НФ-кБ
Ядерный фактор каппа-легкая цепь-энхансер активированных В-клеток NF -κB ) представляет собой семейство белковых комплексов транскрипционных факторов, которые контролируют транскрипцию ДНК ( , выработку цитокинов и выживаемость клеток. NF-κB обнаруживается почти во всех типах клеток животных и участвует в клеточных реакциях на такие стимулы, как стресс, цитокины , свободные радикалы , тяжелые металлы , ультрафиолетовое облучение , окисленные ЛПНП , а также бактериальные или вирусные антигены . [ 2 ] [ 3 ] [ 4 ] [ 6 ] [ 7 ] NF-κB играет ключевую роль в регуляции иммунного ответа на инфекцию. Неправильная регуляция NF-κB связана с раком, воспалительными и аутоиммунными заболеваниями , септическим шоком , вирусной инфекцией и неправильным развитием иммунитета. NF-κB также участвует в процессах синаптической пластичности и памяти. [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ] [ 13 ]
Открытие
[ редактировать ]NF-κB был обнаружен Ранджаном Сеном в лаборатории нобелевского лауреата Дэвида Балтимора посредством его взаимодействия с последовательностью из 11 пар оснований в иммуноглобулина легкой цепи энхансере в В-клетках . [ 14 ] Более поздняя работа Александра Полторака и Бруно Леметра на мышах и дрозофилы плодовых мушках установила, что Toll-подобные рецепторы являются универсально консервативными активаторами передачи сигналов NF-κB. Эти работы в конечном итоге способствовали присуждению Нобелевской премии Брюсу Бойтлеру и Жюлю А. Хоффману , которые были главными исследователями этих исследований. [ 15 ] [ 16 ] [ 17 ]
Структура
[ редактировать ]Все белки семейства NF-κB имеют общий домен Rel-гомологии на N-конце . Подсемейство белков NF-κB, включая RelA, RelB и c-Rel, имеют домен трансактивации на своих С-концах . Напротив, белки NF-κB1 и NF-κB2 синтезируются как крупные предшественники, p105 и p100, которые подвергаются процессингу с образованием зрелых субъединиц p50 и p52 соответственно. Процессинг р105 и р100 опосредован путем убиквитин / протеасома и включает селективную деградацию их С-концевой области, содержащей анкириновые повторы . В то время как генерация р52 из р100 представляет собой строго регулируемый процесс, р50 образуется в результате конститутивного процессинга р105. [ 18 ] [ 19 ] Белки p50 и p52 не обладают внутренней способностью активировать транскрипцию, и поэтому было предложено действовать как репрессоры транскрипции при связывании элементов κB в виде гомодимеров. [ 20 ] [ 21 ] Действительно, это затрудняет интерпретацию исследований с нокаутом p105, в которых генетические манипуляции удаляют IκB (полноразмерный p105) и вероятный репрессор (гомодимеры p50) в дополнение к активатору транскрипции (гетеродимер RelA-p50).
Члены
[ редактировать ]Члены семейства NF-κB имеют структурную гомологию с ретровирусным онкопротеином v-Rel, в результате чего их классифицируют как белки NF-κB/Rel. [ 2 ]
В семействе NF-κB млекопитающих пять белков: [ 22 ]
| Сорт | Белок | Псевдонимы | Ген |
|---|---|---|---|
| я | NF-κB1 | р105 → стр50 | НФКБ1 |
| NF-κB2 | стр100 → стр52 | НФКБ2 | |
| II | РелА | стр.65 | РЕЛА |
| РелБ | РЭЛБ | ||
| c-Rel | РЭЛ |
Белки NF-κB/Rel можно разделить на два класса, которые имеют общие структурные особенности: [ 1 ]

Ниже приведены пять членов семейства NF-κB человека:
| НФКБ1 | |||
|---|---|---|---|
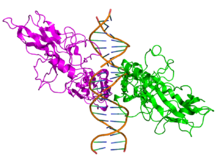
 Вид сверху кристаллографической структуры ( PDB : 1SVC ) гомодимера белка NFKB1 (зеленый и пурпурный), связанного с ДНК (коричневый). | |||
| Идентификаторы | |||
| Символ | НФКБ1 | ||
| ген NCBI | 4790 | ||
| HGNC | 7794 | ||
| МОЙ БОГ | 164011 | ||
| RefSeq | НМ_003998 | ||
| ЮниПрот | P19838 | ||
| Другие данные | |||
| Локус | Хр. 4 q24 | ||
| |||
| РЕЛА | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | РЕЛА | ||
| ген NCBI | 5970 | ||
| HGNC | 9955 | ||
| МОЙ БОГ | 164014 | ||
| RefSeq | НМ_021975 | ||
| ЮниПрот | Q04206 | ||
| Другие данные | |||
| Локус | Хр. 11 q13 | ||
| |||
| НФКБ2 | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | НФКБ2 | ||
| ген NCBI | 4791 | ||
| HGNC | 7795 | ||
| МОЙ БОГ | 164012 | ||
| RefSeq | НМ_002502 | ||
| ЮниПрот | Q00653 | ||
| Другие данные | |||
| Локус | Хр. 10 q24 | ||
| |||
| РЭЛБ | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | РЭЛБ | ||
| ген NCBI | 5971 | ||
| HGNC | 9956 | ||
| МОЙ БОГ | 604758 | ||
| RefSeq | НМ_006509 | ||
| ЮниПрот | Q01201 | ||
| Другие данные | |||
| Локус | Хр. 19 q13.2-19q13 | ||
| |||
| РЭЛ | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | РЭЛ | ||
| ген NCBI | 5966 | ||
| HGNC | 9954 | ||
| МОЙ БОГ | 164910 | ||
| RefSeq | НМ_002908 | ||
| ЮниПрот | Q04864 | ||
| Другие данные | |||
| Локус | Хр. 2 п13-п12 | ||
| |||
Распространение и эволюция видов
[ редактировать ]Помимо млекопитающих, NF-κB обнаружен и у ряда простейших животных. [ 23 ] К ним относятся книдарии (такие как морские анемоны , кораллы и гидры ), пориферы (губки), одноклеточные эукариоты, включая Capsaspora owczarzaki и хоанофлагелляты, а также насекомые (такие как мотыльки , комары и плодовые мухи ). Секвенирование геномов комаров A. aegypti и A. gambiae и плодовой мушки D. melanogaster позволило провести сравнительные генетические и эволюционные исследования NF-κB. У этих видов насекомых активация NF-κB запускается путем Toll (который развился независимо у насекомых и млекопитающих) и путем Imd (иммунодефицит). [ 24 ]
Сигнализация
[ редактировать ]Эффект активации
[ редактировать ]
NF-κB имеет решающее значение в регуляции клеточных ответов, поскольку он принадлежит к категории «быстродействующих» первичных факторов транскрипции, т. е. факторов транскрипции, которые присутствуют в клетках в неактивном состоянии и не требуют синтеза нового белка для активации. (другие члены этого семейства включают факторы транскрипции, такие как c-Jun , STAT и ядерные рецепторы гормонов ). Это позволяет NF-κB первым реагировать на вредные клеточные раздражители. Известные индукторы активности NF-κB сильно варьируются и включают активные формы кислорода ( АФК ), фактор некроза опухоли альфа ( ФНОα ), интерлейкин 1-бета ( ИЛ-1β ), бактериальные липополисахариды ( ЛПС ), изопротеренол , кокаин , эндотелин-1. и ионизирующее излучение . [ 26 ]
Подавление NF-κB цитотоксичности фактора некроза опухоли (апоптоза) происходит за счет индукции антиоксидантных ферментов и устойчивого подавления N-концевых киназ c-Jun (JNK). [ 27 ]
Рецептор-активатор NF-κB ( RANK ), который представляет собой тип TNFR , является центральным активатором NF-κB. Остеопротегерин (OPG), который является рецептора-ловушки гомологом для лиганда RANK ( RANKL ), ингибирует RANK путем связывания с RANKL, и, таким образом, остеопротегерин активно участвует в регуляции активации NF-κB. [ 28 ]
Многие бактериальные продукты и стимуляция широкого спектра рецепторов клеточной поверхности приводят к активации NF-κB и довольно быстрым изменениям в экспрессии генов. [ 2 ] Идентификация Toll-подобных рецепторов (TLR) как специфических молекул распознавания образов и открытие того, что стимуляция TLRs приводит к активации NF-κB, улучшили наше понимание того, как различные патогены активируют NF-κB. Например, исследования выявили TLR4 как рецептор компонента ЛПС грамотрицательных бактерий . [ 29 ] TLRs являются ключевыми регуляторами как врожденного, так и адаптивного иммунного ответа. [ 30 ]
В отличие от RelA, RelB и c-Rel, субъединицы NF-κB p50 и p52 не содержат доменов трансактивации в своих C-концевых половинах. Тем не менее, члены NF-κB p50 и p52 играют решающую роль в модуляции специфичности функции NF-κB. Хотя гомодимеры р50 и р52, как правило, являются репрессорами транскрипции сайта κB, как р50, так и р52 участвуют в трансактивации гена-мишени, образуя гетеродимеры с RelA, RelB или c-Rel. [ 31 ] Кроме того, гомодимеры р50 и р52 также связываются с ядерным белком Bcl-3 , и такие комплексы могут действовать как активаторы транскрипции. [ 32 ] [ 33 ] [ 34 ]
Торможение
[ редактировать ]В нестимулированных клетках димеры NF-κB изолируются в цитоплазме с помощью семейства ингибиторов, называемых IκB (ингибитор κB), которые представляют собой белки, содержащие множественные копии последовательности, называемой анкириновыми повторами . Благодаря своим доменам анкириновых повторов белки IκB маскируют сигналы ядерной локализации (NLS) белков NF-κB и удерживают их изолированными в неактивном состоянии в цитоплазме. [ 35 ]
IκB представляют собой семейство родственных белков, которые имеют N-концевой регуляторный домен, за которым следуют шесть или более анкириновых повторов и домен PEST вблизи их С-конца. Хотя семейство IκB состоит из IκBα , IκBβ , IκBε и Bcl-3 , наиболее изученным и основным белком IκB является IκBα. Благодаря наличию анкириновых повторов в их С-концевых половинах р105 и р100 также функционируют как белки IκB. С-концевая половина p100, которую часто называют IκBδ, также действует как ингибитор. [ 36 ] [ 37 ] Деградация IκBδ в ответ на стимулы развития, такие как те, которые трансдуцируются через LTβR , потенцирует активацию димера NF-κB по NIK-зависимому неканоническому пути. [ 36 ] [ 38 ]
Процесс активации (канонический/классический)
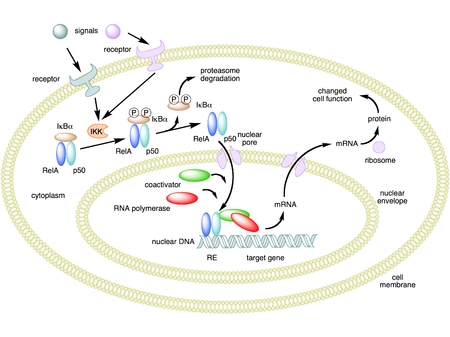
[ редактировать ]Активация NF-κB инициируется сигнал-индуцированной деградацией белков IκB. Это происходит в первую очередь за счет активации киназы, называемой киназой IκB (IKK). IKK состоит из гетеродимера каталитических субъединиц IKKα и IKKβ и «главного» регуляторного белка, называемого NEMO (незаменимый модулятор NF-κB) или IKKγ. При активации сигналами, обычно поступающими извне клетки, киназа IκB фосфорилирует два остатка серина, расположенные в регуляторном домене IκB. При фосфорилировании этих серинов (например, серинов 32 и 36 в человеческом IκBα) белки IκB модифицируются посредством процесса, называемого убиквитинированием , который затем приводит к их разрушению под действием клеточной структуры, называемой протеасомой .
