WWTR1
| WWTR1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | WWTR1 , TAZ, домен WW, содержащий регулятор транскрипции 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 607392 ; МГИ : 1917649 ; Гомологен : 9159 ; GeneCards : WWTR1 ; ОМА : WWTR1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






Белок-регулятор транскрипции, содержащий WW-домен (WWTR1 [ 5 ] ), также известный как коактиватор транскрипции с PDZ-связывающим мотивом (TAZ), представляет собой белок , который у человека кодируется WWTR1 геном . WWTR1 действует как корегулятор транскрипции и не влияет только на транскрипцию . [ 5 ] В комплексе с партнерами по связыванию транскрипционных факторов WWTR1 помогает стимулировать экспрессию генов в путях, связанных с развитием , клеток ростом и выживанием , а также ингибированием апоптоза . [ 6 ] Аберрантная функция WWTR1 связана с ее ролью в возникновении рака . [ 7 ] [ 8 ] [ 9 ] WWTR1 часто называют TAZ из-за его первоначального названия TAZ. Однако WWTR1 (TAZ) не следует путать с белком тафаззином , который первоначально имел официальный символ гена TAZ, а теперь называется TAFAZZIN.
Структура
[ редактировать ]

WWTR1 содержит богатую пролином область, мотив связывания TEAD , домен WW , область спиральной спирали и домен трансактивации (TAD) , содержащий домена PDZ мотив связывания . WWTR1 (TAZ) не имеет ДНК-связывающего домена , поэтому он не может напрямую управлять транскрипцией. WWTR1 демонстрирует консервативную структурную гомологию с другим корегулятором транскрипции, да-ассоциированным белком 1 (YAP) . [ 5 ] И YAP, и TAZ способны образовывать гомодимеры и гетеродимеры друг с другом посредством взаимодействий в домене катушки-клубка. [ 12 ] YAP и TAZ взаимодействуют с факторами транскрипции, способствуя образованию тканей . WWTR1 (TAZ) взаимодействует с множеством партнеров по транскрипции, включая четыре члена семейства доменов TEA ( TEAD1 /2/ 3/ 4 ) через мотив связывания TEAD и несколько других факторов, содержащих мотив PPXY, который состоит из пролина - пролина. -X (любая аминокислота )- тирозина последовательность . Примеры таких партнеров включают Runx/PEBP2, AP2 , C/EBP, c-Jun , Krox-20 , Krox-24 , MEF2B , NF-E2 , Oct-4 и p73 , которые взаимодействуют с WWTR1 через домен WW. [ 6 ] Было показано, что домен трансактивации на С- конце (аминокислоты 165–395) играет важную роль в обеспечении транскрипционных эффектов. [ 6 ]
Функция
[ редактировать ]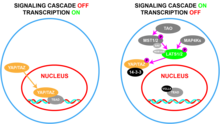
WWTR1 (TAZ) играет важную роль в эмбриогенезе и развитии . [ 13 ] [ 14 ] которые включают регуляцию размера органов, [ 15 ] [ 16 ] [ 17 ] обновление стволовых клеток , [ 18 ] регенерация тканей , [ 19 ] [ 18 ] остеогенез , [ 20 ] и ангиогенез . [ 21 ] Эти функции реализуются посредством совместной активации факторов транскрипции, которые способствуют росту, миграции и дифференцировке клеток. [ 15 ] [ 16 ] [ 17 ] такие как четыре члена семейства транскрипционных факторов TEAD, ген парного бокса 3 (PAX3) и факторы транскрипции, связанные с Runt (RUNX1/) 2 ). [ 11 ] Пролиферативные функции WWTR1 (TAZ) и его паралога YAP ограничиваются сигнальным путем Hippo . [ 22 ] [ 23 ] [ 24 ] Этот супрессивный путь состоит из киназного сигнального каскада, ядро которого состоит из серин-треониновых киназ STK3 , / MST2 и STK4 / MST1 , которые, будучи активными и образуя комплекс с фосфорилируют и регуляторным белком SAV1 активируют LATS1. /2- киназы, которые в комплексе с регуляторным белком MOB1 фосфорилируют и далее инактивируют YAP/TAZ. [ 14 ] [ 15 ] [ 25 ] Таким образом, активация Hippo останавливает рост клеток за счет снижения экспрессии пролиферативных генов, что приводит к снижению гибели клеток в результате ферроптоза. [ 26 ] [ 27 ] и повышенная гибель клеток в результате апоптоза . [ 14 ] [ 15 ]
