Сальмонелла
| Сальмонелла | |
|---|---|
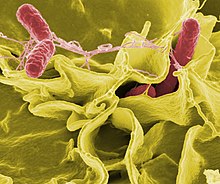 | |
| с улучшенным цветом, Растровая электронная микрофотография показывающая проникновение Salmonella Typhimurium (красный) в культивируемые клетки человека. | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Псевдомонадота |
| Сорт: | Гаммапротеобактерии |
| Заказ: | Энтеробактерии |
| Семья: | Энтеробактерии |
| Род: | Сальмонелла Линьер , 1900 год. |
| Виды и подвиды [1] | |
| |
Сальмонеллы — род палочковидных грамотрицательных (бацилл) бактерий семейства Enterobacteriaceae . Двумя известными видами сальмонеллы являются Salmonella enterica и Salmonella bongori . S. enterica является типовым видом и делится на шесть подвидов. [2] [3] которые включают более 2650 серотипов . [4] Сальмонелла была названа в честь Дэниела Элмера Салмона (1850–1914), американского ветеринарного хирурга .
сальмонелл Виды представляют собой неспорообразующие , преимущественно подвижные энтеробактерии с диаметром клеток от 0,7 до 1,5 мкм , длиной от 2 до 5 мкм и перитрихальными жгутиками (по всему телу клетки, что позволяет им двигаться). [5] Они являются хемотрофами , получающими энергию в результате реакций окисления и восстановления , используя органические источники. Они также являются факультативными анаэробами , способными генерировать аденозинтрифосфат с кислородом («аэробно»), когда он доступен, или использовать другие акцепторы электронов или ферментацию («анаэробно»), когда кислород недоступен. [5]
сальмонелл Виды являются внутриклеточными возбудителями . [6] из которых определенные серотипы вызывают такое заболевание, как сальмонеллез . Большинство инфекций происходит в результате употребления в пищу продуктов, загрязненных фекалиями. Серотипы брюшнотифозной сальмонеллы могут передаваться только между людьми и могут вызывать заболевания пищевого происхождения, а также брюшной тиф и паратиф. Брюшной тиф вызывается брюшнотифозной сальмонеллой, которая проникает в кровоток, а также распространяется по всему организму, проникая в органы и выделяя эндотоксины (септическая форма). Это может привести к опасному для жизни гиповолемическому шоку и септическому шоку и требует интенсивной терапии , включая антибиотики .
Нетифоидные сальмонелл серотипы являются зоонозными и могут передаваться от животных и между людьми. Обычно они проникают только в желудочно-кишечный тракт и вызывают сальмонеллез, симптомы которого можно устранить без антибиотиков. Однако в странах Африки к югу от Сахары нетифоидная сальмонелла может быть инвазивной и вызывать паратиф , что требует немедленного лечения антибиотиками. [7]
Таксономия [ править ]
Род Salmonella входит в семейство Enterobacteriaceae. Его таксономия была пересмотрена и может привести к путанице. Род включает два вида: S. bongori и S. enterica , последний из которых делится на шесть подвидов: S. e. enterica , S.e. саламаэ , С. е. arizonae , S. e. diarizonae , S. e. houtenae и S. e. Индика . [8] [9] Таксономическая группа содержит более 2500 серотипов (также сероваров), определенных на основе соматического О ( липополисахаридного ) и жгутикового Н-антигенов ( классификация Кауфмана-Уайта ). Полное название серотипа указывается, например, Salmonella enterica subsp. enterica серотипа Typhimurium, но может быть сокращенно Salmonella Typhimurium. Дальнейшая дифференциация штаммов для помощи в клинических и эпидемиологических исследованиях может быть достигнута с помощью тестирования чувствительности к антибиотикам и других методов молекулярной биологии, таких как гель-электрофорез в импульсном поле , мультилокусное типирование последовательностей и, все чаще, полногеномное секвенирование . Исторически сальмонеллы клинически подразделялись на инвазивные (брюшной тиф) и неинвазивные (нетифоидные сальмонеллы) в зависимости от предпочтений хозяина и проявлений заболевания у человека. [10]
История [ править ]
Сальмонелла была впервые обнаружена в 1880 году Карлом Эбертом в пейеровых бляшках и селезенке больных брюшным тифом. [11] Четыре года спустя Георг Теодор Гаффки смог вырастить возбудитель в чистой культуре. [12] Год спустя ученый-медик Теобальд Смит обнаружил то, что позже будет известно как Salmonella enterica (var. Choleraesuis). В то время Смит работал лаборантом в ветеринарном отделе Министерства сельского хозяйства США . Подразделением руководил Дэниел Элмер Салмон , ветеринарный патологоанатом. [13] Первоначально является Salmonella считалось, что возбудителем холеры свиней Choleraesuis , поэтому Салмон и Смит назвали ее «бациллой свиной холеры». Название «Сальмонелла» не использовалось до 1900 года, когда Жозеф Леон Линьер предложил назвать возбудитель, открытый группой Сальмона, «Сальмонеллой» в его честь. [14] : 16
В конце 1930-х годов австралийский бактериолог Нэнси Аткинсон основала лабораторию типирования сальмонеллы – одну из трех в мире на тот момент – в Южной Австралии правительства Лаборатории патологии и бактериологии в Аделаиде (позже Институт медицинских и ветеринарных наук). ). Именно здесь Аткинсон описала несколько новых штаммов сальмонеллы, в том числе Salmonella Adelaide, выделенную в 1943 году. Аткинсон опубликовала свою работу по сальмонеллам в 1957 году. [15]
Серотипирование [ править ]
Серотипирование осуществляется путем смешивания клеток с антителами к определенному антигену. Это может дать некоторое представление о риске. Исследование 2014 года показало, что S. Reading очень распространен среди молодых индеек , но не вносит существенного вклада в развитие сальмонеллеза человека. [16] Серотипирование может помочь в выявлении источника заражения путем сопоставления серотипов людей с серотипами предполагаемого источника инфекции. [17] Соответствующее профилактическое лечение можно определить на основе известной устойчивости серотипа к антибиотикам. [18]
Новые методы «серотипирования» включают xMAP и ПЦР в реальном времени — два метода, основанные на последовательностях ДНК, а не на реакциях антител. Эти методы потенциально могут быть быстрее благодаря достижениям в технологии секвенирования. Эти системы «молекулярного серотипирования» фактически выполняют генотипирование генов, определяющих поверхностные антигены. [19] [20]
, культивирования и Условия роста обнаружения

Большинство подвидов сальмонелл выделяют сероводород . [21] которые можно легко обнаружить, выращивая их на среде, содержащей сульфат железа , такой как тот, который используется в тесте с тройным сахаром железа . Большинство изолятов существуют в двух фазах: подвижной и неподвижной. Культуры, которые неподвижны при первичной культуре, можно перевести в подвижную фазу с помощью пробирки Крейги или кюветного планшета . [22] Бульон RVS можно использовать для обогащения видов сальмонелл для обнаружения в клинических образцах. [23]
Сальмонеллу также можно обнаружить и подтипировать с помощью мультиплексного анализа. [24] или полимеразная цепная реакция в реальном времени (qPCR) [25] из выделенной ДНК сальмонеллы .
