Ремоделирование сосудов у эмбриона
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|

Ремоделирование сосудов — это процесс, который происходит, когда незрелое сердце начинает сокращаться, проталкивая жидкость через раннюю сосудистую сеть . Процесс обычно начинается на 22-й день и продолжается до десятой недели эмбриогенеза человека . Это первое прохождение жидкости инициирует сигнальный каскад и движение клеток на основе физических сигналов , включая напряжение сдвига и окружное напряжение , которое необходимо для ремоделирования сосудистой сети, артериально - венозной идентичности, ангиогенеза и регуляции генов посредством механотрансдукции . Этот эмбриональный процесс необходим для будущей стабильности зрелой сосудистой сети. [2]
Васкулогенез — это первоначальное формирование компонентов сети кровеносных сосудов или сосудистого дерева. Это продиктовано генетическими факторами и не имеет никакой внутренней функции, кроме как заложить предварительный контур системы кровообращения . Как только поток жидкости начинается, биомеханические и гемодинамические воздействия применяются к системе, созданной васкулогенезом, и может начаться активный процесс ремоделирования.
Известно, что физические сигналы, такие как давление , скорость , характер потока и напряжение сдвига, действуют на сосудистую сеть разными способами, включая морфогенез ветвления , расширение сосудов в областях с интенсивным потоком, ангиогенез и развитие венозных клапанов. Механотрансдукция этих физических сигналов к эндотелиальным и гладкомышечным клеткам сосудистой стенки может также запускать активацию или подавление определенных генов, которые отвечают за вазодилатацию , выравнивание клеток и другие факторы, смягчающие сдвиговое напряжение. Эта связь между генетикой и окружающей средой не совсем понятна, но исследователи пытаются прояснить ее, комбинируя надежные генетические методы, такие как генетически удаленные модели организмов и тканей , с новыми технологиями, разработанными для измерения и отслеживания режимов потока, профилей скорости и колебаний давления. в естественных условиях . [2]
И in vivo исследование , и моделирование являются необходимыми инструментами для понимания этого сложного процесса. Ремоделирование сосудов имеет отношение к заживлению ран и правильной интеграции трансплантации тканей и донорства органов . Содействие активному процессу ремоделирования в некоторых случаях может помочь пациентам быстрее выздороветь и сохранить функциональное использование донорских тканей. Однако, помимо заживления ран, хроническое ремоделирование сосудов у взрослых часто является симптомом сердечно -сосудистых заболеваний . Таким образом, более глубокое понимание этого биомедицинского явления может помочь в разработке терапевтических или профилактических мер для борьбы с такими заболеваниями, как атеросклероз .
Исторический взгляд
[ редактировать ]Более 100 лет назад Тома заметил, что увеличение местного кровотока приводит к увеличению диаметра сосудов , и даже предположил, что кровоток может быть ответственен за рост и развитие кровеносных сосудов. [3] Впоследствии Чепмен в 1918 году обнаружил, что удаление сердца куриного эмбриона нарушает процесс ремоделирования, но первоначальный рисунок сосудов, заложенный васкулогенезом, остается нетронутым. Затем, в 1926 году, Мюррей предположил, что диаметр сосуда пропорционален величине напряжения сдвига на стенке сосуда; то есть сосуды активно адаптировались к моделям потока на основе физических сигналов окружающей среды, таких как напряжение сдвига.
