Нью-Джерси (2000). « Bangiomorpha pubescens n. gen., n. sp.: Значение для эволюции пола, многоклеточности и мезопротерозойской/неопротерозойской радиации эукариот» . Палеобиология . 26 (3): 386–404. Бибкод : 2000Pbio...26..386B . doi : 10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2 . ISSN 0094-8373 . S2CID 36648568 . Архивировано из оригинала 7 марта 2007 года.
- ^ ТМ Гибсон (2018). «Точный возраст Bangiomorpha pubescens датирует возникновение эукариотического фотосинтеза» . Геология . 46 (2): 135–138. Бибкод : 2018Geo....46..135G . дои : 10.1130/G39829.1 .
- ^ «ВОДОРОСЛИ | Английское значение — Кембриджский словарь» . Проверено 6 апреля 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Нэборс, Мюррей В. (2004). Введение в ботанику . Pearson Education, Inc. Сан-Франциско: ISBN 978-0-8053-4416-5 .
- ↑ Перейти обратно: Перейти обратно: а б с Килинг, Патрик Дж. (2004). «Разнообразие и история эволюции пластид и их хозяев» . Американский журнал ботаники . 91 (10): 1481–1493. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 .
- ^ Палмер, доктор медицинских наук; Солтис, Делавэр; Чейз, М.В. (2004). «Растительное древо жизни: обзор и некоторые точки зрения» . Американский журнал ботаники . 91 (10): 1437–1445. дои : 10.3732/ajb.91.10.1437 . ПМИД 21652302 .
- ^ Смитсоновский национальный музей естественной истории; Кафедра ботаники. «Исследование водорослей» . Архивировано из оригинала 2 июля 2010 года . Проверено 25 августа 2010 г.
- ^ Прингсхайм, Э.Г. 1963. Бесцветные водоросли. Вклад в эволюционные исследования . Густав Фишер Верлаг, Штутгарт. 471 стр., виды: Algae#Pringsheim (1963) .
- ^ Тартар, А.; Бусиас, генеральный директор; Бекнель, Джей Джей; Адамс, Би Джей (2003). «Сравнение генов пластидной 16S рРНК (rrn 16) из видов Helicosporidium: данные, подтверждающие реклассификацию Helicosporidia в зеленые водоросли (Chlorophyta)» . Международный журнал систематической и эволюционной микробиологии . 53 (Часть 6): 1719–1723. дои : 10.1099/ijs.0.02559-0 . ПМИД 14657099 .
- ^ Фигероа-Мартинес, Ф.; Недельку, AM; Смит, доктор медицинских наук; Рейес-Прието, А. (2015). «Когда гаснет свет: эволюционная судьба свободноживущих бесцветных зеленых водорослей» . Новый фитолог . 206 (3): 972–982. дои : 10.1111/nph.13279 . ПМК 5024002 . ПМИД 26042246 .
- ^ Бенгтсон, С.; Беливанова В.; Расмуссен, Б.; Уайтхаус, М. (2009). «Спорные «кембрийские» окаменелости виндхийцев реальны, но старше их более чем на миллиард лет» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (19): 7729–7734. Бибкод : 2009PNAS..106.7729B . дои : 10.1073/pnas.0812460106 . ПМЦ 2683128 . ПМИД 19416859 .
- ^ Пол, Вишал; Чандра Шехарая, PS; Кушваха, Шивбачан; Сапре, Аджит; Дасгупта, Сантану; Саньял, Дебанджан (2020). «Роль водорослей в связывании CO2 в решении проблемы изменения климата: обзор» . В Деб, Дипанкар; Диксит, Амбеш; Чандра, Лалту (ред.). Возобновляемая энергия и изменение климата . Умные инновации, системы и технологии. Том. 161. Сингапур: Спрингер. стр. 257–265. дои : 10.1007/978-981-32-9578-0_23 . ISBN 978-981-329-578-0 . S2CID 202902934 .
- ^ «водоросль, водоросли». Третий новый международный словарь английского языка Вебстера, полный со словарем семи языков . Том. 1. Британская энциклопедия, Inc., 1986.
- ^ Партридж, Эрик (1983). «водоросли». Происхождение . Гринвичский дом. ISBN 9780517414255 .
- ^ Льюис, Чарльтон Т.; Короткий, Чарльз (1879). «Алга» . Латинский словарь . Оксфорд: Кларендон Пресс . Проверено 31 декабря 2017 г.
- ^ Чейн, Томас Келли; Блэк, Джон Сазерленд (1902). Библейская энциклопедия: критический словарь литературной, политической и религиозной истории, археологии, географии и естественной истории Библии . Компания Макмиллан. п. 3525.
- ^ Ли, Роберт Эдвард, изд. (2008), «Основные характеристики водорослей» , Phycology (4-е изд.), Кембридж: Cambridge University Press, стр. 3–30, doi : 10.1017/CBO9780511812897.002 , ISBN 978-1-107-79688-1 , получено 13 сентября 2023 г.
- ^ Ли, RE (2008). Психология . Издательство Кембриджского университета. ISBN 9780521367448 .
- ^ Аллаби, М., изд. (1992). «Водоросли». Краткий словарь ботаники . Издательство Оксфордского университета.
- ^ Сато, Наоки (27 мая 2021 г.). «Являются ли цианобактерии предком хлоропластов или просто одним из доноров генов растений и водорослей?» . Гены . 12 (6): 823. doi : 10.3390/genes12060823 . ISSN 2073-4425 . ПМК 8227023 . ПМИД 34071987 .
- ^ Бхаттачарья, Д.; Медлин, Л. (1998). «Филогения водорослей и происхождение наземных растений» (PDF) . Физиология растений . 116 (1): 9–15. дои : 10.1104/стр.116.1.9 . ПМК 1539170 . Архивировано (PDF) из оригинала 7 февраля 2009 г.
- ^ Лосос, Джонатан Б.; Мейсон, Кеннет А.; Певица, Сьюзен Р. (2007). Биология (8-е изд.). МакГроу-Хилл. ISBN 978-0-07-304110-0 .
