Коактиватор (генетика)
Коактиватор — это тип корегулятора транскрипции который связывается с активатором ( фактором транскрипции ) для увеличения скорости транскрипции гена , или набора генов. [ 1 ] Активатор содержит ДНК-связывающий домен , который связывается либо с участком промотора ДНК , либо со специфической регуляторной последовательностью ДНК, называемой энхансером . [ 2 ] [ 3 ] Связывание комплекса активатор-коактиватор увеличивает скорость транскрипции за счет привлечения к промотору общего механизма транскрипции, тем самым увеличивая экспрессию генов . [ 3 ] [ 4 ] [ 5 ] Использование активаторов и коактиваторов позволяет обеспечить высокоспецифичную экспрессию определенных генов в зависимости от типа клеток и стадии развития. [ 2 ]
Некоторые коактиваторы также обладают активностью гистон-ацетилтрансферазы (HAT). HAT образуют большие мультипротеиновые комплексы, которые ослабляют связь гистонов с ДНК путем ацетилирования N -концевого гистонового хвоста. Это предоставляет больше места транскрипционному аппарату для связывания с промотором, тем самым увеличивая экспрессию генов. [ 1 ] [ 4 ]
Активаторы обнаружены во всех живых организмах , но белки-коактиваторы обычно обнаруживаются только у эукариот, поскольку они более сложны и требуют более сложного механизма регуляции генов. [ 1 ] [ 4 ] У эукариот коактиваторами обычно являются белки, локализованные в ядре. [ 1 ] [ 6 ]
Механизм
[ редактировать ]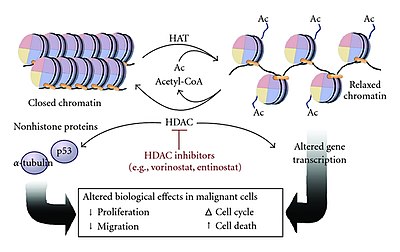
Некоторые коактиваторы косвенно регулируют экспрессию генов, связываясь с активатором и вызывая конформационные изменения , которые затем позволяют активатору связываться с последовательностью энхансера или промотора ДНК. [ 2 ] [ 7 ] [ 8 ] Как только комплекс активатор-коактиватор связывается с энхансером, РНК-полимераза II и другие общие механизмы транскрипции привлекаются к ДНК, и начинается транскрипция. [ 9 ]
Гистон ацетилтрансфераза
[ редактировать ]Ядерная ДНК обычно плотно обернута вокруг гистонов, что затрудняет или делает невозможным доступ транскрипционного аппарата к ДНК. Эта ассоциация обусловлена прежде всего электростатическим притяжением между ДНК и гистонами, поскольку фосфатный остов ДНК заряжен отрицательно, а гистоны богаты остатками лизина, которые заряжены положительно. [ 10 ] Тесная ассоциация ДНК-гистонов предотвращает транскрипцию ДНК в РНК.