математические модели кинетики роста сальмонелл для курицы, свинины, томатов и дынь. Разработаны [26] [27] [28] [29] [30] Сальмонеллы размножаются бесполым путем с интервалом деления клеток 40 минут. [14] [16] [17] [18]
Виды сальмонеллы ведут преимущественно образ жизни, связанный с хозяином, но было обнаружено, что бактерии способны сохраняться в ванной комнате в течение нескольких недель после заражения и часто выделяются из источников воды, которые действуют как резервуары бактерий и могут способствовать облегчению передачи инфекции между хозяевами. [31] Сальмонелла известна своей способностью переносить высыхание и может сохраняться годами в сухой среде и пищевых продуктах. [32]
Бактерии не уничтожаются при замораживании. [33] [34] но ультрафиолетовый свет и тепло ускоряют их разрушение. Они погибают после нагревания до 55 °C (131 °F) в течение 90 минут или до 60 °C (140 °F) в течение 12 минут. [35] хотя, если их инокулировать веществами с высоким содержанием жира и высокой жидкостью, такими как арахисовое масло, они приобретают термостойкость и могут выдерживать температуру до 90 ° C (194 ° F) в течение 30 минут. [36] Для защиты от заражения сальмонеллой рекомендуется нагревать пищу до внутренней температуры 75 °C (167 °F). [37] [38]
Виды сальмонелл можно обнаружить в пищеварительном тракте человека и животных, особенно рептилий. Сальмонелла на коже рептилий или амфибий может передаваться людям, которые имеют дело с животными. [39] Еда и вода также могут быть заражены бактериями, если они вступают в контакт с фекалиями инфицированных людей или животных. [40]
Номенклатура [ править ]
Первоначально каждый «вид» сальмонеллы был назван в соответствии с клиническими соображениями, например Salmonella typhi-murium (мышиный брюшной тиф), S. cholerae-suis (свиная холера). После того, как для многих видов было признано отсутствие специфичности хозяина, новые штаммы получили видовые названия в зависимости от места, в котором новый штамм был изолирован. [41]
В 1987 году Ле Минор и Попофф использовали молекулярные открытия, чтобы доказать, что сальмонелла состоит только из одного вида, S. enterica , превратив прежние названия «видов» в серотипы . [42] В 1989 году Ривз и др. предположил, что серотип V должен оставаться отдельным видом, возродив название S. bongori . [43] Таким образом, сложилась текущая (к 2005 г.) номенклатура с шестью признанными подвидами S. enterica : enterica (серотип I), salamae (серотип II), arizonae (IIIa), diarizonae (IIIb), houtenae (IV) и indica. (VI). [3] [44] [45] [46] Поскольку специалисты по инфекционным заболеваниям не знакомы с новой номенклатурой, традиционная номенклатура остается распространенной. [ нужна ссылка ]
Серотип или серовар — это классификация сальмонелл , основанная на антигенах, которые представляет организм. Схема классификации Кауфмана -Уайта отличает серологические разновидности друг от друга. Серотипы обычно объединяются в группы подвидов после рода и вида, при этом серотипы/серовары пишутся с заглавной буквы, но не курсивом: примером является Salmonella enterica серовар Typhimurium. Более современные подходы к типированию и субтипированию сальмонелл включают методы на основе ДНК, такие как импульсный гель-электрофорез , многолокусный анализ VNTR , мультилокусное типирование последовательностей и методы на основе мультиплексной ПЦР . [47] [48]
В 2005 году был предложен третий вид, Salmonella subterranea , но, по данным Всемирной организации здравоохранения , указанная бактерия не принадлежит к роду Salmonella . [49] В 2016 году S. subterranea было предложено отнести к Atlantibacter subterranea , [50] но LPSN отвергает ее как недействительную публикацию , поскольку она была сделана за пределами IJSB и IJSEM. [51] GTDB и NCBI согласны с переназначением в 2016 году. [52] [53]
GTDB RS202 сообщает, что S. arizonae , S. diarizonae и S. houtenae должны быть отдельными видами. [54]
Патогенность [ править ]
сальмонелл Виды являются факультативными внутриклеточными патогенами . [6] Сальмонелла может проникать в различные типы клеток, включая эпителиальные клетки , М-клетки , макрофаги и дендритные клетки . [55] Будучи факультативным анаэробом , сальмонеллы используют кислород для производства аденозинтрифосфата (АТФ) в аэробной среде (т. е. когда кислород доступен). Однако в анаэробной среде (т. е. когда кислород недоступен) сальмонелла производит АТФ путем ферментации ; путем замены одного или нескольких из четырех менее эффективных акцепторов электронов, чем кислород, в конце цепи переноса электронов: сульфат , нитрат , сера или фумарат . [56]
Большинство инфекций происходит в результате употребления в пищу продуктов питания, загрязненных фекалиями животных или людей, например, работниками общественного питания в коммерческих столовых. Серотипы сальмонелл можно разделить на две основные группы — брюшнотифозные и нетифоидные. К брюшнотифозным серотипам относятся Salmonella Typhi и Salmonella Paratyphi A, которые адаптированы к человеку и не встречаются у других животных. Нетифоидные серотипы встречаются чаще и обычно вызывают самопроизвольное желудочно-кишечное заболевание . Они могут заразить целый ряд животных и являются зоонозными , то есть могут передаваться от человека к другим животным. [57] [ нужна ссылка ]
Патогенность сальмонелл и взаимодействие с хозяином широко изучаются с 2010-х годов. Большинство важных вирулентных генов сальмонеллы закодированы в пяти островах патогенности, так называемых островах патогенности сальмонеллы (SPI). Они кодируются хромосомами и вносят значительный вклад во взаимодействие бактерии-хозяина. Другие признаки, такие как плазмиды, жгутики или белки, связанные с биопленками, могут способствовать инфекции. SPI характеризуются тем, что регулируются сложными и тонко настроенными регуляторными сетями, которые обеспечивают экспрессию генов только при наличии правильных стрессов окружающей среды. [58]
Молекулярное моделирование и анализ активного сайта гомолога SdiA, предполагаемого сенсора кворума патогенности Salmonella typhimurium, выявляет специфические закономерности связывания регуляторов транскрипции AHL. [59] Также известно, что ген вирулентности плазмиды сальмонеллы spvB усиливает вирулентность бактерий за счет ингибирования аутофагии. [60]
Тифоидная сальмонелла [ править ]
Брюшной тиф вызывается серотипами сальмонелл , строго адаптированными к человеку или высшим приматам, к ним относятся Salmonella Typhi , Paratyphi A, Paratyphi B и Paratyphi C. При системной форме заболевания сальмонеллы попадают через лимфатическую систему кишечника в крови больных (тифозная форма) и разносится к различным органам (печень, селезенка, почки) с образованием вторичных очагов (септическая форма). Эндотоксины сначала действуют на сосудистый и нервный аппарат, приводя к повышению проницаемости и снижению тонуса сосудов, нарушению терморегуляции, появлению рвоты и диареи. При тяжелых формах заболевания теряется достаточное количество жидкости и электролитов, что приводит к нарушению водно-солевого обмена, снижению объема циркулирующей крови и артериального давления, развитию гиповолемического шока . Также может развиться септический шок . Шок смешанного характера (с признаками как гиповолемического, так и септического шока) чаще встречается при тяжелом течении сальмонеллеза . В вследствие тяжелых случаях в результате поражения почек гипоксия и токсемия . [ нужна ссылка ]
Нетифоидная сальмонелла [ править ]
Неинвазивный [ править ]
Заражение нетифоидными серотипами сальмонелл обычно приводит к пищевым отравлениям . Заражение обычно происходит, когда человек употребляет в пищу продукты, содержащие высокую концентрацию [ нужны разъяснения ] бактерий. Младенцы и дети младшего возраста гораздо более восприимчивы к инфекции, что легко достигается при проглатывании небольшого количества [ нужны разъяснения ] бактерий. У младенцев возможно заражение при вдыхании пыли, содержащей бактерии. [ нужна ссылка ]
Микроорганизмы проникают через пищеварительный тракт и должны попадать в организм в больших количествах, чтобы вызвать заболевание у здоровых взрослых. Инфекция может начаться только после того, как живые сальмонеллы (а не только токсины, вырабатываемые сальмонеллами ) достигают желудочно-кишечного тракта. Часть микроорганизмов погибает в желудке, а выжившие попадают в тонкий кишечник и размножаются в тканях. Кислотность желудка ответственна за уничтожение большинства проглатываемых бактерий, но сальмонелла развила определенную степень толерантности к кислой среде, что позволяет определенной части проглоченных бактерий выжить. [61] Бактериальные колонии также могут попасть в слизь, вырабатываемую в пищеводе. К концу инкубационного периода близлежащие клетки-хозяева отравляются эндотоксинами, выделяемыми из погибших сальмонелл. Местной реакцией на эндотоксины являются энтерит и желудочно-кишечные расстройства. [ нужна ссылка ]