- ^ Арчибальд, Дж. М.; Килинг, П.Дж. (ноябрь 2002 г.). «Переработанные пластиды:« зеленое движение »в эволюции эукариот». Тенденции в генетике . 18 (11): 577–584. дои : 10.1016/S0168-9525(02)02777-4 . ПМИД 12414188 .
- ^ О'Нил, Эллис С.; Трик, Мартин; Анрисса, Бернар; Филд, Роберт А. (2015). «Эвглена во времени: эволюция, контроль центральных метаболических процессов и многодоменные белки в биохимии углеводов и натуральных продуктов» . Перспективы в науке . 6 : 84–93. Бибкод : 2015PerSc...6...84O . дои : 10.1016/j.pisc.2015.07.002 .
- ^ Понсе-Толедо, Рафаэль И.; Лопес-Гарсия, Purificación; Морейра, Дэвид (октябрь 2019 г.). «Горизонтальный и эндосимбиотический перенос генов в ранней эволюции пластид» . Новый фитолог . 224 (2): 618–624. дои : 10.1111/nph.15965 . ISSN 0028-646X . ПМК 6759420 . ПМИД 31135958 .
- ^ Понсе-Толедо, Рафаэль I; Морейра, Дэвид; Лопес-Гарсия, Purificación; Дешам, Филипп (19 июня 2018 г.). «Вторичные пластиды эвгленид и хлорарахниофитов функционируют благодаря смеси генов происхождения красных и зеленых водорослей» . Молекулярная биология и эволюция . 35 (9): 2198–2204. дои : 10.1093/molbev/msy121 . ISSN 0737-4038 . ПМЦ 6949139 . ПМИД 29924337 .
- ^ Янсон, Свен; Гранели, Эдна (сентябрь 2003 г.). «Генетический анализ гена psbA из отдельных клеток указывает на криптомонадное происхождение пластиды у Dinophys (Dinophyceae)». Психология . 42 (5): 473–477. Бибкод : 2003Phyco..42..473J . дои : 10.2216/i0031-8884-42-5-473.1 . ISSN 0031-8884 . S2CID 86730888 .
- ^ Вегенер Парфри, Лаура ; Барберо, Эрика; Лассер, Элиз; Данторн, Мика; Бхаттачарья, Дебашиш; Паттерсон, Дэвид Дж .; Кац, Лаура А. (декабрь 2006 г.). «Оценка поддержки текущей классификации эукариотического разнообразия» . ПЛОС Генетика . 2 (12): е220. дои : 10.1371/journal.pgen.0020220 . ПМЦ 1713255 . ПМИД 17194223 .
- ^ Бурки, Ф.; Шалчиан-Тебризи, К.; Минге, М.; Скъёвеланд, О.; Николаев, С.И.; и др. (2007). Батлер, Джеральдин (ред.). «Филогеномика меняет супергруппы эукариот» . ПЛОС ОДИН . 2 (8): е790. Бибкод : 2007PLoSO...2..790B . дои : 10.1371/journal.pone.0000790 . ЧВК 1949142 . ПМИД 17726520 .
- ^ Линней, Чарльз (1753). Виды растений Том. 2. За счет Лаврентия Сальвия. п. 1131
- ^ Шарма, ОП (1 января 1986 г.). Учебник Водорослей . Тата МакГроу-Хилл. п. 22. ISBN 9780074519288 .
- ^ Гмелин, С.Г. (1768). История драконов Св. Петербург: Из типографии Академии наук - через Google Книги.
- ^ Сильва, ПК; Бассон, П.В.; Мо, Р.Л. (1996). Каталог бентосных морских водорослей Индийского океана . Издательство Калифорнийского университета. ISBN 9780520915817 – через Google Книги.
- ↑ Перейти обратно: Перейти обратно: а б Медлин, Линда К.; Кооистра, Вибе ХКФ; Поттер, Дэниел; Сондерс, Гэри В.; Андерсон, Роберт А. (1997). «Филогенетические взаимоотношения« золотых водорослей »(гаптофитов, гетероконтных хромофитов) и их пластид» (PDF) . Систематика и эволюция растений : 188. Архивировано (PDF) из оригинала 5 октября 2013 г.
- ^ Диксон, PS (1973). Биология родофитов . Эдинбург: Оливер и Бойд. п. 232. ИСБН 978-0-05-002485-0 .
- ^ Харви, Д. (1836). «Водоросли» (PDF) . В Маккее, Дж. Т. (ред.). Flora hibernica, включающая цветковые растения, папоротники, Characeae Musci Hepaticae, лишайники и водоросли Ирландии, расположенные в соответствии с естественной системой, с кратким описанием родов в соответствии с системой Линнея . стр. 157–254. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 31 декабря 2017 г. .
- ↑ Браун, А. Algarum unicellium genera nova et minus cognita, praemissis Observeibus de Algis unicellibus in Genere (Новые и менее известные роды одноклеточных водорослей, которым предшествуют наблюдения, касающиеся одноклеточных водорослей в целом). Архивировано 20 апреля 2016 года в Wayback Machine . Lipsiae, Apud W. Engelmann, 1855. Перевод: Ланкестер, Э. и Буск, Г. (ред.). Ежеквартальный журнал микроскопической науки , 1857 г., том. 5, (17), 13–16. Архивировано 4 марта 2016 г. в Wayback Machine ; (18), 90–96. Архивировано 5 марта 2016 г. в Wayback Machine ; (19), 143–149. Архивировано 4 марта 2016 г. в Wayback Machine .
- ^ Зибольд, C. Th v. « Одноклеточные растения и животные Ueber (Об одноклеточных растениях и животных). Архивировано 26 ноября 2014 г. в Wayback Machine ». В: Зибольд, К.Т. против. и Кёлликер, А. (1849). Журнал научной зоологии , Том 1, с. 270. Перевод: Ланкестер Э. и Буск Г. (ред.). Ежеквартальный журнал микроскопической науки , 1853 г., том. 1, (2), 111–121. Архивировано 4 марта 2016 г. в Wayback Machine ; (3), 195–206. Архивировано 4 марта 2016 г. в Wayback Machine .
