Жасмонат

Жасмонат ( JA на липидной основе ) и его производные представляют собой растительные гормоны , которые регулируют широкий спектр процессов в растениях, от роста и фотосинтеза до репродуктивного развития. В частности, JA имеют решающее значение для защиты растений от травоядных и реакции растений на плохие условия окружающей среды и другие виды абиотических и биотических проблем. [1] Некоторые JA также могут выделяться в виде летучих органических соединений (ЛОС), чтобы обеспечить связь между растениями в ожидании взаимной опасности. [2]
История
[ редактировать ]Выделение метилжасмоната (MeJA) из жасминового масла, полученного из Jasminum grandiflorum, привело к открытию молекулярной структуры жасмонатов и их названия в 1962 году. [3] [4] в то время как сама жасмоновая кислота была выделена из Lasiodiplodia theobromae Олдериджем и др. в 1971 году. [4]
Биосинтез
[ редактировать ]

Биосинтез рассмотрен Акостой и Фармером, 2010 г., Вастернаком и Хаусом, 2013 г. и Вастернаком и Сонгом, 2017 г. [4] Жасмонаты (ЖА) представляют собой оксилипины , т.е. производные оксигенированных жирных кислот. Они биосинтезируются из линоленовой кислоты в мембранах хлоропластов. Синтез начинается с превращения линоленовой кислоты в 12-оксо-фитодиеновую кислоту (OPDA), которая затем подвергается восстановлению и трем циклам окисления с образованием (+)-7-изо-JA, жасмоновой кислоты. происходит только превращение линоленовой кислоты в ОПДА В хлоропластах ; все последующие реакции происходят в пероксисоме . [5]
Сам JA может далее метаболизироваться в активные или неактивные производные. Метил JA (MeJA) — летучее соединение, которое потенциально отвечает за межрастительные коммуникации . JA, конъюгированный с аминокислотой изолейцином (Ile), приводит к образованию JA-Ile ((+)-7-изо-жасмоноил- L -изолейцина), который, как установил Фонсека и др., 2009, участвует в большинстве передач сигналов JA. [6] - см. также обзор Кацира и др., 2008 г. [5] Однако Van Poecke & Dicke 2003 считает, что не Arabidopsis выбросы летучих веществ требуют ни JA-Ile, ни VanDoorn et al 2011 для Solanum nigrum травоядным животным к устойчивости . [6] JA подвергается декарбоксилированию с образованием цис-жасмона .
Функция
[ редактировать ]Хотя жасмонат (JA) регулирует множество различных процессов в растении, его роль в реакции на рану понятна лучше всего. После механического ранения или травоядности биосинтез JA быстро активируется, что приводит к экспрессии соответствующих генов ответа. Например, у томата ранение производит защитные молекулы, которые подавляют переваривание листьев в кишках насекомых . Другим косвенным результатом передачи сигналов JA является летучая эмиссия соединений, полученных из JA. MeJA на листьях может перемещаться по воздуху к близлежащим растениям и повышать уровень транскриптов, связанных с реакцией на рану. [1] В целом, это излучение может дополнительно активировать биосинтез JA и передачу сигналов клетками , тем самым побуждая близлежащие растения активировать свою защиту в случае травоядности.
JA также участвуют в гибели клеток и старении листьев. JA может взаимодействовать со многими киназами и факторами транскрипции, связанными со старением. JA также может вызывать гибель митохондрий, вызывая накопление активных форм кислорода (АФК). Эти соединения разрушают мембраны митохондрий и ставят под угрозу клетку, вызывая апоптоз или запрограммированную гибель клеток. Роль JA в этих процессах наводит на мысль о методах, с помощью которых растение защищает себя от биотических проблем и ограничивает распространение инфекций . [7]
JA и его производные также участвуют в развитии растений, симбиозе и множестве других процессов, включенных в список ниже.
