Система секреции типа III
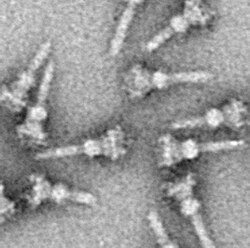
Система секреции типа III ( T3SS или TTSS ) является одной из бактериальных систем секреции, используемых бактериями для секреции своих эффекторных белков в клетки хозяина для содействия вирулентности и колонизации . [1] [2] Хотя система секреции типа III широко рассматривается как эквивалент инъекционной сомы , многие утверждают, что инъекционная система является лишь частью системы секреции типа III, которая также включает в себя такие структуры, как жгутиковый экспортный аппарат. [3] T3SS представляет собой игольчатый белковый комплекс, обнаруженный у нескольких видов патогенных грамотрицательных бактерий .
Обзор
[ редактировать ]Термин «система секреции типа III» был введен в 1993 году. [4] Эта система секреции отличается по крайней мере от пяти других систем секреции, обнаруженных у грамотрицательных бактерий . Многие бактерии, ассоциированные с животными и растениями, обладают сходными T3SS. Эти T3SS схожи в результате конвергентной эволюции, и филогенетический анализ подтверждает модель, в которой грамотрицательные бактерии могут передавать кассету гена T3SS горизонтально другим видам. Некоторые из наиболее изученных T3SS относятся к видам: [ нужна ссылка ]
- Шигеллы (вызывают бактериальную дизентерию ),
- сальмонелла ( брюшной тиф ),
- Escherichia coli ( кишечная флора , некоторые штаммы вызывают пищевое отравление ),
- Вибрионы ( гастроэнтерит и диарея ),
- Буркхолдерия ( сап ),
- Иерсиния ( чума ),
- Хламидиоз ( заболевание, передающееся половым путем ),
- Pseudomonas (заражает человека , животных и растения ) и
- Растительные патогены, такие как Erwinia , Ralstonia и Xanthomonas , а также растительный симбионт Rhizobium .
T3SS состоит примерно из 30 различных белков, что делает его одной из самых сложных систем секреции. Его структура имеет много общего с бактериальными жгутиками (длинными, жесткими внеклеточными структурами, обеспечивающими подвижность ). Некоторые из белков, участвующих в ССТТ, имеют гомологию аминокислотной последовательности с жгутиковыми белками. Некоторые из бактерий, обладающих T3SS, также имеют жгутики и подвижны ( Salmonella например, Shigella ), а некоторые нет ( например, ). С технической точки зрения, секреция типа III используется как для секреции белков, связанных с инфекцией, так и для жгутиковых компонентов. Однако термин «секреция III типа» используется главным образом по отношению к инфекционному аппарату. Бактериальный жгутик имеет общего предка с системой секреции типа III. [5] [6]
T3SS необходимы для патогенности (способности заражать) многих патогенных бактерий. Дефекты T3SS могут сделать бактерию непатогенной. Было высказано предположение, что некоторые неинвазивные штаммы грамотрицательных бактерий утратили T3SS, поскольку энергетически затратная система больше не используется. [7] традиционные антибиотики Хотя в прошлом были эффективны против этих бактерий, постоянно появляются штаммы , устойчивые к антибиотикам . Понимание того, как работает T3SS, и разработка препаратов, специально нацеленных на него, стали важной целью многих исследовательских групп по всему миру с конца 1990-х годов.
Структура
[ редактировать ]| Система секреции типа III | |
|---|---|
 Игольчатый комплекс T3SS | |
| Идентификаторы | |
| Символ | Т3СС |
| TCDB | 1.Б.22 |
| Суперсемейство OPM | 348 |
| белок OPM | 5 TCQ |
Отличительной чертой T3SS является игла. [8] [9] (в более общем смысле, игольчатый комплекс ( NC ) или аппарат T3SS ( T3SA ); также называемый инъектисомой , когда АТФаза исключена; см. ниже). Бактериальные белки, которые необходимо секретировать, переходят из бактериальной цитоплазмы через иглу непосредственно в цитоплазму хозяина. Две цитоплазмы разделяют три мембраны : двойные мембраны (внутренняя и внешняя) грамотрицательных бактерий и эукариотическая мембрана. Игла обеспечивает плавное прохождение через эти высокоселективные и почти непроницаемые мембраны. У одной бактерии может быть несколько сотен игольчатых комплексов, распределенных по мембране. Было высказано предположение, что игольчатый комплекс является универсальным признаком всех ССТТ патогенных бактерий. [10]
Игольчатый комплекс начинается в цитоплазме бактерии, пересекает две мембраны и выступает из клетки. Часть, закрепленная в мембране, является основанием (или базальным тельцем ) ССТТ. Внеклеточная часть — игла. Так называемый внутренний стержень соединяет иглу с основанием. Сама игла, хотя и является самой большой и заметной частью ССТТ, состоит из множества единиц одного белка. Таким образом, большинство различных белков ССТТ — это те, которые создают основу, и те, которые секретируются в организм хозяина. Как упоминалось выше, комплекс игл имеет сходство с бактериальными жгутиками. Точнее, основание игольчатого комплекса структурно очень похоже на основание жгутика; сама игла аналогична жгутиковому крючку — структуре, соединяющей основание с жгутиковой нитью. [11] [12]
Основание состоит из нескольких круглых колец и является первой структурой, построенной в новом игольчатом комплексе. Как только основание готово, оно служит машиной для секреции внешних белков (иглы). Как только весь комплекс завершен, система переключается на секрецию белков, предназначенных для доставки в клетки-хозяева. Предполагается, что игла построена снизу вверх; единицы белка -мономера иглы накапливаются друг на друге, так что единица на кончике иглы добавляется последней. Игольчатая субъединица является одним из самых маленьких белков ССТТ, ее масса составляет около 9 кДа . Каждая игла состоит из 100–150 субъединиц.