- ↑ Перейти обратно: Перейти обратно: а б Рэган, Марк (3 июня 2010 г.). «О разграничении и высшей классификации водорослей» . Европейский журнал психологии . 33 (1): 1–15. дои : 10.1080/09670269810001736483 . Проверено 16 февраля 2024 г.
- ^ де Жюссье, Антуан Лоран (1789). Роды растений, расположенные в естественном порядке . В Париже у вдовы Эриссан и Теофила Барруа. п. 6.
- ^ Хан, Амна Комал; Каусар, Хумера; Джафери, Сияда Самра; и др. (6 ноября 2020 г.). « Взгляд на эволюцию и геномику водорослей» . Биомолекулы . 10 (11): 1524. doi : /biom10111524 . PMC 7694994. . PMID 33172219 10.3390
- ^ Уильямс, бакалавр; Килинг, Пи Джей (2003). «Загадочные органеллы паразитических протистов и грибов» . В Литтлвуде, DTJ (ред.). Эволюция паразитизма . Лондон: Elsevier Academic Press. п. 46. ИСБН 978-0-12-031754-7 .
- ^ Раунд (1981). стр. 398–400, Раунд, FE (8 марта 1984 г.). Экология водорослей . Архив Кубка. ISBN 9780521269063 . Проверено 6 февраля 2015 г. .
- ↑ Перейти обратно: Перейти обратно: а б с Рейес-Прието, Адриан; Вебер, Андреас П.М.; Бхаттачарья, Дебашиш (2007). «Происхождение и формирование пластид в водорослях и растениях» . Ежегодный обзор генетики . 41 : 147–168. дои : 10.1146/annurev.genet.41.110306.130134 . ПМИД 17600460 . Проверено 3 декабря 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Хан, Амна Комал; Каусар, Хумера; Джафери, Сайеда Самра; Друэ, Саманта; Хано, Кристоф; Аббаси, Билал Хайдер; Анджум, Сумайра (6 ноября 2020 г.). «Взгляд на эволюцию и геномику водорослей» . Биомолекулы . 10 (11): 1524. doi : 10.3390/biom10111524 . ПМЦ 7694994 . ПМИД 33172219 .
- ^ Благородный, Иван (18 сентября 2003 г.). «Когда растения завоевали землю» . Би-би-си. Архивировано из оригинала 11 ноября 2006 года.
- ^ Веллман, Швейцария; Остерлофф, Польша; Мохиуддин, У. (2003). «Фрагменты древнейших наземных растений» . Природа . 425 (6955): 282–285. Бибкод : 2003Natur.425..282W . дои : 10.1038/nature01884 . ПМИД 13679913 . S2CID 4383813 . Архивировано из оригинала 30 августа 2017 года.
- ^ Кенрик, П.; Крейн, PR (1997). Происхождение и ранняя диверсификация наземных растений. Кладистическое исследование . Вашингтон: Издательство Смитсоновского института. ISBN 978-1-56098-729-1 .
- ^ Рэйвен, Дж.А.; Эдвардс, Д. (2001). «Корни: эволюционное происхождение и биогеохимическое значение» . Журнал экспериментальной ботаники . 52 (90001): 381–401. doi : 10.1093/jexbot/52.suppl_1.381 . ПМИД 11326045 .
- ^ Кнаут, Л. Пол; Кеннеди, Мартин Дж. (2009). «Позднедокембрийское позеленение Земли». Природа . 460 (7256): 728–732. Бибкод : 2009Natur.460..728K . дои : 10.1038/nature08213 . ПМИД 19587681 . S2CID 4398942 .
- ^ Стротер, Пол К.; Баттисон, Лейла; Брейзер, Мартин Д.; Веллман, Чарльз Х. (2011). «Самые ранние неморские эукариоты Земли». Природа . 473 (7348): 505–509. Бибкод : 2011Natur.473..505S . дои : 10.1038/nature09943 . ПМИД 21490597 . S2CID 4418860 .
- ↑ Перейти обратно: Перейти обратно: а б с д Сяо, С.; Нолл, АХ; Юань, X.; Пюшель, CM (2004). «Фосфатизированные многоклеточные водоросли в неопротерозойской формации Душаньтуо, Китай, и ранняя эволюция флоридеофитных красных водорослей» . Американский журнал ботаники . 91 (2): 214–227. дои : 10.3732/ajb.91.2.214 . ПМИД 21653378 .
- ^ Ваггонер, Бен (1994–2008). «Знакомство с Phaeophyta: ламинарии и бурые «водоросли» » . Музей палеонтологии Калифорнийского университета (UCMP). Архивировано из оригинала 21 декабря 2008 года . Проверено 19 декабря 2008 г.
- ^ Томас, Д.Н. (2002). Морские водоросли . Лондон: Музей естественной истории. ISBN 978-0-565-09175-0 .
- ^ Ваггонер, Бен (1994–2008). «Знакомство с Rhodophyta, красными водорослями » . Музей палеонтологии Калифорнийского университета (UCMP). Архивировано из оригинала 18 декабря 2008 года . Проверено 19 декабря 2008 г.
- ^ «Знакомство с зелеными водорослями» . Беркли.edu . Архивировано из оригинала 13 февраля 2007 года . Проверено 15 февраля 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Коннелл, Шон; Фостер, MS; Айролди, Лаура (9 января 2014 г.). «Что такое водорослевые газоны? К лучшему описанию газонов» . Серия «Прогресс в области морской экологии» . 495 : 299–307. Бибкод : 2014MEPS..495..299C . дои : 10.3354/meps10513 .
- ^ Тазава, Масаси (2010). «Шестидесятилетние исследования клеток харовых: потрясающий материал для биологии растительных клеток» . Прогресс в ботанике 72 . Том. 72. Спрингер. стр. 5–34. дои : 10.1007/978-3-642-13145-5_1 . ISBN 978-3-642-13145-5 . Проверено 7 октября 2012 г.