Функциональное резервирование с помощью YAP
[ редактировать ]Сходства
[ редактировать ]WWTR1 (TAZ) имеет сходную структурную последовательность и мотивы связывания с да-ассоциированным белком 1 (YAP) . [ 11 ] В существующей литературе YAP и TAZ часто считаются функционально избыточными. [ 11 ] Оба играют роль в развитии размеров органов, а также в миграции клеток , заживлении ран , ангиогенезе и метаболизме , особенно в липогенезе . [ 11 ] [ 28 ] Инактивация YAP и TAZ происходит посредством фосфорилирования киназами пути Hippo, а именно LATS1 и LATS2. [ 11 ] Это рекрутирует связывание регуляторного белка 14-3-3 , который предотвращает локализацию YAP/TAZ в ядре и маркирует его для убиквитинирования , что позволяет распознавать его для последующей деградации протеасомами . [ 11 ]
Различия
[ редактировать ]TAZ способен образовывать как гетеродимеры , так и гетеротетрамеры с TEAD для инициации транскрипции (TAZ-TEAD и TAZ-TEAD-TAZ-TEAD), тогда как YAP способен образовывать только гетеродимеры YAP-TEAD . [ 11 ] Эти различия придают TAZ уникальные функции, такие как регуляция дифференцировки адипоцитов посредством взаимодействия с рецептором, активирующим пролифератор пероксисом (PPARγ) , а также остеогенез посредством транскрипционной коактивации костно-специфичных транскрипционных факторов, таких как RUNX2 (также известный как Cbfa1 .) [ 11 ] Кроме того, TAZ независимо взаимодействует с ядерным фактором активированных Т-клеток 5 (NFATC5), подавляя транскрипцию в почечных клетках, подвергающихся осмотическому стрессу . [ 11 ] И YAP, и TAZ связываются с против декапентаплегических семейных транскрипционных факторов (SMAD), материнскими комплексами чтобы способствовать передаче сигналов TGF-бета и стимулировать дифференцировку и развитие, но активация только TAZ происходит при трансдукции этого каскада. [ 11 ] TAZ способен образовывать комплекс только с SMAD2 , SMAD3 или SMAD4 , чтобы способствовать ядерному челноку и транскрипции, но YAP также может взаимодействовать с SMAD1 и SMAD7, кроме того. [ 11 ] Исследования на мышах in vivo показали, что животные, лишенные функционального TAZ, более жизнеспособны, чем животные, лишенные экспрессии YAP. [ 11 ] Напротив, подавление YAP способствовало более сильному эффекту на расширение клеток, поглощение глюкозы и остановку клеточного цикла , чем TAZ. [ 11 ] При анализе на немелкоклеточного рака легких (НМРЛ) клеточных линиях WWTR1 поддерживал организацию и адгезию внеклеточного матрикса (ECM) , а также контролировал миграцию в большей степени, чем YAP, который более точно регулирует гены деления клеток и прогрессирования клеточного цикла. [ 11 ]
Белковые взаимодействия
[ редактировать ]

| Партнер по взаимодействию с белками | Функциональные эффекты |
|---|---|
| ВКЛАД , Ангиомотин | Связывание секвестров YAP/TAZ в цитоплазме, ингибирование их функции [ 12 ] |
| AP-1, белок-активатор 1 | Содействие транскрипции [ 11 ] |