Известно около 2000 серотипов нетифоидной сальмонеллы , которые могут быть причиной до 1,4 миллиона заболеваний в Соединенных Штатах каждый год. К группе риска тяжелого заболевания относятся младенцы, пожилые люди, реципиенты трансплантатов и люди с ослабленным иммунитетом. [40]
Инвазивный [ править ]
В то время как в развитых странах нетифоидные серотипы проявляются главным образом в виде желудочно-кишечных заболеваний, в странах Африки к югу от Сахары эти серотипы могут создавать серьезную проблему при инфекциях кровотока и являются наиболее часто выделяемыми бактериями из крови людей с лихорадкой. В 2012 году сообщалось, что при инфекциях кровотока, вызванных нетифоидными сальмонеллами в Африке, уровень смертности составляет 20–25%. Большинство случаев инвазивной нетифоидной сальмонеллезной инфекции (iNTS) вызывают Salmonella enterica Typhimurium или Salmonella enterica Enteritidis. Новая форма Salmonella Typhimurium (ST313) возникла на юго-востоке Африканского континента 75 лет назад, а 18 лет спустя последовала вторая волна, вышедшая из Центральной Африки. Эта вторая волна iNTS, возможно, возникла в бассейне Конго и на раннем этапе развития приобрела ген, который сделал ее устойчивой к антибиотику хлорамфениколу . Это создало необходимость использования дорогих противомикробных препаратов в очень бедных районах Африки, что затруднило лечение. Считается, что повышенная распространенность iNTS в странах Африки к югу от Сахары по сравнению с другими регионами связана с тем, что большая часть африканского населения имеет некоторую степень иммуносупрессии или нарушений из-за бремени ВИЧ , малярия и недоедание, особенно у детей. Генетическая структура iNTS развивается в более сходную с брюшным тифом бактерию, способную эффективно распространяться по человеческому организму. Сообщается, что симптомы разнообразны, включая лихорадку, гепатоспленомегалию и респираторные симптомы, часто при отсутствии желудочно-кишечных симптомов. [62]
Эпидемиология [ править ]
Поскольку сальмонеллез считается спорадическим, от 60% до 80% случаев заражения сальмонеллезом остаются недиагностированными. [63] В марте 2010 года был завершен анализ данных, согласно которому уровень заболеваемости составил 1140 на 100 000 человеко-лет. Согласно тому же анализу, 93,8 миллиона случаев гастроэнтерита были вызваны инфекциями сальмонеллы. Для 5-го процентиля расчетное количество составило 61,8 миллиона случаев, а для 95-го процентиля расчетное количество составило 131,6 миллиона случаев. По оценкам, число смертей из-за сальмонеллы составило около 155 000 смертей. [64] В 2014 году в таких странах, как Болгария и Португалия, дети до 4 лет имели в 32 и 82 раза больше шансов заболеть сальмонеллезом соответственно. [65] Наиболее восприимчивы к инфекции: дети, беременные женщины, пожилые люди и люди с ослабленной иммунной системой. [66]
Факторы риска заражения сальмонеллой включают различные продукты. Мясо, такое как курица и свинина, может быть заражено. Различные овощи и ростки также могут быть заражены сальмонеллой. Наконец, различные обработанные пищевые продукты, такие как куриные наггетсы и пироги, также могут содержать эти бактерии. [67]
Успешные формы профилактики исходят от существующих организаций, таких как FDA , Министерство сельского хозяйства США и Служба безопасности и инспекции пищевых продуктов . Все эти организации создают стандарты и проводят проверки для обеспечения общественной безопасности в США. Например, агентство FSIS, работающее с Министерством сельского хозяйства США, имеет План действий по борьбе с сальмонеллой. Недавно, в феврале 2016 года, он получил обновленный двухлетний план. В планах представлены их достижения и стратегии по снижению заражения сальмонеллой. [68] Центры по контролю и профилактике заболеваний также предоставляют ценную информацию о профилактическом уходе, например, о том, как безопасно обращаться с сырыми продуктами и как правильно хранить эти продукты. В Европейском Союзе Европейское управление по безопасности пищевых продуктов разработало превентивные меры посредством управления рисками и оценки рисков. С 2005 по 2009 год EFSA разработало подход по снижению воздействия сальмонеллы . Их подход включал оценку рисков и управление рисками для домашней птицы, что привело к снижению случаев заражения вдвое. [69] В Латинской Америке была внедрена пероральная вакцина против сальмонеллы у домашней птицы, разработанная доктором Шерри Лейтон, которая предотвращает заражение птиц бактериями. [70]
Недавняя вспышка Salmonella Typhimurium была связана с шоколадом. [71]
Глобальный мониторинг [ править ]
В Германии необходимо сообщать об инфекциях пищевого происхождения. [72] С 1990 по 2016 год количество официально зарегистрированных случаев снизилось примерно с 200 000 до примерно 13 000 случаев. [73] По оценкам , в Соединенных Штатах сальмонеллой . ежегодно регистрируется около 1 200 000 случаев заражения [74] По оценкам исследования Всемирной организации здравоохранения, в 2000 году произошло 21 650 974 случая брюшного тифа, 216 510 из которых закончились смертью, а также 5 412 744 случая паратифа. [75]
заражения Молекулярные механизмы
Механизмы заражения тифоидных и нетифоидных серотипов различаются из-за их различных целей в организме и различных симптомов, которые они вызывают. Обе группы должны войти, преодолев барьер, созданный клеточной стенкой кишечника, но как только они преодолеют этот барьер, они используют разные стратегии, чтобы вызвать инфекцию. [ нужна ссылка ]
Переключиться на вирулентность [ править ]
Двигаясь к ткани-мишени в желудочно-кишечном тракте, сальмонеллы подвергаются воздействию желудочной кислоты, детергентной активности желчи в кишечнике, снижению поступления кислорода, конкурирующей нормальной кишечной флоре и, наконец, антимикробным пептидам, присутствующим в кишечнике. поверхность клеток, выстилающих стенку кишечника. Все это формирует стрессы, которые сальмонелла может ощущать и на которые реагирует, они образуют факторы вирулентности и, как таковые, регулируют переход от нормального роста в кишечнике к вирулентности . [76]
Переключение на вирулентность дает доступ к нише репликации внутри хозяина (например, человека) и может быть разделено на несколько этапов: [ нужна ссылка ]
- Подход, при котором они перемещаются к клетке-хозяину посредством кишечной перистальтики и посредством активного плавания через жгутики , проникают через слизистый барьер и располагаются близко к эпителию , выстилающему кишечник.
- Адгезия, при которой они прикрепляются к клетке-хозяину с помощью бактериальных адгезинов и системы секреции типа III .
- Инвазия, при которой сальмонелла проникает в клетку-хозяина (см. варианты механизмов ниже),
- Репликация, при которой бактерия может размножаться внутри клетки-хозяина.
- Распространение, при котором бактерия может распространиться на другие органы через клетки крови (если ей удалось обойти иммунную защиту). Альтернативно, бактерии могут вернуться в кишечник, повторно заселяя кишечную популяцию.
- Реинвазия ( вторичная инфекция , если теперь на системном участке) и дальнейшая репликация.
Механизмы проникновения [ править ]
Нетифоидные серотипы преимущественно проникают в М-клетки стенки кишечника посредством бактериально-опосредованного эндоцитоза — процесса, связанного с воспалением кишечника и диареей. Они также способны нарушать плотные соединения между клетками кишечной стенки, ухудшая способность клеток останавливать поток ионов , воды и иммунных клеток в кишечник и из него. Считается, что сочетание воспаления, вызванного бактериально-опосредованным эндоцитозом, и нарушение плотных контактов в значительной степени способствует индукции диареи. [77]
Сальмонеллы также способны преодолевать кишечный барьер посредством фагоцитоза и переноса CD18 -положительными иммунными клетками, что может быть ключевым механизмом возникновения брюшнотифозной сальмонеллезной инфекции. Считается, что это более скрытый способ прохождения кишечного барьера и, следовательно, может способствовать тому, что меньшее количество брюшнотифозных сальмонелл для заражения требуется , чем нетифоидных сальмонелл . [77] Клетки сальмонеллы способны проникать в макрофаги посредством макропиноцитоза . [78] Серотипы брюшного тифа могут использовать это для распространения по организму через систему мононуклеарных фагоцитов , сеть соединительной ткани, которая содержит иммунные клетки и окружает ткани, связанные с иммунной системой, по всему телу. [77]
Большая часть успеха сальмонеллы в возникновении инфекции объясняется двумя системами секреции типа III (T3SS), которые экспрессируются в разное время во время инфекции. T3SS-1 позволяет инъецировать бактериальные эффекторы в цитозоль хозяина. Эти эффекторы T3SS-1 стимулируют образование мембранных складок, позволяющих захватывать сальмонеллу клетками нефагоцитарными . Сальмонелла также находится в мембраносвязанном отсеке, называемом вакуолью, содержащей сальмонеллу (SCV). Подкисление SCV приводит к экспрессии T3SS-2. Секреция эффекторов T3SS-2 сальмонеллой необходима для ее эффективного выживания в цитозоле хозяина и развития системного заболевания. [77] Кроме того, оба T3SS участвуют в колонизации кишечника, индукции кишечных воспалительных реакций и диареи. Эти системы содержат множество генов, которые должны работать совместно для заражения. [ нужна ссылка ]