- ^ Тараховская, Е.Р.; Маслов, Ю. Я.; Шишова, М.Ф. (апрель 2007 г.). «Фитогормоны в водорослях». Российский журнал физиологии растений . 54 (2): 163–170. дои : 10.1134/s1021443707020021 . S2CID 27373543 .
- ^ Бродо, Ирвин М.; Шарнофф, Сильвия Дюран; Шарнофф, Стивен; Лори-Бурк, Сьюзен (2001). Лишайники Северной Америки . Нью-Хейвен: Издательство Йельского университета. п. 8. ISBN 978-0-300-08249-4 .
- ^ Пирсон, Лоренц К. (1995). Разнообразие и эволюция растений . ЦРК Пресс. п. 221. ИСБН 978-0-8493-2483-3 .
- ^ Бродо и др. (2001), с. 6: «Вид лишайника, собранный в любом месте своего ареала, имеет один и тот же лишайниковообразующий гриб и, как правило, один и тот же фотобионт. (Однако конкретный фотобионт может ассоциироваться с множеством различных лишайниковых грибов)».
- ^ Бродо и др. (2001), с. 8.
- ^ Тейлор, Деннис Л. (1983). «Кораллово-водорослевой симбиоз». В Гоффе, Линда Дж. (ред.). Водорослевой симбиоз: континуум стратегий взаимодействия . Архив Кубка. стр. 19–20 . ISBN 978-0-521-25541-7 .
- ^ Найт, Сьюзен (осень 2001 г.). «Есть ли в вашем озере губки?» (PDF) . Озеро Тайдс . 26 (4). Партнерство Висконсин-Лейкс: 4–5. Архивировано из оригинала (PDF) 2 июля 2007 года . Проверено 4 августа 2007 г. - через UWSP.edu.
- ^ Френкель Дж.; Виверман, В.; Понерт, Г. (2014). «Передача сигналов феромонов при половом размножении водорослей» . Плант Дж . 79 (4): 632–644. дои : 10.1111/tpj.12496 . ПМИД 24597605 .
- ^ Бернштейн, Харрис; Байерли, Генри К.; Хопф, Фредерик А.; Мишо, Ричард Э. (20 сентября 1985 г.). «Генетические повреждения, мутации и эволюция пола» . Наука . 229 (4719): 1277–1281. Бибкод : 1985Sci...229.1277B . дои : 10.1126/science.3898363 . ISSN 0036-8075 . ПМИД 3898363 .
- ^ Отто, СП (2009). «Эволюционная загадка секса» . Являюсь. Нат . 174 (Приложение 1): С1–С14. дои : 10.1086/599084 . ПМИД 19441962 . S2CID 9250680 . Архивировано из оригинала 9 апреля 2017 года.
- ^ Хейвуд, П.; Маги, ПТ (1976). «Мейоз у простейших: некоторые структурные и физиологические аспекты мейоза у водорослей, грибов и простейших» . Бактериол Рев . 40 (1): 190–240. дои : 10.1128/MMBR.40.1.190-240.1976 . ПМК 413949 . ПМИД 773364 .
- ^ «Гербарий водорослей» . Национальный музей естественной истории, отдел ботаники. 2008. Архивировано из оригинала 1 декабря 2008 года . Проверено 19 декабря 2008 г.
- ^ Джон (2002), с. 1.
- ^ Хьюсман (2000), с. 25.
- ^ Стегенга (1997).
- ^ Клерк, Оливье (2005). Путеводитель по водорослям Квазулу-Натала . Национальный ботанический сад Бельгии. ISBN 978-90-72619-64-8 .
- ^ Эбботт и Холленберг (1976), с. 2.
- ^ Харди и Гири (2006).
- ^ Гири, Майкл Д. (2012). «Сколько существует видов водорослей?» . Журнал психологии . 48 (5): 1057–1063. Бибкод : 2012JPcgy..48.1057G . дои : 10.1111/j.1529-8817.2012.01222.x . ПМИД 27011267 . S2CID 30911529 .
- ↑ Перейти обратно: Перейти обратно: а б с д Раунд, FE (1981). «Глава 8. Распространение, непрерывность и фитогеография» . Экология водорослей . Архив Кубка. стр. 357–361. ISBN 9780521269063 – через Google Книги.
- ^ Раунд (1981), с. 362.
- ^ Раунд (1981), с. 357.
- ^ Раунд (1981), с. 371.
- ^ Раунд (1981), с. 366.
- ^ Раунд (1981), с. 176.
- ^ «В Гренландии есть загадочная «темная зона», и она становится еще темнее» . Space.com . 10 апреля 2018 г.
- ^ «Альпийский ледник становится розовым из-за водорослей, которые ускоряют изменение климата, - говорят ученые» . Небесные новости . 6 июля 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б с Омар, Ван Мазна Ван (декабрь 2010 г.). «Перспективы использования водорослей в качестве биологических индикаторов для мониторинга и защиты водной среды с особым упором на пресноводные экосистемы Малайзии» . Trop Life Sci Res . 21 (2): 51–67. ПМК 3819078 . ПМИД 24575199 .
- ^ Некки-младший, О. (ред.) (2016). Речные водоросли . Спрингер, Некки, Орландо-младший (2 июня 2016 г.). Речные водоросли . Спрингер. ISBN 9783319319841 . .
- ^ Йохансен, младший (2012). «Диатомовые водоросли: применение в науках об окружающей среде и Земле» . В Смоле, Япония; Стермер, Э.Ф. (ред.). Диатомеи воздушных местообитаний (2-е изд.). Издательство Кембриджского университета. стр. 465–472. ISBN 9781139492621 – через Google Книги.
- ^ Шарма, ОП (1986). стр. 2–6, [1] .
- ^ Хуземанн, М.; Уильямс, П.; Эдмундсон, Скотт Дж.; Чен, П.; Крук, Р.; Куллинан, В.; Кроу, Б.; Лундквист, Т. (сентябрь 2017 г.). «Фотобиореактор лабораторного симулятора пруда с водорослями (LEAPS): валидация с использованием открытых прудовых культур Chlorella sorokiniana и Nannochrominiana salina» . Водорослевые исследования . 26 : 39–46. Бибкод : 2017AlgRe..26...39H . дои : 10.1016/j.algal.2017.06.017 . ISSN 2211-9264 . ОСТИ 1581797 .
