Растительный гормон

Растительные гормоны (или фитогормоны ) — это сигнальные молекулы , вырабатываемые растениями и встречающиеся в чрезвычайно низких концентрациях . Растительные гормоны контролируют все аспекты роста и развития растений, включая эмбриогенез . [1] регуляция размера органов , защита от патогенов , [2] [3] стрессоустойчивость [4] [5] и репродуктивное развитие. [6] В отличие от животных (у которых выработка гормонов ограничена специализированными железами ), каждая растительная клетка способна вырабатывать гормоны. [7] [8] Вент и Тиманн придумали термин «фитогормон» и использовали его в названии своей книги 1937 года. [9]
Фитогормоны встречаются во всем царстве растений и даже в водорослях , где они имеют функции, аналогичные тем, которые наблюдаются у сосудистых растений («высших растений») . [10] Некоторые фитогормоны встречаются и в микроорганизмах , например в одноклеточных грибах и бактериях , однако в этих случаях они не играют гормональной роли и их лучше рассматривать как вторичные метаболиты . [11]
Характеристики
[ редактировать ]
Слово «гормон» происходит от греческого языка и означает «привести в движение» . Растительные гормоны влияют на экспрессию генов и уровни транскрипции , клеточное деление и рост. Они естественным образом производятся внутри растений, хотя очень похожие химические вещества производятся грибами и бактериями, которые также могут влиять на рост растений. [12] Большое количество родственных соединений синтезируется химических человеком. Они используются для регулирования роста культурных растений, сорняков , а также выращенных in vitro растений и растительных клеток; эти искусственные соединения называются регуляторами роста растений ( РГР ). На ранних этапах изучения растительных гормонов обычно использовался термин «фитогормон», но сейчас он применяется менее широко.
Растительные гормоны – это не питательные вещества , а химические вещества , которые в небольших количествах способствуют росту и влияют на него. [13] развитие и дифференцировка клеток и тканей . Биосинтез растительных гормонов в тканях растений часто носит диффузный и не всегда локализованный характер. У растений отсутствуют железы для производства и хранения гормонов, потому что, в отличие от животных, у которых есть две системы кровообращения ( лимфатическая и сердечно-сосудистая ), питаемые сердцем , которое перемещает жидкости по телу, растения используют более пассивные средства для перемещения химических веществ по своему телу. Растения используют простые химические вещества в качестве гормонов, которые легче перемещаются по тканям. Они часто производятся и используются локально в организме растения. Растительные клетки вырабатывают гормоны, которые влияют даже на разные участки клетки, производящие гормон.
Гормоны транспортируются внутри растения с помощью четырех типов движений. Для локализованного движения поток цитоплазмы внутри клеток и медленная диффузия ионов и молекул используется между клетками. Сосудистые ткани используются для перемещения гормонов из одной части растения в другую; к ним относятся ситовидные трубки или флоэма , которые перемещают сахара из листьев к корням и цветам, и ксилема , которая перемещает воду и минеральные растворы от корней к листве .
Не все растительные клетки реагируют на гормоны, но те клетки, которые реагируют, запрограммированы реагировать на определенные моменты цикла роста. Наибольшие эффекты наблюдаются на определенных этапах жизни клетки, а ослабленные эффекты наблюдаются до или после этого периода. Растениям нужны гормоны в определенные моменты роста и в определенных местах. Им также необходимо отключить эффекты гормонов, когда они больше не нужны. Производство гормонов очень часто происходит в местах активного роста внутри меристем , прежде чем клетки полностью дифференцируются. После производства их иногда перемещают в другие части завода, где они вызывают немедленный эффект; или они могут храниться в клетках для последующего высвобождения. Растения используют разные пути для регулирования количества внутренних гормонов и смягчения их воздействия; они могут регулировать количество химических веществ, используемых для биосинтеза гормонов. Они могут хранить их в клетках, инактивировать или уничтожать уже сформированные гормоны путем конъюгируя их с углеводами , аминокислотами или пептидами . Растения также могут химически расщеплять гормоны, эффективно уничтожая их. Растительные гормоны часто регулируют концентрации других растительных гормонов. [14] Растения также перемещают гормоны по растению, разбавляя их концентрацию.
Концентрация гормонов, необходимых для реакций растений, очень низкая (10 −6 до 10 −5 моль / л ). Из-за таких низких концентраций было очень сложно изучать растительные гормоны, и только с конца 1970-х годов ученые смогли начать собирать воедино их влияние и взаимосвязь с физиологией растений. [15] Большая часть ранних работ по растительным гормонам включала изучение растений, у которых был генетический дефицит одного из них, или использование культур тканевых растений, выращенных in vitro , которые подвергались воздействию различных соотношений гормонов, и сравнение полученного роста. Самые ранние научные наблюдения и исследования датируются 1880-ми годами; определение и наблюдение растительных гормонов, а также их идентификация продолжались в течение следующих 70 лет.
Синергизм растительных гормонов означает, что действие двух или более гормонов приводит к эффекту, превышающему эффект отдельных эффектов. Например, ауксины и цитокинины часто действуют совместно во время деления и дифференцировки клеток. Оба гормона играют ключевую роль в регуляции клеточного цикла, но когда они объединяются, их синергетические взаимодействия могут усиливать пролиферацию клеток и органогенез более эффективно, чем любой из них по отдельности.
Классы
[ редактировать ]Различные гормоны можно разделить на разные классы в зависимости от их химической структуры. Внутри каждого класса гормонов химическая структура может различаться, но все представители одного и того же класса оказывают сходное физиологическое действие. Первоначальные исследования растительных гормонов выявили пять основных классов: абсцизовая кислота, ауксины, брассиностероиды, цитокинины и этилен. [16] Позднее этот список был расширен, и теперь основными растительными гормонами считаются брассиностероиды, жасмонаты, салициловая кислота и стриголактоны. Кроме того, существует несколько других соединений, которые выполняют функции, аналогичные основным гормонам, но их статус как настоящих гормонов все еще обсуждается.
Абсцизовая кислота
[ редактировать ]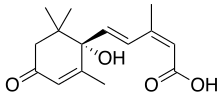
Абсцизовая кислота (также называемая АБК) является одним из наиболее важных ингибиторов роста растений. Он был открыт и исследован под двумя разными названиями: дормин и абсцицин II , прежде чем его химические свойства были полностью известны. Как только было установлено, что эти два соединения одинаковы, их назвали абсцизовой кислотой. Название связано с тем фактом, что он содержится в высоких концентрациях в недавно опадающих или только что опавших листьях.