Токсин AvrA, инъецируемый системой секреции SPI1 типа III S. Typhimurium, ингибирует врожденную иммунную систему благодаря своей активности серин / треонин -ацетилтрансферазы и требует связывания с эукариотических клеток-мишеней фитиновой кислотой (IP6). [79] Это делает хозяина более восприимчивым к инфекции. [ нужна ссылка ]
Клинические симптомы [ править ]
Известно, что сальмонеллез может вызывать боли в спине или спондилез . Он может проявляться пятью клиническими проявлениями: инфекция желудочно-кишечного тракта, кишечная лихорадка, бактериемия, местная инфекция и хроническое резервуарное состояние. Первоначальными симптомами являются, среди прочего, неспецифическая лихорадка, слабость и миалгия. В состоянии бактериемии он может распространиться на любые части тела, что вызывает локализованную инфекцию или образование абсцессов. Формами локализованной сальмонеллезной инфекции являются артрит, инфекция мочевыводящих путей, инфекция центральной нервной системы, инфекция костей, инфекция мягких тканей и др. [80] Инфекция может оставаться в латентной форме в течение длительного времени, а при ухудшении функции ретикулярных эндотелиальных клеток она может активироваться и, как следствие, вторично индуцировать распространение инфекции в кости через несколько месяцев или несколько лет после острого сальмонеллеза. [80]
в 2018 году Исследование Имперского колледжа Лондона также показывает, как сальмонелла разрушает определенные ветви иммунной системы (например, 3 из 5 белков NF-kappaB ), используя семейство эффекторов цинковых металлопротеиназ , оставляя другие нетронутыми. [81] Сообщалось также о сальмонеллезном абсцессе щитовидной железы. [82]
Устойчивость к окислительному взрыву [ править ]
Отличительной чертой патогенеза сальмонеллы является способность бактерии выживать и размножаться внутри фагоцитов . Фагоциты производят агенты, повреждающие ДНК, такие как оксид азота и радикалы кислорода , в качестве защиты от патогенов. Таким образом, виды сальмонелл должны столкнуться с атакой молекул, которые нарушают целостность генома. Бухмайер и др. [83] показали, что мутанты S. enterica, лишенные функции белка RecA или RecBC, очень чувствительны к окислительным соединениям, синтезируемым макрофагами, и, кроме того, эти результаты указывают на то, что для успешной системной инфекции S. enterica требуется RecA- и RecBC-опосредованное рекомбинационное восстановление повреждений ДНК. [83] [84]
Адаптация хоста [ править ]
S. enterica через некоторые из своих серотипов, таких как Typhimurium и Enteritidis, демонстрирует признаки способности инфицировать несколько различных видов млекопитающих-хозяев, в то время как другие серотипы, такие как Typhi, по-видимому, ограничены лишь несколькими хозяевами. [85] Некоторые из способов сальмонеллы серотипов адаптации к своим хозяевам включают потерю генетического материала и мутации. У более сложных видов млекопитающих иммунные системы , включающие патоген-специфические иммунные реакции, нацелены на серовары сальмонеллы посредством связывания антител с такими структурами, как жгутики. Из-за потери генетического материала, который кодирует формирование жгутика, сальмонелла хозяина может уклоняться от иммунной системы . [86] mgtC Лидерная РНК из гена вирулентности бактерий (оперон mgtCBR) снижает выработку флагеллина во время инфекции путем прямого спаривания оснований с мРНК гена fljB , кодирующего флагеллин, и способствует деградации. [87] В исследовании Киселы и др. Было обнаружено , что более патогенные серовары S. enterica имеют определенные общие адгезины, которые развились в результате конвергентной эволюции. [88] Это означает, что, поскольку эти штаммы сальмонеллы подвергались воздействию схожих условий, таких как иммунная система, схожие структуры развивались отдельно, чтобы свести на нет эти схожие, более развитые защитные механизмы у хозяев. Тем не менее, остается много вопросов о том, как сальмонелла развилась во множество различных типов, но сальмонелла, возможно, эволюционировала через несколько фаз. Как Баумлер и др. предположили, что сальмонелла, скорее всего, эволюционировала посредством горизонтального переноса генов , образования новых сероваров за счет дополнительных островов патогенности и путем аппроксимации своего происхождения. [89] Таким образом, сальмонелла могла эволюционировать во множество различных серотипов, получая генетическую информацию от разных патогенных бактерий. Наличие нескольких островов патогенности в геноме разных серотипов подтверждает эту теорию. [89]
Сальмонелла св. У Ньюпорта есть признаки адаптации к образу жизни, связанному с колонизацией растений, что может играть роль в его непропорциональной связи с болезнями пищевого происхождения, связанными с продуктами. Разнообразие функций, выбранных во время св. Сообщалось, что устойчивость Ньюпорта у томатов аналогична стойкости, выбранной для sv. Typhimurium от животных-хозяев. [90] Ген papA , уникальный для sv. Ньюпорт, способствует приспособленности штамма к томатам и имеет гомологи в геномах других энтеробактерий, которые способны колонизировать растения и животных-хозяев. [90]
Исследования [ править ]
Помимо своей важности в качестве патогенов, нетифоидные виды сальмонелл, такие как серовар S. enterica Typhimurium, обычно используются в качестве гомологов брюшнотифозных видов. Многие результаты можно передавать, и это снижает опасность для исследователя в случае заражения, но также и ограничено. Например, с помощью этой модели невозможно изучить конкретные токсины брюшного тифа. [91] Однако сильные исследовательские инструменты, такие как широко используемая модель гастроэнтерита кишечника у мышей , основаны на использовании Salmonella Typhimurium. [92]
В генетике . S. Typhimurium сыграл важную роль в разработке генетических инструментов, которые привели к пониманию фундаментальной физиологии бактерий Эти разработки стали возможны благодаря открытию первого генерализованного трансдуцирующего фага P22. [93] в С. Typhimurium, позволивший быстро и легко редактировать гены . В свою очередь, это сделало возможным генетический анализ тонкой структуры. Большое количество мутантов привело к пересмотру генетической номенклатуры бактерий. [94] Многие из применений транспозонов в качестве генетических инструментов, включая доставку транспозонов, мутагенез и конструирование хромосомных перестроек, также были разработаны S. в Тифимурий. Эти генетические инструменты также привели к созданию простого теста на канцерогены — теста Эймса. [95]
В качестве естественной альтернативы традиционным противомикробным препаратам фаги признаны высокоэффективными средствами борьбы с сальмонеллой и другими бактериями пищевого происхождения. [96]
Древняя ДНК [ править ]
Геномы S. enterica были реконструированы по человеческим останкам возрастом до 6500 лет по всей Западной Евразии, что свидетельствует о географически широко распространенных инфекциях системного S. enterica в доисторические времена и о возможной роли процесса неолитизации в эволюции адаптации хозяина. [97] [98] Дополнительные реконструированные геномы из колониальной Мексики позволяют предположить, что S. enterica является причиной коколизтли , эпидемии в Новой Испании 16-го века . [99]
См. также [ править ]
- 1984 Биотеррористическая атака Раджниши
- Великая катастрофа в Хормеле 1998 года
- Вспышка сальмонеллеза в США в 2008 г.
- Американская ассоциация общественного здравоохранения против Бутца
- Висмутсульфитный агар
- Пищевые тест-полоски
- Взаимодействие хозяина и патогена
- Список вспышек болезней пищевого происхождения
- 2008–2009 гг. Сальмонеллез, переносимый арахисом.
- Яйцо округа Райт
- заказ XLD
Ссылки [ править ]
- ^ « Сальмонелла » . Таксономия NCBI . Бетесда, Мэриленд: Национальный центр биотехнологической информации . Проверено 27 января 2019 г.
- ^ Су Л.Х., Чиу Ч. (2007). «Сальмонелла: клиническое значение и эволюция номенклатуры». Медицинский журнал Чанг Гунг . 30 (3): 210–219. ПМИД 17760271 .
- ↑ Перейти обратно: Перейти обратно: а б Райан член парламента, О'Двайер Дж., Адли CC (2017). «Оценка комплексной номенклатуры клинически и ветеринарно значимого возбудителя сальмонеллы » . БиоМед Исследования Интернэшнл . 2017 : 3782182. doi : 10.1155/2017/3782182 . ПМЦ 5429938 . ПМИД 28540296 .
- ^ Галь-Мор О, Бойл ЕС, Грассл Г.А. (2014). «Один вид, разные заболевания: как и почему различаются брюшнотифозные и нетифозные серовары Salmonella enterica» . Границы микробиологии . 5 : 391. дои : 10.3389/fmicb.2014.00391 . ПМК 4120697 . ПМИД 25136336 .
- ↑ Перейти обратно: Перейти обратно: а б Фабрега А., Вила Дж (апрель 2013 г.). «Навыки серовара Salmonella enterica Typhimurium добиться успеха у хозяина: вирулентность и регуляция» . Обзоры клинической микробиологии . 26 (2): 308–341. дои : 10.1128/CMR.00066-12 . ПМЦ 3623383 . ПМИД 23554419 .