- ^ Лейн, Кэти; Дербишир, Эмма; Ли, Вейли; Бреннан, Чарльз (январь 2014 г.). «Биодоступность и потенциальное использование вегетарианских источников жирных кислот омега-3: обзор литературы». Критические обзоры в области пищевой науки и питания . 54 (5): 572–579. дои : 10.1080/10408398.2011.596292 . ПМИД 24261532 . S2CID 30307483 .
- ^ Уинвуд, Р.Дж. (2013). «Водорослевое масло как источник жирных кислот омега-3». Обогащение пищевых продуктов жирными кислотами Омега-3 . Серия публикаций Woodhead по пищевой науке, технологиям и питанию. стр. 389–404. дои : 10.1533/9780857098863.4.389 . ISBN 978-0-85709-428-5 .
- ^ Ленихан-Гилс, Джорджия; Епископ, Карен; Фергюсон, Линнетт (18 апреля 2013 г.). «Альтернативные источники жиров омега-3: можем ли мы найти устойчивую замену рыбе?» . Питательные вещества . 5 (4): 1301–1315. дои : 10.3390/nu5041301 . ПМЦ 3705349 . ПМИД 23598439 .
- ^ Венкатеш, Г. (1 марта 2022 г.). «Циркулярная биоэкономика — парадигма будущего: систематический обзор публикаций научных журналов с 2015 по 2021 год» . Круговая экономика и устойчивое развитие . 2 (1): 231–279. Бибкод : 2022CirES...2..231В . дои : 10.1007/s43615-021-00084-3 . ISSN 2730-5988 . S2CID 238768104 .
- ↑ Перейти обратно: Перейти обратно: а б Диас, Крисандра Дж.; Дуглас, Кай Дж.; Канг, Калиса; Коларик, Эшлинн Л.; Малиновский, Родеон; Торрес-Тиджи, Ясин; Молино, Жуан В.; Бадари, Амр; Мэйфилд, Стивен П. (2023). «Развитие водорослей как устойчивого источника пищи» . Границы в питании . 9 . дои : 10.3389/fnut.2022.1029841 . ISSN 2296-861X . ПМК 9892066 . ПМИД 36742010 .
- ^ Кратко: Состояние мирового рыболовства и аквакультуры, 2018 г. (PDF) . ФАО. 2018.
- ^ Вердельо Виейра, Витор; Кадоре, Жан-Поль; Асьен, Ф. Габриэль; Бенеманн, Джон (январь 2022 г.). «Разъяснение наиболее важных концепций, связанных с сектором производства микроводорослей» . Процессы . 10 (1): 175. дои : 10.3390/pr10010175 . hdl : 10835/13146 . ISSN 2227-9717 .
- ^ Грин, Чарльз; Скотт-Бюхлер, Селина; Хауснер, Арджун; Джонсон, Закари; Лей, Синь Гэнь; Хантли, Марк (2022). «Преобразование будущего морской аквакультуры: подход экономики замкнутого цикла» . Океанография : 26–34. дои : 10.5670/oceanog.2022.213 . ISSN 1042-8275 .
- Новостная статья об исследовании: «Богатые питательными веществами водоросли могут помочь удовлетворить глобальный спрос на продовольствие: исследователи Корнелла» . CTVNews . 20 октября 2022 г. Проверено 17 ноября 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б Рейнольдс, Даман; Каминити, Джефф; Эдмундсон, Скотт; Гао, Сун; Уик, Макдональд; Хюземанн, Михаэль (12 июля 2022 г.). «Белки морских водорослей являются питательно ценными компонентами рациона человека» . Американский журнал клинического питания . 116 (4): 855–861. дои : 10.1093/ajcn/nqac190 . ISSN 0002-9165 . ПМИД 35820048 .
- ^ «Морские водоросли: растения или водоросли?» . Национальная прибрежная ассоциация Пойнт-Рейес . Проверено 1 декабря 2018 г.
- ^ Чжан, Личжу; Ляо, Вэй; Хуан, Яджун; Вэнь, Юйси; Чу, Яояо; Чжао, Чао (13 октября 2022 г.). «Глобальное выращивание и переработка морских водорослей за последние 20 лет» . Производство продуктов питания, переработка и питание . 4 (1). дои : 10.1186/s43014-022-00103-2 .
- ^ Бушманн, Алехандро Х.; Камю, Каролина; Инфанте, Хавьер; Неори, Амир; Израиль, Альваро; Эрнандес-Гонсалес, Мария К.; Переда, Сандра В.; Гомес-Пинчетти, Хуан Луис; Гольберг, Александр; Тадмор-Шалев, Нива; Кричли, Алан Т. (2 октября 2017 г.). «Производство морских водорослей: обзор глобального состояния эксплуатации, сельского хозяйства и новой исследовательской деятельности». Европейский журнал психологии . 52 (4): 391–406. Бибкод : 2017EJPhy..52..391B . дои : 10.1080/09670262.2017.1365175 . ISSN 0967-0262 . S2CID 53640917 .
- ^ Спросите, Э.И. (1990). Справочник по выращиванию Cottonii и Spinosum . Филиппины: FMC BioPolymer Corporation. п. 52.
- ↑ Перейти обратно: Перейти обратно: а б Джонс, Никола (15 марта 2023 г.). «Опубликованная ставка на водоросли» . Журнал Хакай . Проверено 19 марта 2023 г.
- ^ Ван, Тайпин; Ян, Чжаоцин; Дэвис, Джонатан; Эдмундсон, Скотт Дж. (1 мая 2022 г.). Количественная оценка биоэкстракции азота на фермах по выращиванию морских водорослей – пример моделирования и мониторинга в реальном времени в Худ-Канале, штат Вашингтон (технический отчет). Управление научно-технической информации . дои : 10.2172/1874372 .