Игла T3SS имеет длину около 60–80 нм и внешнюю ширину 8 нм. Он должен иметь минимальную длину, чтобы другие внеклеточные бактериальные структуры ( например, адгезины и слой липополисахарида ) не мешали секреции. Отверстие иглы имеет диаметр 3 нм. Большинство свернутых эффекторных белков слишком велики, чтобы пройти через отверстие иглы, поэтому большинство секретируемых белков должны проходить через иглу в развернутом виде , и эту задачу выполняет АТФаза в основании структуры. [13]
белки ССТТ
[ редактировать ]
Белки T3SS можно разделить на три категории:
- Структурные белки : строят основание, внутренний стержень и иглу.
- Эффекторные белки : секретируются в клетку-хозяина и способствуют инфекции/подавляют защиту клетки-хозяина.
- Шапероны : связывают эффекторы в цитоплазме бактерий, защищают их от агрегации и деградации и направляют к игольчатому комплексу.
Большинство генов ССТТ расположены в оперонах . Эти опероны расположены на бактериальной хромосоме у некоторых видов и на специальной плазмиде у других видов. У сальмонеллы , например, есть хромосомная область, в которой собрано большинство генов ССТТ, так называемый сальмонеллы остров патогенности ( SPI ). Шигеллы , с другой стороны, имеют большую вирулентную плазмиду, на которой расположены все гены ССТТ. Важно отметить, что многие острова патогенности и плазмиды содержат элементы, которые позволяют осуществлять частый горизонтальный перенос генов острова/плазмиды новому виду.
Эффекторные белки, которые должны секретироваться через иглу, должны быть распознаны системой, поскольку они плавают в цитоплазме вместе с тысячами других белков. Распознавание осуществляется посредством сигнала секреции — короткой последовательности аминокислот, расположенной в начале ( N-конце ) белка (обычно в пределах первых 20 аминокислот), которую игольчатый комплекс способен распознавать. В отличие от других систем секреции, сигнал секреции белков T3SS никогда не отщепляется от белка.
Индукция секреции
[ редактировать ]Контакт иглы с клеткой-хозяином запускает секрецию T3SS; [14] Об этом триггерном механизме известно немного (см. ниже). Секрецию также можно индуцировать путем снижения концентрации кальция ионов в питательной среде (для иерсиний и псевдомонад ; это делается путем добавления хелатора, такого как ЭДТА или ЭГТА ) и путем добавления ароматического красителя Конго красный в питательную среду (для шигелл ). например. Эти и другие методы используются в лабораториях для искусственного индуцирования секреции III типа.
Индукция секреции внешними стимулами, помимо контакта с клетками-хозяевами, также происходит in vivo , в инфицированных организмах. Бактерии воспринимают такие сигналы, как температура , pH , осмолярность и уровень кислорода , и используют их, чтобы «решить», активировать ли свой T3SS. Например, сальмонелла может лучше размножаться и проникать в подвздошную кишку , чем в слепую животных кишку . Бактерии могут знать, где они находятся, благодаря различным ионам, присутствующим в этих регионах; подвздошная кишка содержит формиат и ацетат , а слепая кишка - нет. Бактерии чувствуют эти молекулы, определяют, что они находятся в подвздошной кишке, и активируют свой механизм секреции. Молекулы, присутствующие в слепой кишке, такие как пропионат и бутират , оказывают негативное воздействие на бактерии и подавляют секрецию. Холестерин , липид, обнаруженный в мембранах большинства эукариотических клеток, способен индуцировать секрецию шигелл .
Перечисленные выше внешние сигналы регулируют секрецию либо напрямую, либо посредством генетического механизма. несколько транскрипционных факторов , регулирующих экспрессию Известно генов ССТТ. Некоторые из шаперонов, связывающих эффекторы T3SS, также действуют как факторы транскрипции. Был предложен механизм обратной связи: когда бактерия не секретирует, ее эффекторные белки связываются с шаперонами и плавают в цитоплазме. Когда начинается секреция, шапероны отделяются от эффекторов, а последние секретируются и покидают клетку. Одиночные шапероны затем действуют как факторы транскрипции, связываясь с генами, кодирующими их эффекторы, и индуцируя их транскрипцию и, таким образом, производство большего количества эффекторов.
Было предложено, чтобы структуры, подобные инъексомам Type3SS, соединяли наружные и внутренние мембраны грамотрицательных бактерий, чтобы помочь высвободить везикулы внешней мембраны, предназначенные для доставки бактериальных секретов к эукариотическому хозяину или другим клеткам-мишеням in vivo. [15]
T3SS-опосредованная инфекция
[ редактировать ]Эффекторы ССТТ входят в комплекс иглы у основания и продвигаются внутрь иглы к клетке-хозяину. Точный путь проникновения эффекторов в хозяина в основном неизвестен. Ранее предполагалось, что игла сама по себе способна проколоть отверстие в мембране клетки-хозяина; эта теория была опровергнута. Теперь ясно, что некоторые эффекторы, называемые транслокаторами , секретируются первыми и образуют пору или канал ( транслокон ) в мембране клетки-хозяина, через который могут проникнуть другие эффекторы. Мутировавшие бактерии, у которых отсутствуют транслокаторы, способны секретировать белки, но не способны доставлять их в клетки-хозяева. В целом каждый T3SS включает в себя три транслокатора. Некоторые транслокаторы выполняют двойную роль; после участия в образовании пор они проникают в клетку и действуют как настоящие эффекторы.