Этот класс РГР состоит из одного химического соединения, которое обычно вырабатывается в листьях растений и происходит из хлоропластов , особенно когда растения находятся в состоянии стресса. В целом он действует как ингибирующее химическое соединение, влияющее на рост почек , а также на покой семян и почек. Он вызывает изменения в апикальной меристеме , вызывая покой почек и превращение последнего набора листьев в защитные покровы почек. Поскольку он был обнаружен в только что оторванных листьях, первоначально предполагалось, что он играет роль в процессах естественного опадения листьев, но дальнейшие исследования опровергли это. У видов растений из частей мира с умеренным климатом абсцизовая кислота играет роль в состоянии покоя листьев и семян, подавляя рост, но, когда она рассеивается из семян или почек, начинается рост. У других растений, когда уровень АБК снижается, рост начинается с повышения уровня гиббереллина . Без АБК почки и семена начали бы расти в теплые периоды зимы и погибли бы при повторном замерзании. Поскольку АБК медленно рассеивается из тканей, и ее действие требует времени, чтобы компенсироваться другими растительными гормонами, происходит задержка физиологических путей, которая обеспечивает некоторую защиту от преждевременного роста. Абсцизовая кислота накапливается в семенах во время созревания плодов, предотвращая прорастание семян внутри плода или перед зимой. Эффекты абсцизовой кислоты ослабляются в тканях растения при низких температурах или в результате ее удаления водой, промывающей ткани и вымывающими их из них, выводя семена и почки из состояния покоя. [17]
АБК существует во всех частях растения, и ее концентрация в любой ткани, по-видимому, опосредует ее действие и действует как гормон; его деградация или, точнее, катаболизм внутри растения влияет на метаболические реакции, рост клеток и выработку других гормонов. [18] Растения начинают жизнь как семена с высоким уровнем АБК. Непосредственно перед прорастанием семян уровень АБК снижается; в период прорастания и раннего роста рассады уровень АБК снижается еще больше. Когда растения начинают давать побеги с полностью функциональными листьями, уровень АБК снова начинает увеличиваться, замедляя рост клеток в более «зрелых» участках растения. Стресс от воды или хищников влияет на выработку и скорость катаболизма АБК, опосредуя еще один каскад эффектов, вызывающих специфические реакции целевых клеток. Ученые все еще изучают сложные взаимодействия и эффекты этого и других фитогормонов.
У растений, находящихся в условиях водного стресса, АБК играет роль в закрытии устьиц . Вскоре после того, как растения испытывают водный стресс и корни испытывают дефицит воды, сигнал перемещается вверх к листьям, вызывая образование там предшественников АБК, которые затем перемещаются к корням. Затем корни выделяют АБК, которая транспортируется в листву через сосудистую систему. [19] и модулирует поглощение калия и натрия замыкающими клетками , которые затем теряют тургор , закрывая устьица. [20] [21]
Ауксины
[ редактировать ]
Ауксины — это соединения, которые положительно влияют на увеличение клеток, образование почек и зарождение корней. Они также способствуют выработке других гормонов и в сочетании с цитокининами контролируют рост стеблей, корней и плодов, а также превращают стебли в цветы. [22] Ауксины были первым классом обнаруженных регуляторов роста.Голландский биолог Фриц Вармолт Вент впервые описал ауксины. [23] Они влияют на удлинение клеток, изменяя пластичность клеточной стенки. Они стимулируют камбий , подтип клеток меристемы , к делению, а в стеблях вызывают вторичной ксилемы дифференцировку .
Ауксины подавляют рост почек ниже стебля (феномен, известный как апикальное доминирование) , а также способствуют боковых и придаточных корней развитию и росту . Опадение листьев инициируется прекращением выработки ауксинов точкой роста растения. Ауксины в семенах регулируют синтез специфических белков, [24] поскольку они развиваются внутри цветка после опыления , в результате чего из цветка развивается плод, содержащий развивающиеся семена.
В больших концентрациях ауксины часто токсичны для растений; они наиболее токсичны для двудольных и менее токсичны для однодольных . Благодаря этому свойству синтетические ауксиновые гербициды, в том числе 2,4-дихлорфеноксиуксусная кислота (2,4-Д) и 2,4,5-трихлорфеноксиуксусная кислота были разработаны (2,4,5-Т), которые используются для борьбы с сорняками путем дефолиации. . Ауксины, особенно 1-нафталинуксусная кислота (NAA) и индол-3-масляная кислота (IBA), также широко применяются для стимуляции роста корней при черенках растений . Наиболее распространенным ауксином, обнаруженным в растениях, является индол-3-уксусная кислота (ИУК).
Брассиностероиды
[ редактировать ]
Брассиностероиды (БР) представляют собой класс полигидроксистероидов, единственный пример стероидных гормонов в растениях. Брассиностероиды контролируют удлинение и деление клеток, гравитропизм , устойчивость к стрессу и дифференцировку ксилемы . Они подавляют рост корней и опадение листьев. Брассинолид был первым брассиностероидом, который был идентифицирован и был выделен из экстракта пыльцы рапса ( Brassica napus ) в 1979 году. [25] Брассиностероиды — класс стероидных фитогормонов растений, которые регулируют многочисленные физиологические процессы. Этот растительный гормон был идентифицирован Mitchell et al. которые извлекли ингредиенты из пыльцы Brassica и обнаружили, что основным активным компонентом извлеченных ингредиентов был брассинолид . [26] Это открытие означало открытие нового класса растительных гормонов, названных брассиностероидами. Эти гормоны действуют очень похоже на стероидные гормоны животных, способствуя росту и развитию.
