Ядерный рецептор

В области молекулярной биологии ядерные рецепторы представляют собой класс белков, ответственных за восприятие стероидов , щитовидной железы гормонов , витаминов и некоторых других молекул. Эти внутриклеточные рецепторы работают вместе с другими белками, регулируя экспрессию определенных генов , тем самым контролируя развитие , гомеостаз и метаболизм организма.
Ядерные рецепторы связываются непосредственно с ДНК, регулируя экспрессию соседних генов; следовательно, эти рецепторы классифицируются как факторы транскрипции . [2] [3] Регуляция экспрессии генов ядерными рецепторами часто происходит в присутствии лиганда — молекулы, влияющей на поведение рецептора. Связывание лиганда с ядерным рецептором приводит к конформационному изменению, активирующему рецептор. Результатом является повышение или понижение экспрессии генов.
Уникальным свойством ядерных рецепторов, отличающим их от других классов рецепторов, является их прямой контроль над геномной ДНК. Ядерные рецепторы играют ключевую роль как в эмбриональном развитии, так и в гомеостазе взрослых. Как обсуждается ниже, ядерные рецепторы классифицируются по механизму действия. [4] [5] или гомология . [6] [7]
Распространение видов
[ редактировать ]Ядерные рецепторы специфичны для многоклеточных животных (животных) и не встречаются у протистов , водорослей , грибов и растений. [8] Среди рано ветвящихся линий животных с секвенированными геномами две были зарегистрированы от губки Amphimedon Queenslandica , две - от гребневика Mnemiopsis leidyi. [9] четыре от плакозойных Trichoplax adhaerens и 17 от книдарий Nematostella vectensis . [10] имеется 270 ядерных рецепторов Только у круглого червя Caenorhabditis elegans . [11] 21 у плодовой мухи и других насекомых, [12] 73 у рыбок данио . [13] У человека, мыши и крысы имеется по 48, 49 и 47 ядерных рецепторов соответственно. [14]
Лиганды
[ редактировать ]
Лиганды, которые связываются с ядерными рецепторами и активируют их, включают липофильные вещества, такие как эндогенные гормоны , витамины А и D и ксенобиотические гормоны . Поскольку экспрессия большого количества генов регулируется ядерными рецепторами, лиганды, активирующие эти рецепторы, могут оказывать глубокое воздействие на организм. Многие из этих регулируемых генов связаны с различными заболеваниями, что объясняет, почему молекулярные мишени примерно 13% лекарств, одобренных Управлением по контролю за продуктами и лекарствами США (FDA), нацелены на ядерные рецепторы. [15]
Ряд ядерных рецепторов, называемых сиротскими рецепторами , [16] не имеют известных (или, по крайней мере, общепринятых) эндогенных лигандов. Некоторые из этих рецепторов, такие как FXR , LXR и PPAR, связывают ряд промежуточных продуктов метаболизма, таких как жирные кислоты, желчные кислоты и/или стерины, с относительно низким сродством. Следовательно, эти рецепторы могут функционировать как метаболические сенсоры. Другие ядерные рецепторы, такие как CAR и PXR , по-видимому, функционируют как сенсоры ксенобиотиков, повышая экспрессию ферментов цитохрома P450 , которые метаболизируют эти ксенобиотики. [17]
Структура
[ редактировать ]Большинство ядерных рецепторов имеют молекулярную массу от 50 000 до 100 000 дальтон .
Ядерные рецепторы имеют модульную структуру и содержат следующие домены : [18] [19]
- (AB) N-концевой регуляторный домен: содержит функцию активации 1 ( AF-1 ), действие которой не зависит от присутствия лиганда. [20] Транскрипционная активация AF-1 обычно очень слабая, но она взаимодействует с AF-2 в E-домене (см. ниже), вызывая более сильное усиление экспрессии генов. Последовательность домена AB сильно варьируется между различными ядерными рецепторами.
- (C) ДНК-связывающий домен ( DBD ): высококонсервативный домен, содержащий два цинковых пальца , которые связываются со специфическими последовательностями ДНК, называемыми элементами гормонального ответа (HRE). Недавно новый мотив цинковых пальцев (CHC2) идентифицирован у NR паразитических плоских червей. [21]
- (D) Шарнирная область: считается гибким доменом, который соединяет DBD с LBD. Влияет на внутриклеточный транспорт и субклеточное распределение с помощью целевой пептидной последовательности.
- (E) Лигандсвязывающий домен ( LBD ): умеренно консервативен по последовательности и высококонсервативен по структуре между различными ядерными рецепторами. Структура . LBD называется альфа-спиральной сэндвич- складкой , в которой три антипараллельные альфа-спирали («сэндвич-начинка») окружены двумя альфа-спиралями с одной стороны и тремя с другой («хлеб») Полость для связывания лиганда находится внутри LBD и чуть ниже трех антипараллельных альфа-спиральных сэндвич-«наполнителей». Наряду с DBD, LBD участвует в интерфейсе димеризации рецептора и, кроме того, связывает коактиваторные и корепрессорные белки. LBD также содержит функцию активации 2 ( AF-2 ), действие которой зависит от присутствия связанного лиганда, контролируемого конформацией спирали 12 (H12). [20]
- (F) С-концевой домен: последовательность различных ядерных рецепторов сильно варьируется.
N-концевой (A/B), ДНК-связывающий (C) и лигандсвязывающий (E) домены независимо хорошо свернуты и структурно стабильны, в то время как шарнирная область (D) и необязательные C-концевые (F) домены могут быть конформационно гибкий и неупорядоченный. [22] Относительная ориентация доменов сильно различается при сравнении трех известных многодоменных кристаллических структур, две из которых связываются с DR1 (DBD, разделенные 1 п.н.), [1] [23] одно связывание с DR4 (на 4 п.н.). [24]
Вверху – схематическая одномерная аминокислотная последовательность ядерного рецептора. Внизу — трехмерные структуры областей DBD (связанных с ДНК) и LBD (связанных с гормонами) ядерного рецептора. Показанные структуры относятся к рецептору эстрогена . Экспериментальные структуры N-концевого домена (A/B), шарнирной области (D) и C-концевого домена (F) не были определены, поэтому представлены красными, фиолетовыми и оранжевыми пунктирными линиями соответственно. |
|
| ||||||||||||||||||||||||||||||||||||||||||||||

Механизм действия
[ редактировать ]Ядерные рецепторы представляют собой многофункциональные белки, передающие сигналы родственных им лигандов . Ядерные рецепторы (NR) можно разделить на два широких класса в соответствии с механизмом их действия и внутриклеточным распределением в отсутствие лиганда.
Небольшие липофильные вещества, такие как природные гормоны, диффундируют через клеточную мембрану и связываются с ядерными рецепторами, расположенными в цитозоле (NR типа I) или ядре (NR типа II) клетки. Связывание вызывает конформационные изменения в рецепторе, которые, в зависимости от класса рецептора, запускают каскад последующих событий, которые направляют NR к сайтам регуляции транскрипции ДНК, что приводит к усилению или понижению экспрессии генов. Обычно они функционируют как гомо/гетеродимеры. [27] Кроме того, были идентифицированы два дополнительных класса: тип III, который является вариантом типа I, и тип IV, который связывает ДНК в виде мономеров. [4]
Соответственно, ядерные рецепторы можно разделить на следующие четыре механистических класса: [4] [5]
Тип I
[ редактировать ]Связывание лиганда с ядерными рецепторами типа I в цитозоле приводит к диссоциации белков теплового шока , гомодимеризации , транслокации ( т. е . активному транспорту ) из цитоплазмы в ядро клетки и связыванию со специфическими последовательностями ДНК , известными как элементы гормонального ответа. (ОПЧ). Ядерные рецепторы типа I связываются с HRE, состоящими из двух полусайтов, разделенных ДНК переменной длины, причем второй полусайт имеет последовательность, инвертированную по сравнению с первым (инвертированный повтор). Ядерные рецепторы типа I включают представителей подсемейства 3, таких как рецепторы андрогенов , рецепторы эстрогена , рецепторы глюкокортикоидов и рецепторы прогестерона . [28]
Было отмечено, что некоторые из ядерных рецепторов подсемейства 2 NR могут связываться с прямым повтором вместо HRE с инвертированным повтором . Кроме того, некоторые ядерные рецепторы связываются либо в виде мономеров, либо в виде димеров, при этом только один ДНК-связывающий домен рецептора прикрепляется к одному половинному участку HRE. Эти ядерные рецепторы считаются рецепторами-сиротами , поскольку их эндогенные лиганды до сих пор неизвестны.