Эффекторы ССТТ манипулируют клетками-хозяевами несколькими способами. Наиболее поразительным эффектом является усиление поглощения бактерии клеткой-хозяином. Многие бактерии, обладающие T3SS, должны проникать в клетки-хозяева для репликации и распространения инфекции. Эффекторы, которые они вводят в клетку-хозяина, заставляют хозяина поглотить бактерию и практически «съесть» ее. Чтобы это произошло, бактериальные эффекторы манипулируют актина механизмом полимеризации клетки-хозяина. Актин является компонентом цитоскелета , а также участвует в подвижности и изменении формы клеток. Благодаря эффекторам T3SS бактерия может использовать собственные механизмы клетки-хозяина в своих целях. Как только бактерия проникла в клетку, она может легче секретировать другие эффекторы, проникать в соседние клетки и быстро инфицировать всю ткань .
хозяина Также было показано, что эффекторы T3SS вмешиваются в клеточный цикл , и некоторые из них способны индуцировать апоптоз . Одним из наиболее изученных эффекторов ССТТ является IpaB из Shigella flexneri . Он выполняет двойную роль: как транслокатор, создавая поры в мембране клетки-хозяина, так и как эффектор, оказывающий множественные вредные воздействия на клетку-хозяина. Было продемонстрировано, что IpaB индуцирует апоптоз макрофагов животных — клеток иммунной системы — после их поглощения. [16] Позже было показано, что IpaB достигает этого путем взаимодействия с каспазой 1 , основным регуляторным белком в эукариотических клетках. [17]
Другим хорошо охарактеризованным классом эффекторов T3SS являются эффекторы, подобные активаторам транскрипции ( эффекторы TAL ) из Xanthomonas . При введении в растения эти белки могут проникать в ядро растительной клетки, связывать последовательности промотора растения и активировать транскрипцию генов растений, которые способствуют бактериальной инфекции. [18] Недавно было продемонстрировано, что распознавание эффекторной ДНК TAL представляет собой простой код. [19] [20] и это значительно улучшило понимание того, как эти белки могут изменять транскрипцию генов в клетках растения-хозяина.
Нерешенные проблемы
[ редактировать ]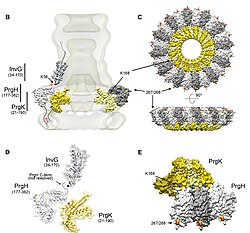
С середины девяностых годов были опубликованы сотни статей о T3SS. Однако многие вопросы, касающиеся системы, остаются нерешенными:
- Белки T3SS . Из примерно 30 белков ССТТ менее 10 в каждом организме были обнаружены непосредственно биохимическими методами. Остальные, возможно, редкие, оказалось трудно обнаружить, и они остаются теоретическими (хотя на многих генах/белках ССТТ проводились генетические, а не биохимические исследования). Локализация каждого белка также до конца не известна.
- Длина иглы . Неизвестно, как бактерия «узнает», что новая игла достигла нужной длины. Существует несколько теорий, в том числе существование «белка-линейки», который каким-то образом соединяет кончик и основание иглы. Добавление новых мономеров к кончику иглы должно растянуть белок-линейку и тем самым сигнализировать о длине иглы до основания.
- Энергетика . Сила, которая управляет прохождением белков внутри иглы, до конца не известна. АТФаза ; связана с основанием ССТТ и участвует в направлении белков в иглу но неясно, поставляет ли он энергию для транспорта.
- Сигнал секреции . Как упоминалось выше, известно существование сигнала секреции в эффекторных белках. Сигнал позволяет системе отличать белки, транспортируемые T3SS, от любых других белков. Его природа, требования и механизм распознавания плохо изучены, но недавно были разработаны методы прогнозирования того, какие бактериальные белки могут транспортироваться системой секреции типа III. [22]
- Активация секреции . Бактерия должна знать, когда настало время секретировать эффекторы. Ненужная секреция, когда поблизости нет клетки-хозяина, является расточительной для бактерии с точки зрения энергии и ресурсов. Бактерия каким-то образом способна распознать контакт иглы с клеткой-хозяином. Как это сделать, все еще изучается, и метод вполне может зависеть от возбудителя. Некоторые теории постулируют деликатное конформационное изменение структуры иглы при контакте с клеткой-хозяином; это изменение, возможно, служит для основания сигналом к началу секреции. Один метод распознавания был обнаружен у сальмонелл , который основан на распознавании цитозольного рН клетки-хозяина посредством T3SS, кодируемого островом патогенности 2, чтобы включить секрецию эффекторов. [23]
- Связывание шаперонов . Неизвестно, когда шапероны связывают свои эффекторы (во время или после трансляции ) и как они диссоциируют от своих эффекторов перед секрецией.
- Эффекторные механизмы . Хотя с начала 21 века было открыто многое о том, как эффекторы ССТТ манипулируют хозяином, большинство эффектов и путей остаются неизвестными.
- Эволюция . Как уже упоминалось, T3SS тесно связан с жгутиком бактерий. [24] Существуют три конкурирующие гипотезы: [25] во-первых, жгутик развился первым, а T3SS произошел от этой структуры, во-вторых, что T3SS развился первым, а жгутик произошел от него, и в-третьих, что эти две структуры произошли от общего предка. Были некоторые разногласия по поводу различных сценариев, [5] [25] поскольку все они объясняют гомологию белков между двумя структурами, а также их функциональное разнообразие. [26] Тем не менее, недавние филогеномические данные подтверждают гипотезу о том, что T3SS происходит из жгутика в результате процесса, включающего первоначальную потерю гена, а затем его приобретение. [27] Ключевым этапом последнего процесса было привлечение секретинов к T3SS, событие, которое происходило по крайней мере трижды из других мембран-ассоциированных систем.