- Изучая мутантов, сверхэкспрессирующих JA, одним из самых ранних открытий было то, что JA ингибирует рост корней. Механизм этого события до сих пор не ясен, но мутанты COI1-зависимого сигнального пути имеют тенденцию проявлять пониженное ингибирование, демонстрируя, что путь COI1 каким-то образом необходим для ингибирования роста корней. [8] [9]
- JA играет множество ролей в развитии цветка . Мутанты по синтезу JA или передаче сигналов JA у Arabidopsis обладают мужской стерильностью, обычно из-за задержки развития. Те же гены, которые способствуют мужской фертильности у арабидопсиса, способствуют женской фертильности у томатов. Сверхэкспрессия 12-OH-JA также может задерживать цветение. [9]
- JA и MeJA подавляют прорастание неспящих семян и стимулируют прорастание спящих семян. [10]
- Высокие уровни JA способствуют накоплению запасных белков; гены, кодирующие вегетативные запасные белки, чувствительны к JA. В частности, тубероновая кислота, производное ЖК, индуцирует образование клубней. [11] [12]
- ЖК также играют роль в симбиозе растений и микроорганизмов; однако его точная роль до сих пор неясна. В настоящее время JA, по-видимому, регулирует обмен сигналами и регуляцию клубеньков между бобовыми и ризобиями. С другой стороны, повышенные уровни JA, по-видимому, регулируют распределение углеводов и устойчивость к стрессу у микоризных растений. [13]

- JA были вовлечены в развитие плотоядных растений, таких как венеринская мухоловка . Исследования показывают, что эволюционное перепрофилирование сигнального пути жасмоната, который обеспечивает защиту от травоядных животных у неплотоядных растений, поддержало эволюцию плотоядных растений. Жасмонаты можно использовать для подачи сигнала о закрытии ловушек и для контроля высвобождения ферментов и переносчиков питательных веществ, которые используются в пищеварении растений. Однако не все плотоядные растения одинаково зависят от жасмонатного пути. Маслянки существенно отличаются от венериных мухоловок и росянки и, возможно, имеют развитые методы регуляции пищеварительных ферментов, независимые от JA. [14] [15]
Роль в патогенезе
[ редактировать ]Pseudomonas syringae вызывает бактериальную пятнистость томатов, захватывая сигнальный путь жасмоната (JA) растения. Эти бактерии используют систему секреции типа III для инъекции смеси вирусных эффекторных белков в клетки-хозяева.
Одной из молекул, входящих в эту смесь, является фитотоксин коронатин (COR). Растения, нечувствительные к JA, обладают высокой устойчивостью к P. syringae и не реагируют на COR; кроме того, применения MeJA было достаточно для восстановления вирулентности мутантных бактерий COR. Зараженные растения также экспрессировали нижестоящие гены JA и реакции на рану, но подавляли уровни генов, связанных с патогенезом (PR). Все эти данные позволяют предположить, что COR действует через путь JA, проникая в растения-хозяева. Предполагается, что активация раневой реакции происходит за счет защиты патогена. Активируя путь ответа на рану JA, P. syringae может отвлекать ресурсы иммунной системы хозяина и более эффективно заражать. [16]
Растения производят N-ациламиды , которые придают устойчивость к некротрофным патогенам путем активации биосинтеза и передачи сигналов JA. Арахидоновая кислота (АК), аналог предшественника ЖК α-LeA, встречающаяся у многоклеточных видов, но не у растений, воспринимается растениями и действует посредством повышения уровня ЖК одновременно с устойчивостью к некротрофным патогенам. АК представляет собой эволюционно консервативную сигнальную молекулу , которая действует у растений в ответ на стресс аналогично тому, как это происходит в животных системах. [17]
Перекрестный разговор с другими путями защиты
[ редактировать ]Хотя жасмонатный (JA) путь имеет решающее значение для реакции на рану, это не единственный сигнальный путь, опосредующий защиту у растений. Чтобы построить оптимальную, но эффективную защиту, различные пути защиты должны быть способны к перекрестному взаимодействию для точной настройки и определения ответов на абиотические и биотические проблемы.
