ЯП1
| ЯП1 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | YAP1 , COB1, YAP, YAP2, YAP65, YKI, Да-связанный белок 1, Yap, Да-связанный регулятор транскрипции, YAP-1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 606608 ; МГИ : 103262 ; Гомологен : 4452 ; Генные карты : YAP1 ; ОМА : YAP1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




YAP1 ( да-ассоциированный белок 1 ), также известный как YAP или YAP65 , представляет собой белок, который действует как корегулятор транскрипции , который способствует транскрипции генов, участвующих в клеточной пролиферации и подавляющих апоптотические гены. YAP1 является компонентом сигнального пути гиппопотама , который регулирует размер органов, регенерацию и онкогенез. идентифицирован благодаря его способности связываться с доменом SH3 протеинтирозинкиназ Yes и Src впервые YAP1 был . [5] YAP1 — мощный онкоген , который усиливается при различных видах рака человека. [6] [7]
Структура
[ редактировать ]
Клонирование гена YAP1 облегчило идентификацию модульного белкового домена , известного как домен WW . [8] [9] [10] Первоначально были идентифицированы две сплайсинговые изоформы продукта гена YAP1, названные YAP1-1 и YAP1-2, которые отличались наличием дополнительных 38 аминокислот , кодирующих домен WW. [11] [12] Помимо домена WW, модульная структура YAP1 содержит богатую пролином область на самом аминоконце , за которой следует TID ( домен, взаимодействующий с транскрипционным фактором TEAD ). [13] Далее, после одного домена WW, который присутствует в изоформе YAP1-1, и двух доменов WW, которые присутствуют в изоформе YAP1-2, следует SH3-BM (мотив связывания Src Homology 3). [5] [14] За SH3-BM следует TAD ( домен трансактивации ) и мотив связывания домена PDZ (PDZ-BM) (рис. 1). [15] [16]
Функция
[ редактировать ]YAP1 является коактиватором транскрипции. [17] а его пролиферативная и онкогенная активность обусловлена его ассоциацией с семейством транскрипционных факторов TEAD , [13] которые активируют гены, которые способствуют росту клеток и ингибируют апоптоз . [18] Были идентифицированы несколько других функциональных партнеров YAP1, включая RUNX, [17] СМАДы , [19] [20] стр73 , [21] ЭрбБ4 , [22] [23] ТП53БП2 , [24] ЛАТС1 / 2 , [25] ПТПН14 , [26] АМОТ , [27] [28] [29] [30] и ЗО1 / 2 . [31] YAP1 и его близкий аналог TAZ ( WWTR1 ) являются основными эффекторами пути супрессора опухоли Hippo. [32] Когда этот путь активирован, YAP1 и TAZ фосфорилируются по остатку серина и изолируются в цитоплазме белками 14-3-3 . [32] Когда путь Hippo не активирован, YAP1/TAZ проникают в ядро и регулируют экспрессию генов . [32]
Сообщается, что несколько генов регулируются YAP1, включая Birc2 , Birc5 , фактор роста соединительной ткани ( CTGF ), амфирегулин (AREG), Cyr61 , Hoxa1 и Hoxc13 .
Также было показано, что YAP/TAZ действуют как датчики жесткости, регулируя механотрансдукцию независимо от сигнального каскада Hippo. [33]
Поскольку YAP и TAZ являются коактиваторами транскрипции, они не имеют ДНК-связывающих доменов. Вместо этого, находясь внутри ядра, они регулируют экспрессию генов посредством TEAD1-4, которые представляют собой специфичные для последовательности факторы транскрипции, которые опосредуют основной транскрипционный результат пути Hippo. [34] Взаимодействие YAP/TAZ и TEAD конкурентно ингибирует и активно диссоциирует взаимодействие TEAD/ VGLL4 , которое действует как репрессор транскрипции. [35] Было показано, что мышиные модели со сверхэкспрессией YAP демонстрируют повышенную регуляцию экспрессии гена-мишени TEAD, что приводит к усилению экспансии клеток-предшественников и избыточному росту тканей. [36]
Регулирование
[ редактировать ]Биохимический
[ редактировать ]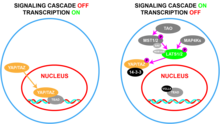
На биохимическом уровне YAP является частью сигнального пути Hippo и регулируется им, где киназный каскад приводит к его «инактивации» вместе с TAZ. [37] В этом сигнальном каскаде киназы ТАО фосфорилируют Ste20-подобные киназы, MST1 / 2 , в их петлях активации (Thr183 для MST1 и Thr180 для MST2). [38] [39] Активный MST1/2 затем фосфорилирует SAV1 и MOB1A / B , которые представляют собой каркасные белки, способствующие рекрутированию и фосфорилированию LATS1/2. [40] [41] LATS1/2 также может фосфорилироваться двумя группами MAP4K . [42] [43] Затем LATS1/2 фосфорилирует YAP и TAZ, что заставляет их связываться с 14-3-3, что приводит к цитоплазматической секвестрации YAP и TAZ. [44] Результатом активации этого пути является ограничение проникновения YAP/TAZ в ядро клетки.