| ASPP2 , белок р53, стимулирующий апоптоз. | Способствует дефосфорилированию и стабилизации WWTR1 (TAZ). [ 12 ] |
| β-катенин | Набирает комплекс уничтожения, деактивирующий ЯП/ТАЗ. [ 12 ] |
| LATS1 / LATS2 , Большие киназы-супрессоры опухолей | Фосфорилирование WWTR1, маркирующее его для убиквинизации [ 11 ] |
| NFATC5 , Ядерный фактор активированных Т-клеток 5 | Подавляет транскрипцию в почечных клетках, подвергающихся осмотическому стрессу. [ 11 ] |
| ПФ, Парафибромин | Стимулирует функцию WWTR1 (TAZ) [ 12 ] |
| PAX3 , ген парного бокса 3 | Продвижение транскрипции [ 11 ] |
| PAX8 ( ген парного бокса 8 ) и NKX2-4 (гомеобокс NK) [ 25 ] | Коактивация транскрипционных факторов, участвующих в регуляции щитовидной железы [ 25 ] |
| PRRG4, богатый пролином и домен Gla 4 [ 25 ] | Подавление транскрипции [ 11 ] |
| RUNX1 /2 , факторы транскрипции, связанные с Runt | Способствует транскрипции [ 11 ] |
| SMAD2 /3 /4 , Матери против транскрипционных факторов семейства декапентаплегии | Ядерный челнок; продвижение транскрипции [ 11 ] |
| STAT1 , преобразователь сигнала и активатор транскрипции | Ингибирование димеризации STAT1/2 в метаболизме [ 11 ] |
| TEAD1 , член семейства доменов TEA [ 25 ] | Транскрипционная активация [ 11 ] |
| TEAD2 , член семейства доменов TEA, [ 25 ] | |
| TEAD3 , член семейства доменов TEA, [ 25 ] | |
| TEAD4 , член семейства доменов TEA [ 25 ] | |
| YAP1 , Да-ассоциированный белок 1 | Регуляция транскрипции, зависимая от димеризации [ 11 ] |
| YWHAE (14-3-3) , белок активации тирозин-3-монооксигеназы/триптофана-5-монооксигеназы Эпсилон [ 25 ] | Ограничивает транслокацию WWTR1 в ядро [ 5 ] |
| ZO-2 , белок плотных соединений 2 | Локализует YAP/TAZ в ядре для повышения активности [ 12 ] |
Клиническое значение
[ редактировать ]Роль в болезни
[ редактировать ]WWTR1 участвует во многих воспалительных заболеваниях, включая рак.
| Болезнь | Клиническое значение WWTR1 |
|---|---|
| Рак | Связан с метастазированием и плохим прогнозом выживаемости при многих типах рака. [ 9 ] |
| Стеатогепатит | Сверхэкспрессия WWTR1 приводит к прогрессированию простого стеатоза до стеатогепатита , способствуя фиброзу. [ 29 ] |
| Атеросклероз | Стимулирует чрезмерную пролиферацию эндотелиальных клеток и воспаление. [ 30 ] |
| Синдром Шегрена | Снижение локализации WWTR1 в ядре приводит к отсутствию функционального развития слюнных/слёзных желез. [ 14 ] |
| Гипертония | Активация YAP/TAZ способствует метаболизму глютамина и повышает легочное артериальное давление. [ 31 ] |
| Псориаз | Активация YAP/TAZ приводит к патологическому ангиогенезу и воспалению, связанным с хроническими заболеваниями кожи. [ 32 ] |
| Атопический дерматит | |
| Розацеа | |
| Хроническая крапивница |
Рак
[ редактировать ]WWTR1 (TAZ) вовлечен в широкий спектр раковых заболеваний, включая меланому, плоскоклеточный рак головы и шеи, рак молочной железы, немелкоклеточный рак легких и другие, из-за его высокой генной и гистологической экспрессии, а также корреляции с повышенным метастазированием и более низкая выживаемость в исследованиях на животных и данных пациентов. [ 9 ] Наряду со структурно сходным корегулятором YAP, во многих исследованиях была описана его роль в стимулировании онкогенеза, изменении неопластического метаболизма и формировании устойчивости к терапевтическому вмешательству. [ 8 ] [ 9 ] [ 33 ] [ 34 ] В частности, сверхэкспрессия TAZ обеспечивала устойчивость к цисплатином химиотерапии , а также к иммунотерапевтическому лечению антителом PD-1 . [ 33 ]