При деградации IκB комплекс NF-κB затем высвобождается и проникает в ядро, где он может «включить» экспрессию специфических генов, которые имеют поблизости сайты связывания ДНК для NF-κB. Активация этих генов с помощью NF-κB затем приводит к заданному физиологическому ответу, например, воспалительному или иммунному ответу, ответу выживания клеток или клеточной пролиферации. Транслокацию NF-κB в ядро можно обнаружить иммуноцитохимически и измерить с помощью лазерной сканирующей цитометрии. [ 39 ] NF-κB включает экспрессию собственного репрессора IκBα. Вновь синтезированный IκBα затем повторно ингибирует NF-κB и, таким образом, образует петлю автоматической обратной связи, что приводит к колебаниям уровней активности NF-κB. [ 40 ] Кроме того, некоторые вирусы, включая вирус СПИДа ВИЧ, имеют сайты связывания для NF-κB, который контролирует экспрессию вирусных генов, которые, в свою очередь, способствуют репликации вируса или вирусной патогенности. В случае ВИЧ-1 активация NF-κB может, по крайней мере частично, участвовать в активации вируса из латентного, неактивного состояния. [ 41 ] YopP — фактор, секретируемый Yersinia pestis , возбудителем чумы, который предотвращает убиквитинирование IκB. Это заставляет этот патоген эффективно ингибировать путь NF-κB и, таким образом, блокировать иммунный ответ человека, инфицированного иерсинией. [ 42 ]
Ингибиторы активности NF-κB
[ редактировать ]Что касается известных белковых ингибиторов активности NF-κB, одним из них является IFRD1 , который подавляет активность NF-κB p65 за счет усиления HDAC -опосредованного деацетилирования субъединицы p65 по лизину 310, способствуя рекрутированию HDAC3 в p65. Фактически IFRD1 образует тримолекулярные комплексы с p65 и HDAC3. [ 43 ] [ 44 ]
НАД + -зависимая протеиндеацетилаза и фактор долголетия SIRT1 ингибируют экспрессию гена NF-κB путем деацетилирования субъединицы RelA/p65 NF-κB по лизину 310. [ 45 ]
Неканонический/альтернативный путь
[ редактировать ]Отобранный набор стимулов клеточной дифференцировки или развития, таких как β-рецептор лимфотоксина (LTβR), BAFF или RANKL , активирует неканонический путь NF-κB, чтобы индуцировать димер NF-κB/RelB:p52 в ядре. В этом пути активация киназы, индуцирующей NF-κB (NIK), при лигировании рецептора приводила к фосфорилированию и последующему протеосомному процессингу белка-предшественника NF-κB2 p100 в зрелую субъединицу p52 IKK1/IKKa-зависимым образом. Затем p52 димеризуется с RelB, проявляя активность связывания ядерной ДНК RelB:p52. RelB:p52 регулирует экспрессию гомеостатических лимфокинов, которые инструктируют лимфоидный органогенез и транспорт лимфоцитов во вторичных лимфоидных органах. [ 46 ] В отличие от канонической передачи сигналов, которая основана на опосредованной NEMO-IKK2 деградации IκBα, -β, -ε, неканоническая передача сигналов зависит от опосредованной NIK обработки p100 в p52. Учитывая различные правила, эти два пути считались независимыми друг от друга. Однако было обнаружено, что синтез компонентов неканонического пути, а именно RelB и p52, контролируется канонической передачей сигналов IKK2-IκB-RelA:p50. [ 47 ] Более того, генерация канонических и неканонических димеров, а именно RelA:p50 и RelB:p52, в клеточной среде механически взаимосвязана. [ 47 ] Эти анализы показывают, что интегрированная сеть системы NF-κB лежит в основе активации как RelA, так и димера, содержащего RelB, и что сбой в каноническом пути приведет к аберрантному клеточному ответу также через неканонический путь. Самое интригующее, недавнее исследование показало, что TNF-индуцированная каноническая передача сигналов подрывает неканоническую активность RelB:p52 в воспаленных лимфоидных тканях, ограничивая проникновение лимфоцитов. [ 48 ] Механистически TNF инактивирует NIK в LTβR-стимулированных клетках и индуцирует синтез мРНК Nfkb2 , кодирующей p100; вместе они мощно накапливали непроцессированный p100, что ослабляло активность RelB. Роль p100/ Nfkb2 в регулировании проникновения лимфоцитов в воспаленную лимфоидную ткань может иметь широкие физиологические последствия.
В дополнение к своей традиционной роли в лимфоидном органогенезе, неканонический путь NF-κB также напрямую усиливает воспалительные иммунные ответы на микробные патогены путем модуляции канонической передачи сигналов NF-κB. Было показано, что p100/ Nfkb2 опосредует стимул-селективные и специфичные для типа клеток перекрестные помехи между двумя путями NF-κB и что Nfkb2 -опосредованные перекрестные помехи защищают мышей от кишечных патогенов. [ 49 ] [ 50 ] С другой стороны, отсутствие регуляции, опосредованной p100, перемещает RelB под контроль TNF-индуцированной канонической передачи сигналов. Фактически, мутационная инактивация p100/ Nfkb2 при множественной миеломе позволила TNF индуцировать длительную активность RelB, которая придавала клеткам миеломы устойчивость к химиотерапевтическому препарату. [ 51 ]
В иммунитете
[ редактировать ]NF-κB является основным фактором транскрипции, который регулирует гены, ответственные как за врожденный , так и за адаптивный иммунный ответ . [ 52 ] При активации рецептора Т- или В-клеток NF-κB активируется посредством различных сигнальных компонентов. При лигировании Т-клеточного рецептора протеинкиназа Lck рекрутируется и фосфорилирует ITAM цитоплазматического хвоста CD3 . Затем ZAP70 рекрутируется в фосфорилированные ITAM и помогает рекрутировать LAT и PLC-γ , что вызывает активацию PKC . Посредством каскада событий фосфорилирования киназный комплекс активируется, и NF-κB может проникать в ядро, чтобы активировать гены, участвующие в развитии, созревании и пролиферации Т-клеток. [ 53 ]
В нервной системе
[ редактировать ]Помимо роли в обеспечении выживания клеток, исследования Марка Мэттсона и других показали, что NF-κB выполняет разнообразные функции в нервной системе, включая роль в пластичности , обучении и памяти. [ 54 ] Помимо стимулов, которые активируют NF-κB в других тканях, NF-κB в нервной системе может быть активирован факторами роста ( BDNF , NGF ) и синаптической передачей, такой как глутамат . [ 9 ] Все эти активаторы NF-κB в нервной системе сходятся на комплексе IKK и каноническом пути.
В последнее время наблюдается большой интерес к роли NF-κB в нервной системе. Текущие исследования показывают, что NF-κB важен для обучения и памяти у многих организмов, включая крабов. [ 11 ] [ 12 ] плодовые мушки, [ 55 ] и мыши. [ 9 ] [ 10 ] NF-κB может регулировать обучение и память, частично модулируя синаптическую пластичность. [ 8 ] [ 56 ] функция синапса, [ 55 ] [ 57 ] [ 58 ] а также регулируя рост дендритов [ 59 ] и дендритные шипики . [ 58 ]
Показано, что гены, имеющие сайты связывания NF-κB, имеют повышенную экспрессию после обучения. [ 10 ] что позволяет предположить, что транскрипционные мишени NF-κB в нервной системе важны для пластичности. Многие гены-мишени NF-κB, которые могут быть важны для пластичности и обучения, включают факторы роста (BDNF, NGF). [ 60 ] цитокины ( ФНО-альфа , ФНОФ ) [ 61 ] и киназы ( PKAc ). [ 56 ]
Несмотря на функциональные доказательства роли транскрипционных факторов Rel-семейства в нервной системе, до сих пор неясно, что неврологические эффекты NF-κB отражают транскрипционную активацию в нейронах. Большинство манипуляций и анализов выполняются в смешанной клеточной среде, обнаруженной in vivo, в культурах «нейрональных» клеток, которые содержат значительное количество глии, или в «нейрональных» клеточных линиях, полученных из опухолей. Когда трансфекции или другие манипуляции нацелены конкретно на нейроны, измеряемыми конечными точками обычно являются электрофизиология или другие параметры, далекие от транскрипции генов. Тщательные тесты NF-κB-зависимой транскрипции в высокоочищенных культурах нейронов обычно практически не обнаруживают активности NF-κB. [ 62 ] [ 63 ]
Некоторые из сообщений о NF-κB в нейронах, по-видимому, являются артефактом неспецифичности антител. [ 64 ] Конечно, артефакты клеточной культуры, например, удаление нейронов из-под влияния глии, также могут привести к ложным результатам. Но эта проблема решается, по крайней мере, в двух подходах к совместной культуре. Моерман и др. [ 65 ] использовали формат совместного культивирования, при котором нейроны и глия могли быть разделены после обработки для анализа EMSA , и они обнаружили, что NF-κB, индуцированный глутаматергическими стимулами, был ограничен глией (и, что интересно, только глией, которая находилась в присутствии нейронов в течение 48 часы). Те же исследователи исследовали эту проблему с помощью другого подхода, используя нейроны трансгенной репортерной мыши NF-κB, культивированные с глией дикого типа; глутаматергические стимулы снова не смогли активироваться в нейронах. [ 66 ] Некоторая часть ДНК-связывающей активности, отмечаемая при определенных условиях (особенно та, которая считается конститутивной), по-видимому, является результатом связывания Sp3 и Sp4 с подмножеством последовательностей энхансера κB в нейронах. [ 67 ] Эта активность фактически ингибируется глутаматом и другими условиями, которые повышают уровень внутринейронного кальция. В конечном счете роль NF-κB в нейронах остается неясной из-за сложности измерения транскрипции в клетках, одновременно идентифицируемых по типу. Конечно, на обучение и память могут влиять транскрипционные изменения в астроцитах и других глиальных элементах. И следует учитывать, что помимо прямой трансактивации генов могут существовать и другие механические эффекты NF-κB.