Механотрансдуктивный
[ редактировать ]Кроме того, YAP регулируется механическими сигналами, такими как жесткость внеклеточного матрикса (ECM), напряжение, напряжение сдвига или площадь адгезии - процессы, которые зависят от целостности цитоскелета. [45] Считается, что эти механически вызванные явления локализации являются результатом изменения размера пор, вызванного уплощением ядра, механочувствительных ионных каналов ядерной мембраны, механической стабильности белка или множества других факторов. [45] Эти механические факторы также связаны с некоторыми раковыми клетками посредством размягчения ядра и более высокой жесткости ЕСМ. [46] [47] [48] Согласно этой теории, фенотип размягчения ядра раковых клеток будет способствовать уплощению ядра в ответ на силу, вызывающую локализацию YAP, что может объяснить его сверхэкспрессию и способствовать пролиферации в онкогенных клетках. [49] Кроме того, более высокий фенотип жесткости ЕСМ обычно наблюдается в опухолях из-за усиленной передачи сигналов интегрина. [48] может сгладить клетку и ядро, что снова приведет к более высокой ядерной локализации YAP. Сходным образом было показано, что противоположный эффект жесткости ядра в результате различных стимулов, таких как сверхэкспрессия ламина А , уменьшает ядерную локализацию YAP. [50] [51]
Клиническое значение
[ редактировать ]Рак
[ редактировать ]Нарушение регуляции транскрипционной активности, опосредованной YAP/TAZ, участвует в развитии аномального роста клеток, а гиперактивация YAP и TAZ наблюдалась при многих видах рака. [49] [52] [53] Следовательно, YAP1 представляет собой потенциальную мишень для лечения рака. [54]
Хотя YAP идентифицирован как протоонкоген, он также может действовать как супрессор опухоли в зависимости от клеточного контекста. [55]
Как мишень для наркотиков
[ редактировать ]Онкоген YAP1 служит мишенью для разработки новых лекарств от рака. [56] Были идентифицированы небольшие соединения, которые разрушают комплекс YAP1-TEAD или блокируют функцию связывания WW-доменов. [57] [58] Эти небольшие молекулы представляют собой ведущие соединения для разработки методов лечения онкологических больных, у которых имеется амплифицированный или сверхэкспрессированный онкоген YAP.
Нейропротекция
[ редактировать ]Сигнальный путь Hippo/YAP может оказывать нейропротекторное действие за счет смягчения нарушения гематоэнцефалического барьера после церебральной ишемии/реперфузионного повреждения. [59]
Мутации
[ редактировать ]Гетерозиготные мутации потери функции в гене YAP1 были идентифицированы в двух семьях с серьезными пороками развития глаз с экстраокулярными особенностями или без них, такими как потеря слуха, заячья губа, умственная отсталость и заболевание почек. [60]
Внешние ссылки
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для UniProt : P46937 (Человеческий транскрипционный коактиватор YAP1) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : P46938 (Коактиватор транскрипции мыши YAP1) в PDBe-KB .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000137693 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000053110 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б Судол М (август 1994 г.). «Да-ассоциированный белок (YAP65) представляет собой богатый пролином фосфопротеин, который связывается с доменом SH3 продукта протоонкогена Yes». Онкоген . 9 (8): 2145–52. ПМИД 8035999 .
- ^ Хуан Дж., Ву С., Баррера Дж., Мэтьюз К., Пан Д. (август 2005 г.). «Сигнальный путь Hippo координально регулирует пролиферацию клеток и апоптоз, инактивируя Йорки, гомолога YAP у дрозофилы» . Клетка . 122 (3): 421–34. дои : 10.1016/j.cell.2005.06.007 . ПМИД 16096061 . S2CID 14139806 .