ядерный рецептор/ДНК Затем комплекс рекрутирует другие белки, которые транскрибируют ДНК ниже HRE в информационную РНК и, в конечном итоге, в белок , что вызывает изменение функции клетки.
Тип II
[ редактировать ]Рецепторы типа II, в отличие от типа I, сохраняются в ядре независимо от статуса связывания лиганда и дополнительно связываются с ДНК в виде гетеродимеров (обычно с RXR ). [27] В отсутствие лиганда ядерные рецепторы типа II часто образуют комплекс с белками -корепрессорами . Связывание лиганда с ядерным рецептором вызывает диссоциацию корепрессора и рекрутирование белков -коактиваторов . Дополнительные белки, включая РНК-полимеразу, затем рекрутируются в комплекс NR/ДНК, которые транскрибируют ДНК в информационную РНК.
Ядерные рецепторы типа II включают в основном подсемейство 1, например, рецептор ретиноевой кислоты , рецептор ретиноида X и рецептор гормона щитовидной железы . [29]
Тип III
[ редактировать ]Ядерные рецепторы типа III (в основном подсемейство 2 NR) аналогичны рецепторам типа I в том, что оба класса связываются с ДНК как гомодимеры. Однако ядерные рецепторы типа III, в отличие от типа I, связываются с прямыми повторами, а не с инвертированными повторами HRE.
Тип IV
[ редактировать ]Ядерные рецепторы типа IV связываются либо в виде мономеров, либо в виде димеров, но только один ДНК-связывающий домен рецептора связывается с одним половинным сайтом HRE. Примеры рецепторов типа IV обнаружены в большинстве подсемейств NR.
Димеризация
[ редактировать ]Ядерные рецепторы человека способны димеризоваться со многими другими ядерными рецепторами (гомотипическая димеризация), как было показано в результате крупномасштабных экспериментов с Y2H и усилий по анализу текста в литературе, которые были сосредоточены на конкретных взаимодействиях. [30] [31] [27] Тем не менее, существует специфичность: члены одного и того же подсемейства имеют очень сходных партнеров по димеризации NR, а лежащая в основе сеть димеризации имеет определенные топологические особенности, такие как наличие сильно связанных концентраторов (RXR и SHP). [27]
Корегуляторные белки
[ редактировать ]Ядерные рецепторы, связанные с элементами гормонального ответа, рекрутируют значительное количество других белков (называемых корегуляторами транскрипции ), которые облегчают или ингибируют транскрипцию соответствующего гена-мишени в мРНК. [32] [33] [34] Функции этих корегуляторов разнообразны и включают ремоделирование хроматина (делание гена-мишени более или менее доступным для транскрипции) или мостиковую функцию для стабилизации связывания других корегуляторных белков. Ядерные рецепторы могут специфически связываться с рядом белков-корегуляторов и тем самым влиять на клеточные механизмы передачи сигнала как напрямую, так и опосредованно. [35]
Коактиваторы
[ редактировать ]Связывание лигандов-агонистов (см. раздел ниже) с ядерными рецепторами индуцирует конформацию рецептора, которая преимущественно связывает белки -коактиваторы . Эти белки часто обладают внутренней активностью гистон-ацетилтрансферазы (HAT), которая ослабляет связь гистонов с ДНК и, следовательно, способствует транскрипции генов.
Корепрессоры
[ редактировать ]Связывание лигандов-антагонистов с ядерными рецепторами, напротив, индуцирует конформацию рецептора, которая преимущественно связывает корепрессорные белки. Эти белки, в свою очередь, рекрутируют деацетилазы гистонов (HDAC), которые усиливают ассоциацию гистонов с ДНК и, следовательно, подавляют транскрипцию генов.
Агонизм против антагонизма
[ редактировать ]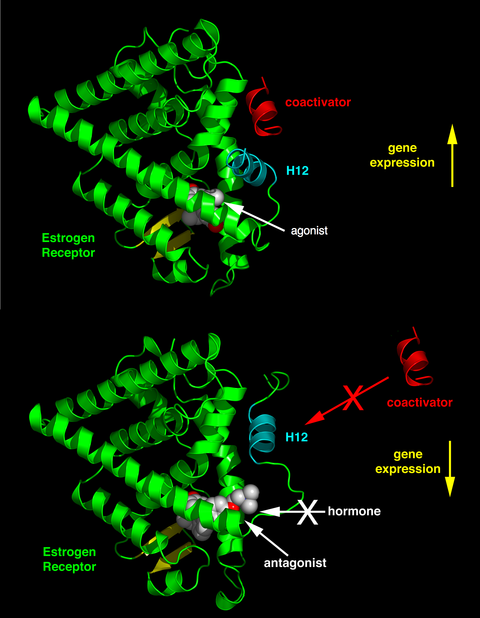
В зависимости от задействованного рецептора, химической структуры лиганда и ткани, на которую воздействуют, лиганды ядерных рецепторов могут проявлять совершенно разнообразные эффекты, варьирующиеся в диапазоне от агонизма до антагонизма и обратного агонизма. [38]
Агонисты
[ редактировать ]Активность эндогенных лигандов (таких как гормоны эстрадиол и тестостерон ) при связывании с родственными им ядерными рецепторами обычно приводит к усилению экспрессии генов. Эта стимуляция экспрессии генов лигандом называется агонистической реакцией. Агонистические эффекты эндогенных гормонов также могут быть имитированы некоторыми синтетическими лигандами, например, глюкокортикоидных рецепторов противовоспалительным препаратом дексаметазоном . Лиганды-агонисты действуют, индуцируя конформацию рецептора, способствующую связыванию коактиватора (см. верхнюю половину рисунка справа).
Антагонисты
[ редактировать ]Другие синтетические лиганды ядерных рецепторов не оказывают очевидного влияния на транскрипцию генов в отсутствие эндогенного лиганда. Однако они блокируют эффект агониста посредством конкурентного связывания с тем же сайтом связывания ядерного рецептора. Эти лиганды называются антагонистами. Примером препарата-антагониста ядерных рецепторов является мифепристон , который связывается с рецепторами глюкокортикоида и прогестерона и, следовательно, блокирует активность эндогенных гормонов кортизола и прогестерона соответственно. Лиганды-антагонисты действуют, индуцируя конформацию рецептора, которая предотвращает связывание коактиватора и способствует связыванию корепрессора (см. нижнюю половину рисунка справа).
Обратные агонисты
[ редактировать ]Наконец, некоторые ядерные рецепторы способствуют низкому уровню транскрипции генов в отсутствие агонистов (также называемому базальной или конститутивной активностью). Синтетические лиганды, которые снижают базальный уровень активности ядерных рецепторов, известны как обратные агонисты . [39]
Селективные модуляторы рецепторов
[ редактировать ]Ряд препаратов, действующих через ядерные рецепторы, вызывают агонистическую реакцию в одних тканях и антагонистическую реакцию в других тканях. Такое поведение может иметь существенные преимущества, поскольку оно может позволить сохранить желаемые полезные терапевтические эффекты лекарства, минимизируя при этом нежелательные побочные эффекты. Препараты со смешанным профилем действия агонистов/антагонистов называются селективными модуляторами рецепторов (SRM). Примеры включают селективные модуляторы рецепторов андрогена ( SARM ), селективные модуляторы рецепторов эстрогена ( SERM ) и селективные модуляторы рецепторов прогестерона ( SPRM ). Механизм действия SRM может варьироваться в зависимости от химической структуры лиганда и задействованного рецептора, однако считается, что многие SRM действуют путем стимулирования конформации рецептора, которая тесно сбалансирована между агонизмом и антагонизмом. В тканях, где концентрация белков -коактиваторов выше, чем корепрессоров , равновесие смещается в сторону агонистов. И наоборот, в тканях, где корепрессоры доминируют, лиганд ведет себя как антагонист. [40]
Альтернативные механизмы
[ редактировать ]
Трансрепрессия
[ редактировать ]Наиболее распространенный механизм действия ядерного рецептора включает прямое связывание ядерного рецептора с элементом ответа на гормон ДНК. Этот механизм называется трансактивацией . Однако некоторые ядерные рецепторы обладают способностью напрямую связываться не только с ДНК, но и с другими факторами транскрипции. Это связывание часто приводит к дезактивации второго транскрипционного фактора в процессе, известном как трансрепрессия . [41] Одним из примеров ядерного рецептора, способного трансрепрессировать, является глюкокортикоидный рецептор (GR). Кроме того, некоторые лиганды GR, известные как селективные агонисты глюкокортикоидных рецепторов ( SEGRA ), способны активировать GR таким образом, что GR более сильно трансрепрессирует, чем трансактивирует. Эта селективность увеличивает разделение между желаемыми противовоспалительными эффектами и нежелательными метаболическими побочными эффектами этих селективных глюкокортикоидов .