Химические основы морфогенеза », написанная в 1952 году математиком и ученым-компьютерщиком Аланом Тьюрингом, выступала за различные биологические модели, основанные на молекулярной диффузии питательных веществ . [4] Однако диффузная модель развития сосудов, по-видимому, не соответствует сложности капиллярного русла и переплетенной сети артерий и вен. [4] [5] В 2000 г. Флёри предположил, что вместо диффузионных молекул, несущих ответственность за морфогенез ветвления сосудистого дерева, морфоген может быть задействован дальнего действия. В этой модели бегущая волна давления будет воздействовать на сосудистую систему посредством напряжения сдвига, перестраивая ветви в конфигурацию с наименьшей энергией, расширяя сосуды, несущие увеличенный кровоток, и перестраивая сети после начала потока жидкости. [4] [6] Известно, что механические силы могут оказывать существенное влияние на морфологию и сложность сосудистого дерева. [5] [6] Однако эти силы сравнительно мало влияют на диффузию питательных веществ, и поэтому кажется маловероятным, что приобретение питательных веществ и кислорода играет существенную роль в ремоделировании сосудов эмбриона. [5]
Сейчас это широко признано [ ласковые слова ] [ кем? ] что ремоделирование сосудов у эмбриона представляет собой процесс, отличный от васкулогенеза; однако эти два процесса неразрывно связаны. Васкулогенез происходит до ремоделирования сосудов, но является необходимым шагом в развитии сети кровеносных сосудов и имеет значение для идентификации сосудов как артериальных или венозных. Как только начинается сокращение сердца, ремоделирование сосудов прогрессирует посредством взаимодействия сил, возникающих в результате биомеханических сигналов и динамики жидкости , которые посредством механотрансдукции преобразуются в изменения на клеточном и генетическом уровнях.
Васкулогенез
[ редактировать ]
Васкулогенез – это формирование ранней сосудистой сети, заложенное генетическими факторами. [7] структуры, называемые островками образуются В мезодермальном слое желточного мешка в результате клеточной дифференцировки гемангиобластов кровяными в эндотелиальные и эритроциты . [7] Затем формируется капиллярное сплетение , когда эндотелиальные клетки мигрируют наружу от островков крови и образуют случайную сеть непрерывных нитей. [7] Затем эти нити подвергаются процессу, называемому люменизацией, — спонтанному перераспределению эндотелиальных клеток из твердого тяжа в полую трубку. [8]
Внутри эмбриона формируется дорсальная аорта , которая в конечном итоге соединяет сердце с капиллярным сплетением желточного мешка. [7] Это образует замкнутую систему жестких эндотелиальных трубок. Даже на такой ранней стадии процесса васкулогенеза, до начала кровотока, участки системы трубок могут экспрессировать эфрины или нейропилины , генетические маркеры артериальной или венозной идентичности соответственно. [7] Эти идентичности все еще несколько гибки, но первоначальная характеристика важна для процесса эмбрионального ремоделирования. [2]
Ангиогенез также способствует усложнению начальной сети; извергающиеся эндотелиальные зачатки формируются в результате процесса, подобного экструзии , который вызывается экспрессией фактора роста эндотелия сосудов (VEGF). [8] Эти эндотелиальные зачатки растут от родительского сосуда, образуя более мелкие дочерние сосуды, достигающие новой территории. [8] Инвагинация , явление расщепления одной трубки с образованием двух ветвящихся трубок, также способствует ангиогенезу. [8] Ангиогенез обычно отвечает за колонизацию отдельных систем органов кровеносными сосудами, тогда как васкулогенез закладывает начальные трубопроводы сети. [9] Известно также, что ангиогенез происходит во время ремоделирования сосудов. [9]
Артериально-венозная идентичность
[ редактировать ]
Классификация ангиобластов на артериальные или венозные клетки необходима для формирования правильной морфологии ветвления. [2] Артериальные сегменты ранней сосудистой сети экспрессируют ephrinB2 и DLL4, тогда как венозные сегменты экспрессируют нейропилин-2 и EPHB4 ; Считается, что это помогает направлять кровоток из артериально-венозных участков петли. [2] Однако механические сигналы, обеспечиваемые первыми сокращениями сердца, по-прежнему необходимы для полного ремоделирования. [2]