Многие коактиваторы обладают активностью гистон-ацетилтрансферазы (HAT), что означает, что они могут ацетилировать определенные остатки лизина на N-концевых хвостах гистонов. [ 4 ] [ 7 ] [ 11 ] В этом методе активатор связывается с сайтом энхансера и рекрутирует комплекс HAT, который затем ацетилирует гистоны, связанные с нуклеосомным промотором, путем нейтрализации положительно заряженных остатков лизина. [ 7 ] [ 11 ] Эта нейтрализация заряда приводит к тому, что гистоны имеют более слабую связь с отрицательно заряженной ДНК, что расслабляет структуру хроматина, позволяя другим факторам транскрипции или механизмам транскрипции связываться с промотором (инициация транскрипции). [ 4 ] [ 11 ] Ацетилирование комплексами HAT может также помочь сохранить хроматин открытым на протяжении всего процесса элонгации, увеличивая скорость транскрипции. [ 4 ]

Ацетилирование N-концевого хвоста гистонов является одной из наиболее распространенных модификаций белков, обнаруженных у эукариот, при этом около 85% всех белков человека ацетилируются. [ 12 ] Ацетилирование имеет решающее значение для синтеза, стабильности, функции, регуляции и локализации белков и транскриптов РНК. [ 11 ] [ 12 ]
HAT действуют аналогично N-концевым ацетилтрансферазам (NAT), но их ацетилирование обратимо, в отличие от NAT. [ 13 ] HAT-опосредованное ацетилирование гистонов обратимо с помощью деацетилазы гистонов (HDAC), которая катализирует гидролиз остатков лизина, удаляя ацетильную группу из гистонов. [ 4 ] [ 7 ] [ 11 ] Это приводит к тому, что хроматин снова закрывается из расслабленного состояния, что затрудняет связывание транскрипционного аппарата с промотором, тем самым подавляя экспрессию генов. [ 4 ] [ 7 ]
Примеры коактиваторов, проявляющих активность HAT, включают CARM1 , CBP и EP300 . [ 14 ] [ 15 ]
Корепрессия
[ редактировать ]Многие коактиваторы также действуют как корепрессоры . при определенных обстоятельствах [ 5 ] [ 9 ] Кофакторы, такие как TAF1 и BTAF1, могут инициировать транскрипцию в присутствии активатора (действовать как коактиватор) и подавлять базальную транскрипцию в отсутствие активатора (действовать как корепрессор). [ 9 ]
Значение
[ редактировать ]Биологическое значение
[ редактировать ]Регуляция транскрипции — один из наиболее распространенных способов изменения экспрессии генов в организме. [ 16 ] Использование активации и коактивации позволяет лучше контролировать, когда, где и сколько белка производится. [ 1 ] [ 7 ] [ 16 ] Это позволяет каждой клетке быстро реагировать на изменения окружающей среды или физиологические изменения и помогает смягчить любой ущерб, который может возникнуть, если бы он не регулировался иным образом. [ 1 ] [ 7 ]
Сопутствующие расстройства
[ редактировать ]Мутации генов-коактиваторов, приводящие к потере или усилению функции белка, связаны с такими заболеваниями и расстройствами, как врожденные дефекты , рак (особенно гормонозависимый рак), нарушения развития нервной системы и умственная отсталость (ИН), среди многих других. [ 17 ] [ 5 ] Нарушение регуляции, приводящее к чрезмерной или недостаточной экспрессии коактиваторов, может пагубно взаимодействовать со многими лекарствами (особенно антигормональными препаратами) и связано с раком, проблемами фертильности , а также нарушениями развития нервной системы и нервно-психическими расстройствами . [ 5 ] Конкретный пример: нарушение регуляции CREB-связывающего белка (CBP), который действует как коактиватор многочисленных факторов транскрипции в центральной нервной системе (ЦНС), репродуктивной системе, тимусе и почках, связано с болезнью Хантингтона , лейкемией , Рубинштейном. -Синдром Тайби , нарушения нервно-психического развития и дефициты иммунной системы , кроветворения и скелетных мышц . функции [ 14 ] [ 18 ]
В качестве мишени для наркотиков
[ редактировать ]Коактиваторы являются многообещающими мишенями для лекарственной терапии при лечении рака, нарушений обмена веществ , сердечно-сосудистых заболеваний и диабета 2 типа , а также многих других заболеваний. [ 5 ] [ 19 ] Например, коактиватор стероидных рецепторов (SCR) NCOA3 часто сверхэкспрессируется при раке молочной железы , поэтому разработка молекулы-ингибитора, которая нацелена на этот коактиватор и снижает его экспрессию, может быть использована в качестве потенциального лечения рака молочной железы. [ 15 ] [ 20 ]
Поскольку факторы транскрипции контролируют множество различных биологических процессов, они являются идеальными мишенями для лекарственной терапии. [ 14 ] [ 21 ] Регулирующие их коактиваторы можно легко заменить синтетическим лигандом, позволяющим контролировать увеличение или уменьшение экспрессии генов. [ 14 ]
Дальнейшие технологические достижения позволят по-новому взглянуть на функции и регуляцию коактиваторов на уровне всего организма и прояснят их роль в заболеваниях человека, что, как мы надеемся, обеспечит лучшие цели для будущих лекарственных препаратов. [ 14 ] [ 15 ]
Известные коактиваторы
[ редактировать ]На сегодняшний день известно более 300 корегуляторов. [ 15 ] Некоторые примеры этих коактиваторов включают: [ 22 ]
- ARA54 нацелен на андрогенные рецепторы
- ATXN7L3 нацелен на нескольких членов ядерных рецепторов . суперсемейства
- BCL3 нацелен на рецептор 9-цис-ретиноевой кислоты (RXR)
- CBP нацелен на многие факторы транскрипции
- CDC25B нацелен на стероидные рецепторы
- COPS5 нацелен на несколько ядерных рецепторов
- DDC нацелен на андрогенные рецепторы
- EP300 нацелен на многие факторы транскрипции
- KAT5 нацелен на многие ядерные рецепторы
- KDM1A нацелен на андрогенные рецепторы
- Семейство коактиваторов стероидных рецепторов (SRC)
- YAP нацелен на факторы транскрипции
- WWTR1 нацелен на факторы транскрипции
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Кури Эй Джей (2008). Механизмы регуляции транскрипции . Молден, Массачусетс: Блэквелл. ISBN 978-1-4051-0370-1 . OCLC 173367793 .