Клиническое значение
[ редактировать ]Рак
[ редактировать ]NF-κB широко используется эукариотическими клетками в качестве регулятора генов, контролирующих пролиферацию и выживание клеток. Таким образом, многие различные типы опухолей человека имеют неправильную регуляцию NF-κB: то есть NF-κB конститутивно активен. Активный NF-κB включает экспрессию генов, которые поддерживают пролиферацию клеток и защищают клетку от условий, которые в противном случае привели бы к ее гибели в результате апоптоза . При раке белки, которые контролируют передачу сигналов NF-κB, мутируют или экспрессируются аберрантно, что приводит к нарушению координации между злокачественной клеткой и остальным организмом. Это проявляется как при метастазировании, так и при неэффективной ликвидации опухоли иммунной системой. [ 68 ]
Нормальные клетки могут погибнуть, когда их удаляют из ткани, к которой они принадлежат, или когда их геном не может работать в гармонии с функцией ткани: эти события зависят от регуляции NF-κB по принципу обратной связи и терпят неудачу при раке. [ 69 ]
Дефекты NF-κB приводят к повышенной восприимчивости к апоптозу, что приводит к повышенной гибели клеток. Это связано с тем, что NF-κB регулирует антиапоптотические гены, особенно TRAF1 и TRAF2 , и, следовательно, отменяет активность ферментов семейства каспаз , которые играют центральную роль в большинстве процессов апоптоза. [ 70 ]
В опухолевых клетках активность NF-κB повышена, как, например, в 41% случаев рака носоглотки . [ 71 ] колоректальный рак , рак простаты и опухоли поджелудочной железы . Это происходит либо из-за мутаций в генах, кодирующих сами факторы транскрипции NF-κB, либо в генах, которые контролируют активность NF-κB (например, гены IκB); кроме того, некоторые опухолевые клетки секретируют факторы, которые вызывают активацию NF-κB. [ 72 ] [ 73 ] Блокирование NF-κB может привести к тому, что опухолевые клетки перестанут пролиферировать, погибнут или станут более чувствительными к действию противоопухолевых препаратов. [ 74 ] [ 75 ] Таким образом, NF-κB является предметом активных исследований фармацевтических компаний в качестве мишени для противораковой терапии. [ 76 ]
Однако, несмотря на то, что убедительные экспериментальные данные идентифицировали NF-κB как критический промотор онкогенеза, что создает веское обоснование для разработки противоопухолевой терапии, основанной на подавлении активности NF-κB, следует проявлять осторожность при рассмотрении анти-NF-κB. Активность -κB как широкая терапевтическая стратегия при лечении рака, поскольку данные также показали, что активность NF-κB повышает чувствительность опухолевых клеток к апоптозу и старению. Кроме того, было показано, что канонический NF-κB является активатором транскрипции Fas, а альтернативный NF-κB является репрессором транскрипции Fas. [ 77 ] Таким образом, NF-κB способствует Fas-опосредованному апоптозу в раковых клетках, и, таким образом, ингибирование NF-κB может подавлять Fas-опосредованный апоптоз, ухудшая подавление опухоли, опосредованное иммунными клетками хозяина.
Воспаление
[ редактировать ]Поскольку NF-κB контролирует многие гены, участвующие в воспалении, неудивительно, что NF-κB оказывается хронически активным при многих воспалительных заболеваниях, таких как воспалительные заболевания кишечника, артрит, сепсис, гастрит, астма, атеросклероз. [ 78 ] и другие. Однако важно отметить, что повышение уровня некоторых активаторов NF-κB, таких как остеопротегерин (ОПГ), связано с повышенной смертностью, особенно от сердечно-сосудистых заболеваний . [ 79 ] [ 80 ] Повышенный уровень NF-κB также связан с шизофренией . [ 81 ] Недавно активация NF-κB была предложена в качестве возможного молекулярного механизма катаболического воздействия сигаретного дыма на скелетные мышцы и саркопению . [ 82 ] Исследования показали, что во время воспаления функция клетки зависит от сигналов, которые она активирует в ответ на контакт с соседними клетками, а также от комбинаций гормонов, особенно цитокинов, которые действуют на нее через специфические рецепторы. [ 83 ] Фенотип клетки внутри ткани развивается посредством взаимной стимуляции сигналов обратной связи, которые координируют ее функцию с другими клетками; это особенно очевидно при перепрограммировании клеточной функции, когда ткань подвергается воспалению, поскольку клетки меняют свой фенотип и постепенно экспрессируют комбинации генов, которые подготавливают ткань к регенерации после устранения причины воспаления. [ 83 ] [ 84 ] Особенно важны реакции обратной связи, которые развиваются между резидентными клетками тканей и циркулирующими клетками иммунной системы. [ 84 ]
Верность ответов обратной связи между различными типами клеток и иммунной системой зависит от целостности механизмов, которые ограничивают диапазон генов, активируемых NF-κB, позволяя экспрессировать только те гены, которые способствуют эффективному иммунному ответу и, следовательно, полному восстановлению ткани. функционируют после разрешения воспаления. [ 84 ] При раке механизмы, регулирующие экспрессию генов в ответ на воспалительные стимулы, изменяются до такой степени, что клетка перестает связывать свое выживание с механизмами, которые координируют ее фенотип и ее функцию с остальной тканью. [ 69 ] Это часто проявляется в серьезном нарушении регуляции активности NF-κB, что позволяет раковым клеткам экспрессировать аномальные когорты генов-мишеней NF-κB. [ 85 ] В результате не только раковые клетки функционируют ненормально: клетки окружающей ткани меняют свою функцию и перестают исключительно поддерживать организм. Кроме того, некоторые типы клеток в микроокружении рака могут менять свои фенотипы, чтобы поддерживать рост рака. [ 86 ] [ 87 ] [ 88 ] Таким образом, воспаление — это процесс, проверяющий соответствие тканевых компонентов, поскольку процесс, ведущий к регенерации тканей, требует координации экспрессии генов между различными типами клеток. [ 83 ] [ 89 ]
НЕМО
[ редактировать ]Синдром дефицита NEMO — редкое генетическое заболевание, связанное с дефектом IKBKG , который, в свою очередь, активирует NF-κB. В основном оно поражает мужчин и имеет весьма разнообразный набор симптомов и прогнозов. [ 90 ]
Старение и ожирение
[ редактировать ]NF-κB все чаще экспрессируется при ожирении и старении. [ 91 ] что приводит к снижению уровня противовоспалительного, проаутофагического и антиинсулинового белка сиртуина 1 . NF-κB увеличивает уровни микроРНК миР -34a , которая ингибирует синтез никотинамидадениндинуклеотида (НАД) путем связывания с ее промоторной областью, [ 92 ] что приводит к снижению уровня сиртуина 1.
NF-κB и интерлейкин 1 альфа взаимно индуцируют друг друга в стареющих клетках по принципу положительной обратной связи , вызывая выработку факторов секреторного фенотипа, ассоциированного со старением (SASP). [ 93 ] , расщепляющий НАД, NF-κB и фермент CD38 также взаимно индуцируют друг друга. [ 94 ]
NF-κB является центральным компонентом клеточного ответа на повреждение. [ 95 ] NF-κB активируется в различных типах клеток, которые подвергаются нормальному или ускоренному старению . [ 95 ] Генетическое или фармакологическое ингибирование активации NF-κB может задержать возникновение многочисленных симптомов и патологий, связанных со старением. [ 95 ] Этот эффект можно частично объяснить тем, что снижение уровня NF-κB снижает выработку митохондриальных активных форм кислорода , которые могут повредить ДНК . [ 95 ]
Зависимость
[ редактировать ]NF-κB является одной из нескольких мишеней индуцируемой транскрипции ΔFosB , которая способствует развитию и поддержанию зависимости от стимула. [ 96 ] [ 97 ] [ 98 ] В хвостатой скорлупе индукция NF-κB связана с усилением локомоции, тогда как в прилежащем ядре индукция NF-κB усиливает положительный усиливающий эффект лекарства за счет сенсибилизации к вознаграждению . [ 97 ]
| Цель ген |
Цель выражение |
Нейронные эффекты | Поведенческие эффекты |
|---|---|---|---|
| с-Фос | ↓ | Молекулярный переключатель, обеспечивающий хроническое индукция ΔFosB [ примечание 1 ] |
– |
| динорфин | ↓ [ примечание 2 ] |
• Понижение уровня κ-опиоида. обратной связи | • Снижение отвращения к наркотикам. |
| НФ-кБ | ↑ | • Расширение NAcc. дендритных процессов • Воспалительная реакция NF-κB в NAcc • Воспалительная реакция NF-κB при ХП |
• Повышенное вознаграждение за лекарства • Повышенное вознаграждение за лекарства • Локомоторная сенсибилизация |
| ГлюР2 | ↑ | • Снижение чувствительности к глутамату. | • Повышенное вознаграждение за лекарства |
| CDK5 | ↑ | • GluR1 Фосфорилирование синаптического белка • Расширение NAcc. дендритных процессов |
Снижение вознаграждения за наркотики (чистый эффект) |
Нелекарственные ингибиторы
[ редактировать ]Было также показано, что многие натуральные продукты (включая антиоксиданты), обладающие противораковой и противовоспалительной активностью, ингибируют NF-κB. Существует спорный патент США (патент США 6 410 516). [ 100 ] это относится к открытию и использованию агентов, которые могут блокировать NF-κB, в терапевтических целях. Этот патент участвует в нескольких судебных процессах, в том числе «Ариад против Лилли» . Последняя работа Карин, [ 101 ] Бен-Нерия [ 102 ] и другие подчеркнули важность связи между NF-κB, воспалением и раком, а также подчеркнули ценность методов лечения, регулирующих активность NF-κB. [ 103 ]
Экстракты ряда трав и пищевых растений являются эффективными ингибиторами активации NF-κB in vitro. [ 104 ] нобилетин , флавоноид, выделенный из кожуры цитрусовых, ингибирует сигнальный путь NF-κB у мышей. Было показано, что [ 105 ] Было показано, что циркумспорозоитный белок Plasmodium falciparum является ингибитором NF-κB. [ 106 ] различные витанолиды Withania somnifera Аналогично, было обнаружено, что (Ashwagandha) оказывают ингибирующее действие на NF-κB за счет ингибирования опосредованной протеасомами деградации убиквитина IκBα. [ 107 ] [ 108 ]
Как мишень для наркотиков
[ редактировать ]Аберрантная активация NF-κB часто наблюдается при многих видах рака. Более того, подавление NF-κB ограничивает пролиферацию раковых клеток. Кроме того, NF-κB играет ключевую роль в воспалительной реакции. Следовательно, методы ингибирования передачи сигналов NF-κB имеют потенциальное терапевтическое применение при раке и воспалительных заболеваниях. [ 109 ] [ 110 ]
Как канонические, так и неканонические пути NF-κB требуют протеосомной деградации компонентов регуляторного пути для осуществления передачи сигналов NF-κB. Ингибитор протеосом вызванной NF-κB , бортезомиб в целом блокирует эту активность и одобрен для лечения лимфомы мантийных клеток, и множественной миеломы . [ 111 ] [ 112 ]
Открытие того, что активацию ядерной транслокации NF-κB можно отделить от усиления окислительного стресса. [ 113 ] открывает многообещающие возможности для разработки стратегий, направленных на ингибирование NF-κB.