- ↑ Перейти обратно: Перейти обратно: а б Янч Дж., Чиккабалли Д., Хенсель М. (март 2011 г.). «Клеточные аспекты иммунитета к внутриклеточной Salmonella enterica». Иммунологические обзоры . 240 (1): 185–195. дои : 10.1111/j.1600-065X.2010.00981.x . ПМИД 21349094 . S2CID 19344119 .
- ^ Райан И.К.Дж., Рэй К.Г., ред. (2004). Медицинская микробиология Шерриса (4-е изд.). МакГроу Хилл. стр. 362–8. ISBN 978-0-8385-8529-0 .
- ^ Бреннер Ф.В., Вильяр Р.Г., Ангуло Ф.Дж., Токс Р., Сваминатан Б. (июль 2000 г.). «Номенклатура сальмонелл» . Журнал клинической микробиологии . 38 (7): 2465–2467. дои : 10.1128/JCM.38.7.2465-2467.2000 . ПМЦ 86943 . ПМИД 10878026 .
- ^ Гиллеспи, Стивен Х., Хоки, Питер М., ред. (2006). Принципы и практика клинической бактериологии (2-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-470-01796-8 .
- ^ Окоро К.К., Кингсли Р.А., Коннор Т.Р., Харрис С.Р., Парри К.М., Аль-Машхадани М.Н., Кариуки С., Мсефула К.Л., Гордон М.А., де Пинна Э., Уэйн Дж., Хейдерман Р.С., Обаро С., Алонсо П.Л., Мандомандо I, МакЛеннан К.А. , Тапиа, доктор медицинских наук, Левин М.М., Теннант С.М., Паркхилл Дж., Дуган Дж. (ноябрь 2012 г.). «Внутриконтинентальное распространение инвазивных для человека патовариантов Salmonella Typhimurium в Африке к югу от Сахары» . Природная генетика . 44 (11): 1215–1221. дои : 10.1038/ng.2423 . ПМК 3491877 . ПМИД 23023330 .
- ^ Эберт CJ (1 июля 1880 г.). «Организмы в органах при брюшном тифе». Архив патологической анатомии и физиологии и клинической медицины (на немецком языке). 81 (1): 58–74. дои : 10.1007/BF01995472 . S2CID 6979333 .
- ^ Харди А. (август 1999 г.). «Еда, гигиена и лаборатория. Краткая история пищевых отравлений в Великобритании, около 1850-1950 годов». Социальная история медицины . 12 (2): 293–311. дои : 10.1093/шм/12.2.293 . ПМИД 11623930 .
- ^ «FDA/CFSAN — Справочное руководство по безопасности пищевых продуктов от А до Я — Сальмонелла» . FDA – Центр безопасности пищевых продуктов и прикладного питания . 3 июля 2008 г. Архивировано из оригинала 2 марта 2009 г. Проверено 14 февраля 2009 г.
- ↑ Перейти обратно: Перейти обратно: а б Хейманн Д.А., Алкамо И.Э., Хейманн Д.Л. (2006). Сальмонелла . Филадельфия: Издательство Chelsea House. ISBN 978-0-7910-8500-4 . Проверено 31 июля 2015 г.
- ^ МакЭвин Э. «Аткинсон, Нэнси (1910–1999)» . Австралийский биографический словарь . Канберра: Национальный центр биографии Австралийского национального университета . ISBN 978-0-522-84459-7 . ISSN 1833-7538 . OCLC 70677943 . Проверено 27 октября 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б «Профиль серотипов изолятов сальмонеллы из продуктов мяса и птицы, с января 1998 г. по декабрь 2014 г.» (PDF) .
- ↑ Перейти обратно: Перейти обратно: а б «Этапы расследования вспышек пищевого происхождения» . 09.11.2018.
- ↑ Перейти обратно: Перейти обратно: а б Юн КБ, Сон БиДжей, Шин МЮ, Лим ХК, Юн ЮХ, Чон ДЮ, Ха Х, Ян СИ, Ким Дж.Б. (июнь 2017 г.). «Схемы устойчивости к антибиотикам и серотипы видов сальмонеллы , выделенных в Чолла-Намдо в Корее» . Осонг Общественное здравоохранение и перспективы исследований . 8 (3): 211–219. дои : 10.24171/j.phrp.2017.8.3.08 . ПМЦ 5525558 . ПМИД 28781944 .
- ^ Луо Ю, Хуан С, Е Дж, Октавия С, Ван Х, Данбар С.А., Цзинь Д, Тан ЮВ, Лан Р (07 сентября 2020 г.). «Сравнение анализа серотипирования сальмонеллы xMAP с традиционным серотипированием и разрешением несоответствий с помощью полногеномного секвенирования» . Границы клеточной и инфекционной микробиологии . 10 : 452. дои : 10.3389/fcimb.2020.00452 . ПМЦ 7504902 . ПМИД 33014887 .
- ^ Наир С., Патель В., Хики Т., Магуайр С., Грейг Д.Р., Ли В., Годбол Дж., Грант К., Чаттэуэй Массачусетс (август 2019 г.). Ледебур Н.А. (ред.). «ПЦР в реальном времени для дифференциации брюшнотифозных и нетифоидных сальмонелл » . Журнал клинической микробиологии . 57 (8): e00167–19. дои : 10.1128/JCM.00167-19 . ПМК 6663909 . ПМИД 31167843 .
- ^ Кларк М.А., Барретт Э.Л. (июнь 1987 г.). «Ген phs и продукция сероводорода Salmonella typhimurium» . Журнал бактериологии . 169 (6): 2391–2397. дои : 10.1128/jb.169.6.2391-2397.1987 . ПМК 212072 . ПМИД 3108233 .
- ^ «Стандарты Великобритании для микробиологических исследований: изменение фазы сальмонеллы » (PDF) . Стандарты Великобритании по микробиологическим исследованиям : 8–10. 8 января 2015 г. Архивировано (PDF) из оригинала 9 октября 2022 г. Проверено 2 августа 2015 г.
- ^ Снайдер Дж.В., Атлас РМ (2006). Справочник по средам для клинической микробиологии . ЦРК Пресс. п. 374. ИСБН 978-0-8493-3795-6 .
- ^ Альварес Дж., Сота М., Виванко А.Б., Пералес И., Цистерна Р., Рементерия А., Гарайзар Дж. (апрель 2004 г.). «Разработка метода мультиплексной ПЦР для выявления и эпидемиологического типирования сальмонелл в клинических образцах человека» . Журнал клинической микробиологии . 42 (4): 1734–1738. doi : 10.1128/JCM.42.4.1734-1738.2004 . ПМЦ 387595 . ПМИД 15071035 .
- ^ Хурфар Дж., Аренс П., Родстрем П. (сентябрь 2000 г.). «Автоматическая 5'-нуклеазная ПЦР для идентификации Salmonella enterica» . Журнал клинической микробиологии . 38 (9): 3429–3435. дои : 10.1128/JCM.38.9.3429-3435.2000 . ПМЦ 87399 . ПМИД 10970396 .
- ^ Домингес С.А., Шаффнер Д.В. (декабрь 2008 г.). «Моделирование роста сальмонеллы в сырой птице, хранившейся в аэробных условиях» . Журнал защиты пищевых продуктов . 71 (12): 2429–2435. дои : 10.4315/0362-028x-71.12.2429 . ПМИД 19244895 .
- ^ Пин C, Авенданьо-Перес Г., Кошани-Кунико Е, Гомес Н., Гунадакич А., Нычас Г.Дж., Скандамис П., Баркер Г. (март 2011 г.). «Моделирование концентрации сальмонеллы во всей цепочке поставок свинины с учетом роста и выживания в условиях колебаний температуры, pH и a(w)». Международный журнал пищевой микробиологии . 145 (Приложение 1): С96-102. doi : 10.1016/j.ijfoodmicro.2010.09.025 . ПМИД 20951457 .
- ^ Пан В., Шаффнер Д.В. (август 2010 г.). «Моделирование роста сальмонеллы в разрезанных красных круглых помидорах в зависимости от температуры» . Журнал защиты пищевых продуктов . 73 (8): 1502–1505. дои : 10.4315/0362-028X-73.8.1502 . ПМИД 20819361 .
- ^ Ли Д., Фридрих Л.М., Данилюк М.Д., Харрис Л.Дж., Шаффнер Д.В. (июнь 2013 г.). «Разработка и обоснование математической модели роста возбудителей болезней в разрезанных дынях» . Журнал защиты пищевых продуктов . 76 (6): 953–958. дои : 10.4315/0362-028X.JFP-12-398 . ПМИД 23726189 .