| Экспрессия белка WWTR1 по типам рака (из Атласа белков) | |||
|---|---|---|---|
| Тип рака | # Образцы со средней/высокой экспрессией | Всего # образцов пациентов | % образцов пациентов со средней/высокой экспрессией |
| Глиома | 11 | 11 | 100% |
| Щитовидная железа | 4 | 4 | 100% |
| Легкое | 12 | 12 | 100% |
| Колоректальный | 11 | 11 | 100% |
| Голова и шея | 4 | 4 | 100% |
| Печень | 12 | 12 | 100% |
| Карциноид | 4 | 4 | 100% |
| поджелудочная | 9 | 9 | 100% |
| Уротелиальный | 9 | 9 | 100% |
| Простата | 10 | 10 | 100% |
| Яичко | 11 | 11 | 100% |
| Грудь | 12 | 12 | 100% |
| шейный | 9 | 9 | 100% |
| эндометриальный | 10 | 10 | 100% |
| яичник | 11 | 11 | 100% |
| Меланома | 12 | 12 | 100% |
| Кожа | 11 | 11 | 100% |
| Желудок | 10 | 11 | 90.1% |
| Реналь | 10 | 11 | 90.1% |
| Лимфома | 9 | 12 | 75% |

Как мишень для наркотиков
[ редактировать ]Функции YAP и TAZ были нацелены в нескольких терапевтических методах лечения рака.
передачи сигналов Hippo Агонист , C19, увеличивает фосфорилирование MST1/2 и LATS1/2, что приводит к большей инактивации YAP/TAZ. Модулирование внеклеточного матрикса жесткости и напряжения с помощью тиазовивина , кукурбитацина I , дазатиниба , флувастатина и пазопаниба показало положительные результаты в клеточных линиях рака молочной железы, предотвращая транслокацию YAP/WWTR1 в ядро. [ 35 ] Было также продемонстрировано, что эндогенные гормональные факторы, которые синтезируются для нормальных физиологических функций, такие как адреналин и глюкагон, оказывают аналогичное ингибирующее действие на функцию YAP/TAZ, способствуя активации пути Hippo. [ 35 ] Было показано, что класс ингибиторов холестерина , статины , ингибирует семейство GTP-аз Rho (Rho-GTPase) , которые представляют собой ферменты , которые сигнализируют об ингибировании восходящего пути Hippo, и проявляют аналогичные эффекты в замедлении роста рака молочной железы и человека. клетки аденокарциномы легких . [ 35 ] Статины ингибируют 3-гидрокси-3-метилглутарил-коэнзим А-редуктазу (HMG-CoA-редуктаза) , которая является предшественником мевалоната в мевалонатном пути , который синтезирует липидные строительные блоки, образующие холестерин, и липидные цепи, ответственные за закрепление Rho-коэнзима. ГТФазы к клеточной мембране. [ 9 ] Rho-GTPase, гомолог A семейства Ras (RhoA) , активируется путем пренлилирования ( посттрансляционная модификация посредством добавления гидрофобных групп ) и частично отвечает за модуляцию элементов цитоскелета , которые снижают активность пути Hippo. [ 9 ] Путем воздействия на киназы Rho с помощью тиазовивина или синтеза липидов через мевалонатный путь с помощью статинов ингибируется RhoA, а повышенная активность киназы Hippo может ограничивать пролиферацию, управляемую YAP/TAZ. [ 9 ] [ 35 ] Тирозинкиназы сигнализируют о пролиферативных путях, некоторые из которых способствуют функции YAP/TAZ, такие как киназы семейства Src , и включают тирозинкиназу Yes, которая связана с функцией YAP. Нацеливание на тирозинкиназы с помощью ингибиторов, таких как дазатиниб и пазопаниб, показало некоторый эффект при раке. [ 9 ]