Один из наиболее изученных примеров перекрестных помех JA возникает при использовании салициловой кислоты (SA). SA, гормон, опосредует защиту от патогенов, индуцируя как экспрессию генов, связанных с патогенезом, так и системную приобретенную устойчивость (SAR), при которой все растение приобретает устойчивость к патогену после локализованного воздействия на него.
Реакция раны и патогена, по-видимому, взаимодействует отрицательно. Например, подавление фенилаланин-аммиаклиазы (PAL), фермента, синтезирующего предшественники SA, снижает SAR, но повышает устойчивость травоядных животных к насекомым. Аналогичным образом, сверхэкспрессия PAL усиливает SAR, но снижает реакцию на рану после травоядных насекомых. [18] В целом было обнаружено, что патогены, живущие в живых растительных клетках, более чувствительны к защите, индуцированной СК, в то время как травоядные насекомые и патогены, получающие пользу от гибели клеток, более восприимчивы к защите ЮК. Таким образом, этот компромисс в путях оптимизирует защиту и экономит растительные ресурсы. [19]
Перекрестные помехи также происходят между JA и другими путями растительных гормонов, такими как абсцизовая кислота (АБК) и этилен (ET). Эти взаимодействия аналогичным образом оптимизируют защиту от патогенов и травоядных животных, ведущих разный образ жизни. Например, активность MYC2 может стимулироваться как путями JA, так и АБК, что позволяет интегрировать сигналы обоих путей. Другие факторы транскрипции, такие как ERF1, возникают в результате передачи сигналов JA и ET. Все эти молекулы могут действовать в сочетании, активируя определенные гены реакции на рану. [19]
Наконец, перекрестные помехи не ограничиваются защитой: взаимодействия JA и ET также имеют решающее значение для развития, и баланс между двумя соединениями необходим для правильного развития апикальных крючков у проростков Arabidopsis . Тем не менее, необходимы дальнейшие исследования для выяснения молекул, регулирующих такие перекрестные помехи. [18]
Механизм сигнализации
[ редактировать ]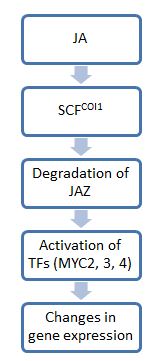
В целом этапы передачи сигналов жасмоната (JA) отражают этапы передачи сигналов ауксина : первый этап включает комплексы убиквитинлигазы E3, которые метят субстраты убиквитином, чтобы пометить их для деградации протеасомами . На втором этапе используются факторы транскрипции для воздействия на физиологические изменения. Одной из ключевых молекул на этом пути является JAZ, которая служит выключателем для передачи сигналов JA. В отсутствие JA белки JAZ связываются с нижестоящими факторами транскрипции и ограничивают их активность. Однако в присутствии JA или его биоактивных производных белки JAZ разрушаются, высвобождая факторы транскрипции для экспрессии генов, необходимых для реакции на стресс . [20]
Поскольку JAZ не исчезал на фоне растений с нулевой мутацией coi1 , было показано, что белок COI1 опосредует деградацию JAZ. COI1 принадлежит к семейству высококонсервативных белков F-box и рекрутирует субстраты для убиквитинлигазы E3 SCF. ИСП1 . Комплексы, которые в конечном итоге образуются, известны как комплексы SCF . [21] Эти комплексы связывают JAZ и направляют его на протеосомную деградацию. Однако, учитывая большой спектр молекул JA, не все производные JA активируют этот путь передачи сигнала, и круг тех, кто участвует в этом пути, неизвестен. [5] На данный момент показано, что только JA-Ile необходим для COI1-опосредованной деградации JAZ11. JA-Ile и структурно родственные производные могут связываться с комплексами COI1-JAZ и способствовать убиквитинированию и, следовательно, деградации последних. [5]
