Ангиогенез
| Ангиогенез | |
|---|---|
 Ангиогенез после васкулогенеза | |
| Идентификаторы | |
| МеШ | Д000096482 |
| Анатомическая терминология | |

Ангиогенез — это физиологический процесс, в ходе которого новые кровеносные сосуды . из ранее существовавших сосудов образуются [1] [2] [3] образуются на более ранней стадии васкулогенеза . Ангиогенез продолжает рост сосудистой сети в основном за счет процессов прорастания и расщепления, но такие процессы, как слияние ангиогенеза , [4] Удлинение сосудов и коопция сосудов также играют роль. [2] Васкулогенез – это эмбриональное образование эндотелиальных клеток из мезодермы . предшественников клеток [5] и от неоваскуляризации , хотя обсуждения не всегда точны (особенно в старых текстах). Первые сосуды в развивающемся эмбрионе формируются посредством васкулогенеза, после чего ангиогенез ответственен за большую часть, если не за весь рост кровеносных сосудов во время развития и при заболевании. [6]
Ангиогенез — это нормальный и жизненно важный процесс роста и развития, а также заживления ран и образования грануляционной ткани . Однако это также фундаментальный шаг в переходе опухолей из доброкачественного состояния в злокачественное , приводящий к использованию ингибиторов ангиогенеза при лечении рака . [7] Существенная роль ангиогенеза в росте опухолей была впервые предположена в 1971 году Джудой Фолкманом , который описал опухоли как «горячие и кровавые». [8] иллюстрируя, что, по крайней мере, для многих типов опухолей приливная перфузия и даже гиперемия характерны .
Типы [ править ]
Прорастающий ангиогенез [ править ]
Прорастающий ангиогенез был первой идентифицированной формой ангиогенеза, и поэтому он гораздо более понятен, чем инвагинальный ангиогенез. Это происходит в несколько четко охарактеризованных стадий. Первоначальный сигнал поступает из участков ткани, лишенных сосудистой сети. Гипоксия , отмечающаяся в этих областях, заставляет ткани требовать присутствия питательных веществ и кислорода, которые позволят тканям осуществлять метаболическую деятельность. Из-за этого паренхиматозные клетки будут секретировать фактор роста эндотелия сосудов ( VEGF-A ), который является проангиогенным фактором роста. [9] Эти биологические сигналы активируют рецепторы эндотелиальных клеток, присутствующих в ранее существовавших кровеносных сосудах. Во-вторых, активированные эндотелиальные клетки, также известные как верхушечные клетки , [10] начинают выделять ферменты, называемые протеазами , которые разрушают базальную мембрану , позволяя эндотелиальным клеткам выйти из исходных (родительских) стенок сосудов. эндотелиальные клетки Затем пролиферируют в окружающий матрикс и образуют твердые отростки, соединяющие соседние сосуды. Клетки, которые пролиферируют, расположены позади верхушечных клеток и известны как стеблевые клетки . [10] Пролиферация этих клеток позволяет капиллярному отростку одновременно увеличиваться в длину.
Когда ростки тянутся к источнику ангиогенного стимула, эндотелиальные клетки мигрируют в тандеме , используя молекулы адгезии, называемые интегринами . Эти отростки затем образуют петли и становятся полноценным просветом сосуда по мере миграции клеток к месту ангиогенеза. Прорастание происходит со скоростью несколько миллиметров в день и позволяет новым сосудам прорастать через разрывы в сосудистой сети . Он заметно отличается от расщепления ангиогенеза, поскольку при нем образуются совершенно новые сосуды, а не расщепляются существующие сосуды.
Инвагинальный ангиогенез [ править ]
Инвагинальный ангиогенез , также известный как расщепляющий ангиогенез , представляет собой образование нового кровеносного сосуда путем разделения существующего кровеносного сосуда на две части.
Инвагинация кишечника впервые наблюдалась у новорожденных крыс. При этом типе формирования сосудов стенка капилляра простирается в просвет, разделяя один сосуд на две части. Выделяют четыре фазы инвагинационного ангиогенеза. Во-первых, две противоположные стенки капилляра создают зону контакта. Во-вторых, соединения эндотелиальных клеток реорганизуются и бислой сосуда перфорируется, что позволяет факторам роста и клеткам проникнуть в просвет. В-третьих, между двумя новыми сосудами в зоне контакта образуется ядро, заполненное перицитами и миофибробластами . Эти клетки начинают откладывать коллагеновые волокна в ядро, чтобы обеспечить внеклеточный матрикс для роста просвета сосуда. Наконец, ядро уточняется без изменений базовой структуры. Инвагинация важна, поскольку представляет собой реорганизацию существующих клеток. Это позволяет значительно увеличить количество капилляров без соответствующего увеличения количества эндотелиальных клеток . Это особенно важно при эмбриональном развитии, поскольку ресурсов для создания богатой среды недостаточно. микроциркуляторное русло новыми клетками каждый раз, когда развивается новый сосуд. [11]
Коалесцентный ангиогенез [ править ]
Коалесцентный ангиогенез — это тип ангиогенеза, который считается противоположностью инвагинационного ангиогенеза, при котором капилляры сливаются или сливаются, образуя более крупный кровеносный сосуд, тем самым увеличивая кровоток и кровообращение. [12] Коалесцентный ангиогенез вышел за пределы области эмбриологии. Предполагается, что он играет роль в формировании новой сосудистой сети, например, в опухоли. [13]
Физиология [ править ]
Механическая стимуляция [ править ]
Механическая стимуляция ангиогенеза изучена недостаточно. Существует значительное количество разногласий относительно напряжения сдвига , действующего на капилляры и вызывающего ангиогенез, хотя современные данные показывают, что усиление мышечных сокращений может усиливать ангиогенез. [14] Это может быть связано с увеличением выработки оксида азота во время физических упражнений. Оксид азота приводит к расширению кровеносных сосудов.
стимуляция Химическая
Химическая стимуляция ангиогенеза осуществляется различными ангиогенными белками, например интегринами и простагландинами, включая несколько факторов роста, например, VEGF, FGF.