У растений эти стероидные гормоны играют важную роль в удлинении клеток посредством передачи сигналов BR. [27] Брассиностероидный рецептор брассиностероидно-нечувствительный 1 (BRI1) является основным рецептором этого сигнального пути. Этот рецептор BRI1 был обнаружен Clouse et al. который сделал открытие, ингибируя BR и сравнив его с диким типом Arabidopsis. У мутанта BRI1 наблюдалось несколько проблем, связанных с ростом и развитием, таких как карликовость , снижение удлинения клеток и другие физические изменения. [26] Эти результаты означают, что растения, правильно экспрессирующие брассиностероиды, растут быстрее, чем их мутантные аналоги. Брассиностероиды связываются с BRI1, локализованным на плазматической мембране. [28] что приводит к сигнальному каскаду, который дополнительно регулирует удлинение клеток. Однако этот сигнальный каскад в настоящее время не совсем понятен. Считается, что происходит то, что BR связывается с комплексом BAK1, что приводит к каскаду фосфорилирования . [29] Этот каскад фосфорилирования затем вызывает деактивацию BIN2, что вызывает высвобождение факторов транскрипции . [29] Эти высвободившиеся факторы транскрипции затем связываются с ДНК, что приводит к процессам роста и развития. [29] и позволяет растениям реагировать на абиотические стрессоры . [30]
Цитокинины
[ редактировать ]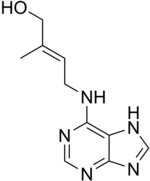
Цитокинины (ЦК) представляют собой группу химических веществ, которые влияют на деление клеток и образование побегов. Они также помогают задержать старение тканей, отвечают за транспорт ауксина по всему растению и влияют на длину междоузлий и рост листьев. Раньше их называли кининами, когда их впервые выделили из дрожжевых клеток. Цитокинины и ауксины часто работают вместе, и соотношение этих двух групп растительных гормонов влияет на большинство основных периодов роста в течение жизни растения. Цитокинины противодействуют апикальному доминированию, индуцированному ауксинами; в сочетании с этиленом они способствуют опадению листьев, частей цветков и плодов. [31]
Среди растительных гормонов известно, что три гормона способствуют иммунологическому взаимодействию: этилен (ET), салицилаты (SA) и жасмонаты (JA), однако было проведено больше исследований для определения роли, которую в этом играют цитокинины. Имеющиеся данные свидетельствуют о том, что цитокинины задерживают взаимодействие с патогенами, демонстрируя признаки того, что они могут вызывать устойчивость к этим патогенным бактериям. Соответственно, уровни ЦК выше у растений, обладающих повышенной устойчивостью к патогенам, по сравнению с теми, которые более восприимчивы. [32] Например, устойчивость к патогенам, включающим цитокинины, была протестирована с использованием видов Arabidopsis путем обработки их природным ЦК (транс-зеатином), чтобы увидеть их реакцию на бактерии Pseudomonas syringa . Исследования табака показывают, что избыточная экспрессия CK, индуцирующая гены IPT, приводит к повышенной устойчивости, тогда как избыточная экспрессия CK-оксидазы приводит к повышенной восприимчивости к патогену, а именно P. syringae .
Хотя между этим гормоном и физическим поведением растений нет особой связи, внутри растения в ответ на него происходят поведенческие изменения. Эффекты цитокининовой защиты могут включать в себя появление и рост микробов (задержку старения листьев), реконфигурацию вторичного метаболизма или даже индуцирование образования новых органов, таких как галлы или узелки. [33] Все эти органы и соответствующие им процессы используются для защиты растений от биотических/абиотических факторов.
Этилен
[ редактировать ]
В отличие от других основных растительных гормонов, этилен представляет собой газ и очень простое органическое соединение, состоящее всего из шести атомов. Он образуется в результате распада метионина , аминокислоты, которая присутствует во всех клетках. Этилен имеет очень ограниченную растворимость в воде и поэтому не накапливается внутри клетки, обычно диффундируя из клетки и покидая растение. Его эффективность как растительного гормона зависит от скорости его производства и скорости выхода в атмосферу. Этилен вырабатывается быстрее в быстро растущих и делящихся клетках, особенно в темноте. Новые побеги и недавно проросшие саженцы производят больше этилена, чем может покинуть растение, что приводит к повышенному количеству этилена, препятствующему расширению листьев (см. Гипонастическая реакция ).
Когда новый побег подвергается воздействию света, реакции, опосредованные фитохромом в клетках растения, дают сигнал о снижении выработки этилена, что позволяет листьям разрастаться. Этилен влияет на рост и форму клеток; Когда растущий побег или корень, находясь под землей, сталкивается с препятствием, выработка этилена значительно увеличивается, предотвращая удлинение клеток и вызывая набухание стебля. В результате более толстый стержень становится более прочным и с меньшей вероятностью прогнется под давлением, поскольку он прижимается к объекту, препятствующему его пути к поверхности. реакцию стебля Если побег не достигает поверхности и стимул этилена становится продолжительным, это влияет на естественную геотропную , который заключается в том, чтобы расти вертикально, позволяя ему расти вокруг объекта. Исследования показывают, что этилен влияет на диаметр и высоту ствола: когда стволы деревьев подвергаются воздействию ветра, вызывающего боковое напряжение, происходит большее производство этилена, в результате чего стволы и ветви деревьев становятся более толстыми и крепкими.
Этилен также влияет на созревание плодов. Обычно, когда семена созревают, выработка этилена увеличивается и накапливается в плодах, что приводит к климактерическому событию непосредственно перед распространением семян. Ядерный белок этилен-нечувствительный2 (EIN2) регулирует выработку этилена и, в свою очередь, регулирует другие гормоны, включая АБК и гормоны стресса. [34] Диффузия этилена из растений под водой сильно затруднена. Это увеличивает внутреннюю концентрацию газа. У многих водных и полуводных видов (например, Callitriche platycarpus , риса и Rumex palustris ) накопленный этилен сильно стимулирует удлинение вверх. Эта реакция является важным механизмом адаптивного выхода из погружения, который позволяет избежать удушья, возвращая побег и листья в контакт с воздухом, одновременно позволяя высвободить захваченный этилен. [35] [36] [37] [38] По крайней мере один вид ( Potamogeton pectinatus ). [39] Было обнаружено, что он не способен производить этилен, сохраняя при этом традиционную морфологию. Это говорит о том, что этилен является настоящим регулятором, а не обязательным требованием для построения основного плана организма растения.