Эта механистическая модель повышает вероятность того, что COI1 служит внутриклеточным рецептором сигналов JA. Недавние исследования подтвердили эту гипотезу, продемонстрировав, что комплекс COI1-JAZ действует как корецептор восприятия JA. В частности, JA-Ile связывается как с лиганд-связывающим карманом в COI1, так и с участком из 20 аминокислот консервативного мотива Jas в JAZ. Этот остаток JAZ действует как заглушка кармана в COI1, удерживая JA-Ile связанным в кармане. Кроме того, Шеард и др., 2010 г. [22] совместная очистка и последующее удаление инозитолпентакисфосфата (InsP 5 ) из COI1, продемонстрировав, что InsP 5 является необходимым компонентом корецептора и играет роль в усилении корецепторного комплекса. Результаты Шеарда могут демонстрировать различную специфичность связывания для различных SCF. ИСП1 -ИнсП 5 -Комплексы ДЖАЗ. [6]
После освобождения от JAZ факторы транскрипции могут активировать гены, необходимые для специфического ответа JA. Наиболее изученные транскрипционные факторы, действующие на этом пути, относятся к семейству транскрипционных факторов MYC, которые характеризуются базовым мотивом связывания ДНК спираль-петля-спираль (bHLH). Эти факторы (их три: MYC2, 3 и 4) имеют тенденцию действовать аддитивно. Например, растение, потерявшее только один мик, становится более восприимчивым к травоядным насекомым, чем обычное растение. Растение, потерявшее все три, будет так же восприимчиво к повреждению, как и мутанты coi1, которые совершенно не реагируют на JA и не могут защититься от травоядных. Однако, хотя все эти молекулы MYC имеют общие функции, они сильно различаются по характеру экспрессии и функциям транскрипции. Например, MYC2 оказывает большее влияние на рост корней по сравнению с MYC3 или MYC4. [8]
Кроме того, MYC2 будет возвращаться вспять и регулировать уровни экспрессии JAZ, что приводит к возникновению петли отрицательной обратной связи . [8] Все эти факторы транскрипции по-разному влияют на уровни JAZ после передачи сигналов JA. Уровни JAZ, в свою очередь, влияют на уровень транскрипционных факторов и экспрессии генов. Другими словами, помимо активации различных генов ответа, факторы транскрипции могут варьировать уровни JAZ для достижения специфичности в ответ на сигналы JA.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Фермер, EE; Райан, Калифорния (1990). «Межрастительная коммуникация: переносимый по воздуху метилжасмонат индуцирует синтез ингибиторов протеиназ в листьях растений» . Proc Natl Acad Sci США . 87 (19): 7713–7716. Бибкод : 1990PNAS...87.7713F . дои : 10.1073/pnas.87.19.7713 . ПМК 54818 . ПМИД 11607107 .
- ^ Болдуин, IT; Халичке, Р.; Пашольд, А.; фон Даль, CC; Престон, Калифорния (2006). «Изменчивая передача сигналов во взаимодействиях растения-растения: «говорящие деревья» в эпоху геномики». Наука . 311 (5762): 812–815. Бибкод : 2006Sci...311..812B . дои : 10.1126/science.1118446 . ПМИД 16469918 . S2CID 9260593 .
- ^ Демоле Е; Ледерер, Э.; Мерсье, Д. (1962). «Выделение и определение структуры метилжасмоната, характерного пахнущего компонента жасминового масла». Хелв Чим Акта . 45 (2): 675–85. дои : 10.1002/hlca.19620450233 .
- ^ Перейти обратно: а б с Пер, Тасир С.; Хан, М. Икбал Р.; Анджум, Насер А.; Масуд, Асим; Хусейн, Софи Джавед; Хан, Нафис А. (2018). «Жасмонаты в растениях в условиях абиотического стресса: влияние перекрестных помех с другими фитогормонами». Экологическая и экспериментальная ботаника . 145 . Эльзевир : 104–120. дои : 10.1016/j.envexpbot.2017.11.004 . ISSN 0098-8472 .