Обзор [ править ]
| Стимулятор | Механизм |
|---|---|
| ФГФ | Стимулирует пролиферацию и дифференцировку эндотелиальных клеток, гладкомышечных клеток и фибробластов. |
| ВЭФР | Влияет на проходимость |
| VEGFR и NRP-1 | Интегрируйте сигналы выживания |
| Анг1 и Анг2 | Стабилизировать сосуды |
| PDGF (BB-гомодимер) и PDGFR | рекрутировать гладкомышечные клетки |
| Рецепторы TGF-β , эндоглина и TGF-β | ↑ внеклеточного матрикса производство |
| CCL2 | Привлекает лимфоциты к местам воспаления. |
| Гистамин | |
| Интегрины α V β 3 , α V β 5 (? [15] ) и α 5 β 1 | Связывают матричные макромолекулы и протеиназы |
| VE-кадгерин и CD31 | эндотелиального соединения молекулы |
| эфрин | Определить образование артерий или вен. |
| активаторы плазминогена | реконструирует внеклеточный матрикс , высвобождает и активирует факторы роста |
| ингибитор активатора плазминогена-1 | стабилизирует близлежащие сосуды |
| eNOS и ЦОГ-2 | |
| AC133 | регулирует ангиобластов дифференцировку |
| ИД1 / ИД3 | Регулирует эндотелиальную трансдифференцировку |
| 3 класса Семафорины | Модулирует адгезию, миграцию, пролиферацию и апоптоз эндотелиальных клеток. Изменяет проницаемость сосудов [16] |
| Nogo-A | Регулирует миграцию и пролиферацию эндотелиальных клеток. [17] Изменяет проницаемость сосудов. [18] |
ФГФ [ править ]
Семейство факторов роста фибробластов (FGF) с членами-прототипами FGF-1 (кислотный FGF) и FGF-2 (основной FGF) состоит на сегодняшний день по меньшей мере из 22 известных членов. [19] Большинство из них представляют собой одноцепочечные пептиды массой 16–18 кДа и обладают высоким сродством к гепарину и гепарансульфату. В целом FGF стимулируют различные клеточные функции путем связывания с FGF-рецепторами клеточной поверхности в присутствии протеогликанов гепарина. Семейство рецепторов FGF состоит из семи членов, и все рецепторные белки представляют собой одноцепочечные рецепторные тирозинкиназы, которые активируются посредством аутофосфорилирования, индуцированного механизмом димеризации рецептора, опосредованной FGF. Активация рецептора приводит к каскаду сигнальной трансдукции, который приводит к активации генов и разнообразным биологическим реакциям, включая дифференцировку клеток, пролиферацию и растворение матрикса, тем самым инициируя процесс митогенной активности, критический для роста эндотелиальных клеток, фибробластов и гладкомышечных клеток. .FGF-1, уникальный среди всех 22 членов семейства FGF, может связываться со всеми семью подтипами рецепторов FGF, что делает его членом семейства FGF с самым широким действием и мощным митогеном для различных типов клеток, необходимых для создания ангиогенного эффекта. ответ в поврежденных (гипоксических) тканях, где происходит активация рецепторов FGF. [20] FGF-1 стимулирует пролиферацию и дифференцировку всех типов клеток, необходимых для построения артериального сосуда, включая эндотелиальные клетки и гладкомышечные клетки; этот факт отличает FGF-1 от других проангиогенных факторов роста , таких как фактор роста эндотелия сосудов (VEGF), который в первую очередь стимулирует образование новых капилляров. [21] [22]
Помимо FGF-1, одной из наиболее важных функций фактора роста фибробластов-2 (FGF-2 или bFGF ) является стимулирование пролиферации эндотелиальных клеток и физическая организация эндотелиальных клеток в трубчатые структуры, тем самым способствуя ангиогенезу. FGF-2 является более мощным ангиогенным фактором, чем VEGF или PDGF ( фактор роста тромбоцитов ); однако он менее эффективен, чем FGF-1. Помимо стимуляции роста кровеносных сосудов, aFGF (FGF-1) и bFGF (FGF-2) играют важную роль в заживлении ран. Они стимулируют пролиферацию фибробластов и эндотелиальных клеток, которые приводят к ангиогенезу и развитию грануляционной ткани; оба увеличивают кровоснабжение и заполняют раневое пространство/полость на ранних стадиях процесса заживления ран.
ВЭФР [ править ]
Было продемонстрировано, что фактор роста эндотелия сосудов (VEGF) вносит основной вклад в ангиогенез, увеличивая количество капилляров в данной сети. Первоначальные исследования in vitro продемонстрировали, что эндотелиальные клетки бычьих капилляров будут пролиферировать и проявлять признаки трубчатых структур при стимуляции VEGF и bFGF , хотя результаты были более выраженными при VEGF. [23] Повышение регуляции VEGF является основным компонентом физиологической реакции на физическую нагрузку, и предполагается, что его роль в ангиогенезе может стать возможным методом лечения сосудистых повреждений. [24] [25] [26] [27] Исследования in vitro ясно демонстрируют, что VEGF является мощным стимулятором ангиогенеза, поскольку в присутствии этого фактора роста покрытые пластинками эндотелиальные клетки будут пролиферировать и мигрировать, в конечном итоге образуя трубчатые структуры, напоминающие капилляры. [14] VEGF вызывает мощный сигнальный каскад в эндотелиальных клетках. Связывание с рецептором VEGF-2 (VEGFR-2) запускает тирозинкиназный сигнальный каскад, который стимулирует выработку факторов, которые по-разному стимулируют проницаемость сосудов (eNOS, продуцирующая NO), пролиферацию/выживание (bFGF), миграцию (ICAM/VCAM/MMP). и, наконец, дифференцировка в зрелые кровеносные сосуды. Механически VEGF активируется мышечными сокращениями в результате увеличения притока крови к пораженным участкам. Увеличение потока также вызывает значительное увеличение продукции мРНК рецепторов VEGF 1 и 2. Увеличение продукции рецепторов означает, что мышечные сокращения могут вызвать активацию сигнального каскада, связанного с ангиогенезом. NO широко считается частью ангиогенного сигнального каскада, внося основной вклад в ангиогенный ответ, поскольку ингибирование NO значительно снижает эффекты ангиогенных факторов роста. Однако ингибирование NO во время тренировки не ингибирует ангиогенез, что указывает на то, что в ангиогенный ответ вовлечены и другие факторы. [14]
Ангиопоэтины [ править ]
Ангиопоэтины на Ang1 и Ang2 необходимы для формирования зрелых кровеносных сосудов, как показали исследования с нокаутом мышах . [28] Ang1 и Ang2 представляют собой белковые факторы роста, которые действуют путем связывания своих рецепторов Tie-1 и Tie-2 ; хотя это несколько спорно, похоже, что клеточные сигналы передаются в основном через Tie-2 ; физиологическая передача сигналов через Tie-1 хотя в некоторых работах показана и . Эти рецепторы представляют собой тирозинкиназы . Таким образом, они могут инициировать передачу сигналов в клетках , когда связывание лиганда вызывает димеризацию, которая инициирует фосфорилирование ключевых тирозинов.