Также изучалось ингибирование функции YAP/TAZ путем нацеливания на их взаимодействие с партнерами по транскрипции в семействе TEAD. [ 35 ] [ 36 ] Сюда входит использование вертепорфина , который исследовался при лечении рака кожи, особенно меланомы, хотя и не вышел за рамки доклинических исследований. [ 35 ]
| Название препарата/молекулы | Класс препарата | Механизм действия |
|---|---|---|
| С19 | Агонист киназы гиппопотама | Увеличивает фосфорилирование с помощью MST1/2 и LATS1/2, инактивируя YAP/TAZ и уменьшая пролиферацию клеток. [ 35 ] |
| Дигидрексидин | Агонист дофамина | Увеличивает активность LATS1/2; Снижает функцию YAP/TAZ и пролиферацию клеток. [ 35 ] |
| Адреналин | Гормональный фактор | Увеличивает активность LATS1/2; Снижает функцию YAP/TAZ и пролиферацию клеток. [ 35 ] |
| глюкагон | Гормональный фактор | |
| Тиазовивин | Ингибитор Rho-киназы | Ингибирует Rho-GTPase; Увеличивает активность LATS1/2; Снижает функцию YAP/TAZ и пролиферацию клеток. [ 9 ] [ 35 ] |
| Кукурбитацин I | Ингибитор JAK/STAT3 | |
| Дазатиниб | Ингибитор тирозинкиназы | |
| Флувастатин | Статины | |
| Пазопаниб | Ингибитор тирозинкиназы | |
| Verteporfin | Ингибитор TEAD | Ингибирует связывание YAP/TAZ с факторами транскрипции семейства TEAD; Снижает пролиферативную транскрипцию и пролиферацию клеток. [ 35 ] |
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000018408 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000027803 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с д Канаи Ф., Мариньяни П.А., Сарбасова Д., Яги Р., Холл Р.А., Доновиц М. и др. (декабрь 2000 г.). «TAZ: новый коактиватор транскрипции, регулируемый взаимодействием с белками домена 14-3-3 и PDZ» . Журнал ЭМБО . 19 (24): 6778–6791. дои : 10.1093/emboj/19.24.6778 . ПМК 305881 . ПМИД 11118213 .
- ^ Перейти обратно: а б с Хун В., Гуань К.Л. (сентябрь 2012 г.). «Коактиваторы транскрипции YAP и TAZ: ключевые нижестоящие эффекторы пути Hippo млекопитающих» . Семинары по клеточной биологии и биологии развития . 23 (7): 785–793. дои : 10.1016/j.semcdb.2012.05.004 . ПМК 3459069 . ПМИД 22659496 .
- ^ Перейти обратно: а б с Моройши Т., Хансен К.Г., Гуань К.Л. (февраль 2015 г.). «Новая роль YAP и TAZ в лечении рака» . Обзоры природы. Рак . 15 (2): 73–79. дои : 10.1038/nrc3876 . ПМЦ 4562315 . ПМИД 25592648 .
- ^ Перейти обратно: а б Чжан X, Чжао Х, Ли Ю, Ся Д, Ян Л, Ма Ю и др. (сентябрь 2018 г.). «Роль активности YAP/TAZ в метаболическом перепрограммировании рака» . Молекулярный рак . 17 (1): 134. дои : 10.1186/s12943-018-0882-1 . ПМК 6122186 . ПМИД 30176928 .
- ^ Перейти обратно: а б с д и ж г час я Занконато Ф, Корденонси М, Пикколо С (июнь 2016 г.). «ЯП/ТАЗ у корней рака» . Раковая клетка . 29 (6): 783–803. doi : 10.1016/j.ccell.2016.05.005 . ПМК 6186419 . ПМИД 27300434 .
- ^ «База данных структуры белков AlphaFold» . Alphafold.ebi.ac.uk . Проверено 23 ноября 2022 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Реджани Ф., Гобби Дж., Чиаррокки А., Санчизи В. (февраль 2021 г.). «ЯП и ТАЗ — не однояйцевые близнецы» Тенденции биохимических наук . 46 (2): 154–168. дои : 10.1016/j.tibs.2020.08.012 . ПМИД 32981815 . S2CID 222166778 .
- ^ Перейти обратно: а б с д и ж Каллус Б.А., Финч-Эдмондсон М.Л., Флетчер С., Уилтон С.Д. (февраль 2019 г.). "Тявкать о ТАЗе и не забывать его" . Письма ФЭБС . 593 (3): 253–276. дои : 10.1002/1873-3468.13318 . ПМИД 30570758 . S2CID 58578804 .