- ^ Оверхольцер М., Чжан Дж., Смолен Г.А., Мьюир Б., Ли В., Сгрой Д.С. и др. (август 2006 г.). «Трансформирующие свойства YAP, кандидата в онкогены на ампликоне хромосомы 11q22» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12405–10. Бибкод : 2006PNAS..10312405O . дои : 10.1073/pnas.0605579103 . ПМК 1533802 . ПМИД 16894141 .
- ^ Борк П., Судол М. (декабрь 1994 г.). «Домен WW: сигнальный сайт в дистрофине?». Тенденции биохимических наук . 19 (12): 531–3. дои : 10.1016/0968-0004(94)90053-1 . ПМИД 7846762 .
- ^ Андре Б., Спрингаэль JY (декабрь 1994 г.). «WWP, новый аминокислотный мотив, присутствующий в одной или нескольких копиях в различных белках, включая дистрофин и SH3-связывающий Yes-ассоциированный белок YAP65». Связь с биохимическими и биофизическими исследованиями . 205 (2): 1201–5. дои : 10.1006/bbrc.1994.2793 . ПМИД 7802651 .
- ^ Хофманн К., Бухер П. (январь 1995 г.). «Домен rsp5 используется белками с различными функциями» . Письма ФЭБС . 358 (2): 153–7. Бибкод : 1995FEBSL.358..153H . дои : 10.1016/0014-5793(94)01415-W . ПМИД 7828727 . S2CID 23110605 .
- ^ Судол М., Борк П., Эйнбонд А., Кастури К., Драк Т., Негрини М. и др. (июнь 1995 г.). «Характеристика гена YAP (Да-ассоциированного белка) млекопитающих и его роль в определении нового белкового модуля, домена WW» . Журнал биологической химии . 270 (24): 14733–41. дои : 10.1074/jbc.270.24.14733 . ПМИД 7782338 .
- ^ Гаффни С.Дж., Ока Т., Мазак В., Хилман Д., Гат У., Мурамацу Т. и др. (ноябрь 2012 г.). «Идентификация, базовая характеристика и эволюционный анализ дифференциально сплайсированных изоформ мРНК человеческого гена YAP1» . Джин . 509 (2): 215–22. дои : 10.1016/j.gene.2012.08.025 . ПМЦ 3455135 . ПМИД 22939869 .
- ^ Перейти обратно: а б Василев А, Канеко К.Дж., Шу Х., Чжао Ю., ДеПамфилис М.Л. (май 2001 г.). «Факторы транскрипции TEAD/TEF используют домен активации YAP65, Src/Yes-ассоциированного белка, локализованного в цитоплазме» . Гены и развитие . 15 (10): 1229–41. дои : 10.1101/gad.888601 . ПМК 313800 . ПМИД 11358867 .
- ^ Рен Р., Майер Б.Дж., Чикетти П., Балтимор Д. (февраль 1993 г.). «Идентификация сайта связывания SH3, богатого пролином и состоящего из десяти аминокислот». Наука . 259 (5098): 1157–61. Бибкод : 1993Sci...259.1157R . дои : 10.1126/science.8438166 . ПМИД 8438166 .
- ^ Ван С., Рааб Р.В., Шац П.Дж., Гуджино В.Б., Ли М. (май 1998 г.). «Консенсус по связыванию пептидов домена NHE-RF-PDZ1 соответствует С-концевой последовательности регулятора трансмембранной проводимости муковисцидоза (CFTR)» . Письма ФЭБС . 427 (1): 103–8. Бибкод : 1998FEBSL.427..103W . дои : 10.1016/S0014-5793(98)00402-5 . ПМИД 9613608 . S2CID 20803242 .
- ^ Молер П.Дж., Креда С.М., Баучер Р.К., Судол М., Статтс М.Дж., Милгрэм С.Л. (ноябрь 1999 г.). «Да-ассоциированный белок 65 локализует p62(c-Да) в апикальном отделе эпителия дыхательных путей путем ассоциации с EBP50» . Журнал клеточной биологии . 147 (4): 879–90. дои : 10.1083/jcb.147.4.879 . ПМК 2156157 . ПМИД 10562288 .