Препарат деносумаб повышает минеральную плотность костной ткани и снижает частоту переломов у многих подгрупп пациентов путем ингибирования RANKL . RANKL действует через свой рецептор RANK , который, в свою очередь, способствует NF-κB, [ 114 ] RANKL обычно работает, обеспечивая дифференцировку остеокластов из моноцитов.
Дисульфирам , олмесартан и дитиокарбаматы могут ингибировать сигнальный каскад NF-κB. [ 115 ] Попытки разработать прямой ингибитор NF-κB были предприняты с использованием таких соединений, как (-)-DHMEQ, PBS-1086, IT-603 и IT-901. [ 116 ] [ 117 ] [ 118 ] (-)-DHMEQ и PBS-1086 являются необратимыми связующими для NF-κB, тогда как IT-603 и IT-901 являются обратимыми связующими. DHMEQ ковалентно связывается с Cys 38 р65. [ 119 ]
Утверждается, что противовоспалительные эффекты анатабина являются результатом модуляции активности NF-κB. [ 120 ] Однако в исследованиях, подтверждающих его пользу, используются аномально высокие дозы в миллимолярном диапазоне (аналогично внеклеточной концентрации калия), которые вряд ли могут быть достигнуты у людей.
BAY 11-7082 также был идентифицирован как препарат, который может ингибировать сигнальный каскад NF-κB. Он способен необратимо предотвращать фосфорилирование IKK-α, что приводит к подавлению активации NF-κB. [ 121 ]
Было показано, что введение BAY 11-7082 восстанавливает функциональность почек у крыс Спраг-Доули, страдающих диабетом, путем подавления окислительного стресса, регулируемого NF-κB. [ 122 ]
Исследования показали, что N-ацилэтаноламин, пальмитоилэтаноламид, способен PPAR-опосредованно ингибировать NF-κB. [ 123 ]
Биологическая цель игуратимода , препарата , продаваемого для лечения ревматоидного артрита в Японии и Китае, по состоянию на 2015 год была неизвестна, но основным механизмом действия, по-видимому, было предотвращение активации NF-κB. [ 124 ]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Другими словами, репрессия c-Fos позволяет ΔFosB быстрее накапливаться в шипиковых нейронах прилежащего ядра, поскольку в этом состоянии он избирательно индуцируется. [ 98 ]
- ^ В различных исследованиях ΔFosB участвовал как в повышении, так и в снижении экспрессии динорфина; [ 97 ] [ 99 ] эта запись в таблице отражает лишь снижение.
Ссылки
[ редактировать ]- ^ Jump up to: а б с Бьянкалана М., Натан Э., Ленардо М.Дж., Фершт А.Р. (сентябрь 2021 г.). «Обмен субъединицей Rel NF-κB в физиологическом масштабе времени» . Белковая наука . 30 (9): 1818–1832. дои : 10.1002/pro.4134 . ПМЦ 8376415 . ПМИД 34089216 .
- ^ Jump up to: а б с д и Гилмор Т.Д. (октябрь 2006 г.). «Введение в NF-kappaB: игроки, пути, перспективы» . Онкоген . 25 (51): 6680–6684. дои : 10.1038/sj.onc.1209954 . ПМИД 17072321 .
- ^ Jump up to: а б с Брасье АР (2006). «Регуляторная сеть NF-kappaB». Сердечно-сосудистая токсикология . 6 (2): 111–130. дои : 10.1385/CT:6:2:111 . ПМИД 17303919 . S2CID 19755135 .
- ^ Jump up to: а б с Перкинс Н.Д. (январь 2007 г.). «Интеграция клеточных сигнальных путей с функцией NF-kappaB и IKK». Обзоры природы. Молекулярно-клеточная биология . 8 (1): 49–62. дои : 10.1038/nrm2083 . ПМИД 17183360 . S2CID 24589510 .
- ^ Jump up to: а б Кончетти Дж., Уилсон С.Л. (сентябрь 2018 г.). «NFKB1 и рак: друг или враг?» . Клетки . 7 (9): 133. doi : 10.3390/cells7090133 . ПМК 6162711 . ПМИД 30205516 .
- ^ Гилмор Т.Д. (ноябрь 1999 г.). «Путь передачи сигнала Rel/NF-kappaB: введение» . Онкоген . 18 (49): 6842–6844. дои : 10.1038/sj.onc.1203237 . ПМИД 10602459 .
- ^ Тиан Б., Бразье А.Р. (2003). «Идентификация генной сети, зависимой от ядерного фактора каппа B» . Последние достижения в исследованиях гормонов . 58 : 95–130. дои : 10.1210/rp.58.1.95 . ПМИД 12795416 .
- ^ Jump up to: а б Альбенси, Британская Колумбия, член парламента от Мэттсона (февраль 2000 г.). «Доказательства участия TNF и NF-kappaB в синаптической пластичности гиппокампа». Синапс . 35 (2): 151–159. doi : 10.1002/(SICI)1098-2396(200002)35:2<151::AID-SYN8>3.0.CO;2-P . ПМИД 10611641 . S2CID 24215807 .
- ^ Jump up to: а б с Мефферт М.К., Чанг Дж.М., Вильтген Б.Дж., Фанселоу М.С., Балтимор Д. (октябрь 2003 г.). «NF-каппа B участвует в синаптической передаче сигналов и поведении» . Природная неврология . 6 (10): 1072–1078. дои : 10.1038/nn1110 . ПМИД 12947408 . S2CID 43284934 .
- ^ Jump up to: а б с Левенсон Дж.М., Чой С., Ли С.Ю., Цао Ю.А., Ан Х.Дж., Уорли К.С. и др. (апрель 2004 г.). «Биоинформатический анализ консолидации памяти выявил участие транскрипционного фактора c-rel» . Журнал неврологии . 24 (16): 3933–3943. doi : 10.1523/JNEUROSCI.5646-03.2004 . ПМК 6729420 . ПМИД 15102909 .
- ^ Jump up to: а б Фрейденталь Р., Локателли Ф., Эрмитт Г., Мальдонадо Х., Лафуркад С., Делоренци А. и др. (февраль 1998 г.). «Активность связывания ДНК, подобная каппа-В, усиливается после интервальной тренировки, которая вызывает долговременную память у краба Chasmagnathus». Письма по неврологии . 242 (3): 143–146. дои : 10.1016/S0304-3940(98)00059-7 . ПМИД 9530926 . S2CID 24577481 .
- ^ Jump up to: а б Мерло Э., Фрейденталь Р., Романо А. (2002). «Ингибитор киназы IkappaB сульфасалазин ухудшает долговременную память у краба Chasmagnathus». Нейронаука . 112 (1): 161–172. дои : 10.1016/S0306-4522(02)00049-0 . ПМИД 12044481 . S2CID 1403544 .
- ^ Пак HJ, Юн HS (март 2013 г.). «Ртуть индуцирует экспрессию циклооксигеназы-2 и индуцибельной синтазы оксида азота». Токсикология и промышленное здоровье . 29 (2): 169–174. дои : 10.1177/0748233711427048 . ПМИД 22080037 . S2CID 25343140 .
- ^ Сен Р., Балтимор Д. (август 1986 г.). «Множественные ядерные факторы взаимодействуют с последовательностями энхансера иммуноглобулина». Клетка . 46 (5): 705–716. дои : 10.1016/0092-8674(86)90346-6 . ПМИД 3091258 . S2CID 37832531 .
- ^ Полторак А., Хе Х, Смирнова И., Лю М.Ю., Ван Хаффель С., Ду Х. и др. (декабрь 1998 г.). «Дефектная передача сигналов LPS у мышей C3H/HeJ и C57BL/10ScCr: мутации в гене Tlr4». Наука . 282 (5396): 2085–2088. дои : 10.1126/science.282.5396.2085 . ПМИД 9851930 .
- ^ Леметр Б., Николя Э., Мишо Л., Райххарт Дж. М., Хоффманн Дж. А. (сентябрь 1996 г.). «Дорсовентральная регуляторная кассета генов spätzle/Toll/cactus контролирует мощный противогрибковый ответ у взрослых дрозофил» . Клетка . 86 (6): 973–983. дои : 10.1016/s0092-8674(00)80172-5 . ПМИД 8808632 . S2CID 10736743 .
- ^ «Нобелевская премия по физиологии и медицине 2011 года» . NobelPrize.org . Проверено 14 июля 2022 г.
- ^ Карин М., Бен-Нерия Ю. (2000). «Фосфорилирование встречается с убиквитинированием: контроль активности NF-каппа B». Ежегодный обзор иммунологии . 18 : 621–663. doi : 10.1146/annurev.immunol.18.1.621 . ПМИД 10837071 .
- ^ Сенфтлебен У., Цао Ю., Сяо Г., Гретен Ф.Р., Крэн Г., Боницци Г. и др. (август 2001 г.). «Активация IKKalpha второго, эволюционно консервативного сигнального пути NF-каппа B». Наука . 293 (5534): 1495–1499. Бибкод : 2001Sci...293.1495S . дои : 10.1126/science.1062677 . ПМИД 11520989 . S2CID 83308790 .
- ^ Плаксин Д., Бойерле П.А., Айзенбах Л. (июнь 1993 г.). «KBF1 (гомодимер p50 NF-каппа B) действует как репрессор экспрессии гена H-2Kb в метастатических опухолевых клетках» . Журнал экспериментальной медицины . 177 (6): 1651–1662. дои : 10.1084/jem.177.6.1651 . ПМК 2191052 . ПМИД 8496683 .
- ^ Гуань Х., Хоу С., Риккарди Р.П. (март 2005 г.). «ДНК-связывание репрессора ядерного фактора каппаВ p50/p50 зависит от фосфорилирования Ser337 каталитической субъединицей протеинкиназы А» . Журнал биологической химии . 280 (11): 9957–9962. дои : 10.1074/jbc.m412180200 . ПМИД 15642694 .
- ^ Набель Г.Дж., Верма И.М. (ноябрь 1993 г.). «Предлагаемая номенклатура семейства NF-каппа B/I каппа B» . Гены и развитие . 7 (11): 2063. doi : 10.1101/gad.7.11.2063 . ПМИД 8224837 .