Негеномный
[ редактировать ]Классическое прямое воздействие ядерных рецепторов на регуляцию генов обычно занимает несколько часов, прежде чем в клетках будет замечен функциональный эффект из-за большого количества промежуточных этапов между активацией ядерных рецепторов и изменениями уровней экспрессии белка. Однако было замечено, что многие эффекты применения ядерных гормонов, такие как изменения активности ионных каналов, происходят в течение нескольких минут, что несовместимо с классическим механизмом действия ядерных рецепторов. Хотя молекулярная мишень для этих негеномных эффектов ядерных рецепторов окончательно не продемонстрирована, было высказано предположение, что существуют варианты ядерных рецепторов, которые связаны с мембраной, а не локализованы в цитозоле или ядре. Более того, эти мембраносвязанные рецепторы функционируют посредством альтернативных механизмов передачи сигнала , не затрагивающих генную регуляцию. [42] [43]
Хотя была выдвинута гипотеза о наличии нескольких мембраносвязанных рецепторов ядерных гормонов, было показано, что для многих быстрых эффектов требуются канонические ядерные рецепторы. [44] [45] Однако проверка относительной важности геномных и негеномных механизмов in vivo невозможна из-за отсутствия специфических молекулярных механизмов негеномных эффектов, которые можно было бы блокировать мутацией рецептора, не нарушая его прямого воздействия на экспрессию генов.
Молекулярный механизм негеномной передачи сигналов через ядерный рецептор гормона щитовидной железы TRβ включает фосфатидилинозитол-3-киназу ( PI3K ). [46] Эта передача сигналов может быть заблокирована единственной заменой тирозина на фенилаланин в TRβ без нарушения прямой регуляции генов. [47] Когда мыши были созданы с этой единственной консервативной аминокислотной заменой в TRβ, [47] Синаптическое созревание и пластичность в гиппокампе нарушались почти так же эффективно, как и полная блокировка синтеза гормонов щитовидной железы. [48] Этот механизм, по-видимому, консервативен у всех млекопитающих, но не у TRα или каких-либо других ядерных рецепторов. Таким образом, фосфотирозин-зависимая ассоциация TRβ с PI3K обеспечивает потенциальный механизм интеграции регуляции развития и метаболизма гормонами щитовидной железы и рецепторными тирозинкиназами. Кроме того, передача сигналов гормонов щитовидной железы через PI3K может изменять экспрессию генов. [49]
Члены семьи
[ редактировать ]Ниже приводится список из 48 известных ядерных рецепторов человека (и их ортологов у других видов). [14] [50] [51] классифицируются по гомологии последовательностей . [6] [7] В список также включены избранные члены семьи, у которых нет человеческих ортологов (символ NRNC выделен желтым цветом).
| Подсемейство | Группа | Член | ||||||
|---|---|---|---|---|---|---|---|---|
| Символ НСРН [6] | Аббревиатура | Имя | Ген | Лиганд(ы) | ||||
| 1 | Подобный рецептору гормона щитовидной железы | А | Рецептор гормона щитовидной железы | NR1A1 | TRα | Рецептор гормона щитовидной железы-α | ТРА | гормон щитовидной железы |
| НР1А2 | ТРβ | Рецептор гормона щитовидной железы-β | ТРБ | |||||
| Б | Рецептор ретиноевой кислоты | NR1B1 | РАРα | Рецептор ретиноевой кислоты-α | НИКОГДА | витамин А и родственные соединения | ||
| NR1B2 | RARβ | Рецептор ретиноевой кислоты-β | РАРБ | |||||
| NR1B3 | РАРγ | Рецептор ретиноевой кислоты-γ | УРОДЛИВЫЙ | |||||
| С | Рецептор, активирующий пролифератор пероксисом | NR1C1 | PPARα | Рецептор-α, активирующий пролифератор пероксисом | ППАРА | жирные кислоты , простагландины | ||
| NR1C2 | PPAR-β/δ | Рецептор-β/δ, активирующий пролифератор пероксисом | ППАРД | |||||
| NR1C3 | PPARγ | Рецептор-γ, активирующий пролифератор пероксисом | ППАРГ | |||||
| Д | Рев-Эрба | НР1Д1 | Рев-ЭрбАα | Рев-ЭрбАα | НР1Д1 | гем | ||
| НР1Д2 | Рев-ЭрбАβ | Рев-ЭрбАα | НР1Д2 | |||||
| И | E78C-подобный (членистоногие, трематоды, муллоски, нематоды) [50] [52] | NR1E1 | Eip78C | Белок 78C, индуцируемый экдизоном | Eip78C | |||
| Ф | RAR-связанный сиротский рецептор | NR1F1 | РОРα | RAR-связанный сиротский рецептор-α | ПЛЕВАТЬ | холестерин , АТРА | ||
| НР1Ф2 | РОРβ | RAR-связанный сиротский рецептор-β | РОРБ | |||||
| NR1F3 | РОРγ | RAR-связанный сиротский рецептор-γ | РОРК | |||||
| Г | CNR14-подобный (нематода) [50] | НР1Г1 | секс-1 | Рецептор стероидных гормонов cnr14 [53] | секс-1 | |||
| ЧАС | печени X- рецептор | NR1H1 | ЭКР | Рецептор экдизона, EcR (членистоногие) | ЭКР | экдистероиды | ||
| NR1H2 | LXRβ | Рецептор X печени-β | NR1H2 | оксистерины | ||||
| NR1H3 | LXRα | Рецептор X печени-α | NR1H3 | |||||
| NR1H4 | FXR | Рецептор фарнезоида X | NR1H4 | |||||
| NR1H5 [54] | FXR-б | Фарнезоид X-рецептор-β (псевдоген у человека) | NR1H5P | |||||
| я | Подобный рецептору витамина D | НР1И1 | ВДР | Рецептор витамина D | ВДР | витамин Д | ||
| НР1И2 | PXR | Рецептор прегнана X | НР1И2 | ксенобиотики | ||||
| НР1И3 | МАШИНА | Конститутивный рецептор андростана | НР1И3 | Андростан | ||||
| Дж | HR96-подобный [50] | NR1J1 | Гр96/ Даф-12 | Ядерный гормональный рецептор HR96 | 96 грн. | холестерин / дафахроновая кислота [55] | ||
| NR1J2 | ||||||||
| NR1J3 | ||||||||
| К | HR1-подобный [50] | НР1К1 | 1 час | Ядерный гормональный рецептор HR1 | ||||
| 2 | Ретиноид X-рецептор-подобный | А | Ядерный фактор гепатоцитов-4 | NR2A1 | HNF4α | Ядерный фактор гепатоцитов-4-α | ХНФ4А | жирные кислоты |
| НР2А2 | HNF4γ | Ядерный фактор гепатоцитов-4-γ | ХНФ4Г | |||||
| Б | Рецептор ретиноида X | NR2B1 | RXRα | Ретиноидный рецептор X-α | РХРА | ретиноиды | ||
| NR2B2 | RXRβ | Ретиноидный рецептор X-β | RXRB | |||||
| NR2B3 | RXRγ | Ретиноид X-рецептор-γ | RXRG | |||||
| NR2B4 | USP | Белок ультрадыхальца (членистоногие) | УСП | фосфолипиды [56] | ||||
| С | Тестикулярный рецептор | NR2C1 | ТР2 | Тестикулярный рецептор 2 | NR2C1 | |||
| NR2C2 | ТР4 | Тестикулярный рецептор 4 | NR2C2 | |||||
| И | TLX/PNR | NR2E1 | ТЛХ | Гомолог гена бесхвостой дрозофилы | NR2E1 | |||
| НР2Е3 | бронирование | Фоторецепторный ядерный рецептор, специфичный для клетки | НР2Е3 | |||||
| Ф | КУПЕР /EAR | НР2Ф1 | КУП-ТФИ | Вышестоящий промотор фактора транскрипции I куриного овальбумина | НР2Ф1 | |||
| НР2Ф2 | КУП-ТФII | Куриный овальбумин, расположенный выше промотора фактора транскрипции II | НР2Ф2 | ретиноевая кислота (слабая) [57] | ||||
| НР2Ф6 | УХ-2 | связанные с V-erbA | НР2Ф6 | |||||
| 3 | Подобно рецептору эстрогена | А | Рецептор эстрогена | NR3A1 | ЭРα | Рецептор эстрогена-α | СОЭ1 | эстрогены |
| NR3A2 | ЭРβ | Рецептор эстрогена-β | СОЭ2 | |||||
| Б | Эстрогенсвязанный рецептор | NR3B1 | ERRα | Эстрогенсвязанный рецептор-α | ЭСРРА | |||
| NR3B2 | ERRβ | Эстрогенсвязанный рецептор-β | ЭСРРБ | |||||
| NR3B3 | ЭРРγ | Эстрогенсвязанный рецептор-γ | ЭСРРГ | |||||