- ^ Перейти обратно: а б Яги Р., Чен Л.Ф., Шигесада К., Мураками Ю., Ито Ю. (май 1999 г.). «WW-домен, содержащий да-ассоциированный белок (YAP), является новым коактиватором транскрипции» . Журнал ЭМБО . 18 (9): 2551–62. дои : 10.1093/emboj/18.9.2551 . ПМЦ 1171336 . ПМИД 10228168 .
- ^ Чжао Б., Ким Дж., Йе X, Лай З.К., Гуань К.Л. (февраль 2009 г.). «Как TEAD-связывающий, так и WW-домены необходимы для стимуляции роста и активности онкогенной трансформации да-ассоциированного белка» . Исследования рака . 69 (3): 1089–98. doi : 10.1158/0008-5472.CAN-08-2997 . ПМИД 19141641 .
- ^ Ферриньо О, Лаллеманд Ф, Верреккья Ф, Л'Хост С, Камонис Дж, Атфи А, Мовьель А (июль 2002 г.). «Да-ассоциированный белок (YAP65) взаимодействует с Smad7 и усиливает его ингибирующую активность в отношении передачи сигналов TGF-бета/Smad» . Онкоген . 21 (32): 4879–84. дои : 10.1038/sj.onc.1205623 . ПМИД 12118366 .
- ^ Арагон Э., Гернер Н., Си Q, Гомес Т., Гао С., Массаге Дж., Масиас М.Дж. (октябрь 2012 г.). «Структурная основа разносторонних взаимодействий Smad7 с регуляторными доменами WW в путях TGF-β» . Структура . 20 (10): 1726–36. дои : 10.1016/j.str.2012.07.014 . ПМЦ 3472128 . ПМИД 22921829 .
- ^ Страно С., Мунаррис Э., Росси М., Кастаньоли Л., Шауль Ю., Сакки А. и др. (май 2001 г.). «Физическое взаимодействие с Yes-ассоциированным белком усиливает транскрипционную активность p73» . Журнал биологической химии . 276 (18): 15164–73. дои : 10.1074/jbc.M010484200 . ПМИД 11278685 .
- ^ Комуро А., Нагай М., Навин Н.Е., Судол М. (август 2003 г.). «Белок YAP, содержащий домен WW, связывается с ErbB-4 и действует как активатор совместной транскрипции для карбоксильно-концевого фрагмента ErbB-4, который транслоцируется в ядро» . Журнал биологической химии . 278 (35): 33334–41. дои : 10.1074/jbc.M305597200 . ПМИД 12807903 .
- ^ Омерович Дж., Пуджиони Э.М., Наполетано С., Виско В., Фрайоли Р., Фрати Л. и др. (апрель 2004 г.). «Лиганд-регулируемая ассоциация ErbB-4 с коактиватором транскрипции YAP65 контролирует транскрипцию на ядерном уровне». Экспериментальные исследования клеток . 294 (2): 469–79. дои : 10.1016/j.yexcr.2003.12.002 . ПМИД 15023535 .
- ^ Эспаньель Х, Судол М (апрель 2001 г.). «Да-ассоциированный белок и p53-связывающий белок-2 взаимодействуют через свои домены WW и SH3» . Журнал биологической химии . 276 (17): 14514–23. дои : 10.1074/jbc.M008568200 . ПМИД 11278422 .
- ^ Ока Т., Мазак В., Судол М. (октябрь 2008 г.). «Киназы Mst2 и Lats регулируют апоптотическую функцию белка, ассоциированного с киназой Yes (YAP)» . Журнал биологической химии . 283 (41): 27534–46. дои : 10.1074/jbc.M804380200 . ПМИД 18640976 .
- ^ Лю X, Ян Н., Фигель С.А., Уилсон К.Э., Моррисон К.Д., Гельман И.Х., Чжан Дж. (март 2013 г.). «PTPN14 взаимодействует с онкогенной функцией YAP и отрицательно регулирует ее» . Онкоген . 32 (10): 1266–73. дои : 10.1038/onc.2012.147 . ПМК 4402938 . ПМИД 22525271 .
- ^ Ван В, Хуан Дж, Чен Дж (февраль 2011 г.). «Ангиомотин-подобные белки связываются с YAP1 и отрицательно регулируют его» . Журнал биологической химии . 286 (6): 4364–70. дои : 10.1074/jbc.C110.205401 . ПМК 3039387 . ПМИД 21187284 .