Номенклатура белков ССТТ
[ редактировать ]
С начала 1990-х годов новые белки ССТТ постоянно обнаруживаются у различных видов бактерий. Сокращения даны независимо для каждой серии белков в каждом организме, и названия обычно мало что говорят о функции белка. Позднее было показано, что некоторые белки, открытые независимо у разных бактерий, гомологичны ; однако исторические названия в основном сохранены, и этот факт может вызвать путаницу. Например, белки SicA, IpgC и SycD являются гомологами Salmonella , Shigella и Yersinia соответственно, но последняя буква («серийный номер») в их названии об этом не свидетельствует.
Ниже приводится краткое изложение наиболее распространенных названий белковых серий у нескольких видов, содержащих T3SS. Обратите внимание, что эти названия включают белки, которые образуют механизм ССТТ, а также секретируемые эффекторные белки :
- Иерсиния
- Yop : иерсинии . внешний белок
- Ysc : секреция иерсинии (компонент)
- Ypk : иерсинии. протеинкиназа
- Сальмонелла
- Спа : Поверхностное представление антигена
- Sic : сальмонеллы Сопровождающий вторжение
- Глоток : сальмонеллы. белок инвазии
- Prg : ген, репрессированный PhoP.
- Инв : Вторжение
- Org : Ген, регулирующий кислород.
- Ssp : сальмонеллами. белок, секретируемый
- Iag : Ген, связанный с инвазией.
- Шигелла
- Ipg : ген плазмидной инвазии.
- Ipa : плазмидный антиген инвазии.
- Mxi : мембранная экспрессия Ipa.
- Спа : Поверхностное представление антигена
- Osp : внешний шигеллы . белок
- Эшерихия
- Tir : транслоцированный рецептор интимина.
- Сентябрь : Секреция E. coli. белков
- Esc : секрет эшерихий (компонент)
- Esp : эшерихий . белок секреции
- Ces : Шаперон E. coli. секреции
- Псевдомонада
- Hrp : Гиперчувствительная реакция и патогенность
- Hrc : гиперчувствительная реакция сохранена (или Hrp сохранена).
- Ризобий
- Nop : узелковый белок
- Rhc : ризобии сохранились.
- В нескольких видах:
- Источник : Вирулентность
- «Протохламидия амебофила»
- "член глоссинидиуса" [28]
После этих сокращений следует буква или цифра. Буквы обычно обозначают «серийный номер», либо хронологический порядок открытия, либо физический порядок появления гена в опероне . Числа, в более редком случае, обозначают молекулярную массу белка в кДа . Примеры: IpaA, IpaB, IpaC; MxiH, MxiG, MxiM; Спа9, Спа47.
Во всех T3SS присутствуют несколько ключевых элементов: мономер иглы, внутренний стержень иглы, кольцевые белки, два транслокатора, белок кончика иглы, белок-линейка (который, как полагают, определяет длину иглы; см. выше) и АТФаза , которая поставляет энергию для секреции. В следующей таблице показаны некоторые из этих ключевых белков четырех бактерий, содержащих T3SS:
| ↓ Функция/Род → | Шигелла | Сальмонелла | Иерсиния | Эшерихия |
|---|---|---|---|---|
| Игольчатый мономер | MxiH | ПргИ | YscF | ЭскФ |
| Внутренний стержень | MxiI | Ответ | YscI | Ты выходишь |
| Белок на кончике иглы | IPAD | СипД | ЛкрВ | Испания |
| Транслокатор | ИпаБ | СипБ | ЙопБ | EspD |
| Транслокатор | IPAC | СипК | ЙопД | ЭспБ |
| Сопровождающий двух транслокаторов | ИПгС | СикА | SycD | ЦЕСД |
| АТФаза | Спа47 | ИнвК | YscN | Сентябрь (EscN) |
| Линейка белка | Спа32 | ИнвДж | YscP | Орф16 |
| Выключатель | Спа40 | СпаС | ЕмГУ | ЭскУ |
| привратник | MxiC | Инв | ЙопН (ТайА) | сентябрь |
Методы, используемые в исследовании T3SS
[ редактировать ]Выделение игольчатых комплексов ССТТ
[ редактировать ]Выделение крупных, хрупких, гидрофобных мембранных структур из клеток на протяжении многих лет представляло собой проблему. Однако к концу 1990-х годов было разработано несколько подходов к изоляции NC T3SS. В 1998 году первые НК были выделены из Salmonella typhimurium . [29]
Для выделения бактерии выращивают в большом объеме жидкой питательной среды до достижения лог-фазы . Затем их центрифугируют ; супернатант , (среду) отбрасывают, а осадок (бактерии) ресуспендируют в буфере для лизиса обычно содержащем лизоцим и иногда детергент, такой как LDAO или Triton X-100 . Этот буфер разрушает клеточную стенку . После нескольких циклов лизиса и промывки вскрывшиеся бактерии подвергают серии ультрацентрифугирования . Эта обработка обогащает крупные макромолекулярные структуры и удаляет более мелкие клеточные компоненты. Необязательно конечный лизат подвергают дальнейшей очистке CsCl градиентом плотности .
Дополнительный подход к дальнейшей очистке использует аффинную хроматографию . Рекомбинантные белки T3SS, несущие белковую метку ( например, гистидиновую метку ), производятся путем молекулярного клонирования , а затем вводятся ( трансформируются ) в исследуемые бактерии. После первоначального выделения НК, как описано выше, лизат пропускают через колонку, покрытую частицами с высоким сродством к метке (в случае гистидиновых меток: никеля ионы ). Меченый белок сохраняется в колонке, а вместе с ним и весь игольчатый комплекс. С помощью таких методов можно достичь высокой степени чистоты. Эта чистота важна для многих деликатных анализов, которые использовались для характеристики NC.