- ^ Ли Д. «Разработка и проверка математической модели роста сальмонеллы в дыне» . Архивировано из оригинала 20 ноября 2013 года.
- ^ Уинфилд, доктор медицинских наук, Гройсман, Е.А. (июль 2003 г.). «Роль среды, не являющейся хозяином, в образе жизни сальмонелл и кишечной палочки» . Прикладная и экологическая микробиология . 69 (7): 3687–3694. Бибкод : 2003ApEnM..69.3687W . дои : 10.1128/aem.69.7.3687-3694.2003 . ПМК 165204 . ПМИД 12839733 .
- ^ Мандал РК, Квон ЮМ (8 сентября 2017 г.). «Глобальный скрининг генов Salmonella enterica Serovar Typhimurium на выживание при высыхании» . Границы микробиологии . 8 (1723): 1723. doi : 10.3389/fmicb.2017.01723 . ПМК 5596212 . ПМИД 28943871 .
- ^ Сорреллс К.М., Спек М.Л., Уоррен Дж.А. (январь 1970 г.). «Патогенность Salmonella Gallinarum после метаболического повреждения при замораживании» . Прикладная микробиология . 19 (1): 39–43. дои : 10.1128/АЕМ.19.1.39-43.1970 . ПМК 376605 . ПМИД 5461164 .
Различия в смертности между полностью непострадавшими и преимущественно травмированными популяциями были небольшими и постоянными (уровень 5%) с гипотезой об отсутствии различий.
- ^ Бешат Л.Р., Хитон Э.К. (июнь 1975 г.). «Выживаемость сальмонеллы на орехах пекан под влиянием условий обработки и хранения» . Прикладная микробиология . 29 (6): 795–801. дои : 10.1128/АЕМ.29.6.795-801.1975 . ПМК 187082 . ПМИД 1098573 .
Небольшое снижение жизнеспособной популяции трех видов было отмечено на инокулированных половинках пекана, хранившихся при -18, -7 и 5°C в течение 32 недель.
- ^ Гудфеллоу С.Дж., Браун В.Л. (август 1978 г.). «Судьба сальмонеллы, инокулированной в говядину для приготовления пищи» . Журнал защиты пищевых продуктов . 41 (8): 598–605. дои : 10.4315/0362-028x-41.8.598 . ПМИД 30795117 .
- ^ Ма Л., Чжан Г., Гернер-Шмидт П., Мантрипрагада В., Изеок И., Дойл, член парламента (август 2009 г.). «Термическая инактивация сальмонеллы в арахисовом масле» . Журнал защиты пищевых продуктов . 72 (8): 1596–1601. дои : 10.4315/0362-028x-72.8.1596 . ПМИД 19722389 .
- ^ Партнерство по образованию в области безопасности пищевых продуктов (PFSE) Боритесь с BAC! Базовая брошюра , заархивированная 31 августа 2013 г. в Wayback Machine .
- ^ Министерства сельского хозяйства США. Таблица внутренних температур приготовления пищи Архивировано 3 мая 2012 г. в Wayback Machine . У Министерства сельского хозяйства США есть другие ресурсы, доступные на странице «Безопасное обращение с пищевыми продуктами» , заархивировано 5 июня 2013 г. на Wayback Machine странице информационного бюллетеня . См. также Национальный центр домашнего хранения продуктов питания .
- ^ «Рептилии, земноводные и сальмонеллы» . Центры по контролю и профилактике заболеваний . Министерство здравоохранения и социальных служб США. 25 ноября 2013 года . Проверено 3 августа 2013 г.
- ↑ Перейти обратно: Перейти обратно: а б Голдрик Б.А. (март 2003 г.). «Пищевые заболевания». Американский журнал медсестер . 103 (3): 105–106. дои : 10.1097/00000446-200303000-00043 . ПМИД 12635640 .
- ^ Ф. Кауфманн: Die Bacteriologie der Salmonella-Gruppe. Мунксгаард, Копенгаген, 1941 год.
- ^ Ле Минор Л, Попов М.Ю. (1987). «Запрос мнения: Обозначение Salmonella enterica sp. nov., nom. rev., как типа и единственного вида рода Salmonella » . Межд. Дж. Сист. Бактериол . 37 (4): 465–468. дои : 10.1099/00207713-37-4-465 .
- ^ Ривз М.В., Эвинс Г.М., Хейба А.А., Пликайтис Б.Д., Фармер Дж.Дж. (февраль 1989 г.). «Клональная природа Salmonella typhi и ее генетическое родство с другими сальмонеллами, как показано с помощью мультилокусного ферментативного электрофореза, и предложение Salmonella bongori comb. nov» . Журнал клинической микробиологии . 27 (2): 313–320. doi : 10.1128/JCM.27.2.313-320.1989 . ПМЦ 267299 . ПМИД 2915026 .
- ^ Янда Дж.М., Эбботт С.Л. (2006). «Энтеробактерии», АСМ Пресс.
- ^ Судебная комиссия Международного комитета по систематике прокариот (январь 2005 г.). «Типовым видом рода Salmonella Lignieres 1900 является Salmonella enterica (ex Kauffmann and Edwards 1952) Le Minor и Popoff 1987, с типовым штаммом LT2T и сохранением эпитета enterica в Salmonella enterica по сравнению со всеми более ранними эпитетами, которые могут быть применены к Мнение этого вида 80» . Международный журнал систематической и эволюционной микробиологии . 55 (Часть 1): 519–520. дои : 10.1099/ijs.0.63579-0 . ПМИД 15653929 .
- ^ Тиндалл Б.Дж., Гримон П.А., Гэррити ГМ, Эзеби Дж.П. (январь 2005 г.). «Номенклатура и систематика рода Salmonella» . Международный журнал систематической и эволюционной микробиологии . 55 (Часть 1): 521–524. дои : 10.1099/ijs.0.63580-0 . ПМИД 15653930 .
- ^ Порволлик С, изд. (2011). Сальмонелла: от генома к функции . Кайстер Академик Пресс . ISBN 978-1-904455-73-8 .
- ^ Ахтман М. , Уэйн Дж., Вейл Ф.К., Наир С., Чжоу З., Сангал В., Крауланд М.Г., Хейл Дж.Л., Харботтл Х., Уэсбек А., Дуган Г. , Харрисон Л.Х., Брисс С. (2012). «Типирование мультилокусных последовательностей как замена серотипирования Salmonella enterica» . ПЛОС Патогены . 8 (6): e1002776. дои : 10.1371/journal.ppat.1002776 . ПМК 3380943 . ПМИД 22737074 .

- ^ Гримонт, Пенсильвания, Ксавье Вейл Ф (2007). Антигенные формулы сероваров сальмонеллы (PDF) (9-е изд.). Институт Пастера, Париж, Франция: Сотрудничающий центр ВОЗ по справочным данным и исследованиям сальмонеллы. п. 7. Архивировано (PDF) из оригинала 9 октября 2022 г. Проверено 26 августа 2015 г.
- ^ Хата Х, Натори Т, Мизуно Т, Канадзава И, Эльдесуки И, Хаяси М, Мията М, Фукунага Х, Оджи С, Хосояма А, Аоно Е, Ямадзоэ А, Цучикане К, Фудзита Н, Эзаки Т (май 2016 г.). «Филогенетика семейства Enterobacteriaceae и предложение реклассифицировать Escherichia hermannii и Salmonella subterranea как Atlantibacter hermannii и Atlantibacter subterranea gen. nov.,comb. nov.» . Микробиология и иммунология . 60 (5): 303–311. дои : 10.1111/1348-0421.12374 . ПМИД 26970508 . S2CID 32594451 .
- ^ «Виды: Atlantibacter subterranea» . lpsn.dsmz.de.
- ^ «GTDB — Дерево у g__Atlantibacter» . gtdb.ecogenomic.org .
- ^ «Таксономический браузер (Атлантибактер)» . www.ncbi.nlm.nih.gov .
- ^ «GTDB — Дерево при г__Сальмонелле» . gtdb.ecogenomic.org .
- ^ ЛаРок Д.Л., Чаудхари А., Миллер С.И. (апрель 2015 г.). «Взаимодействие сальмонелл с процессами хозяина» . Обзоры природы. Микробиология . 13 (4): 191–205. дои : 10.1038/nrmicro3420 . ПМК 5074537 . ПМИД 25749450 .
- ^ Гарай П., Гнанадхас Д.П., Чакравортти Д. (июль 2012 г.). «Серовары Salmonella enterica Typhimurium и Typhi как модельные организмы: выявление парадигмы взаимодействия хозяина и патогена» . Вирулентность . 3 (4): 377–388. дои : 10.4161/viru.21087 . ПМЦ 3478240 . ПМИД 22722237 .