- ^ Чан С.В., Лим С.Дж., Чонг Ю.Ф., Поббати А.В., Хуан С., Хонг В. (март 2011 г.). «Независимое от пути Hippo ограничение TAZ и YAP ангиомотином» . Журнал биологической химии . 286 (9): 7018–26. дои : 10.1074/jbc.C110.212621 . ПМК 3044958 . ПМИД 21224387 .
- ^ Чжао Б., Ли Л., Лу Ц., Ван Л.Х., Лю С.И., Лэй Ц., Гуань К.Л. (январь 2011 г.). «Ангиомотин — это новый компонент пути Hippo, который ингибирует онкопротеин YAP» . Гены и развитие . 25 (1): 51–63. дои : 10.1101/gad.2000111 . ПМК 3012936 . ПМИД 21205866 .
- ^ Ока Т., Шмитт А.П., Судол М. (январь 2012 г.). «Противоположная роль ангиомотин-подобного-1 и зоны окклюденс-2 в проапоптотической функции YAP» . Онкоген . 31 (1): 128–34. дои : 10.1038/onc.2011.216 . ПМИД 21685940 .
- ^ Ока Т., Ремю Э., Меершерт К., Ванлоо Б., Бушери С., Гфеллер Д. и др. (декабрь 2010 г.). «Функциональные комплексы между YAP2 и ZO-2 зависят от домена PDZ и регулируют ядерную локализацию и передачу сигналов YAP2» . Биохимический журнал (Представлена рукопись). 432 (3): 461–72. дои : 10.1042/BJ20100870 . hdl : 1854/LU-1256657 . ПМИД 20868367 .
- ^ Перейти обратно: а б с Пан Д (октябрь 2010 г.). «Сигнальный путь гиппопотама в развитии и раке» . Развивающая клетка . 19 (4): 491–505. дои : 10.1016/j.devcel.2010.09.011 . ПМК 3124840 . ПМИД 20951342 .
- ^ МакМюррей Р.Дж., Далби М.Дж., Цимбури П.М. (май 2015 г.). «Использование биоматериалов для изучения механотрансдукции, роста и дифференцировки стволовых клеток» (PDF) . Журнал тканевой инженерии и регенеративной медицины . 9 (5): 528–39. дои : 10.1002/сем.1957 . ПМИД 25370612 . S2CID 39642567 .
- ^ Чжао Б., Йе Х, Ю Дж, Ли Л, Ли В, Ли С и др. (июль 2008 г.). «TEAD опосредует YAP-зависимую индукцию генов и контроль роста» . Гены и развитие . 22 (14): 1962–71. дои : 10.1101/gad.1664408 . ПМЦ 2492741 . ПМИД 18579750 .
- ^ Кунц Л.М., Лю-Читтенден Ю., Инь Ф., Чжэн Ю., Юй Дж., Хуан Б. и др. (май 2013 г.). «Эффектор бегемота Йорки контролирует нормальный рост тканей, противодействуя репрессии, опосредованной зубцами по умолчанию» . Развивающая клетка . 25 (4): 388–401. дои : 10.1016/j.devcel.2013.04.021 . ПМК 3705890 . ПМИД 23725764 .
- ^ Чен Ц, Чжан Н, Се Р, Ван В, Цай Дж, Чой К.С. и др. (июнь 2015 г.). «Гомеостатический контроль сигнальной активности Hippo, выявленный с помощью эндогенной активирующей мутации в YAP» . Гены и развитие . 29 (12): 1285–97. дои : 10.1101/gad.264234.115 . ПМЦ 4495399 . ПМИД 26109051 .
- ^ Мэн З., Моройши Т., Гуань К.Л. (январь 2016 г.). «Механизмы регуляции пути бегемота» . Гены и развитие . 30 (1): 1–17. дои : 10.1101/gad.274027.115 . ПМК 4701972 . ПМИД 26728553 .
- ^ Богджано Дж.К., Вандерзалм П.Дж., Фехон Р.Г. (ноябрь 2011 г.). «Tao-1 фосфорилирует киназы Hippo/MST, чтобы регулировать путь супрессора опухолей Hippo-Salvador-Warts» . Развивающая клетка . 21 (5): 888–95. дои : 10.1016/j.devcel.2011.08.028 . ПМК 3217187 . ПМИД 22075147 .
- ^ Пун CL, Линь Цзи, Чжан X, Харви К.Ф. (ноябрь 2011 г.). «Стерильная 20-подобная киназа Тао-1 контролирует рост тканей, регулируя путь Сальвадора-Бородавки-Гиппопотама» . Развивающая клетка . 21 (5): 896–906. дои : 10.1016/j.devcel.2011.09.012 . ПМИД 22075148 .