Эффекторы типа III были известны с начала 1990-х годов, но способ их доставки в клетки-хозяева оставался полной загадкой. Гомология между многими жгутиковыми белками и белками T3SS привела исследователей к подозрению в существовании внешней структуры T3SS, похожей на жгутики. Идентификация и последующее выделение структуры иглы позволило исследователям:
- подробно охарактеризовать трехмерную структуру НК и на основе этого сделать выводы относительно механизма секреции (например, что узкая ширина иглы требует разворачивания эффекторов перед секрецией),
- проанализировать белковые компоненты НК, подвергнув изолированные иглы протеомному анализу (см. ниже),
- назначьте роли различным компонентам NC, выключив гены T3SS, изолируя NC от мутировавших бактерий и исследуя изменения, вызванные мутациями.
Микроскопия, кристаллография и ЯМР твердого тела
[ редактировать ]Как и почти все белки, визуализация NC T3SS возможна только с помощью электронной микроскопии . Первые изображения НК (1998 г.) показали игольчатые структуры, выступающие из клеточной стенки живых бактерий, а также плоские двумерные изолированные НК. [29] В 2001 году изображения НК Shigella flexneri были проанализированы в цифровом виде и усреднены, чтобы получить первую полу3D-структуру НК. [8] Спиральная структура НК Shigella flexneri была разрешена с разрешением 16 Å с помощью рентгеновских дифракции лучей в 2003 г. [30] а годом позже размером 17 Å была опубликована 3D-структура НК Salmonella typhimurium . [31] Последние достижения и подходы позволили создавать 3D-изображения NC с высоким разрешением. [32] [33] дальнейшее выяснение сложной структуры НК.
За прошедшие годы было кристаллизовано множество белков ССТТ. К ним относятся структурные белки НК, эффекторы и шапероны. Первой структурой мономера игольчатого комплекса была ЯМР-структура BsaL из «Burkholderia pseudomallei», а затем кристаллическая структура MixH из Shigella flexneri , которые были решены в 2006 году. [34] [35]
В 2012 году сочетание рекомбинантного производства игл дикого типа, твердотельного ЯМР и электронной микроскопии. [36] и моделирование Rosetta выявило супрамолекулярные интерфейсы и, в конечном итоге, полную атомную структуру иглы T3SS Salmonella typhimurium . [37] Было показано, что субъединицы PrgI из 80 остатков образуют правозакрученную спиральную сборку примерно с 11 субъединицами на два витка, аналогичную жгутику Salmonella typhimurium . Модель также выявила расширенный аминоконцевой домен, который расположен на поверхности иглы, в то время как высококонсервативный карбокси-конец направлен в сторону просвета. [37]
Протеомика
[ редактировать ]Для идентификации массива белков, составляющих T3SS, было использовано несколько методов. Изолированные игольчатые комплексы можно разделить с помощью SDS-PAGE . Полосы, появляющиеся после окрашивания, можно отдельно вырезать из геля и проанализировать с помощью секвенирования белков и масс-спектрометрии . Структурные компоненты НК можно отделить друг от друга (например, игольчатую часть от базовой части) и, анализируя эти фракции, можно вывести белки, участвующие в каждой из них. Альтернативно, изолированные NC могут быть непосредственно проанализированы с помощью масс-спектрометрии без предварительного электрофореза , чтобы получить полную картину протеома NC .
Генетические и функциональные исследования
[ редактировать ]Исследователи манипулировали T3SS у многих бактерий. Наблюдение за влиянием отдельных манипуляций можно использовать для понимания роли каждого компонента системы. Примеры манипуляций:
- Делеция одного или нескольких генов ССТТ ( нокаут гена ).
- Сверхэкспрессия одного или нескольких генов СДТ3 (другими словами: производство in vivo белка СТ3 в количествах, больших, чем обычно).
- Точечные или региональные изменения в генах или белках ССТТ. Это делается для того, чтобы определить функцию определенных аминокислот или участков белка.
- Введение гена или белка одного вида бактерий в другой (анализ перекрестной комплементации). Это делается для того, чтобы проверить различия и сходства между двумя T3SS.
Манипуляции с компонентами ССТТ могут влиять на некоторые аспекты функции и патогенности бактерий. Примеры возможных влияний:
- Способность бактерий проникать в клетки-хозяева в случае внутриклеточных патогенов. Это можно измерить с помощью анализа инвазии ( анализ защиты гентамицина ).
- Способность внутриклеточных бактерий мигрировать между клетками-хозяевами.
- Способность бактерий убивать клетки-хозяева. Это можно измерить несколькими методами, например, с помощью анализа высвобождения ЛДГ , в котором фермент ЛДГ, выделяющийся из мертвых клеток, идентифицируется путем измерения его ферментативной активности.
- Способность ССТТ секретировать определенный белок или секретировать вообще. Чтобы это проверить, у бактерий, растущих в жидкой среде, индуцируют секрецию. Затем бактерии и среду разделяют центрифугированием, а фракцию среды (супернатант) затем анализируют на наличие секретируемых белков. Чтобы предотвратить секрецию обычно секретируемого белка, к нему можно искусственно присоединить большую молекулу. Если несекретируемый белок остается «застрявшим» в нижней части игольчатого комплекса, секреция эффективно блокируется.