- ^ Дуарте, Карлос М.; У, Цзяпин; Сяо, Си; Брюн, Аннетт; Краузе-Йенсен, Дорте (2017). «Может ли выращивание морских водорослей сыграть роль в смягчении последствий изменения климата и адаптации?» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00100 . hdl : 10754/623247 . ISSN 2296-7745 .
- ^ Биндофф, Нидерланды; Чунг, WWL; Кайро, JG; Аристеги, Ж.; и др. (2019). «Глава 5: Изменение океана, морских экосистем и зависимых сообществ» (PDF) . Специальный доклад МГЭИК об океане и криосфере в условиях меняющегося климата . стр. 447–587.
- ^ Чжу, Юньхуа; Шмидт, Эндрю; Вальдес, Питер; Сноуден-Свон, Лесли; Эдмундсон, Скотт (21 марта 2022 г.). «Гидротермальное сжижение и модернизация микроводорослей, выращенных в сточных водах: состояние технологий 2021 года» . дои : 10.2172/1855835 . ОСТИ 1855835 . S2CID 247648577 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Льюис, Дж. Г.; Стэнли, Северная Каролина; Гист, Г.Г. (1988). «9. Коммерческое производство гидроколлоидов водорослей». В Лемби, Калифорния; Вааланд, младший (ред.). Водоросли и дела человека . Издательство Кембриджского университета. ISBN 978-0-521-32115-0 .
- ^ «Макроцистис К. Агард 1820: 46» . База водорослей. Архивировано из оригинала 4 января 2009 года . Проверено 28 декабря 2008 г.
- ^ «Вторичные продукты бурых водорослей» . Исследования водорослей . Смитсоновский национальный музей естественной истории. Архивировано из оригинала 13 апреля 2009 года . Проверено 29 декабря 2008 г.
- ^ Чисти, Ю. (май – июнь 2007 г.). «Биодизель из микроводорослей» . Достижения биотехнологии . 25 (3): 294–306. doi : 10.1016/j.biotechadv.2007.02.001 . ПМИД 17350212 . S2CID 18234512 .
- ^ Ян, ЗК; Ню, Ю.Ф.; Ма, Ю.Х.; Сюэ, Дж.; Чжан, Миннесота; Ян, В.Д.; Лю, Дж. С.; Лу, Ш.; Гуань, Ю.; Ли, HY (4 мая 2013 г.). «Молекулярные и клеточные механизмы накопления нейтральных липидов у диатомовых водорослей после дефицита азота» . Биотехнология для биотоплива . 6 (1): 67. дои : 10.1186/1754-6834-6-67 . ПМЦ 3662598 . ПМИД 23642220 .
- ^ Вейффельс, Рене Х.; Барбоза, Мария Дж. (2010). «Взгляд на биотопливо из микроводорослей». Наука . 329 (5993): 796–799. Бибкод : 2010Sci...329..796W . дои : 10.1126/science.1189003 . ПМИД 20705853 . S2CID 206526311 .
- ^ Прочтите, Клэр Сьюэлл (1849). «О сельском хозяйстве Южного Уэльса: отчет о премии» . Журнал Королевского сельскохозяйственного общества Англии . 10 : 142–143.
- ^ Макхью, Деннис Дж. (2003). «9. Другие виды использования морских водорослей» . Руководство по производству морских водорослей: Технический документ ФАО по рыболовству 441 . Рим: Департамент рыболовства и аквакультуры, Продовольственная и сельскохозяйственная организация (ФАО) ООН. ISBN 978-92-5-104958-7 . Архивировано из оригинала 28 декабря 2008 года.
- ^ Юнг, Фредерик; Крюгер-Генге, Энн; Куппер, Ж.-Х.; Вальдек, П. (апрель 2019 г.). «Спирулина Platensis — суперпродукт?» . Исследовательские ворота . 5:43 . Проверено 21 декабря 2020 г. .
- ^ Симунс, Фредерик Дж. (1991). «6. Водоросли и другие водоросли». Еда в Китае: культурное и историческое исследование . ЦРК Пресс. стр. 179–190. ISBN 978-0-936923-29-1 .
- ^ Мортон, Стив Л. «Современное использование культивируемых водорослей» . Этноботанические листовки . Университет Южного Иллинойса в Карбондейле. Архивировано из оригинала 23 декабря 2008 года . Проверено 26 декабря 2008 г.
- ^ Мондрагон, Дженнифер; Мондрагон, Джефф (2003). Водоросли Тихоокеанского побережья . Монтерей, Калифорния: Публикации Sea Challengers. ISBN 978-0-930118-29-7 .
- ^ «Durvillaea antarctica (Chamisso) Hariot» . База водорослей.
- ^ «Как морские водоросли могут помочь накормить мир» . Всемирный экономический форум . 25 октября 2017 г. Проверено 21 июня 2018 г.
- ^ «Одно из решений проблемы глобального голода может быть найдено на дне океана» . Всемирный экономический форум . 15 декабря 2017 года . Проверено 21 июня 2018 г.
- ^ «Водоросли: прудовая нечисть или еда будущего?» . Как все работает . 12 июня 2018 года . Проверено 21 июня 2018 г.
- ^ Рани, Комал; Сандал, Ниди; Саху, ПК (2018). «Всесторонний обзор хлореллы - ее состав, польза для здоровья, рыночный и нормативный сценарий» (PDF) . Журнал фармацевтических инноваций . 7 (7): 585. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 21 декабря 2020 г. .
- ^ Бигоньо, К.; Хозин-Гольдберг И.; Буссиба, С.; Воншак, А.; Коэн, З. (2002). «Липидный и жирнокислотный состав зеленой маслянистой водоросли Parietochromis incisa, богатейшего растительного источника арахидоновой кислоты» . Фитохимия . 60 (5): 497–503. Бибкод : 2002PChem..60..497B . дои : 10.1016/S0031-9422(02)00100-0 . ПМИД 12052516 . Архивировано из оригинала 1 октября 2017 года.
- ^ Обри, Эллисон (1 ноября 2007 г.). «Получение пищи для мозга прямо из источника» . Утренний выпуск . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Архивировано из оригинала 3 ноября 2007 года.