Первое событие биомеханического иерархического ремоделирования происходит сразу после начала сердечного сокращения, когда желточная артерия формируется путем слияния нескольких более мелких капилляров. Впоследствии боковые ветви могут отсоединяться от основной артерии и снова присоединяться к венозной сети, эффективно меняя свою идентичность. [10] Это мысль [ кем? ] Это происходит из-за высокого просветного давления в артериальных линиях, что предотвращает повторное прикрепление ветвей обратно к артериальным сосудам. [10] Это также предотвращает образование шунтов между двумя компонентами сети. [5] Мойон и др. показали, что артериальные эндотелиальные клетки могут стать венозными и наоборот. [11] Они пересадили участки эндотелиальных трубок перепела , в которых ранее были экспрессированы артериальные маркеры, на вены кур (или наоборот), продемонстрировав пластичность системы. Изменение структуры кровотока в артериях и/или венах также может иметь тот же эффект, хотя неясно, связано ли это с различиями в физических или химических свойствах венозного и артериального кровотока (т.е. профиля давления и напряжения кислорода ). [10]
Другим примером изменчивости артериально-венозной идентичности является интерсомитный сосуд. На ранних стадиях этот сосуд соединяется с аортой , делая ее частью артериальной сети. [2] Однако отростки сердечной вены могут сливаться с интерсомитным сосудом, который медленно отделяется от аорты и становится веной. [2] Этот процесс до конца не изучен, но может происходить из-за необходимости сбалансировать механические силы, такие как давление и перфузия . [2]
Артериально-венозная идентичность на ранних стадиях эмбрионального ремоделирования сосудов является гибкой: артериальные сегменты часто рециркулируются в венозные линии, а физическая структура и генетические маркеры сегментов активно ремоделируются вместе с самой сетью. [10] Это указывает на то, что система в целом демонстрирует степень пластичности, которая позволяет ей формироваться с помощью преходящих паттернов кровотока и гемодинамических сигналов, однако генетические факторы действительно играют роль в первоначальной спецификации идентичности сосудов. [2]
Биомеханика
[ редактировать ]Как только сердце начинает биться, механические силы начинают действовать на раннюю сосудистую систему, которая быстро расширяется и реорганизуется для обеспечения тканевого метаболизма . [9] У эмбрионов, лишенных кровотока, эндотелиальные клетки сохраняют недифференцированную морфологию, сходную с ангиобластами (по сравнению с уплощенными эпителиальными клетками, обнаруженными в зрелой сосудистой сети). [2] Как только сердце начинает биться, морфология и поведение эндотелиальных клеток изменяются. [2] [12] Изменяя частоту сердечных сокращений , сердце также может контролировать перфузию или давление, воздействующее на систему, чтобы вызвать прорастание новых сосудов. [2] В свою очередь, прорастание новых сосудов уравновешивается расширением других тканей эмбриона, которые сжимают кровеносные сосуды по мере их роста. [5] Равновесие этих сил играет важную роль в ремоделировании сосудов, но хотя ангиогенные механизмы, необходимые для запуска прорастания новых сосудов, изучены, мало что известно о процессах ремоделирования, необходимых для сдерживания роста ненужных ветвей. [2]

Когда кровь перфузирует систему, она оказывает на стенки сосудов силы сдвига и давления. В то же время рост тканей за пределами сердечно-сосудистой системы отталкивает наружные стенки сосудов. Эти силы должны быть сбалансированы для получения эффективного энергетического состояния для недорогой доставки питательных веществ и кислорода ко всем тканям тела эмбриона. [2] Когда рост желточного мешка (внешней ткани) ограничен, баланс между сосудистыми и тканевыми силами смещается, и некоторые сосудистые ветви могут отсоединяться или уменьшаться во время процесса ремоделирования, поскольку они неспособны прокладывать новые пути через сжатую ткань. [2] В общем, жесткость и сопротивление этих тканей определяют степень их деформации и способ воздействия на них биомеханических сил. [2]