| С | 3-кетостероидные рецепторы | NR3C1 | ГР | Глюкокортикоидный рецептор | NR3C1 | кортизол | ||
| NR3C2 | МИСТЕР | Минералокортикоидный рецептор | NR3C2 | альдостерон | ||||
| NR3C3 | пиар | Рецептор прогестерона | РГР | прогестерон | ||||
| NR3C4 | АР | Андрогенный рецептор | АР | тестостерон | ||||
| Д | Подобно рецептору эстрогена (у лофотрохозозоа ) [58] | НР3Д | ||||||
| И | Подобно рецептору эстрогена (у книдарий ) [59] | НР3Е | ||||||
| Ф | Подобно рецептору эстрогена (у плакозоа ) [59] | НР3Ф | ||||||
| 4 | Фактор роста нервов IB-подобный | А | НГФИБ/НУРР1/НОР1 | NR4A1 | НГФИБ | Фактор роста нервов IB | NR4A1 | |
| NR4A2 | НУРР1 | Связанные с ядерными рецепторами 1 | NR4A2 | |||||
| NR4A3 | НОР1 | Орфанный рецептор 1 нейронного происхождения | NR4A3 | |||||
| 5 | стероидогенный Фактороподобный | А | СФ1/ЛРХ1 | NR5A1 | СФ1 | Стероидогенный фактор 1 | NR5A1 | фосфатидилинозитолы |
| НР5А2 | ЛРХ-1 | Гомолог печеночного рецептора-1 | НР5А2 | фосфатидилинозитолы | ||||
| Б | HR39-подобный | NR5B1 [50] | HR39/ ФТЗ-Ф1 | Рецептор ядерного гормона фуши таразу, фактор I бета | HR39 | |||
| 6 | Подобный ядерному фактору зародышевой клетки | А | ГКНФ | НР6А1 | ГКНФ | Ядерный фактор зародышевой клетки | НР6А1 | |
| 7 | NR с двумя ДНК-связывающими доменами [60] [50] [61] | А | 2DBD-NRα | NR7A1 | 2DBD-НРА2 | |||
| Б | 2DBD-NRβ | NR7B1 | 2DBD-НРА3 | |||||
| С | 2DBD-NRγ | NR7C1 | 2DBD-НРА1 | членистоногое "α/β" | ||||
| 8 | №8 [62] ( эуметазоа ) | А | НР8А | НР8А1 | CgNR8A1 | Ядерный рецептор 8 | AKG49571 | |
| 0 | Разное (отсутствует LBD или DBD) | А | кнр/кнрл/эгон [50] (членистоногие) | NR0A1 | КНИ | Белок зиготической щели tot | кнл | |
| Б | ДАКС/ШП | NR0B1 | DAX1 | Реверс пола, чувствительный к дозировке, критическая область гипоплазии надпочечников, на хромосоме X, ген 1 | NR0B1 | |||
| NR0B2 | МГП | Малый гетеродимерный партнер | NR0B2 | |||||
Из двух 0-семейств 0A имеет DBD, подобное семейству 1, а 0B имеет уникальный LBD. Второй ДБД семейства 7, вероятно, связан с ДБД семейства 1. Три, вероятно, NR семейства 1 из Biomphalaria glabrata обладают DBD наряду с LBD, подобным семейству 0B. [50] Расположение C. elegans nhr-1 ( Q21878 ) оспаривается: хотя большинство источников относят его к NR1K1, [50] ручная аннотация в WormBase считает его членом NR2A. [63] Раньше существовала группа 2D, единственным членом которой была Drosophila HR78/NR1D1 ( Q24142 ) и ортологи, но позже из-за высокого сходства она была объединена в группу 2C, образовав «группу 2C/D». [50] Нокаутные исследования на мышах и плодовых мушках подтверждают существование такой объединенной группы. [64]
Эволюция
[ редактировать ]Темой дебатов был вопрос о том, является ли предковый ядерный рецептор либо лиганд-связывающим, либо сиротским рецептором . Эти дебаты начались более двадцати пяти лет назад, когда первые лиганды были идентифицированы как стероиды млекопитающих и гормоны щитовидной железы. [65] Вскоре после этого идентификация рецептора экдизона у дрозофилы привела к идее, что ядерные рецепторы представляют собой гормональные рецепторы, которые связывают лиганды с наномолярным сродством. В то время тремя известными лигандами ядерных рецепторов были стероиды, ретиноиды и гормоны щитовидной железы, и из этих трех и стероиды, и ретиноиды были продуктами терпеноидного метаболизма. Таким образом, было предположено, что предковый рецептор был лигандирован молекулой терпеноида. [66]
В 1992 году сравнение ДНК-связывающего домена всех известных ядерных рецепторов привело к построению филогенного дерева ядерных рецепторов, которое показало, что все ядерные рецепторы имели общего предка. [67] В результате были предприняты дополнительные усилия по раскрытию состояния первого ядерного рецептора, и к 1997 году была предложена альтернативная гипотеза: предковый ядерный рецептор был сиротским рецептором и со временем приобрел способность связывать лиганды. [7] Эта гипотеза была выдвинута на основании следующих аргументов:
- Последовательности ядерных рецепторов, которые были идентифицированы у самых ранних многоклеточных животных (книдарий и шистосом ), все были членами групп рецепторов COUP-TF, RXR и FTZ-F1. И COUP-TF, и FTZ-F1 являются рецепторами-сиротами, а RXR связывает лиганд только у позвоночных. [68]
- В то время как орфанные рецепторы имели известные гомологи членистоногих, никаких ортологов лигандированных рецепторов позвоночных не было идентифицировано за пределами позвоночных, что позволяет предположить, что орфанные рецепторы старше, чем лигандированные рецепторы. [69]
- Орфанные рецепторы встречаются во всех шести подсемействах ядерных рецепторов, а лиганд-зависимые рецепторы встречаются среди трех. [7] Таким образом, поскольку считалось, что лиганд-зависимые рецепторы преимущественно относятся к недавним подсемействам, казалось логичным, что они приобрели способность независимо связывать лиганды.
- Филогенетическое положение данного ядерного рецептора внутри дерева коррелирует с его ДНК-связывающим доменом и способностями к димеризации, но не установлено связи между лиганд-зависимым ядерным рецептором и химической природой его лиганда. В дополнение к этому, эволюционные взаимоотношения между лиганд-зависимыми рецепторами не имели особого смысла, поскольку близкородственные рецепторы подсемейств связывали лиганды, происходящие из совершенно разных путей биосинтеза (например, TR и RAR). С другой стороны, подсемейства, которые эволюционно не связаны, связывают сходные лиганды (RAR и RXR связывают полностью транс- и 9-цис-ретиноевую кислоту соответственно). [69]
- В 1997 году было обнаружено, что ядерные рецепторы не существуют в статических конформациях «выключено» и «включено», но что лиганд может изменить равновесие между двумя состояниями. Более того, было обнаружено, что ядерные рецепторы могут регулироваться лиганд-независимым образом посредством фосфорилирования или других посттрансляционных модификаций. Таким образом, это обеспечило механизм того, как предковый орфанный рецептор регулировался лиганд-независимым образом, и объяснило, почему лиганд-связывающий домен консервативен. [69]
В течение следующих 10 лет проводились эксперименты для проверки этой гипотезы, и вскоре появились контраргументы:
- Ядерные рецепторы были идентифицированы в недавно секвенированном геноме демогубки Amphimedon Queenslandica , члена Porifera, самого древнего типа многоклеточных животных. Геном A. queenslandica содержит два ядерных рецептора, известные как AqNR1 и AqNR2, и оба они характеризуются способностью связываться и регулироваться лигандами. [70]
- Гомологи лиганд-зависимых рецепторов позвоночных были обнаружены вне позвоночных у моллюсков и Platyhelminthes. Более того, было обнаружено, что ядерные рецепторы, обнаруженные у книдарий, имеют структурные лиганды у млекопитающих, что может отражать наследственную ситуацию.