Гиббереллины
[ редактировать ]
Гиббереллины (ГК) включают в себя широкий спектр химических веществ, которые естественным образом производятся растениями и грибами. Впервые они были обнаружены, когда японские исследователи, в том числе Эйити Куросава, заметили химическое вещество, вырабатываемое грибом Gibberella fujikuroi , которое вызывает аномальный рост растений риса. [40] Позже было обнаружено, что ГК также производятся самими растениями и контролируют множество аспектов развития на протяжении жизненного цикла. Синтез ГК сильно регулируется в семенах при прорастании, и его присутствие необходимо для прорастания. У проростков и взрослых ГК сильно способствуют удлинению клеток. ГК также способствуют переходу между вегетативным и репродуктивным ростом, а также необходимы для функционирования пыльцы во время оплодотворения. [41]
Гиббереллины нарушают состояние покоя (в активной стадии) семян и почек и способствуют увеличению высоты растения. Это помогает в росте стебля [ нужна ссылка ]
Жасмонаты
[ редактировать ]
Жасмонаты (ЖК) — это гормоны на основе липидов, которые первоначально были выделены из жасминового масла. [42] ЖК особенно важны в ответной реакции растений на нападение травоядных и некротрофных патогенов . [43] Наиболее активной ЖК растений является жасмоновая кислота . Жасмоновая кислота может далее метаболизироваться в метилжасмонат (MeJA), который представляет собой летучее органическое соединение . Это необычное свойство означает, что MeJA может действовать как воздушный сигнал, сообщающий о нападении травоядных на другие удаленные листья одного растения, и даже как сигнал соседним растениям. [44] Считается, что помимо своей защитной роли ЖК также играют роль в прорастании семян, хранении белка в семенах и росте корней. [43]
Было показано, что ЖК взаимодействуют с сигнальным путем других гормонов по механизму, описанному как «перекрестные помехи». Классы гормонов могут оказывать как отрицательное, так и положительное влияние на сигнальные процессы друг друга. [45]
метиловый эфир жасмоновой кислоты (JAME) регулирует генетическую экспрессию в растениях. Было показано, что [46] Они действуют на сигнальные пути в ответ на травоядность и усиливают экспрессию защитных генов. [47] Жасмонилизолейцин (JA-Ile) накапливается в ответ на травоядность, что вызывает усиление экспрессии защитных генов за счет высвобождения факторов транскрипции. [47]
Мутанты жасмоната легче потребляются травоядными животными, чем растения дикого типа, что указывает на то, что ЖК играют важную роль в обеспечении защиты растений. Когда травоядные животные перемещаются по листьям растений дикого типа, они достигают такой же массы, что и травоядные животные, которые потребляют только мутантные растения, что означает, что действие ЖА локализовано в местах обитания травоядных животных. [48] Исследования показали, что между путями защиты существуют значительные перекрестные помехи. [49]
Салициловая кислота
[ редактировать ]
Салициловая кислота (СК) — гормон, по структуре родственный бензойной кислоте и фенолу . Первоначально он был выделен из экстракта коры белой ивы ( Salix alba ) и представляет большой интерес для медицины человека, поскольку является предшественником болеутоляющего средства аспирина . У растений СК играет решающую роль в защите от биотрофных патогенов. Подобно JA, SA также может метилироваться . Как и MeJA, метилсалицилат летуч и может подавать соседним растениям сигнал на большие расстояния, предупреждая об атаке патогенов. Помимо своей защитной роли, СК также участвует в реакции растений на абиотический стресс, особенно на засуху, экстремальные температуры, тяжелые металлы и осмотический стресс. [50]
Салициловая кислота (СК) служит ключевым гормоном врожденного иммунитета растений, включая устойчивость как местных, так и системных тканей к биотическим атакам, реакциям гиперчувствительности и гибели клеток. Некоторые из влияний SA на растения включают прорастание семян, рост клеток, дыхание, закрытие устьиц, экспрессию генов, связанных со старением, реакции на абиотические и биотические стрессы, базальную термоустойчивость и урожайность плодов. Возможная роль салициловой кислоты в передаче сигнала об устойчивости к болезням была впервые продемонстрирована путем инъекции листьев устойчивого табака СК. [51] В результате инъекции SA стимулировали накопление патоген-связанного белка (PR) и повышали устойчивость к инфекции вируса табачной мозаики (TMV). Воздействие патогенов вызывает каскад реакций в растительных клетках. Биосинтез СК увеличивается за счет изохоризмат-синтазы (ICS) и фенилаланин-аммиаклиазы (PAL) в пластидах. [52] Было замечено, что во время взаимодействия растений и микробов в рамках защитных механизмов СК сначала накапливается в местной инфицированной ткани, а затем распространяется по всему растению, вызывая системную приобретенную устойчивость в неинфицированных дистальных частях растения. Таким образом, при увеличении внутренней концентрации СК растения смогли создать устойчивые барьеры для патогенов и других неблагоприятных условий окружающей среды. [53]
Стриголактоны
[ редактировать ]
Стриголактоны (SL) были первоначально обнаружены в ходе исследований прорастания паразитического сорняка Striga lutea . Было обнаружено, что прорастание видов Striga стимулировалось присутствием соединения, выделяемого корнями растения-хозяина. [54] Позже было показано, что СЛ, выделяющиеся в почву, также способствуют росту симбиотических арбускулярных микоризных (АМ) грибов. [55] Совсем недавно была выявлена еще одна роль СЛ в ингибировании ветвления побегов. [56] Это открытие роли SL в ветвлении побегов привело к резкому увеличению интереса к этим гормонам, и с тех пор было показано, что SL играют важную роль в старении листьев , реакции на фосфатное голодание, солеустойчивости и передаче световых сигналов. [57]
Другие известные гормоны
[ редактировать ]Другие идентифицированные регуляторы роста растений включают:
- Растительные пептидные гормоны – включают все небольшие секретируемые пептиды, которые участвуют в передаче сигналов от клетки к клетке. Эти небольшие пептидные гормоны играют решающую роль в росте и развитии растений, включая защитные механизмы, контроль деления и расширения клеток, а также самонесовместимость пыльцы. [58] Известно, что небольшой пептид CLE25 действует как сигнал на большие расстояния, передающий водный стресс, ощущаемый в корнях, к устьицам листьев. [59]
- Полиамины – сильноосновные молекулы с низкой молекулярной массой, обнаруженные во всех изученных на данный момент организмах. Они необходимы для роста и развития растений и влияют на процесс митоза и мейоза. У растений полиамины связаны с контролем старения. [60] и запрограммированная гибель клеток . [61]
- Оксид азота (NO) – служит сигналом для гормональных и защитных реакций (например, закрытия устьиц, развития корней, прорастания, фиксации азота, гибели клеток, реакции на стресс). [62] NO может продуцироваться с помощью еще не определенной NO-синтазы, особого типа нитритредуктазы, нитратредуктазы, митохондриальной цитохром-с-оксидазы или неферментативных процессов и регулировать функции органелл растительных клеток (например, синтез АТФ в хлоропластах и митохондриях). [63]
- Каррикины – не являются растительными гормонами, поскольку не производятся самими растениями, а содержатся в дыме горящего растительного сырья. Каррикины могут способствовать прорастанию семян многих видов. [64] Обнаружение того, что растения, у которых отсутствует рецептор каррикина, демонстрируют несколько фенотипов развития (повышенное накопление биомассы и повышенная чувствительность к засухе), привело некоторых к предположениям о существовании еще не идентифицированного каррикин-подобного эндогенного гормона у растений. Клеточный сигнальный путь каррикина имеет много общих компонентов с сигнальным путем стриголактона. [65]
- Триаконтанол – жирный спирт, который действует как стимулятор роста, особенно инициируя новые базальные разрывы у представителей семейства розовых . Он содержится в люцерне (люцерне), пчелином воске и некоторых восковых кутикулах листьев.
Использование в садоводстве
[ редактировать ]Синтетические растительные гормоны или PGR используются в ряде различных методов, включающих растений черенками размножение , прививку , микроразмножение и культуру тканей . Чаще всего они коммерчески доступны в виде «порошка гормона укоренения».
Размножение растений черенками полностью развитых листьев, стеблей или корней садоводы осуществляют, используя ауксин в качестве укореняющего вещества, наносимого на поверхность среза; ауксины попадают в растение и способствуют закладыванию корней. При прививке ауксин способствует образованию каллюсной ткани, которая соединяет поверхности трансплантата вместе . При микроразмножении используются различные регуляторы роста для стимулирования размножения и последующего укоренения новых всходов. При культивировании тканей растительных клеток PGR используются для обеспечения роста каллуса, размножения и укоренения.