- Способность бактерий собирать интактный игольчатый комплекс. НК можно выделить из обработанных бактерий и исследовать под микроскопом. Однако незначительные изменения не всегда могут быть обнаружены с помощью микроскопии.
- Способность бактерий заражать живых животных или растения. Даже если in vitro будет доказано, что манипулируемые бактерии способны инфицировать клетки-хозяева, их способность поддерживать инфекцию в живом организме не может считаться само собой разумеющейся.
- Уровни экспрессии других генов. Это можно проверить несколькими способами, в частности, нозерн-блоттингом и RT-PCR . Уровни экспрессии всего генома можно оценить с помощью микроматрицы . С помощью этих методов были обнаружены многие факторы транскрипции и регуляторные сети типа III.
- Рост и приспособленность бактерий.
Ингибиторы T3SS
[ редактировать ]Было обнаружено несколько соединений, которые ингибируют ССТТ у грамотрицательных бактерий , включая гуадиномины , которые естественным образом продуцируются видами Streptomyces . [38] моноклональные антитела , которые также ингибируют T3SS. Были разработаны [39] Было показано, что Aurodox, антибиотик, способный ингибировать трансляцию белков T3SS, способен предотвращать эффекторы T3SS in vitro и на животных моделях. [40] [41]
Инструменты прогнозирования сигнальных пептидов типа III
[ редактировать ]Ссылки
[ редактировать ]- ^ Лара-Техеро М., Галан Дж.Э. (март 2019 г.). «Инъектисома, сложная наномашина для инъекции белка в клетки млекопитающих» . ЭкоСал Плюс . 8 (2). doi : 10.1128/ecosalplus.ESP-0039-2018 . ПМК 6450406 . ПМИД 30942149 .
- ^ МакХью Р.Э., О'Бойл Н., Коннолли Дж.П., Хоскиссон П.А., Роу А.Дж. (февраль 2019 г.). «Характеристика механизма действия ауродокса, ингибитора системы секреции типа III Streptomyces goldiniensis» . Инфекция и иммунитет . 87 (2): e00595–18. дои : 10.1128/IAI.00595-18 . ПМК 6346137 . ПМИД 30455200 .
- ^ Халте М., Эрхардт М. (январь 2021 г.). «Экспорт белка через систему секреции бактериального жгутика III типа» . Биомолекулы . 11 (2): 186. doi : 10.3390/biom11020186 . ПМЦ 7911332 . ПМИД 33572887 .
- ^ Салмонд ГП, Ривз П.Дж. (январь 1993 г.). «Защитники мембранного движения и секреция белка грамотрицательными бактериями». Тенденции биохимических наук . 18 (1): 7–12. дои : 10.1016/0968-0004(93)90080-7 . ПМИД 8438237 .
- ^ Перейти обратно: а б Гофна У, Рон Э.З., Граур Д. (июль 2003 г.). «Бактериальные системы секреции типа III являются древними и развиваются в результате множественных событий горизонтального переноса». Джин . 312 : 151–163. дои : 10.1016/S0378-1119(03)00612-7 . ПМИД 12909351 .
- ^ Нгуен Л., Полсен И.Т., Чиу Дж., Хьюк С.Дж., Сайер М.Х. (апрель 2000 г.). «Филогенетический анализ компонентов систем секреции белков типа III». Журнал молекулярной микробиологии и биотехнологии . 2 (2): 125–144. ПМИД 10939240 .
- ^ Гонг Х, Ву ГП, Бай Ю, Ян Э, Лю Ф, Лу С (январь 2010 г.). «Дифференциальная экспрессия факторов системы секреции Salmonella типа III InvJ, PrgJ, SipC, SipD, SopA и SopB в культурах и у мышей» . Микробиология . 156 (Часть 1): 116–127. дои : 10.1099/mic.0.032318-0 . ПМЦ 2889428 . ПМИД 19762438 .
- ^ Перейти обратно: а б Блокер А, Жуири Н., Ларке Э., Гунон П., Эбель Ф., Парсот С. и др. (февраль 2001 г.). «Структура и состав «игольчатого комплекса» Shigella flexneri, входящего в состав его секретона III типа» . Молекулярная микробиология . 39 (3): 652–663. дои : 10.1046/j.1365-2958.2001.02200.x . ПМИД 11169106 .
- ^ Галан Дж. Э., Вольф-Ватц Х. (ноябрь 2006 г.). «Доставка белка в эукариотические клетки с помощью секреторных машин III типа». Природа . 444 (7119): 567–573. Бибкод : 2006Natur.444..567G . дои : 10.1038/nature05272 . ПМИД 17136086 . S2CID 4411244 .
- ^ Паллен М.Дж., Бейли К.М., Beatson SA (апрель 2006 г.). «Эволюционные связи между FliH/YscL-подобными белками из бактериальных систем секреции типа III и компонентами второго стебля FoF1 и вакуолярными АТФазами» . Белковая наука . 15 (4): 935–941. дои : 10.1110/ps.051958806 . ПМК 2242474 . ПМИД 16522800 .
- ^ Айзава С.И. (август 2001 г.). «Бактериальные жгутики и системы секреции III типа» . Письма FEMS по микробиологии . 202 (2): 157–164. дои : 10.1111/j.1574-6968.2001.tb10797.x . ПМИД 11520608 .
- ^ Дулиттл В.Ф., Жаксыбаева О. (июль 2007 г.). «Эволюция: уменьшаемая сложность – случай бактериальных жгутиков» . Современная биология . 17 (13): 510–512 Р5. дои : 10.1016/j.cub.2007.05.003 . ПМИД 17610831 . S2CID 17452659 .