- С помощью структурного и масс-спектрометрического анализа было обнаружено, что два предполагаемых орфанных рецептора , HNF4 и USP , связывают жирные кислоты и фосфолипиды соответственно. [56]
- Ядерные рецепторы и лиганды оказались гораздо менее специфичными, чем считалось ранее. Ретиноиды могут связывать рецепторы млекопитающих, отличные от RAR и RXR, такие как PPAR, RORb или COUP-TFII. Кроме того, RXR чувствителен к широкому спектру молекул, включая ретиноиды, жирные кислоты и фосфолипиды. [71]
- Исследование эволюции стероидных рецепторов показало, что предковые стероидные рецепторы могли связывать лиганд эстрадиол. И наоборот, рецептор эстрогена, обнаруженный у моллюсков, конститутивно активен и не связывает эстроген-родственные гормоны. Таким образом, это стало примером того, как предковый лиганд-зависимый рецептор может потерять способность связывать лиганды. [72]
Сочетание этих недавних данных, а также углубленного изучения физической структуры лигандсвязывающего домена ядерного рецептора привело к появлению новой гипотезы относительно наследственного состояния ядерного рецептора. Эта гипотеза предполагает, что предковый рецептор может действовать как липидный сенсор со способностью связывать, хотя и довольно слабо, несколько различных гидрофобных молекул, таких как ретиноиды, стероиды, гемы и жирные кислоты. Благодаря своей способности взаимодействовать с различными соединениями этот рецептор посредством дупликации либо утрачивает способность к лиганд-зависимой активности, либо специализируется на высокоспецифичный рецептор для конкретной молекулы. [71]
История
[ редактировать ]Ниже приводится краткая подборка ключевых событий в истории исследований ядерных рецепторов. [73]
- 1905 – Эрнест Старлинг придумал слово «гормон».
- 1926 – Эдвард Кэлвин Кендалл и Тадеус Райхштейн выделили и определили структуру кортизона и тироксина.
- 1929 – Адольф Бутенандт и Эдвард Адельберт Дойзи – независимо выделили и определили структуру эстрогена.
- 1958 – Элвуд Дженсен – выделил рецептор эстрогена.
- 1980-е годы — клонирование рецепторов эстрогена, глюкокортикоидов и гормонов щитовидной железы Пьером Шамбоном , Рональдом Эвансом и Бьёрном Веннстремом соответственно.
- 2004 — Пьер Шамбон, Рональд Эванс и Элвуд Дженсен были награждены премией Альберта Ласкера за фундаментальные медицинские исследования , наградой, которая часто предшествует Нобелевской премии в области медицины.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б ПДБ : 3E00 ; Чандра В., Хуанг П., Хамуро Ю., Рагурам С., Ван Ю., Беррис Т.П., Растинеджад Ф. (ноябрь 2008 г.). «Структура интактного комплекса PPAR-гамма-RXR-ядерный рецептор на ДНК» . Природа . 456 (7220): 350–6. дои : 10.1038/nature07413 . ПМЦ 2743566 . ПМИД 19043829 .
- ^ Эванс Р.М. (май 1988 г.). «Суперсемейство рецепторов стероидных и тиреоидных гормонов» . Наука . 240 (4854): 889–95. Бибкод : 1988Sci...240..889E . дои : 10.1126/science.3283939 . ПМК 6159881 . ПМИД 3283939 .
- ^ Олефский Ю.М. (октябрь 2001 г.). «Серия миниобзоров ядерных рецепторов» . Журнал биологической химии . 276 (40): 36863–4. дои : 10.1074/jbc.R100047200 . ПМИД 11459855 . S2CID 5497175 .
- ^ Jump up to: а б с Мангельсдорф Д.Д., Таммел С., Беато М., Херрлих П., Шютц Г., Умесоно К., Блумберг Б., Кастнер П., Марк М., Шамбон П., Эванс Р.М. (декабрь 1995 г.). «Суперсемейство ядерных рецепторов: второе десятилетие» . Клетка . 83 (6): 835–9. дои : 10.1016/0092-8674(95)90199-X . ПМК 6159888 . ПМИД 8521507 .
- ^ Jump up to: а б Новак Н., Хайнцель Т. (декабрь 2004 г.). «Ядерные рецепторы: обзор и классификация». Текущие цели по борьбе с наркотиками. Воспаление и аллергия . 3 (4): 335–46. дои : 10.2174/1568010042634541 . ПМИД 15584884 .
- ^ Jump up to: а б с Комитет по номенклатуре ядерных рецепторов (апрель 1999 г.). «Единая система номенклатуры суперсемейства ядерных рецепторов» . Клетка . 97 (2): 161–3. дои : 10.1016/S0092-8674(00)80726-6 . ПМИД 10219237 . S2CID 36659104 .
- ^ Jump up to: а б с д Лаудет V (декабрь 1997 г.). «Эволюция суперсемейства ядерных рецепторов: ранняя диверсификация от предкового рецептора-сироты». Журнал молекулярной эндокринологии . 19 (3): 207–26. дои : 10.1677/jme.0.0190207 . ПМИД 9460643 . S2CID 16419929 .
- ^ Эскрива Х., Ланглуа М.К., Мендонса Р.Л., Пирс Р., Лаудет В. (май 1998 г.). «Эволюция и диверсификация суперсемейства ядерных рецепторов». Анналы Нью-Йоркской академии наук . 839 (1): 143–6. Бибкод : 1998NYASA.839..143E . дои : 10.1111/j.1749-6632.1998.tb10747.x . ПМИД 9629140 . S2CID 11164838 .
- ^ Райтцель А.М., Панг К., Райан Дж.Ф., Малликин Дж.К., Мартиндейл М.К., Баксеванис А.Д., Таррант А.М. (февраль 2011 г.). «Ядерные рецепторы гребневика Mnemiopsis leidyi лишены ДНК-связывающего домена с цинковыми пальцами: специфичная для линии потеря или наследственное состояние при возникновении суперсемейства ядерных рецепторов?» . ЭвоДево . 2 (1): 3. дои : 10.1186/2041-9139-2-3 . ПМК 3038971 . ПМИД 21291545 .
- ^ Бриджэм Дж.Т., Эйк Г.Н., Ларру С., Дешпанде К., Хармс М.Дж., Готье М.Е., Ортлунд Э.А., Дегнан Б.М., Торнтон Дж.В. (октябрь 2010 г.). «Эволюция белков путем молекулярных манипуляций: диверсификация суперсемейства ядерных рецепторов от лиганд-зависимого предка» . ПЛОС Биология . 8 (10): e1000497. дои : 10.1371/journal.pbio.1000497 . ПМК 2950128 . ПМИД 20957188 .
- ^ Слудер А.Э., Майна К.В. (апрель 2001 г.). «Ядерные рецепторы нематод: темы и вариации». Тенденции в генетике . 17 (4): 206–13. дои : 10.1016/S0168-9525(01)02242-9 . ПМИД 11275326 .
- ^ Читл Джарвела AM, Pick L (2017). «Функция и эволюция ядерных рецепторов в эмбриональном развитии насекомых». Актуальные темы биологии развития . 125 : 39–70. дои : 10.1016/bs.ctdb.2017.01.003 . ISBN 9780128021729 . ПМИД 28527580 .
- ^ Шааф М.Ю. (2017). «Исследование ядерных рецепторов у рыбок данио» . Журнал молекулярной эндокринологии . 59 (1): Р65–Р76. doi : 10.1530/JME-17-0031 . ПМИД 28438785 .
- ^ Jump up to: а б Чжан З., Берч П.Е., Куни А.Дж., Ланц Р.Б., Перейра Ф.А., Ву Дж., Гиббс Р.А., Вайнсток Г., Уилер Д.А. (апрель 2004 г.). «Геномный анализ семейства ядерных рецепторов: новое понимание структуры, регуляции и эволюции генома крысы» . Геномные исследования . 14 (4): 580–90. дои : 10.1101/гр.2160004 . ПМЦ 383302 . ПМИД 15059999 .
- ^ Оверингтон Дж. П., Аль-Лазикани Б., Хопкинс А. Л. (декабрь 2006 г.). «Сколько существует целей по борьбе с наркотиками?». Обзоры природы. Открытие наркотиков . 5 (12): 993–6. дои : 10.1038/nrd2199 . ПМИД 17139284 . S2CID 11979420 .