- ^ Гош С., Мэй М.Дж., Копп Э.Б. (1998). «Белки NF-каппа B и Rel: эволюционно консервативные медиаторы иммунных ответов». Ежегодный обзор иммунологии . 16 : 225–260. doi : 10.1146/annurev.immunol.16.1.225 . ПМИД 9597130 .
- ^ Уотерхаус Р.М., Кривенцева Е.В., Мейстер С., Кси З, Альварес К.С., Бартоломай Л.С. и др. (июнь 2007 г.). «Эволюционная динамика генов и путей, связанных с иммунитетом, у комаров-переносчиков болезней» . Наука . 316 (5832): 1738–1743. Бибкод : 2007Sci...316.1738W . дои : 10.1126/science.1139862 . ПМК 2042107 . ПМИД 17588928 .
- ^ PDB : 3do7 ; Фуско А.Дж., Хуанг Д.Б., Миллер Д., Ван В.Ю., Ву Д., Гош Г. (февраль 2009 г.). «Гетеродимер NF-kappaB p52:RelB распознает два класса сайтов kappaB с двумя различными режимами» . Отчеты ЭМБО . 10 (2): 152–159. дои : 10.1038/embor.2008.227 . ПМК 2637311 . ПМИД 19098713 .
- ^ (а) Чандель Н.С., Тшина В.К., МакКлинток Д.С., Шумакер П.Т. (июль 2000 г.). «Роль оксидантов в активации NF-каппа B и транскрипции гена TNF-альфа, индуцированной гипоксией и эндотоксином» . Журнал иммунологии . 165 (2): 1013–1021. doi : 10.4049/jimmunol.165.2.1013 . ПМИД 10878378 . ; (б) Фицджеральд Д.К., Мид К.Г., МакЭвой А.Н., Лиллис Л., Мерфи Э.П., МакХью Д.Э. и др. (март 2007 г.). «Фактор некроза опухоли-альфа (TNF-альфа) увеличивает активность ядерного фактора каппаB (NFkappaB) и высвобождение интерлейкина-8 (IL-8) из эпителиальных клеток молочной железы крупного рогатого скота». Ветеринарная иммунология и иммунопатология . 116 (1–2): 59–68. дои : 10.1016/j.vetimm.2006.12.008 . ПМИД 17276517 . ; (с) Ренард П., Закари М.Д., Бужелет С., Миро М.Э., Хэгеман Г., Ремакл Дж. и др. (январь 1997 г.). «Влияние модуляции антиоксидантных ферментов на индуцированную интерлейкином-1 активацию ядерного фактора каппа B». Биохимическая фармакология . 53 (2): 149–160. дои : 10.1016/S0006-2952(96)00645-4 . ПМИД 9037247 . ; (г) Цинь Х, Уилсон К.А., Ли С.Дж., Чжао X, Бенвенист Э.Н. (ноябрь 2005 г.). «ЛПС индуцирует экспрессию гена CD40 посредством активации NF-каппаВ и STAT-1альфа в макрофагах и микроглии» . Кровь . 106 (9): 3114–3122. дои : 10.1182/кровь-2005-02-0759 . ПМК 1895321 . ПМИД 16020513 . ; (е) Такемото Ю., Ёсияма М., Такеучи К., Омура Т., Комацу Р., Изуми Ю. и др. (ноябрь 1999 г.). «Повышение активности связывания ДНК JNK, AP-1 и NF-каппа B при ремоделировании сердца, индуцированном изопротеренолом». Журнал молекулярной и клеточной кардиологии . 31 (11): 2017–2030. дои : 10.1006/jmcc.1999.1033 . ПМИД 10591028 . ; (е) Харгрейв БАЙ, Тиангко Д.А., Латтанцио Ф.А., Биб С.Дж. (2003). «Кокаин, а не морфин, вызывает выработку активных форм кислорода и активацию NF-kappaB в временно котрансфицированных клетках сердца». Сердечно-сосудистая токсикология . 3 (2): 141–151. дои : 10.1385/CT:3:2:141 . ПМИД 14501032 . S2CID 35240781 . ; (г) Нойхофер В., Питтроу Д. (сентябрь 2006 г.). «Роль эндотелина и антагонистов рецепторов эндотелина при заболеваниях почек». Европейский журнал клинических исследований . 36 (Дополнительный 3): 78–88. дои : 10.1111/j.1365-2362.2006.01689.x . ПМИД 16919017 . S2CID 30687039 . ; (час) Басу С., Розенцвейг К.Р., Юмелл М., Прайс Б.Д. (июнь 1998 г.). «ДНК-зависимая протеинкиназа участвует в активации NF каппа B после повреждения ДНК». Связь с биохимическими и биофизическими исследованиями . 247 (1): 79–83. дои : 10.1006/bbrc.1998.8741 . ПМИД 9636658 .
- ^ Папа С., Бубичи С., Заззерони Ф., Фам К.Г., Кунцен С., Кнабб Дж.Р. и др. (май 2006 г.). «NF-kappaB-опосредованный контроль каскада JNK в антагонизме запрограммированной гибели клеток в норме и болезни» . Смерть клеток и дифференцировка . 13 (5): 712–729. дои : 10.1038/sj.cdd.4401865 . ПМИД 16456579 .
- ^ Бодуэн М., Ламуре Ф., Дюпломб Л., Редини Ф., Хейманн Д. (сентябрь 2007 г.). «РАНКЛ, РАНК, остеопротегерин: ключевые партнеры остеоиммунологии и сосудистых заболеваний» . Клеточные и молекулярные науки о жизни . 64 (18): 2334–2350. дои : 10.1007/s00018-007-7104-0 . ПМЦ 11149428 . ПМИД 17530461 . S2CID 32179220 .
- ^ Дойл С.Л., О'Нил Л.А. (октябрь 2006 г.). «Toll-подобные рецепторы: от открытия NFkappaB до нового понимания регуляции транскрипции врожденного иммунитета». Биохимическая фармакология . 72 (9): 1102–1113. дои : 10.1016/j.bcp.2006.07.010 . ПМИД 16930560 .
- ^ Хайден М.С., West AP, Гош С. (октябрь 2006 г.). «NF-каппаВ и иммунный ответ» . Онкоген . 25 (51): 6758–6780. дои : 10.1038/sj.onc.1209943 . ПМИД 17072327 .
- ^ Ли Кью, Верма IM (октябрь 2002 г.). «Регуляция NF-каппаВ в иммунной системе». Обзоры природы. Иммунология . 2 (10): 725–734. дои : 10.1038/nri910 . ПМИД 12360211 . S2CID 6962119 .
- ^ Фудзита Т., Нолан ГП, Лиу ХК, Скотт МЛ, Балтимор Д. (июль 1993 г.). «Кандидат в протоонкоген bcl-3 кодирует коактиватор транскрипции, который активируется через гомодимеры NF-каппа B p50» . Гены и развитие . 7 (7Б): 1354–1363. дои : 10.1101/gad.7.7b.1354 . ПМИД 8330739 .
- ^ Франзосо Г., Бурс В., Парк С., Томита-Ямагути М., Келли К., Зибенлист Ю. (сентябрь 1992 г.). «Кандидатный онкопротеин Bcl-3 является антагонистом ингибирования, опосредованного p50/NF-каппа B» . Природа . 359 (6393): 339–342. Бибкод : 1992Natur.359..339F . дои : 10.1038/359339a0 . ПМИД 1406939 . S2CID 4322739 .
- ^ Бурс В., Францосо Г., Азаренко В., Парк С., Канно Т., Браун К. и др. (март 1993 г.). «Онкопротеин Bcl-3 напрямую трансактивируется через мотивы каппа B посредством ассоциации с ДНК-связывающими гомодимерами p50B» . Клетка . 72 (5): 729–739. дои : 10.1016/0092-8674(93)90401-Б . ПМИД 8453667 .
- ^ Джейкобс, доктор медицинских наук, Харрисон, Южная Каролина (декабрь 1998 г.). «Структура комплекса IkappaBalpha/NF-kappaB» . Клетка . 95 (6): 749–758. дои : 10.1016/S0092-8674(00)81698-0 . ПМИД 9865693 . S2CID 7003353 .
- ^ Jump up to: а б Басак С., Ким Х., Кернс Дж.Д., Тергаонкар В., О'Ди Э., Вернер С.Л. и др. (январь 2007 г.). «Четвертый белок IkappaB в сигнальном модуле NF-kappaB» . Клетка . 128 (2): 369–381. дои : 10.1016/j.cell.2006.12.033 . ПМЦ 1831796 . ПМИД 17254973 . .
- ^ Добжански П., Райсек Р.П., Браво Р. (март 1995 г.). «Специфическое ингибирование транскрипционной активности RelB/p52 С-концевым доменом p100». Онкоген . 10 (5): 1003–1007. ПМИД 7898917 .
- ^ Ло Дж.К., Басак С., Джеймс Э.С., Квиамбо Р.С., Кинселла М.К., Алегре М.Л. и др. (февраль 2006 г.). «Координация между членами семейства NF-kappaB p50 и p52 важна для передачи сигналов LTbetaR в развитии и организации вторичных лимфоидных тканей» . Кровь . 107 (3): 1048–1055. дои : 10.1182/кровь-2005-06-2452 . ПМК 1895903 . ПМИД 16195333 .
- ^ Дептала А., Беднер Э., Горчица В., Дажинкевич З. (ноябрь 1998 г.). «Активация ядерного фактора каппа B (NF-kappaB) с помощью лазерной сканирующей цитометрии (LSC)» . Цитометрия . 33 (3): 376–382. doi : 10.1002/(SICI)1097-0320(19981101)33:3<376::AID-CYTO13>3.0.CO;2-Q . ПМЦ 3874872 . ПМИД 9822350 .
- ^ Нельсон Д.Э., Ихекваба А.Е., Эллиот М., Джонсон Дж.Р., Гибни К.А., Форман Б.Е. и др. (октябрь 2004 г.). «Колебания передачи сигналов NF-kappaB контролируют динамику экспрессии генов». Наука . 306 (5696): 704–708. Бибкод : 2004Sci...306..704N . дои : 10.1126/science.1099962 . ПМИД 15499023 . S2CID 86055964 .
- ^ Хискотт Дж., Квон Х., Генен П. (январь 2001 г.). «Враждебные поглощения: вирусное присвоение пути NF-kappaB» . Журнал клинических исследований . 107 (2): 143–151. дои : 10.1172/JCI11918 . ЧВК 199181 . ПМИД 11160127 .
- ^ Адкинс И., Шульц С., Боргманн С., Аутенрит И.Б., Грёбнер С. (февраль 2008 г.). «Дифференциальная роль внешнего белка P иерсинии в ингибировании ядерного фактора каппа B в индукции гибели клеток в дендритных клетках и макрофагах» . Журнал медицинской микробиологии . 57 (Часть 2): 139–144. дои : 10.1099/jmm.0.47437-0 . ПМИД 18201977 .