- ^ У Цзы, Гуань К.Л. (январь 2021 г.). «Передача сигналов гиппопотама в эмбриогенезе и развитии» . Тенденции биохимических наук . 46 (1): 51–63. дои : 10.1016/j.tibs.2020.08.008 . ПМЦ 7749079 . ПМИД 32928629 .
- ^ Перейти обратно: а б с д Чжэн Ю, Пан Д. (август 2019 г.). «Сигнальный путь бегемотов в развитии и заболеваниях» . Развивающая клетка . 50 (3): 264–282. дои : 10.1016/j.devcel.2019.06.003 . ПМК 6748048 . ПМИД 31386861 .
- ^ Перейти обратно: а б с д Пикколо С., Дюпон С., Корденонси М. (октябрь 2014 г.). «Биология YAP/TAZ: передача сигналов гиппопотамов и не только». Физиологические обзоры . 94 (4): 1287–1312. doi : 10.1152/physrev.00005.2014 . ПМИД 25287865 .
- ^ Перейти обратно: а б Покатерра А., Романи П., Дюпон С. (январь 2020 г.). «Краткий обзор функций ЯП/ТАЗ и их регулирования». Журнал клеточной науки . 133 (2): jcs230425. дои : 10.1242/jcs.230425 . hdl : 11577/3317485 . ПМИД 31996398 . S2CID 210945848 .
- ^ Перейти обратно: а б Тотаро А., Пансьера Т., Пикколо С. (август 2018 г.). «Сигналы восходящего потока YAP/TAZ и ответы нисходящего потока» . Природная клеточная биология . 20 (8): 888–899. дои : 10.1038/s41556-018-0142-z . ПМК 6186418 . ПМИД 30050119 .
- ^ Перейти обратно: а б Моя И.М., Гальдер Г. (апрель 2019 г.). «Передача сигналов Hippo-YAP/TAZ в регенерации органов и регенеративной медицине» . Обзоры природы. Молекулярно-клеточная биология . 20 (4): 211–226. дои : 10.1038/s41580-018-0086-y . ПМИД 30546055 . S2CID 54820180 .
- ^ Дрискилл Дж. Х., Пан Д. (январь 2021 г.). «Путь бегемота в гомеостазе и патофизиологии печени» . Ежегодный обзор патологии . 16 : 299–322. doi : 10.1146/annurev-pathol-030420-105050 . ПМЦ 8594752 . ПМИД 33234023 .
- ^ Ковар Х., Бирбаумер Л., Радич-Сарикас Б. (апрель 2020 г.). «Путь YAP/TAZ в остеогенезе и патогенезе костной саркомы» . Клетки . 9 (4): 972. doi : 10.3390/cells9040972 . ПМК 7227004 . ПМИД 32326412 .
- ^ Бупатия GT, Хонг В (2019). «Роль передачи сигналов Hippo Pathway-YAP/TAZ в ангиогенезе» . Границы клеточной биологии и биологии развития . 7 : 49. дои : 10.3389/fcell.2019.00049 . ПМК 6468149 . ПМИД 31024911 .
- ^ Хэн BC, Чжан X, Обель Д., Бай Y, Ли X, Вэй Y и др. (январь 2021 г.). «Обзор сигнальных путей, регулирующих активность YAP/TAZ» . Клеточные и молекулярные науки о жизни . 78 (2): 497–512. дои : 10.1007/ s00018-020-03579-8 ПМК 11071991 . ПМИД 32748155 . S2CID 220930261 .
- ^ Ма С., Мэн З., Чен Р., Гуань К.Л. (июнь 2019 г.). «Путь бегемота: биология и патофизиология». Ежегодный обзор биохимии . 88 : 577–604. doi : 10.1146/annurev-biochem-013118-111829 . ПМИД 30566373 . S2CID 58642425 .
- ^ Мэн З., Моройши Т., Гуань К.Л. (январь 2016 г.). «Механизмы регуляции пути бегемота» . Гены и развитие . 30 (1): 1–17. дои : 10.1101/gad.274027.115 . ПМК 4701972 . ПМИД 26728553 .