- ^ «Переосмысление водорослей» . Австралийская радиовещательная корпорация. 12 октября 2016 г. Архивировано из оригинала 2 февраля 2017 г. Проверено 26 января 2017 г.
- ^ Моррисси, Дж.; Джонс, М.С.; Харриотт, В. (1988). «Круговорот питательных веществ в аквариуме Большого Барьерного рифа – материалы 6-го Международного симпозиума по коралловым рифам, Австралия» . РифБаза. Архивировано из оригинала 23 февраля 2015 года.
- ^ Вераарт, Аннелис Дж.; Романи, Анна М.; Торнес, Элизабет; Сабатер, Сержи (2008). «Реакция водорослей на обогащение питательными веществами в лесном олиготрофном ручье» . Журнал психологии . 44 (3): 564–572. Бибкод : 2008JPcgy..44..564V . дои : 10.1111/j.1529-8817.2008.00503.x . ПМИД 27041416 . S2CID 2040067 . Архивировано из оригинала 1 октября 2010 года.
- ^ «Водоросли: злая зеленая машина для чистки» . Служба сельскохозяйственных исследований Министерства сельского хозяйства США. 7 мая 2010 г. Архивировано из оригинала 19 октября 2010 г.
- ^ «Биополимеры водорослей, компании, производство, рынок – Масляные водоросли – Масло из водорослей» . Oilgae.com . Проверено 18 ноября 2017 г.
- ^ «Возобновляемые шлепанцы: ученые производят обувь № 1 в мире из водорослей» . ЗМЭ Наука . 9 октября 2017 года . Проверено 18 ноября 2017 г.
- ^ «Первая в мире доска для серфинга с водорослями поднимает волны в Сан-Диего» . Energy.gov.ru . Проверено 18 ноября 2017 г.
- ^ Каппителли, Франческа; Сорлини, Клаудия (2008). «Микроорганизмы атакуют синтетические полимеры в предметах, представляющих наше культурное наследие» . Прикладная и экологическая микробиология . 74 (3): 564–569. Бибкод : 2008ApEnM..74..564C . дои : 10.1128/АЕМ.01768-07 . ПМК 2227722 . ПМИД 18065627 .
- ^ Арад, Шошана; Сфарим, Ишай (1998). «Производство ценных продуктов из микроводорослей: развивающаяся агроиндустрия». В Альтмане, Ари (ред.). Сельскохозяйственная биотехнология . Книги по почвам, растениям и окружающей среде. Том. 61. ЦРК Пресс. п. 638. ИСБН 978-0-8247-9439-2 .
- ^ Ратбун, К.; Дойл, А.; Уотерхаус, Т. (июнь 1994 г.). «Измерение хлорофиллов и каротиноидов водорослей с помощью ВЭЖХ» (PDF) . Протоколы совместных исследований глобальных потоков океана . 13 : 91–96. Архивировано из оригинала (PDF) 4 марта 2016 года . Проверено 7 июля 2014 г.
- ^ Латаса, М.; Бидигаре, Р. (1998). «Сравнение популяций фитопланктона Аравийского моря во время весеннего межмуссонного периода и юго-западного муссона 1995 года, как описано с помощью пигментов, проанализированных методом ВЭЖХ». Глубоководные исследования. Часть II . 45 (10–11): 2133–2170. Бибкод : 1998DSRII..45.2133L . дои : 10.1016/S0967-0645(98)00066-6 .
Библиография [ править ]
Общие [ править ]
- Чепмен, виджей (1950). Морские водоросли и их использование . Лондон: Метуэн. ISBN 978-0-412-15740-0 .
- Фрич, Ф.Е. (1945) [1935]. Строение и размножение водорослей . Том. Я и II. Издательство Кембриджского университета.
- ван ден Хук, К.; Манн, Д.Г.; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета.
- Кассинджер, Рут (2020). Слизь: как водоросли создали нас, досаждают нам и могут нас спасти . Маринер.
- Лемби, Калифорния; Уолланд, младший (1988). Водоросли и дела человека . Издательство Кембриджского университета. ISBN 978-0-521-32115-0 .
- Мамфорд, ТФ; Миура, А. (1988). « Порфира как продукт питания: выращивание и экономика». В Лемби, Калифорния; Вааланд, младший (ред.). Водоросли и дела человека . Издательство Кембриджского университета. стр. 87–117. ISBN 978-0-521-32115-0 . .
- Раунд, FE (1981). Экология водорослей . Лондон: Издательство Кембриджского университета. ISBN 978-0-521-22583-0 .
- Смит, GM (1938). Криптогаминая ботаника . Том. Я. Нью-Йорк: МакГроу-Хилл.
- Спросите, Э.И. (1990). Справочник по выращиванию Cottonii и Spinosum . FMC BioPolymer Corporation. Филиппины.
Региональный [ править ]
Великобритания и Ирландия [ править ]
- Броди, Джульетта; Берроуз, Элси М.; Чемберлен, Ивонн М.; Кристенсен, Тайге; Диксон, Питер Стэнли; Флетчер, РЛ; Хоммерсанд, Макс Х.; Ирвин, Линда М.; Мэггс, Кристин А. (1977–2003). Морские водоросли Британских островов: совместный проект Британского психологического общества и Британского музея (естественной истории) . Лондон / Андовер: Британский музей естественной истории, HMSO / Intercept. ISBN 978-0-565-00781-2 .
- Куллинан, Джон П. (1973). Психология южного побережья Ирландии . Корк: Издательство Коркского университета.
- Харди, ФГ; Аспиналл, Р.Дж. (1988). Атлас морских водорослей Нортумберленда и Дарема . Музей Хэнкока, Университет Ньюкасл-апон-Тайн: Центр биологических записей Нортумберленда. ISBN 978-0-9509680-5-6 .
- Харди, ФГ; Гири, Майкл Д .; Арнольд, Генри Р. (2006). Контрольный список и Атлас морских водорослей Великобритании и Ирландии (пересмотренная редакция). Лондон: Британское психологическое общество. ISBN 978-3-906166-35-3 .