ММП [ править ]
Другим важным фактором ангиогенеза является матриксная металлопротеиназа (ММП). ММП помогают разрушать белки, которые поддерживают прочность стенок сосудов. Этот протеолиз позволяет эндотелиальным клеткам выйти в интерстициальный матрикс, как это видно при прорастании ангиогенеза. Ингибирование ММП предотвращает образование новых капилляров . [29] Эти ферменты строго регулируются в процессе формирования сосудов, поскольку разрушение внеклеточного матрикса может снизить целостность микроциркуляторного русла. [14]
Dll4 [ править ]
Дельта-подобный лиганд 4 (Dll4) представляет собой белок с отрицательным регуляторным действием на ангиогенез. [30] [31] Dll4 представляет собой трансмембранный лиганд семейства рецепторов notch . Было проведено множество исследований, которые помогли определить последствия действия дельта-подобного лиганда 4. В одном исследовании, в частности, оценивалось влияние Dll4 на васкуляризацию и рост опухоли. [32] Для того чтобы опухоль росла и развивалась, она должна иметь соответствующую сосудистую сеть. Путь VEGF жизненно важен для развития сосудистой сети, которая, в свою очередь, способствует росту опухолей. Комбинированная блокада VEGF и Dll4 приводит к ингибированию прогрессирования опухоли и ангиогенеза по всей опухоли. Это происходит из-за затруднения передачи сигналов в эндотелиальных клетках, что останавливает пролиферацию и прорастание этих эндотелиальных клеток. При таком торможении клетки не растут бесконтрольно, поэтому рак на этом этапе останавливается. Однако если блокаду снять, клетки снова начнут пролиферацию. [33]
3 класса Семафорины
Семафорины класса 3 (SEMA3) регулируют ангиогенез путем модуляции адгезии эндотелиальных клеток , миграции, пролиферации, выживаемости и рекрутирования перицитов . [16] Кроме того, семафорины могут мешать VEGF-опосредованному ангиогенезу, поскольку как SEMA3, так и VEGF-A конкурируют за связывание рецептора нейропилина в эндотелиальных клетках. [34] [35] Следовательно, относительные уровни экспрессии SEMA3 и VEGF-A могут быть важны для ангиогенеза. [16]
ингибирование Химическое
Ингибитор ангиогенеза может быть эндогенным или поступать извне в виде лекарственного средства или компонента диеты .
Применение в медицине [ править ]
мишень терапевтическая как Ангиогенез
Ангиогенез может быть целью борьбы с такими заболеваниями, как болезни сердца, характеризующиеся либо плохой васкуляризацией, либо аномальной сосудистой сетью. [36] Применение специфических соединений, которые могут ингибировать или стимулировать создание новых кровеносных сосудов в организме, может помочь в борьбе с такими заболеваниями. Наличие кровеносных сосудов там, где их не должно быть, может повлиять на механические свойства ткани, увеличивая вероятность неудачи. Отсутствие кровеносных сосудов в восстанавливающейся или иным образом метаболически активной ткани может препятствовать восстановлению или другим важным функциям. Некоторые заболевания, такие как ишемические хронические раны , являются результатом нарушения или недостаточного образования кровеносных сосудов и могут лечиться путем местного расширения кровеносных сосудов, что приводит к доставке новых питательных веществ к месту повреждения и облегчению заживления. Другие заболевания, такие как возрастная дегенерация желтого пятна , могут быть вызваны локальным расширением кровеносных сосудов, препятствующим нормальным физиологическим процессам.
Современное клиническое применение принципа ангиогенеза можно разделить на два основных направления: антиангиогенная терапия, с которой начались ангиогенные исследования, и проангиогенная терапия. В то время как антиангиогенная терапия используется для борьбы с раком и злокачественными новообразованиями, [37] [38] Для размножения которых требуется обилие кислорода и питательных веществ, проангиогенная терапия изучается как вариант лечения сердечно-сосудистых заболеваний , которые являются причиной смерти номер один в западном мире . Одним из первых применений проангиогенных методов на людях было немецкое исследование по использованию фактора роста фибробластов 1 (FGF-1) для лечения ишемической болезни сердца. [21] [39] [40]
Что касается механизма действия , проангиогенные методы можно разделить на три основные категории: генная терапия , направленная на интересующие гены для амплификации или ингибирования; белково-заместительная терапия , которая в первую очередь воздействует на ангиогенные факторы роста, такие как FGF-1 или фактор роста эндотелия сосудов , VEGF; и клеточная терапия, которая включает имплантацию определенных типов клеток.