можно применять растительные гормоны или смеси, в их состав При использовании в полевых условиях в качестве биостимуляторов . [66]
Покой семян
[ редактировать ]Растительные гормоны влияют на прорастание и покой семян, воздействуя на различные части семян.
Покой эмбриона характеризуется высоким соотношением АБК:ГК, тогда как семена имеют высокую чувствительность к абсцизовой кислоте и низкую чувствительность к ГК. Чтобы вывести семена из этого типа покоя и инициировать прорастание семян, должно произойти изменение биосинтеза гормонов и деградация в сторону низкого соотношения АБК/ГК, а также снижение чувствительности АБК и повышение чувствительности ГК.
АБК контролирует покой эмбриона и прорастание эмбрионов ГА.Покой семенной кожуры предполагает механическое ограничение семенной кожуры. Это, наряду с низким потенциалом роста зародыша, эффективно приводит к покою семян. ГК освобождает этот период покоя, увеличивая потенциал роста эмбриона и/или ослабляя семенную оболочку, чтобы радикал проростка мог прорваться через семенную оболочку.Различные типы семенных оболочек могут состоять из живых или мертвых клеток, и на оба типа могут влиять гормоны; на те, которые состоят из живых клеток, воздействуют после образования семян, тогда как на семенные кожуры, состоящие из мертвых клеток, могут влиять гормоны во время формирования семенной кожуры. АБК влияет на характеристики роста семенников или семенных оболочек, включая толщину, и влияет на потенциал роста эмбриона, опосредованный ГК. Эти условия и эффекты возникают во время формирования семян, часто в ответ на условия окружающей среды. Гормоны также опосредуют покой эндосперма: эндосперм большинства семян состоит из живой ткани, которая может активно реагировать на гормоны, вырабатываемые зародышем. Эндосперм часто действует как барьер для прорастания семян, играя роль в состоянии покоя семенной кожуры или в процессе прорастания. Живые клетки реагируют на соотношение АБК: ГК и влияют на него, а также опосредуют клеточную чувствительность; Таким образом, ГК увеличивает потенциал роста эмбриона и может способствовать ослаблению эндосперма. ГК также влияет как на АБК-независимые, так и на АБК-ингибирующие процессы в эндосперме. [67]
Использование человеком
[ редактировать ]Салициловая кислота
[ редактировать ]Кора ивы веками использовалась как болеутоляющее средство. Активным ингредиентом коры ивы, обеспечивающим эти эффекты, является гормон салициловая кислота (СК). В 1899 году фармацевтическая компания Bayer начала продавать производное SA под названием аспирин . [68] Помимо использования в качестве болеутоляющего средства, SA также используется при местном лечении некоторых кожных заболеваний, включая прыщи, бородавки и псориаз. [69] другое производное СК, салицилат натрия, подавляет пролиферацию раковых клеток лимфобластного лейкоза, простаты, молочной железы и меланомы человека. Было обнаружено, что [70]
Жасминовая кислота
[ редактировать ]Жасмоновая кислота (JA) может вызывать гибель клеток лимфобластного лейкоза . Было показано, что метилжасмонат (производное JA, также обнаруженное в растениях) ингибирует пролиферацию ряда линий раковых клеток. [70] хотя до сих пор ведутся споры по поводу его использования в качестве противоракового препарата из-за его потенциального негативного воздействия на здоровые клетки. [71]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Мендес-Эрнандес Х.А., Ледезма-Родригес М., Авилес-Монтальво Р.Н., Хуарес-Гомес Ю.Л., Скит А., Авилес-Монтальво Дж. и др. (2019). «Обзор сигнализации соматического эмбриогенеза растений» . Границы в науке о растениях . 10:77 . дои : 10.3389/fpls.2019.00077 . ПМК 6375091 . ПМИД 30792725 .
- ^ Сигенага А.М., Аргуесо Коннектикут (август 2016 г.). «Нет гормона, который мог бы управлять ими всеми: взаимодействие растительных гормонов во время реакции растений на патогены». Семинары по клеточной биологии и биологии развития . 56 : 174–189. дои : 10.1016/j.semcdb.2016.06.005 . ПМИД 27312082 .
- ^ Бюргер М., Чори Дж. (август 2019 г.). «Стресс из-за гормонов: как растения управляют иммунитетом» . Клетка-хозяин и микроб . 26 (2): 163–172. дои : 10.1016/j.chom.2019.07.006 . ПМЦ 7228804 . ПМИД 31415749 .
- ^ Ку Ю.С., Синтаха М., Чунг М.Ю., Лам Х.М. (октябрь 2018 г.). «Перекрестные помехи, передающие сигналы растительных гормонов, между реакциями на биотический и абиотический стресс» . Международный журнал молекулярных наук . 19 (10): 3206. doi : 10.3390/ijms19103206 . ПМК 6214094 . ПМИД 30336563 .
- ^ Улла А., Мангвар Х., Шабан М., Хан А.Х., Акбар А., Али У и др. (ноябрь 2018 г.). «Фитогормоны повышают устойчивость растений к засухе: стратегия борьбы». Международное исследование наук об окружающей среде и загрязнении окружающей среды . 25 (33): 33103–33118. Бибкод : 2018ESPR...2533103U . дои : 10.1007/s11356-018-3364-5 . ПМИД 30284160 . S2CID 52913388 .
- ^ Пьер-Жером Э., Драпек С., Бенфей П.Н. (октябрь 2018 г.). «Регуляция деления и дифференцировки стволовых клеток растений» . Ежегодный обзор клеточной биологии и биологии развития . 34 : 289–310. doi : 10.1146/annurev-cellbio-100617-062459 . ПМК 6556207 . ПМИД 30134119 .
- ^ «Растительные гормоны» . НКС Пирсон. Архивировано из оригинала 27 ноября 2021 г. Проверено 14 августа 2018 г.
- ^ «Растительные гормоны» . Архивировано из оригинала 18 декабря 2019 г. Проверено 14 августа 2018 г.
- ^ Пошел FW, Тиманн КВ (1937). Фитогормоны . Нью-Йорк: Компания Macmillan.
- ^ Тараховская Э.Р., Маслов Ю., Шишова М.Ф. (2007). «Фитогормоны в водорослях». Российский журнал физиологии растений . 54 (2): 163–170. дои : 10.1134/s1021443707020021 . S2CID 27373543 .
- ^ Радемахер В. (1994). «Образование гиббереллина у микроорганизмов». Регулирование роста растений . 15 (3): 303–314. дои : 10.1007/BF00029903 . S2CID 33138732 .