- ^ Перейти обратно: а б с д Кацир, Л.; Чанг, HS; Ку, AJK; Хау, Джорджия (2008). «Передача сигналов жасмоната: консервативный механизм восприятия гормонов» . Современное мнение в области биологии растений . 11 (4). Эльзевир : 428–435. дои : 10.1016/j.pbi.2008.05.004 . ПМК 2560989 . ПМИД 18583180 .
- ^ Перейти обратно: а б с Шуман, Мередит К.; Болдуин, Ян Т. (11 марта 2016 г.). «Слои реакции растений на насекомых-травоядных» . Ежегодный обзор энтомологии . 61 (1). Годовые обзоры : 373–394. doi : 10.1146/annurev-ento-010715-023851 . ISSN 0066-4170 . ПМИД 26651543 . S2CID 24720368 .
- ^ Рейнбот, К; Спрингер, А; Самол, я; Рейнбот, С. (сентябрь 2009 г.). «Растительные оксилипины: роль жасмоновой кислоты в процессе запрограммированной гибели клеток, защиты и старения листьев» . Журнал ФЭБС . 276 (17): 4666–81. дои : 10.1111/j.1742-4658.2009.07193.x . ПМИД 19663906 . S2CID 1349010 .
- ^ Перейти обратно: а б с Фернандес-Кальво, П.; Чини, А.; Фернандес-Барберо, Г.; Чико, Ж.-М.; Хименес-Ибанес, С.; Геринк, Дж.; Икхаут, Д.; Швейцер, Ф.; Годой, М.; Франко-Соррилья, Ж.М.; Пауэлс, Л.; Уиттерс, Э.; Пуга, Мичиган; Пас-Арес, Дж.; Гуссенс, А.; Реймонд, П.; Де Джагер, Г.; Солано, Р. (18 февраля 2011 г.). «Факторы транскрипции bHLH арабидопсиса MYC3 и MYC4 являются мишенями репрессоров JAZ и действуют дополнительно с MYC2 при активации реакций жасмоната» . Растительная клетка онлайн . 23 (2): 701–715. дои : 10.1105/tpc.110.080788 . ПМК 3077776 . ПМИД 21335373 .
- ^ Перейти обратно: а б Вастернак, К. (18 мая 2007 г.). «Жасмонаты: обновленная информация о биосинтезе, передаче сигналов и действии при реакции растений на стресс, росте и развитии» . Анналы ботаники . 100 (4): 681–697. дои : 10.1093/aob/mcm079 . ПМЦ 2749622 . ПМИД 17513307 .
- ^ Крилман, Роберт А.; Маллет, Джон Э. (1 июня 1997 г.). «Биосинтез и действие жасмонатов в растениях». Ежегодный обзор физиологии растений и молекулярной биологии растений . 48 (1): 355–381. дои : 10.1146/annurev.arplant.48.1.355 . ПМИД 15012267 .
- ^ Андерсон, Дж.М. (1988). «Зависимое от жасмоновой кислоты повышение уровня специфических полипептидов в суспензионных культурах и проростках сои». Журнал роста и регулирования растений . 7 (4): 203–11. дои : 10.1007/BF02025263 . S2CID 37785073 .
- ^ Пелачо, AM; Минго-Кастель, AM. (1991). «Жасмоновая кислота индуцирует клубнеобразование столонов картофеля, культивируемых in vitro» . Физиология растений . 97 (3): 1253–55. дои : 10.1104/стр.97.3.1253 . ПМК 1081150 . ПМИД 16668517 .
- ^ Хаус, Беттина; Шаршмидт, Сара (1 сентября 2009 г.). «Роль жасмонатов в мутуалистическом симбиозе растений и почвенных микроорганизмов». Фитохимия . 70 (13–14): 1589–1599. doi : 10.1016/j.phytochem.2009.07.003 . ПМИД 19700177 .