Есть еще серьезные, нерешенные проблемы, связанные с генной терапией. Трудности включают эффективную интеграцию терапевтических генов в геном клеток-мишеней, снижение риска нежелательного иммунного ответа, потенциальную токсичность, иммуногенность , воспалительные реакции и онкогенез , связанные с вирусными векторами, используемыми при имплантации генов, а также явную сложность генетического механизма. основе ангиогенеза. Наиболее часто встречающиеся у людей расстройства, такие как болезни сердца, высокое кровяное давление, диабет и болезнь Альцгеймера , скорее всего, вызваны комбинированным эффектом вариаций многих генов, и, таким образом, введение одного гена не может принести существенной пользы при лечении людей. такие заболевания. [ нужна ссылка ]
Напротив, проангиогенная белковая терапия использует четко определенные, точно структурированные белки с заранее определенными оптимальными дозами отдельного белка для болезненных состояний и с хорошо известными биологическими эффектами. [1] С другой стороны, препятствием для белковой терапии является способ доставки. Пероральный, внутривенный, внутриартериальный или внутримышечный пути введения белка не всегда столь эффективны, поскольку терапевтический белок может метаболизироваться или выводится из организма до того, как он попадет в ткань-мишень. Клеточная проангиогенная терапия все еще находится на ранней стадии исследований, и остается много открытых вопросов относительно лучших типов клеток и дозировок для использования.
Опухолевой ангиогенез
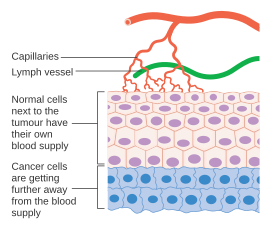
Раковые клетки — это клетки, которые потеряли способность контролируемо делиться. Злокачественная опухоль состоит из популяции быстро делящихся и растущих раковых клеток, в которых постепенно накапливаются мутации . Тем не менее, опухоли нуждаются в специальном кровоснабжении, чтобы обеспечить кислород и другие необходимые питательные вещества, необходимые им для того, чтобы вырасти за пределы определенного размера (обычно 1–2 мм). 3 ). [41] [42]
Опухоли вызывают рост кровеносных сосудов (ангиогенез) путем секреции различных факторов роста (например, VEGF ) и белков. Факторы роста, такие как bFGF и VEGF, могут вызывать рост капилляров в опухоли, которые, как подозревают некоторые исследователи, поставляют необходимые питательные вещества, позволяя опухоли расширяться. В отличие от нормальных кровеносных сосудов, кровеносные сосуды опухоли расширены и имеют неправильную форму. [43] Другие клиницисты полагают, что ангиогенез на самом деле служит путем отходов, удаляя конечные биологические продукты, выделяемые быстро делящимися раковыми клетками. В любом случае ангиогенез является необходимым и необходимым шагом для перехода от небольшого безвредного скопления клеток, размером с металлический шарик на конце шариковой ручки, в большую опухоль. Ангиогенез также необходим для распространения опухоли или метастазирования . [7] Одиночные раковые клетки могут оторваться от сформировавшейся солидной опухоли, проникнуть в кровеносный сосуд и быть перенесены в отдаленное место, где они могут имплантироваться и начать рост вторичной опухоли. Имеющиеся данные свидетельствуют о том, что кровеносные сосуды в данной солидной опухоли на самом деле могут представлять собой мозаичные сосуды, состоящие из эндотелиальных клеток и опухолевых клеток. [7] Эта мозаичность позволяет существенно выделять опухолевые клетки в сосудистую сеть, что, возможно, способствует появлению циркулирующих опухолевых клеток в периферической крови пациентов со злокачественными новообразованиями. [44] Последующий рост таких метастазов также потребует снабжения питательными веществами и кислородом , а также путей удаления отходов.
Эндотелиальные клетки долгое время считались генетически более стабильными, чем раковые клетки. Эта геномная стабильность дает преимущество нацеливания на эндотелиальные клетки с помощью антиангиогенной терапии по сравнению с химиотерапией , направленной на раковые клетки, которые быстро мутируют и приобретают лекарственную устойчивость к лечению. По этой причине эндотелиальные клетки считаются идеальной мишенью для терапии, направленной против них. [45]
Образование опухолевых кровеносных сосудов [ править ]
Механизм образования кровеносных сосудов путем ангиогенеза инициируется спонтанным делением опухолевых клеток вследствие мутации. Затем опухолевые клетки высвобождают ангиогенные стимуляторы. Затем они попадают в уже сформировавшиеся близлежащие кровеносные сосуды и активируют рецепторы их эндотелиальных клеток. Это вызывает высвобождение протеолитических ферментов из сосудистой сети. Эти ферменты воздействуют на определенную точку кровеносного сосуда и вызывают образование пор. Это точка, из которой вырастет новый кровеносный сосуд. Причина, по которой опухолевые клетки нуждаются в кровоснабжении, заключается в том, что они не могут вырасти более чем на 2-3 миллиметра в диаметре без налаженного кровоснабжения, что эквивалентно примерно 50-100 клеткам. [46] Некоторые исследования показали, что сосуды, образующиеся внутри опухолевой ткани, имеют более неравномерную форму и большие размеры, что также связано с худшим прогнозом. [47] [48]
при сердечно- Ангиогенез сосудистых заболеваниях
Ангиогенез представляет собой превосходную терапевтическую мишень для лечения сердечно-сосудистых заболеваний. Это мощный физиологический процесс, лежащий в основе естественного ответа нашего организма на уменьшение кровоснабжения жизненно важных органов, а именно неоангиогенез : образование новых коллатеральных сосудов для преодоления ишемического инсульта. [21] Было проведено большое количество доклинических исследований белковой, генной и клеточной терапии на животных моделях ишемии сердца, а также на моделях заболеваний периферических артерий. Воспроизводимые и заслуживающие доверия успехи в этих ранних исследованиях на животных привели к большому энтузиазму по поводу того, что этот новый терапевтический подход может быть быстро воплощен в клиническую пользу для миллионов пациентов в западном мире с этими расстройствами. Однако десятилетие клинических испытаний генной и белковой терапии, предназначенной для стимуляции ангиогенеза в тканях и органах с недостаточной перфузией, привело к одному разочарованию за другим. Хотя все эти доклинические данные, которые давали большие надежды на переход терапии ангиогенеза от животных к человеку, были так или иначе включены в ранние стадии клинических испытаний, FDA до сих пор (2007 г.) настаивало на том, что первичные Конечной точкой для одобрения ангиогенного агента должно быть улучшение физических показателей пролеченных пациентов. [49]
Эти неудачи позволили предположить, что либо это неправильные молекулярные мишени для индукции неоваскуляризации, либо что их можно эффективно использовать только при правильном формулировании и введении, либо что их представление в контексте общего клеточного микроокружения может играть жизненно важную роль в их полезности. Может оказаться необходимым представить эти белки таким образом, чтобы имитировать естественные сигнальные события, включая концентрацию , пространственные и временные профили, а также их одновременное или последовательное представление с другими соответствующими факторами. [50]
Упражнение [ править ]
Ангиогенез обычно связан с аэробными упражнениями и упражнениями на выносливость . В то время как артериогенез вызывает изменения в сети, которые позволяют значительно увеличить общий объем кровотока в сети, ангиогенез вызывает изменения, которые обеспечивают большую доставку питательных веществ в течение длительного периода времени. Капилляры предназначены для обеспечения максимальной эффективности доставки питательных веществ, поэтому увеличение количества капилляров позволяет сети доставлять больше питательных веществ за тот же промежуток времени. Большее количество капилляров также обеспечивает больший кислородный обмен в сети. Это жизненно важно для тренировки выносливости, поскольку позволяет человеку продолжать тренировки в течение длительного периода времени. Однако нет экспериментальных данных, свидетельствующих о том, что при упражнениях на выносливость для увеличения максимальной доставки кислорода требуется усиление капиллярности. [14]
Дегенерация желтого пятна [ править ]
Сверхэкспрессия VEGF помимо стимуляции ангиогенеза вызывает повышение проницаемости кровеносных сосудов. При влажной дегенерации желтого пятна VEGF вызывает пролиферацию капилляров сетчатки. Поскольку увеличение ангиогенеза также вызывает отек , кровь и другие жидкости сетчатки попадают в сетчатку , вызывая потерю зрения. Антиангиогенные препараты, действующие на пути VEGF, в настоящее время успешно используются для лечения этого типа дегенерации желтого пятна.