- ^ Шривастава Л.М. (2002). Рост и развитие растений: гормоны и окружающая среда . Академическая пресса. п. 140. ИСБН 978-0-12-660570-9 .
- ^ Эпик Х., Рольф С.А., Уиллис Дж.А., Street HE (2005). Физиология цветковых растений (4-е изд.). Издательство Кембриджского университета. п. 191. ИСБН 978-0-521-66251-2 . [ постоянная мертвая ссылка ]
- ^ Сваруп Р., Перри П., Хагенбек Д., Ван Дер Страетен Д., Бимстер Г.Т., Сандберг Г. и др. (июль 2007 г.). «Этилен усиливает биосинтез ауксина в проростках арабидопсиса, усиливая ингибирование удлинения корневых клеток» . Растительная клетка . 19 (7): 2186–96. дои : 10.1105/tpc.107.052100 . ЧВК 1955695 . ПМИД 17630275 .
- ^ Шривастава 2002 , с. 143
- ^ Ботаника: краткое введение в биологию растений . Нью-Йорк: Уайли. 1979. стр. 155–170 . ISBN 978-0-471-02114-8 .
- ^ Феуртадо Дж.А., Эмброуз С.Дж., Катлер А.Дж., Росс А.Р., Абрамс С.Р., Кермод А.Р. (февраль 2004 г.). «Прекращение покоя семян сосны западной белой (Pinus monticola Dougl. Ex D. Don) связано с изменениями в метаболизме абсцизовой кислоты». Планта . 218 (4): 630–9. Бибкод : 2004Plant.218..630F . дои : 10.1007/s00425-003-1139-8 . ПМИД 14663585 . S2CID 25035678 .
- ^ Кермод, А.Р. (декабрь 2005 г.). «Роль абсцизовой кислоты в состоянии покоя семян» . J Регулирование роста растений . 24 (4): 319–344. дои : 10.1007/s00344-005-0110-2 .
- ^ Рен Х., Гао З., Чен Л., Вэй К., Лю Дж., Фань Ю. и др. (2007). «Динамический анализ накопления АБК в зависимости от скорости катаболизма АБК в тканях кукурузы при дефиците воды» . Журнал экспериментальной ботаники . 58 (2): 211–9. дои : 10.1093/jxb/erl117 . ПМИД 16982652 .
- ^ Эльс М.А., Коупленд Д., Даттон Л., Джексон М.Б. (январь 2001 г.). «Снижение гидравлической проводимости корней снижает потенциал воды в листьях, инициирует закрытие устьиц и замедляет расширение листьев у растений, залитых касторовым маслом ( Ricinus communis ), несмотря на уменьшение доставки АБК от корней к побегам в соке ксилемы». Физиология Плантарум . 111 (1): 46–54. дои : 10.1034/j.1399-3054.2001.1110107.x .
- ^ Ян Дж., Цуичихара Н., Это Т., Иваи С. (октябрь 2007 г.). «Активные формы кислорода и оксид азота участвуют в ингибировании АБК открытия устьиц». Растение, клетка и окружающая среда . 30 (10): 1320–5. дои : 10.1111/j.1365-3040.2007.01711.x . ПМИД 17727421 .
- ^ Осборн диджей , Макманус М.Т. (2005). Гормоны, сигналы и клетки-мишени в развитии растений . Издательство Кембриджского университета. п. 158 . ISBN 978-0-521-33076-3 .
- ^ Томич С., Габдуллин Р.Р., Койич-Продич Б., Уэйд Р.К. (1998). «Классификация соединений, родственных ауксину, на основе сходства полей их взаимодействия: распространение на новый набор соединений». Интернет-журнал химии . 1 (26): ЦП1–У21.
- ^ Уолц А., Парк С., Словин Дж. П., Людвиг-Мюллер Дж., Момоноки Ю. С., Коэн Дж. Д. (февраль 2002 г.). «Ген, кодирующий белок, модифицированный фитогормоном индолуксусной кислотой» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1718–23. Бибкод : 2002PNAS...99.1718W . дои : 10.1073/pnas.032450399 . ПМЦ 122257 . ПМИД 11830675 .
- ^ Гроув М.Д., Спенсер Г.Ф., Роведдер В.К., Мандава Н., Уорли Дж.Ф., Вартен Дж.Д. и др. (1979). «Брассинолид, стероид, способствующий росту растений, выделенный из пыльцы Brassica napus». Природа . 281 (5728): 216–217. Бибкод : 1979Natur.281..216G . дои : 10.1038/281216a0 . S2CID 4335601 .
- ^ Перейти обратно: а б Тан Дж, Хан З, Чай Дж (декабрь 2016 г.). «Вопросы и ответы: что такое брассиностероиды и как они действуют на растения?» . БМК Биология . 14 (1): 113. дои : 10.1186/s12915-016-0340-8 . ПМК 5180403 . ПМИД 28007032 .
- ^ Ямагами А., Сайто С., Наказава М., Фудзиока С., Уемура Т., Мацуи М. и др. (июль 2017 г.). «Эволюционно консервативный BIL4 подавляет деградацию брассиностероидного рецептора BRI1 и регулирует удлинение клеток» . Научные отчеты . 7 (1): 5739. Бибкод : 2017НацСР...7.5739Y . дои : 10.1038/s41598-017-06016-2 . ПМК 5515986 . ПМИД 28720789 .
- ^ Ямагами А., Наказава М., Мацуи М., Туджимото М., Сакута М., Асами Т., Накано Т. (февраль 2009 г.). «Химическая генетика выявила новый трансмембранный белок BIL4, который опосредует удлинение растительных клеток при передаче сигналов брассиностероидов» . Бионауки, биотехнологии и биохимия . 73 (2): 415–421. дои : 10.1271/bbb.80752 . ПМИД 19202280 . S2CID 35568960 .
- ^ Перейти обратно: а б с Планас-Риверола А, Гупта А, Бетегон-Путце I, Бош Н, Ибаньес М, Каньо-Дельгадо А.И. (март 2019 г.). «Передача сигналов брассиностероидов в развитии растений и адаптации к стрессу» . Разработка . 146 (5): dev151894. дои : 10.1242/dev.151894 . ПМЦ 6432667 . ПМИД 30872266 .
- ^ Ахаммед Г.Дж., Ли Х, Лю А, Чен С. (19 марта 2020 г.). «Брассиностероиды в устойчивости растений к абиотическому стрессу» . Журнал регулирования роста растений . 39 (4): 1451–1464. дои : 10.1007/s00344-020-10098-0 . ISSN 0721-7595 . S2CID 213166792 .
- ^ Сайпс Д.Л., Эйнсет Дж.В. (август 1983 г.). «Цитокининовая стимуляция опадения эксплантатов пестика лимона». J Регулирование роста растений . 2 (1–3): 73–80. дои : 10.1007/BF02042235 . S2CID 43997977 .