- ^ Микели Л., Леонарди Л., Конти Ф., Буанне П., Кану Н., Карузо М. и др. (март 2005 г.). «PC4 коактивирует MyoD, снимая опосредованное гистондеацетилазой 4 ингибирование фактора 2C энхансера миоцитов» . Молекулярная и клеточная биология . 25 (6): 2242–2259. дои : 10.1128/MCB.25.6.2242-2259.2005 . ПМЦ 1061592 . ПМИД 15743821 .
- ^ Микели Л., Леонарди Л., Конти Ф., Мареска Г., Колазингари С., Маттеи Е. и др. (февраль 2011 г.). «PC4/Tis7/IFRD1 стимулирует регенерацию скелетных мышц и участвует в дифференцировке миобластов в качестве регулятора MyoD и NF-kappaB» . Журнал биологической химии . 286 (7): 5691–5707. дои : 10.1074/jbc.M110.162842 . ПМК 3037682 . ПМИД 21127072 .
- ^ Юнг Ф., Хоберг Дж.Э., Рэмси К.С., Келлер М.Д., Джонс Д.Р., Фрай Р.А. и др. (июнь 2004 г.). «Модуляция NF-kappaB-зависимой транскрипции и выживания клеток с помощью деацетилазы SIRT1» . Журнал ЭМБО . 23 (12): 2369–2380. дои : 10.1038/sj.emboj.7600244 . ПМК 423286 . ПМИД 15152190 .
- ^ Боницци Г., Бебьен М., Отеро Д.С., Джонсон-Врум К.Е., Цао Ю., Ву Д. и др. (октябрь 2004 г.). «Активация генов-мишеней IKKalpha зависит от узнавания специфических сайтов связывания kappaB димерами RelB:p52» . Журнал ЭМБО . 23 (21): 4202–4210. дои : 10.1038/sj.emboj.7600391 . ПМК 524385 . ПМИД 15470505 .
- ^ Jump up to: а б Басак С., Ши В.Ф., Хоффманн А. (май 2008 г.). «Генерация и активация множественных димерных факторов транскрипции в сигнальной системе NF-kappaB» . Молекулярная и клеточная биология . 28 (10): 3139–3150. дои : 10.1128/MCB.01469-07 . ПМЦ 2423155 . ПМИД 18299388 .
- ^ Мукерджи Т., Чаттерджи Б., Дхар А., Байс С.С., Чавла М., Рой П. и др. (декабрь 2017 г.). «Путь TNF-p100 подрывает неканоническую передачу сигналов NF-κB в воспаленных вторичных лимфоидных органах» . Журнал ЭМБО . 36 (23): 3501–3516. дои : 10.15252/embj.201796919 . ПМК 5709727 . ПМИД 29061763 .
- ^ Банот Б., Чаттерджи Б., Виджаярагаван Б., Прасад М.В., Рой П., Басак С. (апрель 2015 г.). Чакраборти А.К. (ред.). «Избирательные к стимулам перекрестные помехи через сигнальную систему NF-κB усиливают врожденный иммунный ответ, облегчая кишечную инфекцию» . электронная жизнь . 4 : e05648. doi : 10.7554/eLife.05648 . ПМЦ 4432492 . ПМИД 25905673 .
- ^ Чаттерджи Б., Банот Б., Мукерджи Т., Тайе Н., Виджаярагаван Б., Чаттопадхьяй С. и др. (декабрь 2016 г.). «Поздняя фаза синтеза IκBα изолирует TLR4-активируемый канонический путь NF-κB от неканонической передачи сигналов NF-κB в макрофагах» . Научная сигнализация . 9 (457): ра120. дои : 10.1126/scisignal.aaf1129 . ПМК 5260935 . ПМИД 27923915 .
- ^ Рой П., Мукерджи Т., Чаттерджи Б., Виджаярагаван Б., Банот Б., Басак С. (март 2017 г.). «Неканонические мутации NFκB усиливают способствующий выживанию ответ TNF при множественной миеломе посредством ауторегуляторного пути RelB:p50 NFκB» . Онкоген . 36 (10): 1417–1429. дои : 10.1038/onc.2016.309 . ПМЦ 5346295 . ПМИД 27641334 .
- ^ Смит Э.М., Грегг М., Хашеми Ф., Шотт Л., Хьюз Т.К. (1 июля 2006 г.). «Активация рилизинг-фактором кортикотропина (CRF) NF-kappaB-направленной транскрипции в лейкоцитах». Клеточная и молекулярная нейробиология . 26 (4–6): 1021–1036. дои : 10.1007/s10571-006-9040-1 . ПМИД 16633893 . S2CID 22544468 .
- ^ Ливолси А., Бусуттил В., Имберт В., Абрахам Р.Т., Пейрон Дж.Ф. (март 2001 г.). «Зависимая от фосфорилирования тирозина активация NF-каппа B. Потребность в протеинтирозинкиназах p56 LCK и ZAP-70». Европейский журнал биохимии . 268 (5): 1508–1515. дои : 10.1046/j.1432-1327.2001.02028.x . ПМИД 11231305 .
- ^ Член парламента Мэттсона, член парламента Мефферта (май 2006 г.). «Роль NF-kappaB в выживании, пластичности и заболевании нервных клеток» . Смерть клеток и дифференцировка . 13 (5): 852–860. дои : 10.1038/sj.cdd.4401837 . ПМИД 16397579 .
- ^ Jump up to: а б Хекшер Э.С., Феттер Р.Д., Марек К.В., Альбин С.Д., Дэвис Г.В. (сентябрь 2007 г.). «NF-kappaB, IkappaB и IRAK контролируют плотность глутаматных рецепторов в NMJ дрозофилы» . Нейрон . 55 (6): 859–873. дои : 10.1016/j.neuron.2007.08.005 . ПМК 2701504 . ПМИД 17880891 .
- ^ Jump up to: а б Кальчмидт Б., Ндиай Д., Корте М., Потион С., Арбибе Л., Прюллаге М. и др. (апрель 2006 г.). «NF-kappaB регулирует формирование пространственной памяти и синаптическую пластичность посредством передачи сигналов протеинкиназы A/CREB» . Молекулярная и клеточная биология . 26 (8): 2936–2946. дои : 10.1128/MCB.26.8.2936-2946.2006 . ПМК 1446931 . ПМИД 16581769 .
- ^ Ван Дж, Фу XQ, Лэй В.Л., Ван Т, Шэн А.Л., Луо З.Г. (август 2010 г.). «Ядерный фактор каппаВ контролирует кластеризацию рецепторов ацетилхолина в нервно-мышечном соединении» . Журнал неврологии . 30 (33): 11104–11113. doi : 10.1523/JNEUROSCI.2118-10.2010 . ПМК 6633475 . ПМИД 20720118 .
- ^ Jump up to: а б Boersma MC, Dresselhaus EC, De Biase LM, Mihalas AB, Bergles DE, Meffert MK (апрель 2011 г.). «Потребность в ядерном факторе каппаВ в синаптогенезе, связанном с развитием и пластичностью» . Журнал неврологии . 31 (14): 5414–5425. doi : 10.1523/JNEUROSCI.2456-10.2011 . ПМК 3113725 . ПМИД 21471377 .
- ^ Гутьеррес Х., Хейл В.А., Дольсе Икс, Дэвис А. (апрель 2005 г.). «Передача сигналов NF-kappaB регулирует рост нервных процессов в развивающейся ПНС и ЦНС» . Разработка . 132 (7): 1713–1726. дои : 10.1242/dev.01702 . ПМИД 15743881 .
- ^ Захир А., Йорек М.А., Лим Р. (декабрь 2001 г.). «Влияние сверхэкспрессии фактора созревания глии в первичных астроцитах на активацию киназы MAP, активацию фактора транскрипции и секрецию нейротрофинов». Нейрохимические исследования . 26 (12): 1293–1299. дои : 10.1023/А:1014241300179 . ПМИД 11885780 . S2CID 26418384 .
- ^ Цю Дж., Ху Х., Несич О., Граф М.Р., Рассин Д.К., Вуд Т.Г. и др. (июль 2004 г.). «Влияние олигонуклеотида NF-kappaB «приманки» на экспрессию генов в гиппокампе крыс P7 после гипоксии / ишемии». Журнал нейробиологических исследований . 77 (1): 108–118. дои : 10.1002/мл.20156 . ПМИД 15197744 . S2CID 25522763 .
- ^ Листвак С.Дж., Ратор П., Херкенхэм М. (октябрь 2013 г.). «Минимальная активность NF-κB в нейронах» . Нейронаука . 250 : 282–299. doi : 10.1016/j.neuroscience.2013.07.013 . ПМК 3785079 . ПМИД 23872390 .
- ^ Яросинский К.В., Уитни Л.В., Масса П.Т. (сентябрь 2001 г.). «Специфический дефицит активации ядерного фактора каппаВ в нейронах центральной нервной системы» . Лабораторные исследования; Журнал технических методов и патологии . 81 (9): 1275–1288. дои : 10.1038/labinvest.3780341 . ПМИД 11555675 .
- ^ Херкенхэм М., Ратор П., Браун П., Листвак С.Дж. (октябрь 2011 г.). «Предостережения по использованию антител NF-κB p65 и p50 для исследований ЦНС» . Журнал нейровоспаления . 8 : 141. дои : 10.1186/1742-2094-8-141 . ПМК 3210105 . ПМИД 21999414 .
- ^ Моерман А.М., Мао X, Лукас М.М., Баргер С.В. (апрель 1999 г.). «Характеристика нейронального фактора, связывающего каппаВ, отличного от NF-каппаВ». Исследования мозга. Молекулярные исследования мозга . 67 (2): 303–315. дои : 10.1016/s0169-328x(99)00091-1 . ПМИД 10216229 .
- ^ Мао XR, Моерман-Герцог А.М., Чэнь Ю, Баргер С.В. (май 2009 г.). «Уникальные аспекты регуляции транскрипции в нейронах - нюансы факторов, связанных с NFkappaB и Sp1» . Журнал нейровоспаления . 6:16 . дои : 10.1186/1742-2094-6-16 . ПМК 2693111 . ПМИД 19450264 .
- ^ Мао X, Ян С.Х., Симпкинс Дж.В., Баргер С.В. (март 2007 г.). «Активация глутаматных рецепторов вызывает опосредованную кальпаином деградацию Sp3 и Sp4, известных транскрипционных факторов семейства Sp в нейронах» . Журнал нейрохимии . 100 (5): 1300–1314. дои : 10.1111/j.1471-4159.2006.04297.x . ЧВК 1949346 . ПМИД 17316402 .
- ^ Влахопулос С.А. (август 2017 г.). «Аберрантный контроль NF-κB при раке обеспечивает транскрипционную и фенотипическую пластичность, чтобы уменьшить зависимость от ткани хозяина: молекулярный режим» . Биология и медицина рака . 14 (3): 254–270. дои : 10.20892/j.issn.2095-3941.2017.0029 . ПМК 5570602 . ПМИД 28884042 .