Тканеинженерные конструкции [ править ]
Ангиогенез сосудов из организма хозяина в имплантированные тканеинженерные конструкции имеет важное значение. Успешная интеграция часто зависит от тщательной васкуляризации конструкции, поскольку она обеспечивает кислород и питательные вещества и предотвращает некроз в центральных областях имплантата. [51] Было показано, что PDGF стабилизирует васкуляризацию в коллаген- гликозаминогликановых каркасах. [52]
История [ править ]
Первое сообщение об ангиогенезе можно отнести к книге «Трактат о крови, воспалении и огнестрельных ранениях», опубликованной в 1794 году, где шотландского анатома Джона Хантера были обобщены результаты исследований . В своем исследовании Хантер наблюдал процесс роста новых кровеносных сосудов у кроликов. Однако он не придумал термин «ангиогенез», который сейчас широко используется учеными. Хантер также ошибочно приписывал процесс роста новых кровеносных сосудов эффекту врожденного жизненного принципа крови. Считается, что термин «ангиогенез» появился только в 1900-х годах. Начало современных исследований ангиогенеза ознаменовано докладом Джуды Фолкмана о ключевой роли ангиогенеза в опухолевом росте. [8] [53] [54]
Количественная оценка [ править ]
Количественная оценка параметров сосудистой сети, таких как плотность микрососудов, имеет различные сложности из-за преимущественного окрашивания или ограниченного представления тканей на гистологических срезах. Недавние исследования показали полную трехмерную реконструкцию сосудистой структуры опухоли и количественную оценку сосудистых структур в целых опухолях на животных моделях. [55]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Сантулли Дж., изд. (2013). Понимание ангиогенеза из систематического обзора . Нью-Йорк: Нова Сайенс. ISBN 978-1-62618-114-4 .
- ^ Jump up to: Перейти обратно: а б Дадли AC, Гриффиоен AW (август 2023 г.). «Патологический ангиогенез: механизмы и терапевтические стратегии» . Ангиогенез . 26 (3): 313–347. дои : 10.1007/s10456-023-09876-7 . ПМЦ 10105163 . ПМИД 37060495 .
- ^ Бирбрайр А., Чжан Т., Ван З.М., Месси М.Л., Олсон Дж.Д., Минц А., Дельбоно О. (июль 2014 г.). «Перициты типа 2 участвуют в нормальном и опухолевом ангиогенезе» . Американский журнал физиологии. Клеточная физиология . 307 (1): C25–C38. doi : 10.1152/ajpcell.00084.2014 . ПМК 4080181 . ПМИД 24788248 .
- ^ Ницше Б., Ронг В.В., Геде А., Хоффманн Б., Скарпа Ф., Кюблер В.М. и др. (февраль 2022 г.). «Коалесцентный ангиогенез – свидетельство новой концепции созревания сосудистой сети» . Ангиогенез . 25 (1): 35–45. дои : 10.1007/s10456-021-09824-3 . ПМЦ 8669669 . ПМИД 34905124 .
- ^ Рисау В., Фламме I (1995). «Васкулогенез». Ежегодный обзор клеточной биологии и биологии развития . 11 : 73–91. дои : 10.1146/annurev.cb.11.110195.000445 . ПМИД 8689573 .
- ^ Фламме И, Фрелих Т, Рисау В (ноябрь 1997 г.). «Молекулярные механизмы васкулогенеза и эмбрионального ангиогенеза». Журнал клеточной физиологии . 173 (2): 206–210. doi : 10.1002/(SICI)1097-4652(199711)173:2<206::AID-JCP22>3.0.CO;2-C . ПМИД 9365523 . S2CID 36723610 .
- ^ Jump up to: Перейти обратно: а б с Милошевич В., Эдельманн Р.Дж., Фосс Дж.Х., Остман А., Акслен Л.А. (2022). «Молекулярные фенотипы эндотелиальных клеток злокачественных опухолей». В Акслен Л.А., Уотник Р.С. (ред.). Биомаркеры микроокружения опухоли . Чам: Международное издательство Springer. стр. 31–52. дои : 10.1007/978-3-030-98950-7_3 . ISBN 978-3-030-98950-7 .
- ^ Jump up to: Перейти обратно: а б Пенн Дж.С. (11 марта 2008 г.). Ангиогенез сетчатки и хориоидеи . Спрингер. стр. 119–. ISBN 978-1-4020-6779-2 . Проверено 26 июня 2010 г.