- ^ Акеда Ю., Галан Дж.Э. (октябрь 2005 г.). «Высвобождение шаперонов и развертывание субстратов при секреции III типа». Природа . 437 (7060): 911–915. Бибкод : 2005Natur.437..911A . дои : 10.1038/nature03992 . ПМИД 16208377 . S2CID 4355750 .
- ^ Кимбро Т.Г., Миллер С.И. (сентябрь 2000 г.). «Вклад компонентов секрета Salmonella typhimurium типа III в образование игольчатых комплексов» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (20): 11008–11013. Бибкод : 2000PNAS...9711008K . дои : 10.1073/pnas.200209497 . ПМК 27139 . ПМИД 10984518 .
- ^ ЯшРой РЦ (2003). «Интоксикация эукариотических клеток грамотрицательными патогенами: новая модель бактериального нановезикулярного экзоцитоза, связанного с внешней мембраной, для системы секреции типа III» . Международная токсикология . 10 (1): 1–9.
- ^ Зыхлинский А., Кенни Б., Менар Р., Прево М.К., Холланд И.Б., Сансонетти П.Дж. (февраль 1994 г.). «IpaB опосредует апоптоз макрофагов, индуцированный Shigella flexneri». Молекулярная микробиология . 11 (4): 619–627. дои : 10.1111/j.1365-2958.1994.tb00341.x . ПМИД 8196540 . S2CID 40167923 .
- ^ Хилби Х., Мосс Дж.Э., Херш Д., Чен Ю., Арондел Дж., Банерджи С. и др. (декабрь 1998 г.). «Апоптоз, индуцированный шигеллами, зависит от каспазы-1, которая связывается с IpaB» . Журнал биологической химии . 273 (49): 32895–32900. дои : 10.1074/jbc.273.49.32895 . ПМИД 9830039 .
- ^ Бох Дж., Бонас У (2010). «Эффекторы семейства Xanthomonas AvrBs3 типа III: открытие и функция». Ежегодный обзор фитопатологии . 48 : 419–436. doi : 10.1146/annurev-phyto-080508-081936 . ПМИД 19400638 .
- ^ Москва М.Ю., Богданов А.Ю. (декабрь 2009 г.). «Простой шифр управляет распознаванием ДНК эффекторами TAL». Наука . 326 (5959): 1501. Бибкод : 2009Sci...326.1501M . дои : 10.1126/science.1178817 . ПМИД 19933106 . S2CID 6648530 .
- ^ Бох Дж., Шольце Х., Шорнак С., Ландграф А., Хан С., Кей С. и др. (декабрь 2009 г.). «Взлом кода специфичности связывания ДНК эффекторов TAL-типа III». Наука . 326 (5959): 1509–1512. Бибкод : 2009Sci...326.1509B . дои : 10.1126/science.1178811 . ПМИД 19933107 . S2CID 206522347 .
- ^ Шрайдт О., Лефебр М.Д., Бруннер М.Дж., Шмид В.Х., Шмидт А., Радикс Дж. и др. (апрель 2010 г.). Стеббинс CE (ред.). «Топология и организация компонентов игольчатого комплекса секреции Salmonella typhimurium III типа» . ПЛОС Патогены . 6 (4): e1000824. doi : 10.1371/journal.ppat.1000824 . ПМЦ 2848554 . ПМИД 20368966 .
- ^ Гринберг М., Годзик А. (апрель 2009 г.). Стеббинс CE (ред.). «Сигнал для сигнализации найден» . ПЛОС Патогены . 5 (4): e1000398. дои : 10.1371/journal.ppat.1000398 . ПМК 2668190 . ПМИД 19390616 .
- ^ Ю XJ, МакГурти К., Лю М., Ансворт К.Е., Холден Д.В. (май 2010 г.). «Ощущение pH внутриклеточными сальмонеллами индуцирует эффекторную транслокацию» . Наука . 328 (5981): 1040–1043. Бибкод : 2010Sci...328.1040Y . дои : 10.1126/science.1189000 . hdl : 10044/1/19679 . ПМК 6485629 . ПМИД 20395475 .
- ^ Медини Д., Коваччи А., Донати С. (декабрь 2006 г.). «Семейства сетей гомологии белков демонстрируют поэтапную диверсификацию систем секреции типа III и типа IV» . PLOS Вычислительная биология . 2 (12): е173. Бибкод : 2006PLSCB...2..173M . дои : 10.1371/journal.pcbi.0020173 . ПМК 1676029 . ПМИД 17140285 .
- ^ Перейти обратно: а б Сайер М.Х. (март 2004 г.). «Эволюция систем секреции белков бактериального типа III». Тенденции в микробиологии . 12 (3): 113–115. дои : 10.1016/j.tim.2004.01.003 . ПМИД 15001186 .
- ^ Макканн ХК, Гуттман Д.С. (2008). «Эволюция системы секреции типа III и ее эффекторов в растительно-микробных взаимодействиях» . Новый фитолог . 177 (1): 33–47. дои : 10.1111/j.1469-8137.2007.02293.x . ПМИД 18078471 .
- ^ Эбби СС, Роча EP (сентябрь 2012 г.). «Нежгутиковая система секреции III типа произошла из бактериального жгутика и диверсифицировалась в системы, адаптированные к клетке-хозяину» . ПЛОС Генетика . 8 (9): e1002983. дои : 10.1371/journal.pgen.1002983 . ПМК 3459982 . ПМИД 23028376 .
- ^ Моран Н.А. (февраль 2001 г.). «Бактериальные зверинцы внутри насекомых» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (4): 1338–1340. Бибкод : 2001PNAS...98.1338M . дои : 10.1073/pnas.98.4.1338 . ПМК 33380 . ПМИД 11171951 .