- ^ «В чем разница между нетифоидными сальмонеллами и S typhi или S paratyphi?» . www.medscape.com . Проверено 28 сентября 2021 г.
- ^ Фабрега А., Вила Дж (апрель 2013 г.). «Навыки серовара Salmonella enterica Typhimurium добиться успеха у хозяина: вирулентность и регуляция» . Обзоры клинической микробиологии . 26 (2): 308–341. дои : 10.1128/CMR.00066-12 . ПМЦ 3623383 . ПМИД 23554419 .
- ^ Гнанендра С., Анусуя С., Натараджан Дж. (октябрь 2012 г.). «Молекулярное моделирование и анализ активного сайта гомолога SdiA, предполагаемого сенсора кворума патогенности Salmonella typhimurium, выявляет специфические закономерности связывания регуляторов транскрипции AHL». Журнал молекулярного моделирования . 18 (10): 4709–4719. дои : 10.1007/s00894-012-1469-1 . ПМИД 22660944 . S2CID 25177221 .
- ^ Ли Ю, Ван Т, Гао С, Сюй ГМ, Ню Х, Хуан Р, Ву Си (февраль 2016 г.). «Ген вирулентности плазмиды сальмонеллы spvB повышает вирулентность бактерий путем ингибирования аутофагии на модели инфекции рыбок данио». Иммунология рыб и моллюсков . 49 : 252–259. дои : 10.1016/j.fsi.2015.12.033 . ПМИД 26723267 .
- ^ Гарсиа-дель Портильо Ф., Фостер Дж.В., Финли Б.Б. (октябрь 1993 г.). «Роль генов реакции толерантности к кислоте в вирулентности Salmonella typhimurium» . Инфекция и иммунитет . 61 (10): 4489–4492. дои : 10.1128/IAI.61.10.4489-4492.1993 . ПМК 281185 . ПМИД 8406841 .
- ^ Физи Н.А., Дуган Дж., Кингсли Р.А., Хейдерман Р.С., Гордон М.А. (июнь 2012 г.). «Инвазивная нетифоидная сальмонеллезная болезнь: новая и забытая тропическая болезнь в Африке» . Ланцет . 379 (9835): 2489–2499. дои : 10.1016/S0140-6736(11)61752-2 . ПМК 3402672 . ПМИД 22587967 .
- ^ «Сальмонелла (нетифоидная)» . www.who.int . Проверено 5 декабря 2019 г.
- ^ Майович С.Е., Мусто Дж., Скаллан Э., Ангуло Ф.Дж., Кирк М., О'Брайен С.Дж., Джонс Т.Ф., Фазиль А., Хоекстра Р.М. (март 2010 г.). «Глобальное бремя нетифоидного сальмонеллезного гастроэнтерита» . Клинические инфекционные болезни . 50 (6): 882–889. дои : 10.1086/650733 . ПМИД 20158401 .
- ^ «Сальмонеллез – Ежегодный эпидемиологический отчет 2016 г. [данные за 2014 г.]» . Европейский центр профилактики и контроля заболеваний . 31 января 2016 г. Проверено 5 декабря 2019 г.
- ^ «Узнайте факты о сальмонелле!» . Центр ветеринарной медицины, FDA. 06.06.2019.
- ^ «Профилактика | Общая информация | Сальмонелла | CDC» . www.cdc.gov . 06.03.2019 . Проверено 5 декабря 2019 г.
- ^ «Сальмонелла» . fsis.usda.gov .
- ^ «Сальмонелла» . Европейское агентство по безопасности пищевых продуктов . Проверено 5 декабря 2019 г.
- ^ «Содержимое не найдено» . 3 марта 2022 г.
- ^ Самарасекера У (июль 2022 г.). «Вспышка Salmonella Typhimurium связана с шоколадом». «Ланцет». Инфекционные болезни . 22 (7): 947. doi : 10.1016/S1473-3099(22)00351-6 . ПМИД 35636448 . S2CID 249136373 .
- ^ § 6 и § 7 закона Германии о профилактике инфекционных заболеваний, Infektionsschutzgesetz.
- ^ «Количество заболеваний сальмонеллезом, ежегодно регистрируемых в Германии до 2016 г.» . Статистика.
- ^ Сальмонелла . Центры по контролю и профилактике заболеваний
- ^ Крамп Дж.А., Луби С.П., Минц Э.Д. (май 2004 г.). «Глобальное бремя брюшного тифа» . Бюллетень Всемирной организации здравоохранения . 82 (5): 346–353. ПМЦ 2622843 . ПМИД 15298225 .
- ^ Рычлик И., Барроу П.А. (ноябрь 2005 г.). «Управление стрессом, вызванным сальмонеллой, и его значение для поведения во время кишечной колонизации и инфекции» . Обзоры микробиологии FEMS . 29 (5): 1021–1040. дои : 10.1016/j.femsre.2005.03.005 . ПМИД 16023758 .
- ↑ Перейти обратно: Перейти обратно: а б с д Харага А., Олсон М.Б., Миллер С.И. (январь 2008 г.). «Взаимодействие сальмонелл с клетками-хозяевами». Обзоры природы. Микробиология . 6 (1): 53–66. дои : 10.1038/nrmicro1788 . ПМИД 18026123 . S2CID 2365666 .
- ^ Керр MC, Ван Дж.Т., Кастро Н.А., Гамильтон Н.А., Таун Л., Браун Д.Л., Менье Ф.А., Браун Н.Ф., Стоу Дж.Л., Тисдейл Р.Д. (апрель 2010 г.). «Ингибирование киназы PtdIns(5) PIKfyve нарушает внутриклеточную репликацию сальмонеллы» . Журнал ЭМБО . 29 (8): 1331–1347. дои : 10.1038/emboj.2010.28 . ПМЦ 2868569 . ПМИД 20300065 .
- ^ Миттал Р., Пик-Чью С.Ю., Шаде Р.С., Валлис Ю., МакМахон Х.Т. (июнь 2010 г.). «Ацетилтрансферазная активность бактериального токсина YopJ иерсинии активируется эукариотической клеткой-хозяином инозитолгексакисфосфатом» . Журнал биологической химии . 285 (26): 19927–19934. дои : 10.1074/jbc.M110.126581 . ПМЦ 2888404 . ПМИД 20430892 .
- ↑ Перейти обратно: Перейти обратно: а б Чой Ю.С., Чо В.Дж., Юн Ш., Ли С.И., Пак Ш.Х., Пак Дж.К., Чан Э.Х., Шин ХИ (декабрь 2010 г.). «Случай боли в спине, вызванной сальмонеллезным спондилитом. Отчет о случае» . Корейский журнал анестезиологии . 59 (Дополнение): S233–S237. дои : 10.4097/kjae.2010.59.S.S233 . ПМК 3030045 . ПМИД 21286449 .
- ^ «Бактериальный белок имитирует ДНК, саботируя защиту клеток: исследование раскрывает подробности инфекций сальмонеллы» . Sciencedaily .
- ^ Сахли М., Хеммауи Б., Эррами Н., Бенариба Ф. (январь 2022 г.). «Сальмонеллезный абсцесс щитовидной железы» . Европейские анналы оториноларингологии . 139 (1): 51–52. дои : 10.1016/j.anorl.2021.05.004 . ПМИД 34053890 . S2CID 235256840 .
- ↑ Перейти обратно: Перейти обратно: а б Бухмайер Н.А., Липпс С.Дж., Со М.И., Хеффрон Ф. (март 1993 г.). «Мутанты Salmonella typhimurium с дефицитом рекомбинации авирулентны и чувствительны к окислительному взрыву макрофагов». Молекулярная микробиология . 7 (6): 933–936. дои : 10.1111/j.1365-2958.1993.tb01184.x . ПМИД 8387147 . S2CID 25127127 .
- ^ Кано Д.А., Пуччарелли М.Г., Гарсиа-дель Портильо Ф., Касадесус Х. (январь 2002 г.). «Роль пути рекомбинации RecBCD в вирулентности сальмонеллы» . Журнал бактериологии . 184 (2): 592–595. дои : 10.1128/jb.184.2.592-595.2002 . ПМК 139588 . ПМИД 11751841 .