- ^ Каллус Б.А., Верхаген А.М., Во Д.Л. (сентябрь 2006 г.). «Ассоциация стерильных двадцати киназ млекопитающих, Mst1 и Mst2, с hSalvador через C-концевые спиральные домены приводит к его стабилизации и фосфорилированию» . Журнал ФЭБС . 273 (18): 4264–76. дои : 10.1111/j.1742-4658.2006.05427.x . ПМИД 16930133 . S2CID 8261982 .
- ^ Праскова М., Ся Ф., Авруч Дж. (март 2008 г.). «Фосфорилирование MOBKL1A/MOBKL1B с помощью MST1 и MST2 ингибирует пролиферацию клеток» . Современная биология . 18 (5): 311–21. Бибкод : 2008CBio...18..311P . дои : 10.1016/j.cub.2008.02.006 . ПМЦ 4682548 . ПМИД 18328708 .
- ^ Мэн З., Моройши Т., Моттье-Пави В., Плуфф С.В., Хансен К.Г., Хонг А.В. и др. (октябрь 2015 г.). «Киназы семейства MAP4K действуют параллельно с MST1/2, активируя LATS1/2 в пути Hippo» . Природные коммуникации . 6 : 8357. Бибкод : 2015NatCo...6.8357M . дои : 10.1038/ncomms9357 . ПМЦ 4600732 . ПМИД 26437443 .
- ^ Чжэн Ю, Ван В, Лю Б, Дэн Х, Устер Э, Пан Д (сентябрь 2015 г.). «Идентификация Happyhour/MAP4K как альтернативных Hpo/Mst-подобных киназ в каскаде киназ бегемота» . Развивающая клетка . 34 (6): 642–55. дои : 10.1016/j.devcel.2015.08.014 . ПМЦ 4589524 . ПМИД 26364751 .
- ^ Чжао Б., Вэй Икс, Ли В., Удан Р.С., Ян К., Ким Дж. и др. (ноябрь 2007 г.). «Инактивация онкопротеина YAP по пути Hippo участвует в ингибировании клеточного контакта и контроле роста тканей» . Гены и развитие . 21 (21): 2747–61. дои : 10.1101/gad.1602907 . ПМК 2045129 . ПМИД 17974916 .
- ^ Перейти обратно: а б Элосеги-Артола А., Андреу И., Бидл А.Е., Лезамиз А., Уроз М., Космальска А.Дж. и др. (ноябрь 2017 г.). «Сила вызывает проникновение YAP в ядерную зону, регулируя транспорт через ядерные поры» . Клетка . 171 (6): 1397–1410.e14. дои : 10.1016/j.cell.2017.10.008 . ПМИД 29107331 .
- ^ Кросс С.Е., Джин Ю.С., Рао Дж., Гимжевски Дж.К. (декабрь 2007 г.). «Наномеханический анализ клеток онкологических больных». Природные нанотехнологии . 2 (12): 780–3. Бибкод : 2007NatNa...2..780C . дои : 10.1038/nnano.2007.388 . ПМИД 18654431 .
- ^ Гак Дж., Шинкингер С., Линкольн Б., Воттава Ф., Эберт С., Ромейк М. и др. (май 2005 г.). «Оптическая деформируемость как неотъемлемый клеточный маркер для проверки злокачественной трансформации и метастатической компетентности» . Биофизический журнал . 88 (5): 3689–98. Бибкод : 2005BpJ....88.3689G . дои : 10.1529/biophysj.104.045476 . ПМЦ 1305515 . ПМИД 15722433 .
- ^ Перейти обратно: а б Фридл П., Александр С. (ноябрь 2011 г.). «Раковая инвазия и микроокружение: пластичность и взаимность» . Клетка . 147 (5): 992–1009. дои : 10.1016/j.cell.2011.11.016 . ПМИД 22118458 .
- ^ Перейти обратно: а б Симомура Т., Миямура Н., Хата С., Миура Р., Хираяма Дж., Нишина Х. (январь 2014 г.). «PDZ-связывающий мотив Yes-ассоциированного белка необходим для его совместной активации TEAD-опосредованной транскрипции CTGF и активности по преобразованию онкогенных клеток». Связь с биохимическими и биофизическими исследованиями . 443 (3): 917–23. дои : 10.1016/j.bbrc.2013.12.100 . ПМИД 24380865 .