- Джон, DM; Уиттон, бакалавр; Брук, Дж. А. (2002). Пресноводная водорослевая флора Британских островов . Кембридж / Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-77051-4 .
- Найт, Марджери; Парк, Мэри В. (1931). Мэнские водоросли: исследование водорослей южной оконечности острова Мэн . Мемуары Ливерпульского комитета морской биологии о типичных британских морских растениях и животных. Том. ХХХ. Ливерпуль: Университетское издательство.
- Мортон, Осборн (1994). Морские водоросли Северной Ирландии . Белфаст: Ольстерский музей. ISBN 978-0-900761-28-7 .
- Мортон, Осборн (1 декабря 2003 г.). «Морские макроводоросли графства Донегол, Ирландия». Бюллетень Ирландского биогеографического общества . 27 : 3–164.
Австралия [ править ]
- Хейсман, Дж. М. (2000). Морские растения Австралии . Издательство Университета Западной Австралии. ISBN 978-1-876268-33-6 .
Новая Зеландия [ править ]
- Чепмен, Валентин Джексон; Линдауэр, Фольксваген; Эйкен, М.; Дромгул, Ф.И. (1970) [1900, 1956, 1961, 1969]. Морские водоросли Новой Зеландии . Лондон / Лер, Германия: Лондонское Линнеевское общество / Крамер.
Европа [ править ]
- Кабиок, Жаклин; Флок, Жан-Ив; Ле Токен, Ален; Будуреск, Шарль-Франсуа; Мейнес, Александр; Верлак, Марк (1992). Путеводитель по морским водорослям Европы: Ла-Манш/Атлантика-Средиземноморье (на французском языке). Лозанна, Швейцария: Делашо и Нистле. ISBN 978-2-603-00848-5 .
- Гайрал, Полетт (1966). Водоросли французских побережий (Ланш и Атлантика), фундаментальные представления по экологии, биологии и систематике морских водорослей (на французском языке). Париж: Дуэн, Дерен и др. Cie.
- Гири, Майкл. Д .; Бланден, Г. (1991). Ресурсы морских водорослей в Европе: использование и потенциал . Джон Уайли и сыновья. ISBN 978-0-471-92947-5 .
- Мигес Родригес, Луис (1998). Морские водоросли из Галичины: Биология, гастрономия, промышленность (на галисийском языке). Виго: Общие издания Галисии. ISBN 978-84-8302-263-4 .
- Отеро, Дж. (2002). Путеводитель по макроводорослям Галиции (на галисийском языке). Ла-Корунья: Baía Editions. ISBN 978-84-89803-22-0 .
- Барбара, И.; Кремадес, Дж. (1993). Путеводитель по водорослям галисийского побережья (на испанском языке). Ла-Корунья: Совет Ла-Коруньи - Дом наук.
Арктика [ править ]
- Кьельман, Франс Рейнхольд (1883). Водоросли Северного Ледовитого моря: обзор видов с изложением общих признаков и развития флоры . Том. 20. Стокгольм: Кунгл. Шведская ветенскапсакадемическая академия. стр. 1–350.
Гренландия [ править ]
- Лунд, Сорен Йенсен (1959). Морские водоросли Восточной Гренландии . Копенгаген: CA Reitzel. 9584734.
Фарерские острова [ править ]
- Боргесен, Фредерик (1970) [1903]. «Морские водоросли». В Утеплении, Юджин (ред.). Ботаника Фарерских островов на основе датских исследований, Часть II . Копенгаген: Издательство Северных стран. стр. 339–532. .
Канарские острова [ править ]
- Боргесен, Фредерик (1936) [1925, 1926, 1927, 1929, 1930]. Морские водоросли с Канарских островов . Копенгаген: Бьянко Лунос.
Марокко [ править ]
- Гайрал, Полетт (1958). Водоросли марокканского атлантического побережья (на французском языке). Касабланка: Рабат [Общество естественных и физических наук Марокко].
Южная Африка [ править ]
- Стегенга, Х.; Болтон, Джей-Джей; Андерсон, Р.Дж. (1997). Морские водоросли западного побережья Южной Африки . Болюсный гербарий, Кейптаунский университет. ISBN 978-0-7992-1793-3 .
Северная Америка [ править ]
- Эбботт, Айова; Холленберг, Дж.Дж. (1976). Морские водоросли Калифорнии . Калифорния: Издательство Стэнфордского университета. ISBN 978-0-8047-0867-8 .
- Грисон, Филип Э. (1982). Аннотированный ключ к идентификации часто встречающихся и доминирующих родов водорослей, наблюдаемых в фитопланктоне США . Вашингтон, округ Колумбия: Министерство внутренних дел США, Геологическая служба . Проверено 19 декабря 2008 г.
- Тейлор, Уильям Рэндольф (1969) [1937, 1957, 1962]. Морские водоросли северо-восточного побережья Северной Америки . Анн-Арбор: Издательство Мичиганского университета. ISBN 978-0-472-04904-2 .
- Вер, доктор медицинских наук; Шит, Р.Г. (2003). Пресноводные водоросли Северной Америки: экология и классификация . Академическая пресса. ISBN 978-0-12-741550-5 .
Внешние ссылки [ править ]
- Гири, Майкл ; Гири, Венди. «АлгаБаза» . – база данных всех названий водорослей, включая изображения, номенклатуру, таксономию, распространение, библиографию, использование, экстракты.
- «Водоросли – клеточно-центрированная база данных» . CCDb.UCSD.edu . Сан-Диего: Калифорнийский университет.
- Андерсон, Дон; Кифер, Брюс; Кляйндинст, Джуди; Шонесси, Кэти; Джойс, Кэтрин; Фино, Даниэль; Шеперд, Адам (2007). «Вредные водоросли» . Национальное управление США по вредному цветению водорослей. Архивировано из оригинала 5 декабря 2008 года . Проверено 19 декабря 2008 г.
- «О водорослях» . NMH.ac.uk. Музей естественной истории, Великобритания.