- ^ Адэр TH, Монтани JP. Ангиогенез. Сан-Рафаэль (Калифорния): Morgan & Claypool Life Sciences; 2010. Глава 1, Обзор ангиогенеза. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK53238/.
- ^ Jump up to: Перейти обратно: а б Уиверс Х., Скаер Х. (июль 2014 г.). «Концевые клетки: главные регуляторы тубулогенеза?» . Семинары по клеточной биологии и биологии развития . 31 (100): 91–99. дои : 10.1016/j.semcdb.2014.04.009 . ПМК 4071413 . ПМИД 24721475 .
- ^ Бурри П.Х., Глущук Р., Джонов В. (ноябрь 2004 г.). «Инвагинальный ангиогенез: его возникновение, характеристики и значение» . Динамика развития . 231 (3): 474–488. дои : 10.1002/dvdy.20184 . ПМИД 15376313 . S2CID 35018922 .
- ^ Ницше Б., Ронг В.В., Геде А., Хоффманн Б., Скарпа Ф., Кюблер В.М. и др. (февраль 2022 г.). «Коалесцентный ангиогенез – свидетельство новой концепции созревания сосудистой сети» . Ангиогенез . 25 (1): 35–45. дои : 10.1007/s10456-021-09824-3 . ПМЦ 8669669 . ПМИД 34905124 .
- ^ Пеццелла Ф, Кербель РС (февраль 2022 г.). «О коалесцентном ангиогенезе и удивительной гибкости кровеносных сосудов» . Ангиогенез . 25 (1): 1–3. дои : 10.1007/s10456-021-09825-2 . ПМИД 34993716 . S2CID 254188870 .
- ^ Jump up to: Перейти обратно: а б с д и Прайор Б.М., Ян Х.Т., Терджунг Р.Л. (сентябрь 2004 г.). «Что заставляет сосуды расти при тренировках?». Журнал прикладной физиологии . 97 (3): 1119–1128. doi : 10.1152/japplphysicalol.00035.2004 . ПМИД 15333630 .
- ^ Возможно, ингибитор ангиогенеза: Шеппард Д. (октябрь 2002 г.). «Эндотелиальные интегрины и ангиогенез: уже не так просто» . Журнал клинических исследований . 110 (7): 913–914. дои : 10.1172/JCI16713 . ПМЦ 151161 . ПМИД 12370267 .
- ^ Jump up to: Перейти обратно: а б с Меколлари В., Ньювенхейс Б., Верхааген Дж. (2014). «Взгляд на роль передачи сигналов семафоринов класса III при травмах центральной нервной системы» . Границы клеточной нейронауки . 8 : 328. дои : 10.3389/fncel.2014.00328 . ПМК 4209881 . ПМИД 25386118 .
- ^ Руст Р., Грённерт Л., Гантнер С., Энцлер А., Малдерс Г., Вебер Р.З. и др. (июль 2019 г.). «Таргетная терапия Nogo-A способствует восстановлению сосудов и функциональному восстановлению после инсульта» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (28): 14270–14279. Бибкод : 2019PNAS..11614270R . дои : 10.1073/pnas.1905309116 . ПМК 6628809 . ПМИД 31235580 .
- ^ Руст Р., Вебер Р.З., Грённерт Л., Малдерс Г., Маурер М.А., Хофер А.С. и др. (декабрь 2019 г.). «Анти-Nogo-A антитела предотвращают утечку из сосудов и действуют как проангиогенные факторы после инсульта» . Научные отчеты . 9 (1): 20040. Бибкод : 2019НатСР...920040Р . дои : 10.1038/s41598-019-56634-1 . ПМЦ 6934709 . ПМИД 31882970 .
- ^ Орниц Д.М., Ито Н. (2001). «Факторы роста фибробластов» . Геномная биология . 2 (3): ОБЗОРЫ3005. doi : 10.1186/gb-2001-2-3-reviews3005 . ПМК 138918 . ПМИД 11276432 .
- ^ Блабер М., ДиСальво Дж., Томас К.А. (февраль 1996 г.). «Рентгеновская кристаллическая структура человеческого кислого фактора роста фибробластов». Биохимия . 35 (7): 2086–2094. CiteSeerX 10.1.1.660.7607 . дои : 10.1021/bi9521755 . ПМИД 8652550 .
- ^ Jump up to: Перейти обратно: а б с Стегманн Т.Дж. (декабрь 1998 г.). «FGF-1: фактор роста человека в индукции неоангиогенеза». Экспертное мнение об исследуемых лекарствах . 7 (12): 2011–2015. дои : 10.1517/13543784.7.12.2011 . ПМИД 15991943 .
- ^ Хурана Р., Саймонс М. (апрель 2003 г.). «Итоги исследований ангиогенеза с использованием фактора роста фибробластов при запущенных атеросклеротических заболеваниях». Тенденции сердечно-сосудистой медицины . 13 (3): 116–122. дои : 10.1016/S1050-1738(02)00259-1 . ПМИД 12691676 .
- ^ Гото Ф, Гото К, Вайндел К, Фолкман Дж (ноябрь 1993 г.). «Синергетическое воздействие фактора роста эндотелия сосудов и основного фактора роста фибробластов на пролиферацию и образование тяжей эндотелиальных клеток бычьих капилляров в коллагеновых гелях». Лабораторные исследования; Журнал технических методов и патологии . 69 (5): 508–517. ПМИД 8246443 .
- ^ Дин Ю.Х., Луан К.Д., Ли Дж., Рафолс Дж.А., Гутинконда М., Диас Ф.Г., Дин Ю. (декабрь 2004 г.). «Вызванная физическими упражнениями сверхэкспрессия ангиогенных факторов и уменьшение ишемии/реперфузии при инсульте» . Текущие нейроваскулярные исследования . 1 (5): 411–420. дои : 10.2174/1567202043361875 . ПМИД 16181089 . S2CID 22015361 . Архивировано из оригинала 19 апреля 2012 года.
- ^ Гэвин Т.П., Робинсон С.Б., Йегер Р.С., Англия Дж.А., Нифонг Л.В., Хикнер Р.К. (январь 2004 г.). «Реакция ангиогенного фактора роста на острую системную тренировку скелетных мышц человека». Журнал прикладной физиологии . 96 (1): 19–24. doi : 10.1152/japplphysicalol.00748.2003 . ПМИД 12949011 . S2CID 12750224 .