- ^ Перейти обратно: а б Пейн, Стефани (2 марта 2022 г.). «Как растения стали хищниками» . Знающий журнал . doi : 10.1146/knowable-030122-1 . Проверено 11 марта 2022 г.
- ^ Хедрих, Райнер; Фукусима, Кенджи (17 июня 2021 г.). «О происхождении плотоядности: молекулярная физиология и эволюция растений на животной диете» . Ежегодный обзор биологии растений . 72 (1): 133–153. doi : 10.1146/annurev-arplant-080620-010429 . ISSN 1543-5008 . ПМИД 33434053 . S2CID 231595236 . Проверено 11 марта 2022 г.
- ^ Чжао, Ю; Тилмони, Р; Бендер, КЛ; Шаллер, А; Он, С.Ю.; Хоу, Джорджия (ноябрь 2003 г.). «Системы вирулентности томатов Pseudomonas syringae pv. способствуют развитию бактериальной пятнистости у томатов, воздействуя на сигнальный путь жасмоната» . Заводской журнал . 36 (4): 485–99. дои : 10.1046/j.1365-313x.2003.01895.x . ПМИД 14617079 .
- ^ Вастернак, К.; Хаус, Б. (июнь 2013 г.). «Жасмонаты: биосинтез, восприятие, передача сигнала и действие при реакции растений на стресс, рост и развитие. Обновление обзора 2007 года в Annals of Botany» . Анналы ботаники . 111 (6): 1021–1058. дои : 10.1093/aob/mct067 . ISSN 0305-7364 . ПМЦ 3662512 . ПМИД 23558912 .
- ^ Перейти обратно: а б Тернер, Дж. Г.; Эллис, К.; Девото, А. (2002). «Сигнальный путь жасмоната» . Растительная клетка . 14 Приложение (7): 153–164. дои : 10.1105/tpc.000679 . ПМК 151253 . ПМИД 12045275 .
- ^ Перейти обратно: а б Курниф, А.; Питерс, CMJ (1 марта 2008 г.). «Перекрестные помехи в оборонной сигнализации» . Физиология растений . 146 (3): 839–844. дои : 10.1104/стр.107.112029 . ПМК 2259093 . ПМИД 18316638 .
- ^ Чини, А.; Фонсека, С.; Фернандес, Г.; Ади, Б.; Чико, Дж. М.; Лоренцо, О.; Гарсиа-Касадо, Г.; Лопес-Видриеро, И.; Лозано, FM; Понсе, MR; Миколь, Дж. Л.; Солано, Р. (2007). «Семейство репрессоров JAZ является недостающим звеном в передаче сигналов жасмоната». Природа . 448 (7154): 666–671. Бибкод : 2007Natur.448..666C . дои : 10.1038/nature06006 . ПМИД 17637675 . S2CID 4383741 .
- ^ Девото, А; Ньето-Ростро, М; Се, Д; Эллис, К; Хармстон, Р.; Патрик, Э; Дэвис, Дж; Шерратт, Л; Коулман, М; Тернер, Дж. Г. (ноябрь 2002 г.). «COI1 связывает передачу сигналов жасмоната и фертильность с комплексом убиквитин-лигазы SCF у арабидопсиса». Заводской журнал . 32 (4): 457–66. дои : 10.1046/j.1365-313x.2002.01432.x . ПМИД 12445118 .
- ^ Шеард, Лаура Б.; Тан, Сюй; Мао, Хайбинь; Уизерс, Джон; Бен-Ниссан, Гили; Хиндс, Томас Р.; Кобаяши, Юичи; Сюй, Фонг-Фу; Шарон, Михал; Просматривай, Джон; Он, Шэн Ян; Ризо, Хосеп; Хоу, Грегг А.; Чжэн, Нин (6 октября 2010 г.). «Восприятие жасмоната корецептором COI1–JAZ, усиленным инозитол-фосфатом» . Природа . 468 (7322): 400–405. Бибкод : 2010Natur.468..400S . дои : 10.1038/nature09430 . ПМК 2988090 . ПМИД 20927106 .