- ^ Бенуа Дж., Куни А., Жигер В., Ингрэм Х. , Лазар М., Маскат Г., Перлманн Т., Рено Дж. П., Швабе Дж., Сладек Ф., Цай М. Дж. , Лаудет В. (декабрь 2006 г.). «Международный союз фармакологии. LXVI. Орфанные ядерные рецепторы». Фармакологические обзоры . 58 (4): 798–836. дои : 10.1124/пр.58.4.10 . ПМИД 17132856 . S2CID 2619263 .
- ^ Мохан Р., Хейман Р.А. (2003). «Модуляторы орфанных ядерных рецепторов». Актуальные темы медицинской химии . 3 (14): 1637–47. дои : 10.2174/1568026033451709 . ПМИД 14683519 .
- ^ Кумар Р., Томпсон Э.Б. (май 1999 г.). «Строение ядерных рецепторов гормонов». Стероиды . 64 (5): 310–9. дои : 10.1016/S0039-128X(99)00014-8 . ПМИД 10406480 . S2CID 18333397 .
- ^ Клинге CM (май 2000 г.). «Взаимодействие рецептора эстрогена с коактиваторами и корепрессорами». Стероиды . 65 (5): 227–51. дои : 10.1016/S0039-128X(99)00107-5 . ПМИД 10751636 . S2CID 41160722 .
- ^ Jump up to: а б Вернмарк А., Трейтер Э., Райт А.П., Густафссон Дж.А. (октябрь 2003 г.). «Функции активации 1 и 2 ядерных рецепторов: молекулярные стратегии активации транскрипции» . Молекулярная эндокринология . 17 (10): 1901–9. дои : 10.1210/me.2002-0384 . ПМИД 12893880 . S2CID 31314461 .
- ^ Ву В, ЛоВерде PT (2021). «Идентификация и эволюция ядерных рецепторов у платихельминтов» . ПЛОС ОДИН . 16(8): e0250750 (8): e0250750. Бибкод : 2021PLoSO..1650750W . дои : 10.1371/journal.pone.0250750 . ПМК 8363021 . ПМИД 34388160 .
- ^ Weatherman RV, Флеттерик Р.Дж., Сканлан Т.С. (1999). «Лиганды ядерных рецепторов и лигандсвязывающие домены». Ежегодный обзор биохимии . 68 : 559–81. doi : 10.1146/annurev.biochem.68.1.559 . ПМИД 10872460 .
- ^ Чандра В., Хуанг П., Потлури Н., Ву Д., Ким Й., Растинежад Ф. (март 2013 г.). «Мультидоменная интеграция в структуру ядерного рецепторного комплекса HNF-4α» . Природа . 495 (7441): 394–8. Бибкод : 2013Natur.495..394C . дои : 10.1038/nature11966 . ПМК 3606643 . ПМИД 23485969 .
- ^ Лу X, Торессон Дж., Бенод С., Су Дж.Х., Филипс К.Дж., Уэбб П., Густафссон Дж.А. (март 2014 г.). «Структура гетеродимера ретиноидного рецептора X α-рецептора X печени β (RXRα-LXRβ) на ДНК». Структурная и молекулярная биология природы . 21 (3): 277–81. дои : 10.1038/nsmb.2778 . ПМИД 24561505 . S2CID 23226682 .
- ^ ВВП : 2C7A ; Ремер С.К., Донхэм, округ Колумбия, Шерман Л., Пон В.Х., Эдвардс Д.П., Черчилль М.Е. (декабрь 2006 г.). «Структура комплекса рецептор прогестерона-дезоксирибонуклеиновая кислота: новые взаимодействия, необходимые для связывания с элементами ответа половинного участка» . Молекулярная эндокринология . 20 (12): 3042–52. дои : 10.1210/me.2005-0511 . ПМЦ 2532839 . ПМИД 16931575 .
- ^ ПДБ : 3L0L ; Джин Л., Мартыновски Д., Чжэн С., Вада Т., Се В., Ли Ю. (май 2010 г.). «Структурная основа гидроксихолестеринов как природных лигандов сиротского ядерного рецептора RORgamma» . Молекулярная эндокринология . 24 (5): 923–9. дои : 10.1210/me.2009-0507 . ПМЦ 2870936 . ПМИД 20203100 .
- ^ Jump up to: а б с д Амуциас Г.Д., Пихлер Э.Э., Миан Н., Де Грааф Д., Имсириду А., Робинсон-Рехави М., Борнберг-Бауэр Э., Робертсон Д.Л., Оливер С.Г. (июль 2007 г.). «Атлас белкового взаимодействия ядерных рецепторов: свойства и качество сети димеризации на основе узлов» . Системная биология BMC . 1:34 . дои : 10.1186/1752-0509-1-34 . ПМК 1971058 . ПМИД 17672894 .
- ^ Линья М.Ю., Поркка К.П., Канг З., Савинайнен К.Дж., Янне О.А., Таммела Т.Л., Весселла Р.Л., Палвимо Дж.Дж., Висакорпи Т. (февраль 2004 г.). «Экспрессия корегуляторов андрогенных рецепторов при раке простаты». Клинические исследования рака . 10 (3): 1032–40. дои : 10.1158/1078-0432.CCR-0990-3 . ПМИД 14871982 . S2CID 8038717 .
- ^ Клинге С.М., Боденнер Д.Л., Десаи Д., Найлз Р.М., Трейш А.М. (май 1997 г.). «Связывание ядерных рецепторов типа II и рецептора эстрогена с элементами полного и половинного эстрогенового ответа in vitro» . Исследования нуклеиновых кислот . 25 (10): 1903–12. дои : 10.1093/нар/25.10.1903 . ПМК 146682 . ПМИД 9115356 .
- ^ Руаль, Жан-Франсуа; Венкатесан, Кавита; Хао, Тонг; Хиродзанэ-Кисикава, Томоко; Дрико, Амели; Оболочка; Беррис, Габриэль Ф.; Гиббонс, Фрэнсис Д.; Дрезе, Матия; Айиви-Гедеуссу, Ноно; Клитгорд, Нильс (20 октября 2005 г.). «К карте белково-белковых взаимодействий человека в масштабе протеома» . Природа . 437 (7062): 1173–1178. Бибкод : 2005Natur.437.1173R . дои : 10.1038/nature04209 . ISSN 1476-4687 . ПМИД 16189514 . S2CID 4427026 .
- ^ Альберс, Майкл; Кранц, Харальд; Кобер, Инго; Кайзер, Кармен; Клинк, Мартин; Суков, Йорг; Керн, Райнер; Кегль, Манфред (февраль 2005 г.). «Автоматизированный двухгибридный скрининг дрожжей на наличие белков, взаимодействующих с ядерными рецепторами» . Молекулярная и клеточная протеомика . 4 (2): 205–213. дои : 10.1074/mcp.M400169-MCP200 . ISSN 1535-9476 . ПМИД 15604093 . S2CID 14876486 .
- ^ Маккенна, Нью-Джерси, Ланц Р.Б., О'Мэлли Б.В. (июнь 1999 г.). «Корегуляторы ядерных рецепторов: клеточная и молекулярная биология» . Эндокринные обзоры . 20 (3): 321–344. дои : 10.1210/edrv.20.3.0366 . PMID 10368774 . S2CID 10182146 .
- ^ Гласс С.К., Розенфельд М.Г. (январь 2000 г.). «Корегуляторный обмен в транскрипционных функциях ядерных рецепторов» . Гены и развитие . 14 (2): 121–41. дои : 10.1101/gad.14.2.121 . ПМИД 10652267 . S2CID 12793980 .
- ^ Аранда А., Паскуаль А. (июль 2001 г.). «Рецепторы ядерных гормонов и экспрессия генов». Физиологические обзоры . 81 (3): 1269–304. дои : 10.1152/physrev.2001.81.3.1269 . hdl : 10261/79944 . ПМИД 11427696 . S2CID 5972234 .
- ^ Копленд Дж.А., Шеффилд-Мур М., Колджич-Зиванович Н., Джентри С., Лампроу Г., Цорцату-Статопулу Ф., Зумпурлис В., Урбан Р.Дж., Влахопулос С.А. (июнь 2009 г.). «Рецепторы половых стероидов при дифференцировке скелета и эпителиальной неоплазии: возможно ли тканеспецифическое вмешательство?». Биоэссе . 31 (6): 629–41. doi : 10.1002/bies.200800138 . ПМИД 19382224 . S2CID 205469320 .