- ^ Краус Р.М., Столлингс Х.В., Йегер Р.К., Гэвин Т.П. (апрель 2004 г.). «Реакция VEGF циркулирующей плазмы на физические упражнения у мужчин, ведущих сидячий образ жизни и тренирующихся на выносливость». Журнал прикладной физиологии . 96 (4): 1445–1450. doi : 10.1152/japplphysicalol.01031.2003 . ПМИД 14660505 . S2CID 21090407 .
- ^ Ллойд П.Г., Прайор Б.М., Ян Х.Т., Терджунг Р.Л. (май 2003 г.). «Экспрессия ангиогенного фактора роста в скелетных мышцах крыс в ответ на тренировку». Американский журнал физиологии. Физиология сердца и кровообращения . 284 (5): H1668–H1678. дои : 10.1152/ajpheart.00743.2002 . ПМИД 12543634 .
- ^ Терстон Дж. (октябрь 2003 г.). «Роль ангиопоэтинов и тирозинкиназ Tie-рецепторов в ангиогенезе и лимфангиогенезе». Исследования клеток и тканей . 314 (1): 61–68. дои : 10.1007/s00441-003-0749-6 . ПМИД 12915980 . S2CID 2529783 .
- ^ Хаас Т.Л., Милькевич М., Дэвис С.Дж., Чжоу А.Л., Эггинтон С., Браун М.Д. и др. (октябрь 2000 г.). «Активность матриксной металлопротеиназы необходима для индуцированного активностью ангиогенеза в скелетных мышцах крыс». Американский журнал физиологии. Физиология сердца и кровообращения . 279 (4): H1540–H1547. дои : 10.1152/ajpheart.2000.279.4.H1540 . ПМИД 11009439 . S2CID 2543076 .
- ^ Лобов И.Б., Ренард Р.А., Пападопулос Н., Гейл Н.В., Терстон Г., Янкопулос Г.Д., Виганд С.Дж. (февраль 2007 г.). «Дельта-подобный лиганд 4 (Dll4) индуцируется VEGF как негативный регулятор ангиогенного прорастания» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (9): 3219–3224. Бибкод : 2007PNAS..104.3219L . дои : 10.1073/pnas.0611206104 . ПМК 1805530 . ПМИД 17296940 .
- ^ Хеллстрем М., Фнг Л.К., Хофманн Дж.Дж., Валлгард Э., Коултас Л., Линдблом П. и др. (февраль 2007 г.). «Передача сигналов Dll4 через Notch1 регулирует образование кончиковых клеток во время ангиогенеза». Природа . 445 (7129): 776–780. Бибкод : 2007Natur.445..776H . дои : 10.1038/nature05571 . ПМИД 17259973 . S2CID 4407198 .
- ^ Сегарра М., Уильямс К.К., Сьерра Мде Л., Бернардо М., Маккормик П.Дж., Марик Д. и др. (сентябрь 2008 г.). «Активация Dll4 передачи сигналов Notch снижает васкуляризацию опухоли и ингибирует рост опухоли» . Кровь . 112 (5): 1904–11. дои : 10.1182/blood-2007-11-126045 . ПМК 2518892 . ПМИД 18577711 .
- ^ Ли Д., Ким Д., Чхве Ю.Б., Кан К., Сунг Э.С., Ан Дж.Х. и др. (июль 2016 г.). «Одновременная блокада VEGF и Dll4 с помощью HD105, биспецифического антитела, ингибирует прогрессирование опухоли и ангиогенез» . МАБ . 8 (5): 892–904. дои : 10.1080/19420862.2016.1171432 . ПМЦ 4968104 . ПМИД 27049350 .
- ^ Сокер С., Такашима С., штаб-квартира Мяо, Нойфельд Г., Клагсбрун М. (март 1998 г.). «Нейуропилин-1 экспрессируется эндотелиальными и опухолевыми клетками как специфичный для изоформы рецептор фактора роста эндотелия сосудов» . Клетка . 92 (6): 735–745. дои : 10.1016/s0092-8674(00)81402-6 . ПМИД 9529250 . S2CID 547080 .
- ^ Херцог Б., Пелле-Мэни С., Бриттон Г., Харцулакис Б., Закари И.С. (август 2011 г.). «Связывание VEGF с NRP1 необходимо для стимуляции VEGF миграции эндотелиальных клеток, образования комплекса между NRP1 и VEGFR2 и передачи сигналов посредством фосфорилирования FAK Tyr407» . Молекулярная биология клетки . 22 (15): 2766–2776. дои : 10.1091/mbc.E09-12-1061 . ПМК 3145551 . ПМИД 21653826 .
- ^ Феррара Н., Кербель РС (декабрь 2005 г.). «Ангиогенез как терапевтическая мишень». Природа . 438 (7070): 967–974. Бибкод : 2005Natur.438..967F . дои : 10.1038/nature04483 . ПМИД 16355214 . S2CID 1183610 .
- ^ Фолкман Дж., Клагсбрун М. (январь 1987 г.). «Ангиогенные факторы». Наука . 235 (4787): 442–447. Бибкод : 1987Sci...235..442F . дои : 10.1126/science.2432664 . ПМИД 2432664 .
- ^ Фолкман Дж. (сентябрь 1996 г.). «Борьба с раком путем воздействия на его кровоснабжение». Научный американец . 275 (3): 150–154. Бибкод : 1996SciAm.275c.150F . doi : 10.1038/scientificamerican0996-150 . ПМИД 8701285 .
- ^ Стегманн Т.Дж., Хопперт Т., Шнайдер А., Гемейнхардт С., Кехер М., Ибинг Р., Штрупп Г. (сентябрь 2000 г.). «[Индукция неоангиогенеза миокарда факторами роста человека. Новый терапевтический подход при ишемической болезни сердца]». Герц (на немецком языке). 25 (6): 589–599. дои : 10.1007/PL00001972 . ПМИД 11076317 . S2CID 21240045 .