- ^ Jump up to: а б с «Общий фактор транскрипции/фактор транскрипции» . Изучайте науку в Scitable . Проверено 29 ноября 2017 г.
- ^ Jump up to: а б Пеннаккио Л.А., Бикмор В., Дин А., Нобрега М.А., Бехерано Дж. (апрель 2013 г.). «Усилители: пять основных вопросов» . Обзоры природы Генетика . 14 (4): 288–95. дои : 10.1038/nrg3458 . ПМЦ 4445073 . ПМИД 23503198 .
- ^ Jump up to: а б с д и ж г час Браун С.Э., Лехнер Т., Хоу Л., Уоркман Дж.Л. (январь 2000 г.). «Множество HAT коактиваторов транскрипции». Тенденции биохимических наук . 25 (1): 15–9. дои : 10.1016/S0968-0004(99)01516-9 . ПМИД 10637607 .
- ^ Jump up to: а б с д и Кумар Р., О'Мэлли Б.В. (2008). Корегуляторы NR и болезни человека . Хакенсак, Нью-Джерси: World Scientific. ISBN 978-981-270-536-5 . OCLC 261137374 .
- ^ Воснакис Н., Кох М., Шеер Э., Кесслер П., Мели Ю., Дидье П., Тора Л. (сентябрь 2017 г.). «Коактиваторы и общие факторы транскрипции имеют две отдельные динамические популяции, зависящие от транскрипции» . Журнал ЭМБО . 36 (18): 2710–2725. дои : 10.15252/embj.201696035 . ПМК 5599802 . ПМИД 28724529 .
- ^ Jump up to: а б с д и ж г Шпигельман Б.М., Генрих Р. (октябрь 2004 г.). «Биологический контроль посредством регулируемых коактиваторов транскрипции» . Клетка . 119 (2): 157–67. дои : 10.1016/j.cell.2004.09.037 . ПМИД 15479634 . S2CID 14668705 .
- ^ Скоулз Н.С., Вайнцирль Р.О. (май 2016 г.). «Молекулярная динамика «нечетких» транскрипционных активатор-коактиваторных взаимодействий» . PLOS Вычислительная биология . 12 (5): e1004935. Бибкод : 2016PLSCB..12E4935S . дои : 10.1371/journal.pcbi.1004935 . ПМЦ 4866707 . ПМИД 27175900 .
- ^ Jump up to: а б с Томас MC, Чан CM (май 2006 г.). «Общий механизм транскрипции и общие кофакторы». Критические обзоры по биохимии и молекулярной биологии . 41 (3): 105–78. CiteSeerX 10.1.1.376.5724 . дои : 10.1080/10409230600648736 . ПМИД 16858867 . S2CID 13073440 .
- ^ Дешер, Геро (29 августа 1997 г.). «Нечеткие наносборки: на пути к слоистым полимерным мультикомпозитам». Наука . 277 (5330): 1232–1237. дои : 10.1126/science.277.5330.1232 . ISSN 0036-8075 .