- ^ Ахтар СС, Мекурея М.Ф., Панди С., Ройч Т. (2020). «Роль цитокининов во взаимодействии растений с микробными возбудителями и насекомыми-вредителями» . Границы в науке о растениях . 10 : 1777. doi : 10.3389/fpls.2019.01777 . ПМК 7042306 . ПМИД 32140160 .
- ^ «Цитокинин - обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 10 июня 2021 г.
- ^ Ван Ю, Лю С, Ли К, Сунь Ф, Ху Х, Ли Х и др. (август 2007 г.). «EIN2 арабидопсиса модулирует реакцию на стресс через путь реакции абсцизовой кислоты». Молекулярная биология растений . 64 (6): 633–44. дои : 10.1007/s11103-007-9182-7 . ПМИД 17533512 . S2CID 42139177 .
- ^ Джексон МБ (1985). «Этилен и реакция растений на переувлажнение и затопление почвы». Ежегодный обзор физиологии растений . 36 (1): 145–174. дои : 10.1146/annurev.pp.36.060185.001045 . ISSN 0066-4294 .
- ^ Джексон МБ (январь 2008 г.). «Удлинение, стимулируемое этиленом: адаптация к стрессу при погружении» . Анналы ботаники . 101 (2): 229–48. дои : 10.1093/aob/mcm237 . ПМК 2711016 . ПМИД 17956854 .
- ^ Джексон МБ, Рам ПК (январь 2003 г.). «Физиологические и молекулярные основы восприимчивости и устойчивости растений риса к полному затоплению» . Анналы ботаники . 91 Спецификация № (2): 227–41. дои : 10.1093/aob/mcf242 . ПМЦ 4244997 . ПМИД 12509343 .
- ^ Воесенек Л.А., Беншоп Дж.Дж., Боу Дж., Кокс М.К., Груневельд Х.В., Милленаар Ф.Ф. и др. (январь 2003 г.). «Взаимодействие между растительными гормонами регулирует удлинение побегов, вызванное погружением в воду, у устойчивых к наводнениям двудольных растений Rumex palustris» . Анналы ботаники . 91 Спецификация № (2): 205–11. дои : 10.1093/aob/mcf116 . ПМК 4244986 . ПМИД 12509341 .
- ^ Саммерс Дж. Э., Воесенек Л., Блом С., Льюис М. Дж., Джексон М. Б. (июль 1996 г.). «Potamogeton pectinatus принципиально не способен синтезировать этилен и не имеет оксидазы 1-аминоциклопропан-1-карбоновой кислоты» . Физиология растений . 111 (3): 901–908. дои : 10.1104/стр.111.3.901 . ПМК 157909 . ПМИД 12226336 .
- ^ Греннан АК (июнь 2006 г.). «Ферменты метаболизма гиббереллина в рисе» . Физиология растений . 141 (2): 524–6. дои : 10.1104/стр.104.900192 . ПМЦ 1475483 . ПМИД 16760495 .
- ^ Цай Ф.Ю., Линь К.С., Као Ч. (январь 1997 г.). «Сравнительное исследование влияния абсцизовой кислоты и метилжасмоната на рост рассады риса». Регулирование роста растений . 21 (1): 37–42. дои : 10.1023/А:1005761804191 . S2CID 34099546 .
- ^ Демоле Э., Ледерер Э., Мерсье Д. (1962). «Выделение и определение структуры метилжасмоната — ароматной составляющей, характерной для жасминового масла». Helvetica Chimica Acta (на французском языке). 45 (2): 675–685. дои : 10.1002/hlca.19620450233 .
- ^ Перейти обратно: а б Просмотрите J (2005). «Жасмонат: сигнал оксилипина, играющий множество ролей в растениях». Растительные гормоны . Витамины и гормоны. Том. 72. стр. 431–56. дои : 10.1016/S0083-6729(05)72012-4 . ISBN 9780127098722 . ПМИД 16492478 .
- ^ Кацир Л., Чунг Х.С., Ку А.Дж., Хоу Г.А. (август 2008 г.). «Передача сигналов жасмоната: консервативный механизм восприятия гормонов» . Современное мнение в области биологии растений . 11 (4): 428–35. дои : 10.1016/j.pbi.2008.05.004 . ПМК 2560989 . ПМИД 18583180 .
- ^ Лоренцо О., Солано Р. (октябрь 2005 г.). «Молекулярные игроки, регулирующие сигнальную сеть жасмоната». Современное мнение в области биологии растений . 8 (5): 532–40. дои : 10.1016/j.pbi.2005.07.003 . ПМИД 16039901 .
- ^ Вастернак С. (октябрь 2007 г.). «Жасмонаты: обновленная информация о биосинтезе, передаче сигналов и действии при реакции растений на стресс, их росте и развитии» . Анналы ботаники . 100 (4): 681–97. дои : 10.1093/aob/mcm079 . ПМЦ 2749622 . ПМИД 17513307 .
- ^ Перейти обратно: а б Хау Г.А., Джандер Дж. (2008). «Иммунитет растений к насекомым-травоядным». Анну. Преподобный Плант Биол . 59 : 41–66. doi : 10.1146/annurev.arplant.59.032607.092825 . ПМИД 18031220 .
- ^ Пашольд А., Халичке Р., Болдуин И.Т. (июль 2007 г.). «Ко(i)-координирующая защита: NaCOI1 опосредует устойчивость, индуцированную травоядными животными, у Nicotiana attenuata и раскрывает роль движения травоядных животных в избегании защиты» . Журнал растений: для клеточной и молекулярной биологии . 51 (1): 79–91. дои : 10.1111/j.1365-313X.2007.03119.x . ПМИД 17561925 .
- ^ Сарате С.И., Кеммпема Л.А., Уоллинг Л.Л. (февраль 2007 г.). «Белокрылка серебристая индуцирует защиту салициловой кислоты и подавляет эффективную защиту жасмоновой кислоты» . Физиология растений . 143 (2): 866–75. дои : 10.1104/стр.106.090035 . ПМК 1803729 . ПМИД 17189328 .
- ^ Ривас-Сан-Висенте М., Пласенсиа Дж. (июнь 2011 г.). «Салициловая кислота беззащитна: ее роль в росте и развитии растений» . Журнал экспериментальной ботаники . 62 (10): 3321–38. дои : 10.1093/jxb/err031 . ПМИД 21357767 .
- ^ Демпси Д.А., Клессиг Д.Ф. (март 2017 г.). «Как многогранный растительный гормон салициловая кислота борется с болезнями растений и используются ли аналогичные механизмы у людей?» . БМК Биология . 15 (1): 23. дои : 10.1186/s12915-017-0364-8 . ПМЦ 5364617 . ПМИД 28335774 .