- ^ Jump up to: а б Влахопулос С.А., Сен О, Хенген Н., Аган Дж., Мошови М., Крицелис Е. и др. (август 2015 г.). «Динамический аберрантный NF-κB стимулирует онкогенез: новая модель, охватывающая микроокружение» . Обзоры цитокинов и факторов роста . 26 (4): 389–403. doi : 10.1016/j.cytogfr.2015.06.001 . ПМЦ 4526340 . ПМИД 26119834 .
- ^ Шейх М.С., Хуан И (2003). «Комплексы активации рецептора смерти: для активации рецептора TNF 1 нужны двое» . Клеточный цикл . 2 (6): 550–552. дои : 10.4161/cc.2.6.566 . ПМИД 14504472 .
- ^ Ли Й.Ю., Чунг Г.Т., Луи В.В., То К.Ф., Ма Б.Б., Чоу С. и др. (январь 2017 г.). «Секвенирование экзома и генома рака носоглотки выявляет мутации, активирующие путь NF-κB» . Природные коммуникации . 8 : 14121. Бибкод : 2017NatCo...814121L . дои : 10.1038/ncomms14121 . ПМЦ 5253631 . ПМИД 28098136 .
- ^ Sun SC (январь 2011 г.). «Неканонический сигнальный путь NF-κB» . Клеточные исследования . 21 (1): 71–85. дои : 10.1038/cr.2010.177 . ПМК 3193406 . ПМИД 21173796 .
- ^ Нури М., Масса С., Карадек Дж., Любик А.А., Ли Н., Труонг С. и др. (апрель 2020 г.). «Транзиентная экспрессия Sox9 способствует устойчивости к андрогенной терапии при раке простаты» . Клинические исследования рака . 26 (7): 1678–1689. дои : 10.1158/1078-0432.CCR-19-0098 . ПМИД 31919137 .
- ^ Танигучи К., Карин М. (май 2018 г.). «NF-κB, воспаление, иммунитет и рак: достижение совершеннолетия». Обзоры природы. Иммунология . 18 (5): 309–324. дои : 10.1038/nri.2017.142 . ПМИД 29379212 . S2CID 3701398 .
- ^ Сунь Л., Мэтьюз Л.А., Кабаркас С.М., Чжан Х, Ян А, Чжан Ю и др. (август 2013 г.). «Эпигенетическая регуляция SOX9 сигнальным путем NF-κB в стволовых клетках рака поджелудочной железы» . Стволовые клетки . 31 (8): 1454–1466. дои : 10.1002/stem.1394 . ПМЦ 3775871 . ПМИД 23592398 .
- ^ Эскарсега Р.О., Фуэнтес-Александро С., Гарсиа-Карраско М., Гатика А., Самора А. (март 2007 г.). «Ядерный фактор транскрипции-каппа B и рак». Клиническая онкология . 19 (2): 154–161. дои : 10.1016/j.clon.2006.11.013 . ПМИД 17355113 .
- ^ Лю Ф., Бардхан К., Ян Д., Тангараджу М., Ганапати В., Уоллер Дж.Л. и др. (июль 2012 г.). «NF-κB напрямую регулирует транскрипцию Fas, модулируя Fas-опосредованный апоптоз и подавление опухоли» . Журнал биологической химии . 287 (30): 25530–25540. дои : 10.1074/jbc.M112.356279 . ПМК 3408167 . ПМИД 22669972 .
- ^ Монако С., Андреакос Е., Кириакидис С., Маури С., Бикнелл С., Фоксвелл Б. и др. (апрель 2004 г.). «Канонический путь активации ядерного фактора каппа B избирательно регулирует провоспалительные и протромботические реакции при атеросклерозе человека» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (15): 5634–5639. Бибкод : 2004PNAS..101.5634M . дои : 10.1073/pnas.0401060101 . ПМЦ 397455 . ПМИД 15064395 .
- ^ Венураджу С.М., Йеррамасу А., Кордер Р., Лахири А. (май 2010 г.). «Остеопротегерин как предиктор ишемической болезни сердца, сердечно-сосудистой смертности и заболеваемости» . Журнал Американского колледжа кардиологов . 55 (19): 2049–2061. дои : 10.1016/j.jacc.2010.03.013 . ПМИД 20447527 .
- ^ Либ В., Гона П., Ларсон М.Г., Массаро Дж.М., Липинска И., Кини Дж.Ф. и др. (сентябрь 2010 г.). «Биомаркеры пути остеопротегерина: клинические корреляты, субклинические заболевания, случаи сердечно-сосудистых заболеваний и смертность» . Атеросклероз, тромбоз и сосудистая биология . 30 (9): 1849–1854. дои : 10.1161/ATVBAHA.109.199661 . ПМК 3039214 . ПМИД 20448212 .
- ^ Сун XQ, Lv LX, Ли WQ, Хао YH, Чжао JP (март 2009 г.). «Взаимодействие ядерного фактора каппа B и цитокинов связано с шизофренией». Биологическая психиатрия . 65 (6): 481–488. doi : 10.1016/j.biopsych.2008.10.018 . ПМИД 19058794 . S2CID 10836374 .
- ^ Кайсари С., Ром О., Айзенбуд Д., Резник А.З. (2013). «Участие NF-κB и специфической для мышц Е3 убиквитинлигазы MuRF1 в катаболизме, индуцированном сигаретным дымом, в мышечных трубках C2». Нейробиология дыхания . Достижения экспериментальной медицины и биологии. Том. 788. стр. 7–17. дои : 10.1007/978-94-007-6627-3_2 . ISBN 978-94-007-6626-6 . ПМИД 23835952 .
- ^ Jump up to: а б с Хаджишенгаллис Г., Чавакис Т. (январь 2013 г.). «Эндогенные модуляторы набора воспалительных клеток» . Тенденции в иммунологии . 34 (1): 1–6. дои : 10.1016/j.it.2012.08.003 . ПМЦ 3703146 . ПМИД 22951309 .
- ^ Jump up to: а б с Видал П.М., Лемменс Э., Дули Д., Хендрикс С. (февраль 2013 г.). «Роль «противовоспалительных» цитокинов в регенерации аксонов». Обзоры цитокинов и факторов роста . 24 (1): 1–12. doi : 10.1016/j.cytogfr.2012.08.008 . ПМИД 22985997 .
- ^ Гривенников С.И., Карин М. (февраль 2010 г.). «Опасные связи: сотрудничество STAT3 и NF-kappaB и перекрестные помехи при раке» . Обзоры цитокинов и факторов роста . 21 (1): 11–19. doi : 10.1016/j.cytogfr.2009.11.005 . ПМЦ 2834864 . ПМИД 20018552 .
- ^ Бонавита Э., Галдиеро М.Р., Джайон С., Мантовани А. (2015). «Фагоциты как коррумпированные полицейские при воспалении, связанном с раком». Достижения в области исследований рака . 128 : 141–171. дои : 10.1016/bs.acr.2015.04.013 . ISBN 978-0-12-802316-7 . ПМИД 26216632 .
- ^ Сионов Р.В., Фридлендер З.Г., Гранот З. (декабрь 2015 г.). «Многогранная роль нейтрофилов в микроокружении опухоли» . Микроокружение рака . 8 (3): 125–158. дои : 10.1007/s12307-014-0147-5 . ПМЦ 4714999 . ПМИД 24895166 .
- ^ Конг X, Ли Л, Ли З, Се К (декабрь 2012 г.). «Направленное разрушение структуры стромы поджелудочной железы и опухолевых клеток в случаях рака поджелудочной железы: молекулярная основа для терапевтических последствий» . Обзоры цитокинов и факторов роста . 23 (6): 343–356. doi : 10.1016/j.cytogfr.2012.06.006 . ПМЦ 3505269 . ПМИД 22749856 .
- ^ Меколлари В., Ньювенхейс Б., Верхааген Дж. (2014). «Взгляд на роль передачи сигналов семафоринов класса III при травмах центральной нервной системы» . Границы клеточной нейронауки . 8 : 328. дои : 10.3389/fncel.2014.00328 . ПМК 4209881 . ПМИД 25386118 .
- ^ Информация о синдроме дефицита NEMO. Информация о синдроме дефицита NEMO], Детская больница Грейт-Ормонд-стрит.
- ^ Кауппинен А, Сууронен Т, Оджала Дж, Каарниранта К, Салминен А (октябрь 2013 г.). «Антагонистические перекрестные помехи между NF-κB и SIRT1 в регуляции воспаления и метаболических нарушений». Сотовая сигнализация . 25 (10): 1939–1948. doi : 10.1016/j.cellsig.2013.06.007 . ПМИД 23770291 .
- ^ де Грегорио Э., Колелл А., Моралес А., Мари М. (май 2020 г.). «Значение оси SIRT1-NF-κB как терапевтической мишени для облегчения воспаления при заболевании печени» . Международный журнал молекулярных наук . 21 (11): 3858. doi : 10.3390/ijms21113858 . ПМК 7312021 . ПМИД 32485811 .
- ^ Ван Р., Ю З., Сунчу Б., Шоаф Дж., Данг И., Чжао С. и др. (июнь 2017 г.). «Рапамицин ингибирует секреторный фенотип стареющих клеток с помощью Nrf2-независимого механизма» . Стареющая клетка . 16 (3): 564–574. дои : 10.1111/acel.12587 . ПМК 5418203 . ПМИД 28371119 .
- ^ Ярбро-младший, Эммонс Р.С., Пенс Б.Д. (июнь 2020 г.). «Макрофагальный иммунометаболизм и воспаление: роль митохондриальной дисфункции, клеточного старения, CD38 и НАД» . Иммунометаболизм . 2 (3): e200026. doi : 10.20900/immunometab20200026 . ПМЦ 7409778 . ПМИД 32774895 .
- ^ Jump up to: а б с д Тилстра Дж.С., Робинсон А.Р., Ван Дж., Грегг С.К., Клаусон К.Л., Рей Д.П. и др. (июль 2012 г.). «Ингибирование NF-κB задерживает старение и старение, вызванное повреждением ДНК, у мышей» . Джей Клин Инвест . 122 (7): 2601–12. дои : 10.1172/JCI45785 . ПМК 3386805 . ПМИД 22706308 .
- ^ Робисон А.Дж., Нестлер Э.Дж. (октябрь 2011 г.). «Транскрипционные и эпигенетические механизмы зависимости» . Обзоры природы. Нейронаука . 12 (11): 623–637. дои : 10.1038/nrn3111 . ПМЦ 3272277 . ПМИД 21989194 .
- ^ Jump up to: а б с д Раффл Дж.К. (ноябрь 2014 г.). «Молекулярная нейробиология зависимости: о чем вообще (Δ)FosB?». Американский журнал о злоупотреблении наркотиками и алкоголем . 40 (6): 428–437. дои : 10.3109/00952990.2014.933840 . ПМИД 25083822 . S2CID 19157711 .