- ^ Бжозовский А.М., Пайк А.К., Даутер З., Хаббард Р.Э., Бонн Т., Энгстрем О., Оман Л., Грин Г.Л., Густафссон Дж.А., Карлквист М. (октябрь 1997 г.). «Молекулярные основы агонизма и антагонизма рецептора эстрогена». Природа . 389 (6652): 753–8. Бибкод : 1997Natur.389..753B . дои : 10.1038/39645 . ПМИД 9338790 . S2CID 4430999 .
- ^ Шиау А.К., Барстад Д., Лория П.М., Ченг Л., Кушнер П.Дж., Агард Д.А., Грин Г.Л. (декабрь 1998 г.). «Структурная основа распознавания рецептора/коактиватора эстрогена и антагонизм этого взаимодействия тамоксифена» . Клетка . 95 (7): 927–37. дои : 10.1016/S0092-8674(00)81717-1 . ПМИД 9875847 . S2CID 10265320 .
- ^ Гронемейер Х., Густафссон Дж.А., Лаудет В. (ноябрь 2004 г.). «Принципы модуляции суперсемейства ядерных рецепторов». Обзоры природы. Открытие наркотиков . 3 (11): 950–64. дои : 10.1038/nrd1551 . ПМИД 15520817 . S2CID 205475111 .
- ^ Буш Б.Б., Стивенс В.К., Мартин Р., Ордентлих П., Чжоу С., Сапп Д.В., Хорлик Р.А., Мохан Р. (ноябрь 2004 г.). «Идентификация селективного обратного агониста альфа-рецептора сиротского ядерного рецептора, связанного с эстрогеном». Журнал медицинской химии . 47 (23): 5593–6. дои : 10.1021/jm049334f . ПМИД 15509154 .
- ^ Смит CL, О'Мэлли BW (февраль 2004 г.). «Функция корегулятора: ключ к пониманию тканевой специфичности селективных модуляторов рецепторов» . Эндокринные обзоры . 25 (1): 45–71. дои : 10.1210/er.2003-0023 . ПМИД 14769827 .
- ^ Паскуаль Дж., Гласс С.К. (октябрь 2006 г.). «Ядерные рецепторы против воспаления: механизмы трансрепрессии». Тенденции в эндокринологии и обмене веществ . 17 (8): 321–7. дои : 10.1016/j.tem.2006.08.005 . ПМИД 16942889 . S2CID 19612552 .
- ^ Бьорнстрем Л., Сьоберг М. (июнь 2004 г.). «Эстрогензависимая рецептор-зависимая активация AP-1 посредством негеномной передачи сигналов» . Ядерный рецептор . 2 (1): 3. дои : 10.1186/1478-1336-2-3 . ПМК 434532 . ПМИД 15196329 .
- ^ Живадинович Д., Гаметчу Б., Уотсон К.С. (2005). «Уровни мембранного рецептора эстрогена-альфа в клетках рака молочной железы MCF-7 предсказывают реакцию цАМФ и пролиферацию» . Исследование рака молочной железы . 7 (1): Р101–12. дои : 10.1186/bcr958 . ПМК 1064104 . ПМИД 15642158 .
- ^ Кустени С., Беллидо Т., Плоткин Л.И., О'Брайен К.А., Боденнер Д.Л., Хан Л., Хан К., ДиГрегорио ГБ, Каценелленбоген Дж.А., Каценелленбоген Б.С., Роберсон П.К. , Вайнштейн Р.С., Джилка Р.Л., Манолагас СК (март 2001 г.). «Ненгенотропная, неспецифическая для пола передача сигналов через рецепторы эстрогена или андрогена: диссоциация от транскрипционной активности» . Клетка . 104 (5): 719–30. дои : 10.1016/S0092-8674(01)00268-9 . ПМИД 11257226 . S2CID 10642274 .
- ^ Стори Н.М., Джентиле С., Улла Х., Руссо А., Мюссель М., Эркслебен С., Армстронг Д.Л. (март 2006 г.). «Быстрая передача сигналов на плазматическую мембрану ядерным рецептором гормона щитовидной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (13): 5197–201. Бибкод : 2006PNAS..103.5197S . дои : 10.1073/pnas.0600089103 . ПМЦ 1458817 . ПМИД 16549781 .
- ^ Стори Н.М., О'Брайан Дж.П., Армстронг Д.Л. (январь 2002 г.). «Rac и Rho опосредуют противоположную гормональную регуляцию калиевого канала, связанного с эфиром» . Современная биология . 12 (1): 27–33. дои : 10.1016/S0960-9822(01)00625-X . ПМИД 11790300 . S2CID 8608805 .
- ^ Jump up to: а б Мартин Н.П., Маррон Фернандес де Веласко Э., Мизуно Ф., Скаппини Э.Л., Глосс Б., Эркслебен С., Уильямс Дж.Г., Стэплтон Х.М., Джентиле С., Армстронг Д.Л. (сентябрь 2014 г.). «Быстрый цитоплазматический механизм регуляции киназы PI3 с помощью ядерного рецептора гормона щитовидной железы, TRβ, и генетические доказательства его роли в созревании синапсов гиппокампа мыши in vivo» . Эндокринология . 155 (9): 3713–24. дои : 10.1210/en.2013-2058 . ПМК 4138568 . ПМИД 24932806 .
- ^ Гилберт М.Е. (январь 2004 г.). «Изменения в синаптической передаче и пластичности в области CA1 гиппокампа взрослых после гипотиреоза развития» . Исследования мозга. Исследования развития мозга . 148 (1): 11–8. дои : 10.1016/j.devbrainres.2003.09.018 . ПМИД 14757514 .
- ^ Мёллер Л.С., Брокер-Пройсс М. (август 2011 г.). «Регуляция транскрипции неклассическим действием гормона щитовидной железы» . Исследование щитовидной железы . 4 (Приложение 1): S6. дои : 10.1186/1756-6614-4-S1-S6 . ПМК 3155112 . ПМИД 21835053 .
- ^ Jump up to: а б с д и ж г час я дж к Каур С., Джоблинг С., Джонс К.С., Нобл Л.Р., Рутледж Э.Дж., Локьер А.Э. (7 апреля 2015 г.). «Ядерные рецепторы Biomphalaria glabrata и Lottia gigantea: значение для разработки новых модельных организмов» . ПЛОС ОДИН . 10 (4): e0121259. Бибкод : 2015PLoSO..1021259K . дои : 10.1371/journal.pone.0121259 . ПМЦ 4388693 . ПМИД 25849443 .
- ^ Беррис Т.П., де Вера ИМ, Кот I, Флавени К.А., Ваннинаяке США, Чаттерджи А., Уокер Дж.К., Штайнауэр Н., Чжан Дж., Кунс Л.А., Корах К.С., Каин Д.В., Холленберг А.Н., Уэбб П., Форрест Д., Джеттен А.М., Эдвардс Д.П., Гримм С.Л., Хартиг С., Ланге К.А., Ричер Дж.К., Сарториус К.А., Тетель М., Биллон С., Элгенди Б., Хегази Л., Гриффетт К., Пейнетти Н., Бернштейн К.Л., Хьюз Т.С., Ситаула С., Стейрук К.Р., Калвер А, Мюррей М.Х., Финк Б.Н., Цидловски Дж.А. (ноябрь 2023 г.). Ольштейн Э (ред.). «Международный союз фундаментальной и клинической фармакологии CXIII: Суперсемейство ядерных рецепторов — обновление 2023 г.» . Фармакологические обзоры . 75 (6): 1233–1318. дои : 10.1124/pharmrev.121.000436 . ISSN 0031-6997 . ПМЦ 10595025 . PMID 37586884 .
- ^ Кроссгроув К., Лаудет В., Майна К.В. (февраль 2002 г.). «Dirofilaria immitis кодирует Di-nhr-7, предполагаемый ортолог гена E78, регулируемого экдизоном дрозофилы». Молекулярная и биохимическая паразитология . 119 (2): 169–77. дои : 10.1016/s0166-6851(01)00412-1 . ПМИД 11814569 .
- ^ «секс-1 (ген)» . WormBase: Информационный ресурс по нематодам .