- ^ Jump up to: а б с д и Хермансон О., Гласс К.К., Розенфельд М.Г. (2002). «Корегуляторы ядерных рецепторов: множественные способы модификации». Тенденции в эндокринологии и обмене веществ . 13 (2): 55–60. дои : 10.1016/s1043-2760(01)00527-6 . ПМИД 11854019 . S2CID 38649132 .
- ^ Jump up to: а б Ван Дамм П., Хоул К., Пимента-Маркес А., Хелсенс К., Вандекеркхове Дж., Мартиньо Р.Г., Геверт К., Арнесен Т. (июль 2011 г.). «NatF способствует эволюционному сдвигу в N-концевом ацетилировании белка и важен для нормальной сегрегации хромосом» . ПЛОС Генетика . 7 (7): e1002169. дои : 10.1371/journal.pgen.1002169 . ПМК 3131286 . ПМИД 21750686 .
- ^ Стархейм К.К., Геварт К., Арнесен Т. (апрель 2012 г.). «Белковые N-концевые ацетилтрансферазы: когда начало имеет значение». Тенденции биохимических наук . 37 (4): 152–61. дои : 10.1016/j.tibs.2012.02.003 . ПМИД 22405572 .
- ^ Jump up to: а б с д и Лонард DM, О'Мэлли BW (октябрь 2012 г.). «Корегуляторы ядерных рецепторов: модуляторы патологии и терапевтические мишени» . Обзоры природы. Эндокринология . 8 (10): 598–604. дои : 10.1038/nrendo.2012.100 . ПМК 3564250 . ПМИД 22733267 .
- ^ Jump up to: а б с д Ся Е.Ю., Гудсон М.Л., Цзоу Дж.С., Привальский М.Л., Чен Х.В. (октябрь 2010 г.). «Корегуляторы ядерных рецепторов как новая парадигма терапевтического таргетинга» . Обзоры расширенной доставки лекарств . 62 (13): 1227–37. дои : 10.1016/j.addr.2010.09.016 . ПМЦ 5004779 . ПМИД 20933027 .
- ^ Jump up to: а б «Усилитель» . Изучайте науку в Scitable . Проверено 29 ноября 2017 г.
- ^ Берк А., Зипурски С.Л., Мацудайра П.Т., Балтимор Д., Дарнелл Дж. (2000). Лодиш Х.Ф. (ред.). Молекулярно-клеточная биология (4-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3136-8 . OCLC 41266312 .
- ^ Бекнель Л.Б., Дарлингтон Ю.Ф., Орекснер С., Истон-Маркс Дж., Уоткинс К.А., МакОвити А., Канканамге В.Х., Дехарт М., Сильва К.М. «ЦБП». НУРСА Молекулы . doi : 10.1621/8egsudafco .
- ^ «Ядерные рецепторы» . курсы.washington.edu . Проверено 29 ноября 2017 г.
- ^ Тянь Дж. К., Сюй Дж. (ноябрь 2012 г.). «Коактиватор стероидных рецепторов-3 как потенциальная молекулярная мишень для терапии рака» . Мнение экспертов о терапевтических целях . 16 (11): 1085–96. дои : 10.1517/14728222.2012.718330 . ПМК 3640986 . ПМИД 22924430 .
- ^ Сладек FM (октябрь 2003 г.). «Ядерные рецепторы как мишени для лекарств: новые разработки в области корегуляторов, орфанных рецепторов и основных терапевтических областей». Мнение экспертов о терапевтических целях . 7 (5): 679–84. дои : 10.1517/14728222.7.5.679 . ПМИД 14498828 . S2CID 16891519 .
- ^ «НУРСА – Молекулы» . nursa.org . Проверено 30 ноября 2017 г.
Внешние ссылки
[ редактировать ]- Атлас сигналов ядерных рецепторов (исследовательский консорциум и база данных, финансируемый НИЗ; включает журнал с открытым доступом, индексируемый PubMed, «Сигнализация ядерных рецепторов» )
- TcoF - база данных Dragon о кофакторах транскрипции и белках, взаимодействующих с факторами транскрипции