- ^ Свифт Дж., Ивановска И.Л., Буксбойм А., Харада Т., Дингал П.С., Пинтер Дж. и др. (август 2013 г.). «Ядерный ламин-А масштабируется с жесткостью тканей и усиливает дифференцировку, направленную на матрикс» . Наука . 341 (6149): 1240104. doi : 10.1126/science.1240104 . ПМЦ 3976548 . ПМИД 23990565 .
- ^ Гьоревски Н., Сакс Н., Манфрин А., Гигер С., Брагина М.Е., Ордоньес-Моран П. и др. (ноябрь 2016 г.). «Конструктор матриц для кишечных стволовых клеток и культур органоидов». Природа . 539 (7630): 560–564. дои : 10.1038/nature20168 . ПМИД 27851739 . S2CID 4470849 .
- ^ Харви К.Ф., Чжан X, Томас Д.М. (апрель 2013 г.). «Путь бегемота и рак человека». Обзоры природы. Рак . 13 (4): 246–57. дои : 10.1038/nrc3458 . ПМИД 23467301 . S2CID 2008641 .
- ^ Джонсон Р., Гальдер Дж. (январь 2014 г.). «Два лица Hippo: выбор пути Hippo для регенеративной медицины и лечения рака» . Обзоры природы. Открытие наркотиков . 13 (1): 63–79. дои : 10.1038/nrd4161 . ПМК 4167640 . ПМИД 24336504 .
- ^ Моройши Т., Хансен К.Г., Гуань К.Л. (февраль 2015 г.). «Новая роль YAP и TAZ в лечении рака» . Обзоры природы. Рак . 15 (2): 73–79. дои : 10.1038/nrc3876 . ПМЦ 4562315 . ПМИД 25592648 .
- ^ Джо Э (ноябрь 2018 г.). «Двойная роль YAP: онкопротеин и супрессор опухоли» . Журнал торакальных заболеваний . 10 (Приложение 33): S3895–S3898. дои : 10.21037/jtd.2018.10.70 . ПМК 6297531 . ПМИД 30631509 .
- ^ Судол М., Шилдс, округ Колумбия, Фарук А. (сентябрь 2012 г.). «Структуры белковых доменов YAP открывают многообещающие цели для разработки новых лекарств от рака» . Семинары по клеточной биологии и биологии развития . 23 (7): 827–33. дои : 10.1016/j.semcdb.2012.05.002 . ПМЦ 3427467 . ПМИД 22609812 .
- ^ Лю-Читтенден И., Хуан Б., Шим Дж.С., Чен К., Ли С.Дж., Андерс Р.А. и др. (июнь 2012 г.). «Генетическое и фармакологическое нарушение комплекса TEAD-YAP подавляет онкогенную активность YAP» . Гены и развитие . 26 (12): 1300–5. дои : 10.1101/gad.192856.112 . ПМЦ 3387657 . ПМИД 22677547 .
- ^ Кан С.Г., Хуинь Т., Чжоу Р. (2012). «Неразрушающее ингибирование металлофуллеренола Gd@C(82)(OH)(22) в домене WW: влияние на путь передачи сигнала» . Научные отчеты . 2 : 957. Бибкод : 2012НатСР...2Е.957К . дои : 10.1038/srep00957 . ПМК 3518810 . ПМИД 23233876 .
- ^ Гонг П., Чжан З., Цзоу С., Тянь Ц., Чэнь Х., Хун М. и др. (январь 2019 г.). «Сигнальный путь Hippo/YAP смягчает нарушение гематоэнцефалического барьера после церебральной ишемии/реперфузионного повреждения» . Поведенческие исследования мозга . 356 : 8–17. дои : 10.1016/j.bbr.2018.08.003 . ПМК 6193462 . ПМИД 30092249 .
- ^ Уильямсон К.А., Рейнгер Дж., Флойд Дж.А., Ансари М., Мейнерт А., Олдридж К.В. и др. (февраль 2014 г.). «Гетерозиготные мутации потери функции в YAP1 вызывают как изолированные, так и синдромальные дефекты закрытия зрительной щели» . Американский журнал генетики человека . 94 (2): 295–302. дои : 10.1016/j.ajhg.2014.01.001 . ПМЦ 3928658 . ПМИД 24462371 .
галерея PDB |
|---|