- ^ Томсон Н.Р., Клейтон Д.Д., Виндхорст Д., Верникос Г., Дэвидсон С., Черчер С., Куэйл М.А., Стивенс М., Джонс М.А., Уотсон М., Бэррон А., Лейтон А., Пикард Д., Кингсли Р.А., Бигнелл А., Кларк Л., Харрис Б. , Ормонд Д., Абделла З., Брукс К., Черевач И., Чиллингворт Т., Вудворд Дж., Норберчак Х., Лорд А., Эроусмит С., Джагелс К., Мул С., Мангалл К., Сандерс М., Уайтхед С., Чабалгоити Дж.А., Маскелл Д., Хамфри Т., Робертс М., Барроу П.А., Дуган Г., Паркхилл Дж. (октябрь 2008 г.). «Сравнительный анализ генома Salmonella Enteritidis PT4 и Salmonella Gallinarum 287/91 дает представление об эволюционных путях адаптации хозяина» . Геномные исследования . 18 (10): 1624–1637. дои : 10.1101/гр.077404.108 . ПМЦ 2556274 . ПМИД 18583645 .
- ^ ден Баккер ХК, Морено Свитт А.И., Говони Г., Каммингс К.А., Раньери М.Л., Дегориция Л., Хельцер К., Родригес-Ривера Л.Д., Браун С., Больчакова Е., Фуртадо М.Р., Видманн М. (август 2011 г.). «Секвенирование генома выявляет диверсификацию содержания факторов вирулентности и возможную адаптацию хозяина в отдельных субпопуляциях Salmonella enterica» . БМК Геномика . 12 : 425. дои : 10.1186/1471-2164-12-425 . ПМК 3176500 . ПМИД 21859443 .
- ^ Чой Э, Хан Ю, Чо Ю.Дж., Нам Д., Ли Э.Дж. (сентябрь 2017 г.). « Транс -действующая лидерная РНК из гена вирулентности сальмонеллы » . Труды Национальной академии наук Соединенных Штатов Америки . 114 (38): 10232–10237. Бибкод : 2017PNAS..11410232C . дои : 10.1073/pnas.1705437114 . ПМЦ 5617274 . ПМИД 28874555 .
- ^ Кисела Д.И., Чаттопадьяй С., Либби С.Дж., Карлинси Дж.Э., Фанг ФК, Чеснокова В, Крамер Дж.Дж., Бесхлебная В., Самадпур М., Гржимайло К., Угорски М., Ланкау Э.В., Маки Р.И., Клегг С., Сокуренко Е.В. (2012). «Эволюция вирулентности Salmonella enterica посредством точечных мутаций фимбриального адгезина» . ПЛОС Патогены . 8 (6): e1002733. дои : 10.1371/journal.ppat.1002733 . ПМК 3369946 . ПМИД 22685400 .
- ↑ Перейти обратно: Перейти обратно: а б Боймлер А.Дж., Цолис Р.М., Фихт Т.А., Адамс Л.Г. (октябрь 1998 г.). «Эволюция адаптации хозяина у Salmonella enterica» . Инфекция и иммунитет . 66 (10): 4579–4587. дои : 10.1128/IAI.66.10.4579-4587.1998 . ПМЦ 108564 . ПМИД 9746553 .
- ↑ Перейти обратно: Перейти обратно: а б де Мораес М.Х., Сото Э.Б., Салас Гонсалес И., Десаи П., Чу В., Порволлик С., Макклелланд М., Теплицкий М. (2018). «Полногеномный сравнительный функциональный анализ выявляет адаптацию Salmonella sv. Newport к образу жизни, связанному с колонизацией растений» . Границы микробиологии . 9 : 877. дои : 10.3389/fmicb.2018.00877 . ПМЦ 5968271 . ПМИД 29867794 .
- ^ Джонсон Р., Милона Э., Франкель Г. (сентябрь 2018 г.). «Тифоидная сальмонелла: отличительные факторы вирулентности и патогенез» . Клеточная микробиология . 20 (9): e12939. дои : 10.1111/cmi.12939 . ПМИД 30030897 . S2CID 51705854 .
- ^ Хапфельмайер С., Хардт В.Д. (октябрь 2005 г.). «Мышиная модель энтероколита, вызванного S. typhimurium». Тенденции в микробиологии . 13 (10): 497–503. дои : 10.1016/j.tim.2005.08.008 . ПМИД 16140013 .
- ^ Зиндер Н.Д., Ледерберг Дж. (ноябрь 1952 г.). «Генетический обмен при сальмонелле» . Журнал бактериологии . 64 (5): 679–699. дои : 10.1128/JB.64.5.679-699.1952 . ПМК 169409 . ПМИД 12999698 .
- ^ Демерек М., Адельберг Э.А., Кларк А.Дж., Хартман П.Е. (июль 1966 г.). «Предложение о единой номенклатуре в генетике бактерий» . Генетика . 54 (1): 61–76. дои : 10.1093/генетика/54.1.61 . ПМЦ 1211113 . ПМИД 5961488 .
- ^ Эймс Б.Н., Макканн Дж., Ямасаки Э. (декабрь 1975 г.). «Методы обнаружения канцерогенов и мутагенов с помощью теста на мутагенность сальмонеллы/микросом млекопитающих». Мутационные исследования . 31 (6): 347–364. дои : 10.1016/0165-1161(75)90046-1 . ПМИД 768755 .
- ^ Гэ Х, Линь С, Сюй Ю, Ху М, Сюй Z, Гэн С, Цзяо X, Чэнь X (июнь 2022 г.). «Фаг для борьбы с сальмонеллой у домашней птицы и уменьшения биопленок». Ветеринарная микробиология . 269 : 109432. doi : 10.1016/j.vetmic.2022.109432 . ПМИД 35489296 . S2CID 248195843 .
- ^ Ки ФМ, Пост С, Эскивель-Гомес ЛР, Хюблер Р, Спироу М.А., Нейман Г.Ю., Фуртвенглер А, Сабин С., Бурри М., Виссготт А., Ланкапалли А.К., Воген О.Дж., Мейер М., Нагель С., Тухбатова Р., Хохлов А., Чижевский А, Хансен С, Белинский А.Б., Калмыков А., Канторович А.Р., Маслов В.Е., Штокхаммер П.В., Вай С., Заваттаро М., Рига А., Карамелли Д., Скитс Р., Беккет Дж., Градоли М.Г., Стеури Н., Хафнер А., Рамштайн М. , Зибке И., Лёш С., Эрдал Ю.С., Алихан Н.Ф., Чжоу З., Ахтман М., Бос К., Райнхольд С., Хаак В., Кюнерт Д., Хербиг А., Краузе Дж. (март 2020 г.). «Появление адаптированной к человеку Salmonella enterica связано с процессом неолитизации» . Экология и эволюция природы . 4 (3): 324–333. Бибкод : 2020NatEE...4..324K . дои : 10.1038/s41559-020-1106-9 . ПМК 7186082 . ПМИД 32094538 .
- ^ Чжоу З, Лундстрём И, Тран-Дьен А, Дюшен С, Алихан Н.Ф., Сержант М.Дж., Лангридж Г., Фотакис А.К., Наир С., Стенойен Х.К., Хамре СС, Касьенс С., Кристоферсен А., Айва С., Томсон Н.Р., Вейл Ф.С., Хо С.Ю., Гилберт М.Т., Ахтман М. (август 2018 г.). «Пангеномный анализ древней и современной Salmonella enterica демонстрирует геномную стабильность инвазивной линии Para C на протяжении тысячелетий» . Современная биология . 28 (15): 2420–2428.e10. Бибкод : 2018CBio...28E2420Z . дои : 10.1016/j.cub.2018.05.058 . ПМК 6089836 . ПМИД 30033331 .
- ^ Вогене О.Дж., Хербиг А., Кампана М.Г., Роблес Гарсия Н.М., Вариннер С., Сабин С., Спиру М.А., Андрадес Валтуэна А., Хусон Д., Туросс Н., Бос К.И., Краузе Дж. (март 2018 г.). «Геномы Salmonella enterica жертв крупной эпидемии шестнадцатого века в Мексике». Экология и эволюция природы . 2 (3): 520–528. Бибкод : 2018NatEE...2..520В . дои : 10.1038/s41559-017-0446-6 . ПМИД 29335577 . S2CID 3358440 .
Внешние ссылки [ править ]
- Сведения о сальмонелле предоставлены Службой безопасности пищевых продуктов и инспекции Министерства сельского хозяйства США.
- Геномы сальмонеллы и соответствующая информация в PATRIC , ресурсном центре биоинформатики, финансируемом NIAID.
- Вопросы и ответы о коммерческих и институциональных методах дезинфекции. Архивировано 29 июня 2017 г. на Wayback Machine.
- Сальмонелла как новый патоген по данным МФСА
- Примечания по сальмонелл номенклатуре
- о подвижности сальмонеллы Видео
- Птичья сальмонелла. Архивировано 9 декабря 2011 г. в Wayback Machine.
- Обзор сальмонеллеза — Ветеринарное руководство компании Merck