- ^ Перейти обратно: а б с д и ж г час я дж к "UniProt - Q9GZV5 · WWTR1_HUMAN" . www.uniprot.org . Проверено 17 ноября 2022 г.
- ^ Сунь Т, Чи Дж.Т. (май 2021 г.). «Регуляция ферроптоза в раковых клетках с помощью путей YAP/TAZ и Hippo: терапевтические последствия» . Гены и болезни . 8 (3): 241–249. дои : 10.1016/j.gendis.2020.05.004 . ПМЦ 8093643 . ПМИД 33997171 .
- ^ Дай С., Чен Икс, Ли Дж., Комиш П., Кан Р., Тан Д. (сентябрь 2020 г.). «Факторы транскрипции при ферроптотической гибели клеток» . Генная терапия рака . 27 (9): 645–656. дои : 10.1038/s41417-020-0170-2 . ПМИД 32123318 . S2CID 211728890 .
- ^ Ку Дж. Х., Гуань К. Л. (август 2018 г.). «Взаимодействие между YAP/TAZ и обменом веществ» . Клеточный метаболизм . 28 (2): 196–206. дои : 10.1016/j.cmet.2018.07.010 . ПМИД 30089241 . S2CID 51939438 .
- ^ Кучай М.С., Чоудхари Н.С., Мишра С.К. (01.11.2020). «Патофизиологические механизмы, лежащие в основе MAFLD». Диабет и метаболический синдром . 14 (6): 1875–1887. дои : 10.1016/j.dsx.2020.09.026 . ПМИД 32998095 . S2CID 222166643 .
- ^ Ню Н, Сюй С, Сюй Ю, Литтл ПиДжей, Джин ЗГ (апрель 2019 г.). «Нацеливание на механочувствительные факторы транскрипции при атеросклерозе» . Тенденции в фармакологических науках . 40 (4): 253–266. дои : 10.1016/j.tips.2019.02.004 . ПМК 6433497 . ПМИД 30826122 .
- ^ Чжан X, Чжао Х, Ли Ю, Ся Д, Ян Л, Ма Ю и др. (сентябрь 2018 г.). «Роль активности YAP/TAZ в метаболическом перепрограммировании рака» . Молекулярный рак . 17 (1): 134. дои : 10.1186/s12943-018-0882-1 . ПМК 6122186 . ПМИД 30176928 .
- ^ Ли Х.Дж., Хонг Ю.Дж., Ким М. (ноябрь 2021 г.). «Ангиогенез при хронических воспалительных заболеваниях кожи» . Международный журнал молекулярных наук . 22 (21): 12035. doi : 10.3390/ijms222112035 . ПМЦ 8584589 . ПМИД 34769465 .
- ^ Перейти обратно: а б Томпсон Би Джей (май 2020 г.). «YAP/TAZ: факторы роста опухоли, метастазирования и устойчивости к терапии». Биоэссе . 42 (5): e1900162. doi : 10.1002/bies.201900162 . hdl : 1885/211659 . ПМИД 32128850 . S2CID 212405819 .
- ^ Занконато Ф, Корденонси М, Пикколо С (август 2019 г.). «ЯП и ТАЗ: сигнальный узел микроокружения опухоли». Обзоры природы. Рак . 19 (8): 454–464. дои : 10.1038/s41568-019-0168-y . ПМИД 31270418 . S2CID 195791034 .
- ^ Перейти обратно: а б с д и ж г час я дж к Андл Т., Чжоу Л., Ян К., Кадекаро А.Л., Чжан Ю. (июнь 2017 г.). «YAP и WWTR1: новые цели в лечении рака кожи». Письма о раке . 396 : 30–41. дои : 10.1016/j.canlet.2017.03.001 . ПМИД 28279717 .
- ^ Поббати А.В., Хонг В. (2020). «Борьба с онкопротеинами YAP/TAZ-TEAD в терапии рака» . Тераностика . 10 (8): 3622–3635. дои : 10.7150/thno.40889 . ПМК 7069086 . ПМИД 32206112 .