- ^ Кумар Д. (ноябрь 2014 г.). «Передача сигналов салициловой кислоты в устойчивости к болезням». Наука о растениях . 228 : 127–34. doi : 10.1016/j.plantsci.2014.04.014 . ПМИД 25438793 .
- ^ Дин П, Дин Ю (июнь 2020 г.). «Истории о салициловой кислоте: защитном гормоне растений». Тенденции в науке о растениях . 25 (6): 549–565. doi : 10.1016/j.tplants.2020.01.004 . ПМИД 32407695 . S2CID 213666131 .
- ^ Се Икс, Ёнеяма К, Ёнеяма К (01 июля 2010 г.). «История стриголактона». Ежегодный обзор фитопатологии . 48 (1): 93–117. doi : 10.1146/annurev-phyto-073009-114453 . ПМИД 20687831 . S2CID 27305711 .
- ^ Акияма К., Мацузаки К., Хаяси Х. (июнь 2005 г.). «Растительные сесквитерпены вызывают ветвление гиф у арбускулярных микоризных грибов». Природа . 435 (7043): 824–7. Бибкод : 2005Natur.435..824A . дои : 10.1038/nature03608 . ПМИД 15944706 . S2CID 4343708 .
- ^ Гомес-Ролдан В., Фермас С., Брюэр П.Б., Пуэх-Пажес В., Дун Э.А., Пиллот Дж.П. и др. (сентябрь 2008 г.). «Ингибирование стриголактоном ветвления побегов». Природы . 455 (7210): 189–94. Бибкод : 2008Natur.455..189G . дои : 10.1038/nature07271 . ПМИД 18690209 . S2CID 205214618 .
- ^ Саид В., Насим С., Али З. (28 августа 2017 г.). «Биосинтез стриголактонов и их роль в устойчивости растений к абиотическому стрессу: критический обзор» . Границы в науке о растениях . 8 : 1487. doi : 10.3389/fpls.2017.01487 . ПМК 5581504 . ПМИД 28894457 .
- ^ Линдси К., Кассон С., Чилли П. (февраль 2002 г.). «Пептиды: новые сигнальные молекулы в растениях» . Тенденции в науке о растениях . 7 (2): 78–83. дои : 10.1016/S0960-9822(01)00435-3 . ПМИД 11832279 . S2CID 5064533 .
- ^ Такахаши Ф., Сузуки Т., Осакабе Ю., Бецуяку С., Кондо Ю., Домаэ Н. и др. (апрель 2018 г.). «Небольшой пептид модулирует устьичный контроль посредством абсцизовой кислоты при передаче сигналов на большие расстояния». Природа . 556 (7700): 235–238. Бибкод : 2018Natur.556..235T . дои : 10.1038/s41586-018-0009-2 . ПМИД 29618812 . S2CID 4598494 .
- ^ Панди С., Ранаде С.А., Нагар П.К., Кумар Н. (сентябрь 2000 г.). «Роль полиаминов и этилена как модуляторов старения растений». Журнал биологических наук . 25 (3): 291–9. дои : 10.1007/BF02703938 . ПМИД 11022232 . S2CID 21925829 .
- ^ Мошу П.Н., Рубелакис-Ангелакис К.А. (март 2014 г.). «Полиамины и запрограммированная смерть клеток» . Журнал экспериментальной ботаники . 65 (5): 1285–96. дои : 10.1093/jxb/ert373 . ПМИД 24218329 .
- ^ Шапиро А.Д. (2005). «Передача сигналов оксида азота в растениях». Растительные гормоны . Витамины и гормоны. Том. 72. С. 339–98. дои : 10.1016/S0083-6729(05)72010-0 . ISBN 9780127098722 . ПМИД 16492476 . S2CID 13308295 .
- ^ Росзер Т (2012). «Синтез оксида азота в хлоропластах». В Roszer T (ред.). Биология субклеточного оксида азота . Нью-Йорк, Лондон, Гейдельберг: Springer. ISBN 978-94-007-2818-9 .
- ^ Чивоча С.Д., Диксон К.В., Флематти Г.Р., Гисалберти Э.Л., Мерритт Д.Д., Нельсон Д.С. и др. (01.10.2009). «Каррикины: новое семейство дымовых регуляторов роста растений». Наука о растениях . 177 (4): 252–256. doi : 10.1016/j.plantsci.2009.06.007 .
- ^ Ли В., Нгуен К.Х., Чу Х.Д., Ха К.В., Ватанабе Й., Осакабе Й. и др. (ноябрь 2017 г.). «Каррикиновый рецептор KAI2 способствует устойчивости Arabidopsis thaliana к засухе» . ПЛОС Генетика . 13 (11): e1007076. дои : 10.1371/journal.pgen.1007076 . ПМК 5703579 . ПМИД 29131815 .
- ^ Яхин О.И., Лубянов А.А., Яхин И.А., Браун П.Х. (26 января 2017 г.). «Биостимуляторы в науке о растениях: глобальная перспектива» . Границы в науке о растениях . 7 : 2049. doi : 10.3389/fpls.2016.02049 . ПМЦ 5266735 . ПМИД 28184225 .
- ^ Лейбнер Г. (2000). «Покой семян» . Место биологии семян . Лондонский Королевский университет Холлоуэй.
- ^ Диармуид Дж. (декабрь 2008 г.). Аспирин: замечательная история чудо-лекарства . Нью-Йорк, штат Нью-Йорк. ISBN 9781596918160 . OCLC 879610692 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Мадан Р.К., Левитт Дж. (апрель 2014 г.). «Обзор токсичности препаратов салициловой кислоты для местного применения». Журнал Американской академии дерматологии . 70 (4): 788–792. дои : 10.1016/j.jaad.2013.12.005 . ПМИД 24472429 .
- ^ Перейти обратно: а б Финрут О, Флешер Э (апрель 2002 г.). «Гормоны стресса растений подавляют пролиферацию и вызывают апоптоз раковых клеток человека» . Лейкемия . 16 (4): 608–16. дои : 10.1038/sj.leu.2402419 . ПМИД 11960340 .
- ^ Чжан М., Чжан М.В., Чжан Л., Чжан Л. (24 июля 2015 г.). «Метилжасмонат и его потенциал в терапии рака» . Сигнализация и поведение растений . 10 (9): e1062199. Бибкод : 2015PlSiB..10E2199Z . дои : 10.1080/15592324.2015.1062199 . ПМЦ 4883903 . ПМИД 26208889 .
Внешние ссылки
[ редактировать ]- Простая таблица растительных гормонов с указанием места синтеза и эффектов применения — именно такой формат используется в шаблонах описаний внизу статей Википедии о растительных гормонах.
- Гормональная регуляция экспрессии и развития генов . Подробное введение в растительные гормоны, включая генетическую информацию.