- ^ Перейти обратно: а б Кубори Т., Мацусима Ю., Накамура Д., Уралил Дж., Лара-Техеро М., Сухан А. и др. (апрель 1998 г.). «Супрамолекулярная структура системы секреции белка Salmonella typhimurium типа III». Наука . 280 (5363): 602–605. Бибкод : 1998Sci...280..602K . дои : 10.1126/science.280.5363.602 . ПМИД 9554854 .
- ^ Кордес Ф.С., Комория К., Ларкет Э., Ян С., Эгельман Э.Х., Блокер А., Леа С.М. (май 2003 г.). «Спиральная структура иглы секреторной системы типа III Shigella flexneri» . Журнал биологической химии . 278 (19): 17103–17107. дои : 10.1074/jbc.M300091200 . ПМИД 12571230 .
- ^ Марловиц Т.С., Кубори Т., Сухан А., Томас Д.Р., Галан Дж.Е., Унгер В.М. (ноябрь 2004 г.). «Структурные данные о сборке комплекса секреторной иглы типа III» . Наука . 306 (5698): 1040–1042. Бибкод : 2004Sci...306.1040M . дои : 10.1126/science.1102610 . ПМК 1459965 . ПМИД 15528446 .
- ^ Сани М., Аллауи А., Фузетти Ф., Остергетель Г.Т., Кигстра В., Букема Э.Дж. (2007). «Структурная организация игольчатого комплекса секреторного аппарата III типа Shigella flexneri» (PDF) . Микрон . 38 (3): 291–301. дои : 10.1016/j.micron.2006.04.007 . hdl : 11370/9ee8c380-a931-4313-89cf-d9faa49cdf3b . ПМИД 16920362 .
- ^ Ходжкинсон Дж.Л., Хорсли А., Стабат Д., Саймон М., Джонсон С., да Фонсека ПК и др. (май 2009 г.). «Трехмерная реконструкция трансмембранных областей T3SS шигеллы обнаруживает 12-кратную симметрию и новые особенности во всем» . Структурная и молекулярная биология природы . 16 (5): 477–485. дои : 10.1038/nsmb.1599 . ПМЦ 2681179 . ПМИД 19396171 .
- ^ Чжан Л., Ван Ю, Пикинг В.Л., Пикинг В.Д., Де Гузман Р.Н. (июнь 2006 г.). «Структура раствора мономерного BsaL, игольчатого белка секреции III типа Burkholderia pseudomallei». Журнал молекулярной биологии . 359 (2): 322–330. дои : 10.1016/j.jmb.2006.03.028 . ПМИД 16631790 .
- ^ Дин Дж.Э., Роверси П., Кордес Ф.С., Джонсон С., Кенджейл Р., Дэниел С. и др. (август 2006 г.). «Молекулярная модель иглы системы секреции типа III: значение для восприятия клеток-хозяев» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12529–12533. Бибкод : 2006PNAS..10312529D . дои : 10.1073/pnas.0602689103 . ПМК 1567912 . ПМИД 16888041 .
- ^ Галкин В.Е., Шмид В.Х., Шрайдт О., Марловиц Т.С., Эгельман Э.Х. (март 2010 г.). «Структура иглы секреторной системы Salmonella typhimurium типа III демонстрирует отличие от жгутиковой системы» . Журнал молекулярной биологии . 396 (5): 1392–1397. дои : 10.1016/j.jmb.2010.01.001 . ПМК 2823972 . ПМИД 20060835 .
- ^ Перейти обратно: а б Локе А., Сгуракис Н.Г., Гупта Р., Гиллер К., Ридель Д., Гусманн С. и др. (май 2012 г.). «Атомная модель иглы системы секреции III типа» . Природа . 486 (7402): 276–279. Бибкод : 2012Natur.486..276L . дои : 10.1038/nature11079 . ПМЦ 3598588 . ПМИД 22699623 .
- ^ Холмс Т.К., Мэй А.Е., Салета-Ривера К., Руби Дж.Г., Скьюс-Кокс П., Фишбах М.А. и др. (октябрь 2012 г.). «Молекулярные данные о биосинтезе гуадиномина: ингибитора системы секреции III типа» . Журнал Американского химического общества . 134 (42): 17797–17806. дои : 10.1021/ja308622d . ПМЦ 3483642 . ПМИД 23030602 .
- ^ Тойретцбахер Ю, Пиддок LJ (июль 2019 г.). «Нетрадиционные антибактериальные терапевтические возможности и проблемы» . Клетка-хозяин и микроб . 26 (1): 61–72. дои : 10.1016/j.chom.2019.06.004 . ПМИД 31295426 .
- ^ Пюлкко Т., Илина П., Таммела П. (май 2021 г.). «Разработка и валидация скринингового анализа высокого содержания ингибиторов энтеропатогенной адгезии E. coli» . Журнал микробиологических методов . 184 : 106201. doi : 10.1016/j.mimet.2021.106201 . ПМИД 33713725 .
- ^ Кимура К., Ивацуки М., Нагаи Т., Мацумото А., Такахаши Ю., Сиоми К. и др. (февраль 2011 г.). «Низкомолекулярный ингибитор системы секреции бактерий III типа защищает от заражения in vivo Citrobacter Rodentium» . Журнал антибиотиков . 64 (2): 197–203. дои : 10.1038/ja.2010.155 . ПМИД 21139624 .
Дальнейшее чтение
[ редактировать ]- Мгновенное понимание химического состава инъекционного препарата от Королевского химического общества.
- Взаимодействие хозяин-патоген у Pseudomonas syringae pv. помидоры и томатные растения приводят к бактериальной пятнистости.