- ^ Фолкман Дж. (февраль 1998 г.). «Ангиогенная терапия сердца человека» . Тираж . 97 (7): 628–629. дои : 10.1161/01.CIR.97.7.628 . ПМИД 9495294 .
- ^ Макдугалл С.Р., Андерсон А.Р., капеллан М.А. (август 2006 г.). «Математическое моделирование динамического адаптивного ангиогенеза, индуцированного опухолью: клинические последствия и терапевтические стратегии». Журнал теоретической биологии . 241 (3): 564–589. Бибкод : 2006JThBi.241..564M . дои : 10.1016/j.jtbi.2005.12.022 . ПМИД 16487543 .
- ^ Спилл Ф, Герреро П., Аларкон Т., Майни ПК, Бирн Х.М. (февраль 2015 г.). «Мезоскопическое и континуальное моделирование ангиогенеза» . Журнал математической биологии . 70 (3): 485–532. arXiv : 1401.5701 . дои : 10.1007/s00285-014-0771-1 . ПМЦ 5320864 . ПМИД 24615007 .
- ^ Гонсалес-Перес Р.Р., Руэда Б.Р. (2013). Регуляторы опухолевого ангиогенеза (первое изд.). Бока-Ратон: Тейлор и Фрэнсис. п. 347. ИСБН 978-1-4665-8097-8 . Проверено 2 октября 2014 г.
- ^ Аллард В.Дж., Матера Дж., Миллер М.К., Реполлет М., Коннелли М.К., Рао С. и др. (октябрь 2004 г.). «Опухолевые клетки циркулируют в периферической крови всех основных карцином, но не у здоровых людей или пациентов с доброкачественными заболеваниями» . Клинические исследования рака . 10 (20): 6897–6904. дои : 10.1158/1078-0432.CCR-04-0378 . ПМИД 15501967 .
- ^ Багри А., Курос-Мехр Х., Леонг К.Г., Пахарь Г.Д. (март 2010 г.). «Использование адъювантной терапии против VEGF при раке: проблемы и обоснование». Тенденции молекулярной медицины . 16 (3): 122–132. doi : 10.1016/j.molmed.2010.01.004 . ПМИД 20189876 .
- ^ Нисида Н., Яно Х., Нисида Т., Камура Т., Кодзиро М. (сентябрь 2006 г.). «Ангиогенез при раке» . Сосудистое здоровье и управление рисками . 2 (3): 213–219. дои : 10.2147/vhrm.2006.2.3.213 . ЧВК 1993983 . ПМИД 17326328 .
- ^ Милошевич В., Эдельманн Р.Дж., Винге И., Стрелл С., Межеевский А., Кнутсвик Г. и др. (июль 2023 г.). «Размер сосуда как маркер выживаемости при раке молочной железы, положительном по рецепторам эстрогена» . Исследование и лечение рака молочной железы . 200 (2): 293–304. дои : 10.1007/s10549-023-06974-4 . ПМЦ 10241708 . ПМИД 37222874 .
- ^ Микалсен Л.Т., Дакал Х.П., Бруланд О.С., Науме Б., Борген Э., Несланд Дж.М., Олсен Д.Р. (11 октября 2013 г.). Аоки I (ред.). «Клиническое влияние среднего размера и прочности сосудов у пациентов с раком молочной железы» . ПЛОС ОДИН . 8 (10): е75954. Бибкод : 2013PLoSO...875954M . дои : 10.1371/journal.pone.0075954 . ПМЦ 3795733 . ПМИД 24146798 .
- ^ Хариавала, доктор медицинских наук, Селлке, FW (июнь 1997 г.). «Ангиогенез и сердце: терапевтические последствия» . Журнал Королевского медицинского общества . 90 (6): 307–311. дои : 10.1177/014107689709000604 . ПМК 1296305 . ПМИД 9227376 .
- ^ Цао Л., Mooney DJ (ноябрь 2007 г.). «Пространственно-временной контроль над передачей сигналов факторов роста для терапевтической неоваскуляризации» . Обзоры расширенной доставки лекарств . 59 (13): 1340–1350. дои : 10.1016/j.addr.2007.08.012 . ПМК 2581871 . ПМИД 17868951 .
- ^ Рувкема Дж., Хадемоссейни А. (сентябрь 2016 г.). «Васкуляризация и ангиогенез в тканевой инженерии: помимо создания статических сетей» . Тенденции в биотехнологии . 34 (9): 733–745. дои : 10.1016/j.tibtech.2016.03.002 . ПМИД 27032730 .
- ^ до Амарал Р.Дж., Кавана Б., О'Брайен Ф.Дж., Кирни С.Дж. (февраль 2019 г.). «Фактор роста тромбоцитов стабилизирует васкуляризацию в коллагеново-гликозаминогликановых каркасах in vitro» . Журнал тканевой инженерии и регенеративной медицины . 13 (2): 261–273. дои : 10.1002/терм.2789 . ПМИД 30554484 . S2CID 58767660 .
- ^ Лензи П., Боччи Дж., Натале Дж. (апрель 2016 г.). «Джон Хантер и происхождение термина «ангиогенез» ». Ангиогенез . 19 (2). ООО «Спрингер Сайенс энд Бизнес Медиа»: 255–256. дои : 10.1007/s10456-016-9496-7 . hdl : 11568/795270 . ПМИД 26842740 . S2CID 254189385 .
- ^ Адэр Т.Х., Монтани Дж.П. (2010). «История» . Ангиогенез . Морган и Клейпул Науки о жизни. doi : 10.4199/C00017ED1V01Y201009ISP009 (неактивен 31 января 2024 г.). ПМИД 21452444 . Проверено 20 июля 2023 г.
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Чиен С.С., Кемпсон И.М., Ван С.Л., Чен Х.Х., Хву Ю, Чен Нью-Йорк и др. (май – июнь 2013 г.). «Полное микромасштабное профилирование опухолевого микроангиогенеза: микрорадиологическая методология раскрывает фундаментальные аспекты опухолевого ангиогенеза и дает ряд количественных параметров для его характеристики». Достижения биотехнологии . 31 (3): 396–401. doi : 10.1016/j.biotechadv.2011.12.001 . ПМИД 22193280 .