- ^ Jump up to: а б Нестлер Э.Дж. (декабрь 2013 г.). «Клеточная основа памяти при наркомании» . Диалоги в клинической неврологии . 15 (4): 431–443. doi : 10.31887/DCNS.2013.15.4/enestler . ПМЦ 3898681 . ПМИД 24459410 .
- ^ Jump up to: а б Нестлер Э.Дж. (октябрь 2008 г.). «Обзор. Транскрипционные механизмы привыкания: роль DeltaFosB» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 363 (1507): 3245–3255. дои : 10.1098/rstb.2008.0067 . ПМК 2607320 . ПМИД 18640924 .
Таблица 3 - ^ US 6410516 , Балтимор Д., Сен Р., Шарп П.А., Сингх Х., Штаудт Л., Лебовиц Дж.Х., Болдуин-младший А.С., Клерк Р.Г., Коркоран Л.М., Бауерле П.А., Ленардо М.Дж., Фан СМ, Маниатис Т.П., «Ядерные факторы, связанные с регуляцией транскрипции». ", выдано 25 июня 2002 г., передано Гарвардскому колледжу, Институту биомедицинских исследований Уайтхеда, Массачусетскому технологическому институту.
- ^ Карин М. (март 2008 г.). «Киназа IkappaB – мост между воспалением и раком» . Клеточные исследования . 18 (3): 334–342. дои : 10.1038/cr.2008.30 . ПМИД 18301380 .
- ^ Пикарский Э., Бен-Нерия Ю. (апрель 2006 г.). «Ингибирование NF-каппаB: палка о двух концах при раке?». Европейский журнал рака . 42 (6): 779–784. дои : 10.1016/j.ejca.2006.01.011 . ПМИД 16530406 .
- ^ Мантовани А., Маркези Ф., Портал С., Аллавена П., Сика А. (2008). «Связь воспалительных реакций с раком: новые цели для терапевтических стратегий». Таргетная терапия рака . Достижения экспериментальной медицины и биологии. Том. 610. стр. 112–127. дои : 10.1007/978-0-387-73898-7_9 . ISBN 978-0-387-73897-0 . ПМИД 18593019 .
- ^ Паур И., Бальстад Т.Р., Кольберг М., Педерсен М.К., Аустенаа Л.М., Джейкобс Д.Р. и др. (май 2010 г.). «Экстракт орегано, кофе, тимьяна, гвоздики и грецких орехов ингибирует NF-каппаВ в моноцитах и у трансгенных репортерных мышей» . Исследования по профилактике рака . 3 (5): 653–663. дои : 10.1158/1940-6207.CAPR-09-0089 . ПМИД 20424131 .
- ^ Линь З, Ву Д, Хуан Л, Цзян С, Пан Т, Кан Икс и др. (2019). «Нобилетин ингибирует IL-1β-индуцированное воспаление в хондроцитах посредством подавления передачи сигналов NF-κB и ослабляет остеоартрит у мышей» . Границы в фармакологии . 10 : 570. дои : 10.3389/fphar.2019.00570 . ПМК 6554687 . ПМИД 31214026 .
- ^ Дин Ю, Хуан Х, Лю Т, Фу Ю, Тан З, Чжэн Х и др. (октябрь 2012 г.). «Белок циркумспорозоита Plasmodium, новый ингибитор NF-κB, подавляет рост SW480». Патологические и онкологические исследования . 18 (4): 895–902. дои : 10.1007/s12253-012-9519-7 . ПМИД 22678765 . S2CID 15823271 .
- ^ Уайт П.Т., Субраманиан С., Мотивала Х.Ф., Коэн М.С. (2016). «Природные витанолиды в лечении хронических заболеваний». В Гупта С.К., Прасад С., Аггарвал Б.Б. (ред.). Противовоспалительные нутрицевтики и хронические заболевания . Достижения экспериментальной медицины и биологии. Том. 928. Международное издательство Springer. стр. 329–373. дои : 10.1007/978-3-319-41334-1_14 . ISBN 978-3-319-41332-7 . ПМК 7121644 . ПМИД 27671823 .
- ^ Вэй Цзи, Ли Т, Куанг Х, Су Х, Ван Ц (11 февраля 2020 г.). «Фармакологические эффекты витанолидов» . Биомедицинский журнал научно-технических исследований . 25 (3): 19243–19248. дои : 10.26717/BJSTR.2020.25.004218 . ISSN 2574-1241 .
- ^ Гарг А., Аггарвал Б.Б. (июнь 2002 г.). «Ядерный фактор транскрипции каппаB как мишень для разработки лекарств от рака» . Лейкемия . 16 (6): 1053–1068. дои : 10.1038/sj.leu.2402482 . ПМИД 12040437 .
- ^ Сетхи Дж., Сунг Б., Аггарвал Б.Б. (январь 2008 г.). «Активация ядерного фактора каппаB: от скамьи до постели». Экспериментальная биология и медицина . 233 (1): 21–31. дои : 10.3181/0707-MR-196 . ПМИД 18156302 . S2CID 86359181 .
- ^ Член парламента Каррана, Маккидж К. (2009). «Бортезомиб: обзор его применения у пациентов с множественной миеломой». Наркотики . 69 (7): 859–888. дои : 10.2165/00003495-200969070-00006 . ПМИД 19441872 .
- ^ Рэдлер Л. (март 2015 г.). «Велкейд (Бортезомиб) получил два новых показания FDA: для повторного лечения пациентов с множественной миеломой и для лечения первой линии пациентов с мантийно-клеточной лимфомой» . Американские льготы по здоровью и лекарствам . 8 (спецификация): 135–140. ПМК 4665054 . ПМИД 26629279 .
- ^ Влахопулос С., Болдох И., Касола А., Брейзер А.Р. (сентябрь 1999 г.). «Ядерный фактор-каппаВ-зависимая индукция экспрессии гена интерлейкина-8 с помощью фактора некроза опухоли альфа: доказательства наличия пути активации, чувствительного к антиоксидантам, отличного от ядерной транслокации». Кровь . 94 (6): 1878–1889. дои : 10.1182/blood.V94.6.1878.418k03_1878_1889 . ПМИД 10477716 . S2CID 25974629 .
- ^ Хамди Н.А. (январь 2008 г.). «Деносумаб: ингибирование RANKL в борьбе с потерей костной массы». Наркотики сегодня . 44 (1): 7–21. дои : 10.1358/точка.2008.44.1.1178467 . ПМИД 18301800 .
- ^ Цвек Б, Дворжак З (2007). «Нацеливание на ядерный фактор-каппаВ и протеасому с помощью дитиокарбаматных комплексов с металлами». Текущий фармацевтический дизайн . 13 (30): 3155–3167. дои : 10.2174/138161207782110390 . ПМИД 17979756 .
- ^ Блейкли CM, Пазаренцос Э, Оливас В, Астхана С, Ян Дж. Дж., Тан И. и др. (апрель 2015 г.). «Комплекс, активирующий NF-κB, участвующий в ответ на ингибирование онкогена EGFR, способствует выживанию опухолевых клеток и остаточным заболеваниям при раке легких» . Отчеты по ячейкам . 11 (1): 98–110. дои : 10.1016/j.celrep.2015.03.012 . ПМК 4394036 . ПМИД 25843712 .
- ^ Фабр С., Мимура Н., Бобб К., Конг С.Ю., Горгун Г., Цирстеа Д. и др. (сентябрь 2012 г.). «Двойное ингибирование канонических и неканонических путей NF-κB демонстрирует значительную противоопухолевую активность при множественной миеломе» . Клинические исследования рака . 18 (17): 4669–4681. дои : 10.1158/1078-0432.CCR-12-0779 . ПМК 4456190 . ПМИД 22806876 .
- ^ Шоно Ю., Такетт А.З., Лиу Х.К., Дубровина Е., Дерензини Е., Оук С. и др. (январь 2016 г.). «Характеристика ингибитора c-Rel, который опосредует противораковые свойства при гематологических злокачественных новообразованиях путем блокирования NF-κB-контролируемых реакций окислительного стресса» . Исследования рака . 76 (2): 377–389. дои : 10.1158/0008-5472.CAN-14-2814 . ПМЦ 4715937 . ПМИД 26744524 .
- ^ Ямамото М., Хори Р., Такейри М., Козава И., Умедзава К. (сентябрь 2008 г.). «Инактивация компонентов NF-kappaB путем ковалентного связывания (-)-дегидроксиметилэпоксихиномицина со специфическими остатками цистеина». Журнал медицинской химии . 51 (18): 5780–5788. дои : 10.1021/jm8006245 . ПМИД 18729348 .
- ^ «Роль RCP006 как противовоспалительного средства» . Институт Роскампа. Архивировано из оригинала 23 октября 2011 г. Проверено 6 сентября 2011 г.
- ^ Колати С.Р., Касала Э.Р., Боддулуру Л.Н., Махаредди Дж.Р., Уппулапу С.К., Гогои Р. и др. (март 2015 г.). «BAY 11-7082 облегчает диабетическую нефропатию за счет ослабления опосредованного гипергликемией окислительного стресса и воспаления почек через путь NF-κB». Экологическая токсикология и фармакология . 39 (2): 690–699. дои : 10.1016/j.etap.2015.01.019 . ПМИД 25704036 .
- ^ Кумар А., Неги Г., Шарма С.С. (май 2012 г.). «Подавление окислительного стресса и нейровоспаления, регулируемых NF-κB и NF-κB, с помощью BAY 11-7082 (ингибитор фосфорилирования IκB) при экспериментальной диабетической нейропатии». Биохимия . 94 (5): 1158–1165. дои : 10.1016/j.biochi.2012.01.023 . ПМИД 22342224 .
- ^ Дана Н., Васеги Г., Хаджой Джаванмард С. (февраль 2019 г.). «Перекрестные помехи между рецепторами, активируемыми пролифератором пероксисом, и Toll-подобными рецепторами: систематический обзор» . Расширенный фармацевтический бюллетень . 9 (1): 12–21. дои : 10.15171/apb.2019.003 . ПМК 6468223 . ПМИД 31011554 .
- ^ Танака К., Ямагути Т., Хара М. (май 2015 г.). «Игуратимод для лечения ревматоидного артрита в Японии». Экспертное обозрение клинической иммунологии . 11 (5): 565–573. дои : 10.1586/1744666X.2015.1027151 . ПМИД 25797025 . S2CID 25134255 .
Внешние ссылки
[ редактировать ]- NF-каппа + B в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Санкар Гош (2006). Справочник по фактору транскрипции NF-κB . Бока-Ратон: CRC. ISBN 978-0-8493-2794-0 .
- Томас Д. Гилмор. «Путь передачи сигнала Rel/NF-κB» . Бостонский университет . Проверено 2 декабря 2007 г.