- ^ Отте К., Кранц Х., Кобер И., Томпсон П., Хоефер М., Хаубольд Б., Реммель Б., Восс Х., Кайзер С., Альберс М., Черуваллат З., Джексон Д., Казари Г., Коегль М., Паабо С., Мус Дж., Кремозер К. , Дойшле У (февраль 2003 г.). «Идентификация бета-рецептора фарнезоида X как нового ядерного рецептора млекопитающих, чувствительных к ланостерину» . Молекулярная и клеточная биология . 23 (3): 864–72. doi : 10.1128/mcb.23.3.864-872.2003 . ПМК 140718 . ПМИД 12529392 .
- ^ «Отчет о генах FlyBase: Dmel\Hr96» . ФлайБейс . Проверено 14 августа 2019 г.
- ^ Jump up to: а б Швабе Й.В., Тейхманн С.А. (январь 2004 г.). «Ядерные рецепторы: эволюция разнообразия». СТКЭ науки . 2004 (217): пе4. дои : 10.1126/stke.2172004pe4 . ПМИД 14747695 . S2CID 20835274 .
- ^ Крузе С.В., Суино-Пауэлл К., Чжоу Х.Е., Кречман Дж.Е., Рейнольдс Р., Фонрейн С. и др. (сентябрь 2008 г.). «Идентификация орфанного ядерного рецептора COUP-TFII как рецептора, активируемого ретиноевой кислотой» . ПЛОС Биология . 6 (9): е227. дои : 10.1371/journal.pbio.0060227 . ПМЦ 2535662 . ПМИД 18798693 .
- ^ Марков Г.В., Гутьеррес-Мазариегос Дж., Питрат Д., Биллас И.М., Боннетон Ф., Морас Д. и др. (март 2017 г.). «Происхождение древней пары гормон/рецептор, выявленное путем воскрешения предкового эстрогена» . Достижения науки . 3 (3): e1601778. Бибкод : 2017SciA....3E1778M . дои : 10.1126/sciadv.1601778 . ПМЦ 5375646 . ПМИД 28435861 .
- ^ Jump up to: а б Халтурин К., Биллас И., Чебаро Ю., Райтцель А.М., Таррант А.М., Лаудет В., Марков Г.В. (ноябрь 2018 г.). «Рецепторы NR3E у книдарий: новое семейство родственников стероидных рецепторов расширяет возможные механизмы связывания лигандов» . J Стероид Биохим Мол Биол . 184 : 11–19. дои : 10.1016/j.jsbmb.2018.06.014 . ПМК 6240368 . ПМИД 29940311 .
- ^ Ву, В., ЛоВерде PT (сентябрь 2023 г.). «Обновленные знания и предлагаемая номенклатура ядерных рецепторов с двумя ДНК-связывающими доменами (2DBD-NR)» . ПЛОС ОДИН . 18 (9): e0286107. Бибкод : 2023PLoSO..1886107W . дои : 10.1371/journal.pone.0286107 . ПМЦ 10497141 . ПМИД 37699039 .
- ^ Ву В., Найлз Э.Г., Хираи Х., ЛоВерде П.Т. (февраль 2007 г.). «Эволюция нового подсемейства ядерных рецепторов, члены которого содержат два ДНК-связывающих домена» . БМК Эвол Биол . 7 (27): 27. дои : 10.1186/1471-2148-7-27 . ПМК 1810520 . ПМИД 17319953 .
- ^ Хуан В., Сюй Ф., Ли Дж., Ли Л., Цюэ Х., Чжан Г. (август 2015 г.). «Эволюция нового подсемейства ядерных рецепторов с акцентом на представителя тихоокеанской устрицы Crassostrea gigas». Джин . 567 (2): 164–72. дои : 10.1016/j.gene.2015.04.082 . ПМИД 25956376 .
- ^ «nhr-1 (ген)» . WormBase: Информационный ресурс по нематодам .
- ^ Марксрайтер С., Таммел К.С. (февраль 2018 г.). «Функции взрослых для ядерного рецептора DHR78 дрозофилы» . Динамика развития . 247 (2): 315–322. дои : 10.1002/dvdy.24608 . ПМК 5771960 . ПМИД 29171103 .
- ^ Эванс Р.М. (май 1988 г.). «Суперсемейство рецепторов стероидных и тиреоидных гормонов» . Наука . 240 (4854): 889–95. Бибкод : 1988Sci...240..889E . дои : 10.1126/science.3283939 . ПМК 6159881 . ПМИД 3283939 .
- ^ Мур Д.Д. (январь 1990 г.). «Разнообразие и единство ядерных рецепторов гормонов: суперсемейство терпеноидных рецепторов». Новый биолог . 2 (1): 100–5. ПМИД 1964083 .
- ^ Лаудет В., Ханни С., Колл Дж., Катцефлис Ф., Стехелин Д. (март 1992 г.). «Эволюция суперсемейства генов ядерных рецепторов» . Журнал ЭМБО . 11 (3): 1003–13. дои : 10.1002/j.1460-2075.1992.tb05139.x . ПМК 556541 . ПМИД 1312460 .
- ^ Эскрива Х., Сафи Р., Ханни С., Ланглуа М.С., Саумиту-Лапрад П., Стехелин Д., Капрон А., Пирс Р., Лаудет В. (июнь 1997 г.). «Связывание лиганда было приобретено в ходе эволюции ядерных рецепторов» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (13): 6803–8. Бибкод : 1997PNAS...94.6803E . дои : 10.1073/pnas.94.13.6803 . ПМК 21239 . ПМИД 9192646 .
- ^ Jump up to: а б с Эскрива Х., Делоне Ф., Лоде В. (август 2000 г.). «Связывание лигандов и эволюция ядерных рецепторов». Биоэссе . 22 (8): 717–27. doi : 10.1002/1521-1878(200008)22:8<717::AID-BIES5>3.0.CO;2-I . ПМИД 10918302 . S2CID 45891497 .
- ^ Бриджэм Дж.Т., Эйк Г.Н., Ларру С., Дешпанде К., Хармс М.Дж., Готье М.Е., Ортлунд Э.А., Дегнан Б.М., Торнтон Дж.В. (октябрь 2010 г.). «Эволюция белков путем молекулярных манипуляций: диверсификация суперсемейства ядерных рецепторов от лиганд-зависимого предка» . ПЛОС Биология . 8 (10): e1000497. дои : 10.1371/journal.pbio.1000497 . ПМК 2950128 . ПМИД 20957188 .
- ^ Jump up to: а б Марков Г.В., Лаудет В (март 2011 г.). «Происхождение и эволюция лигандсвязывающей способности ядерных рецепторов». Молекулярная и клеточная эндокринология . Эволюция рецепторов ядерных гормонов. 334 (1–2): 21–30. дои : 10.1016/j.mce.2010.10.017 . ПМИД 21055443 . S2CID 33537979 .
- ^ Торнтон Дж.В., Need E, Crews D (сентябрь 2003 г.). «Возрождение предкового стероидного рецептора: древнее происхождение передачи сигналов эстрогена». Наука . 301 (5640): 1714–7. Бибкод : 2003Sci...301.1714T . дои : 10.1126/science.1086185 . ПМИД 14500980 . S2CID 37628350 .
- ^ Тата-младший (июнь 2005 г.). «Сто лет гормонов» . Отчеты ЭМБО . 6 (6): 490–6. дои : 10.1038/sj.embor.7400444 . ПМЦ 1369102 . ПМИД 15940278 .
Внешние ссылки
[ редактировать ]- Ядерные + рецепторы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Винсент Лоде (2006). «Сборник фармакологии и классификации суперсемейства ядерных рецепторов IUPHAR 2006E» . Справочник ядерных рецепторов . Международный союз фундаментальной и клинической фармакологии. Архивировано из оригинала 02 апреля 2015 г. Проверено 21 февраля 2008 г.
- « « Ядерные рецепторы Интернет-журнал » . Домашняя страница . опубликовано BioMed Central (с мая 2007 г. материалы больше не принимаются) . Проверено 21 февраля 2008 г.
- «Ресурс ядерных рецепторов» . Джорджтаунский университет. Архивировано из оригинала 11 мая 2008 г. Проверено 21 февраля 2008 г.
- «Атлас сигналов ядерных рецепторов (рецепторы, коактиваторы, корепрессоры и лиганды)» . Консорциум НУРСА . Проверено 21 февраля 2008 г.
исследовательский консорциум и база данных, финансируемые НИЗ; включает журнал с открытым доступом, индексируемый PubMed, «Сигнализация ядерных рецепторов».
- «Ресурс ядерных рецепторов» . Джек Ванден Хьювел . Проверено 21 сентября 2009 г.
| Базы данных органов управления : Национальные |
|---|