Развитие сосудистой сети самоорганизуется в каждой точке ткани за счет баланса между сжимающими силами расширения ткани и окружным растяжением стенок сосудов. [5] Со временем это означает, что мигрирующие линии становятся прямыми, а не изогнутыми; это похоже на представление двух движущихся границ, давящих друг на друга. [5] Прямые сосуды обычно параллельны линиям изодавления, поскольку их границы уравновешивают градиенты давления . [5] Кроме того, направление сосуда имеет тенденцию следовать направлению нормали к самому крутому градиенту напряжения. [5]
Кроме того, биомеханические силы внутри эмбриональных сосудов оказывают важное ремоделирующее воздействие. Колебания давления приводят к колебаниям напряжения и напряжения, которые могут «приучить» сосуды выдерживать нагрузки на более поздних этапах развития организма. [9] Слияние нескольких мелких сосудов также может привести к образованию крупных сосудов в тех участках сосудистого дерева, где артериальное давление и скорость кровотока выше. [10] Закон Мюррея представляет собой соотношение между радиусом материнских сосудов и радиусом ветвей, справедливое для системы кровообращения. Это определяет баланс между наименьшим сопротивлением потоку, обусловленным размером сосудов (поскольку в сосудах большого диаметра наблюдается низкий перепад давления ) и сохранением самой крови как живой ткани, которая не может диффундировать до бесконечности . [2] Следовательно, для снабжения кровью систем органов требуется сложное разветвление, поскольку одна только диффузия не может быть ответственна за это. [ по мнению кого? ] [ оригинальное исследование? ]
Биомеханика воздействует также на соединения сосудистой сети. Было показано, что люминальное давление направляет переработку сегментов сосудов в области высокого давления. [5] и управляют отсоединением сегментов сосудов от артериальных линий и повторным присоединением к венозным линиям для формирования сети. [7] Этот тип разрыва сосудов может даже быть косвенно ответственным за развитие некоторых систем органов и эволюцию более крупных организмов, поскольку без отслоения и миграции большие массы тканей эмбриона оставались бы отключенными от кровоснабжения. [5] Как только сосуды отрываются от материнской артерии, они также могут подвергаться ангиогенезу и инфицировать ткани, дистальные по отношению к остальной части сети. [2]
Гидродинамика
[ редактировать ]
Динамика жидкости также играет важную роль в ремоделировании сосудов. Напряжение сдвига, приложенное к стенкам сосуда, пропорционально вязкости и характеру течения жидкости. Нарушенный характер потока может способствовать образованию клапанов, а повышение давления может повлиять на радиальный рост сосудов. [9] Примитивное сердце в течение первых нескольких дней сокращения лучше всего описать как перистальтический насос , однако через три дня кровоток становится пульсирующим. [9] Пульсирующий поток играет важную роль в ремоделировании сосудов, поскольку характер потока может влиять на механотрансдукцию стресса в эндотелиальные клетки. [7] [13]
Безразмерные соотношения, такие как число Рейнольдса и число Уомерсли, можно использовать для описания кровотока в ранней сосудистой сети. [7] Низкое число Рейнольдса, присутствующее во всех ранних сосудах, означает, что поток можно считать ползучим и ламинарным . [7] Низкое число Уомерсли означает, что вязкие эффекты доминируют в структуре потока и что пограничные слои можно считать несуществующими. [7] Это позволяет гидродинамическим расчетам основываться на определенных предположениях , которые упрощают математику . [ оригинальное исследование? ]
На первых стадиях эмбрионального ремоделирования сосудов высокоскоростной кровоток присутствует не только в сосудах большого диаметра, но это корректируется само собой благодаря эффектам ремоделирования сосудов в течение первых двух дней кровотока. [14] Известно [ кем? ] что эмбриональные сосуды реагируют на повышение давления увеличением диаметра сосудов. [9] Из-за отсутствия гладкомышечных клеток и гликокаликса , обеспечивающих эластичную поддержку сосудов взрослого человека, кровеносные сосуды развивающегося эмбриона гораздо более устойчивы к кровотоку. [7] Это означает, что на увеличение потока или давления можно ответить только быстрым, полупостоянным увеличением диаметра сосуда, а не более постепенным растяжением и расширением, наблюдаемым у взрослых кровеносных сосудов. [7]
Перестановка соотношений Лапласа и Пуазейля предполагает, что радиальный рост происходит в результате окружного растяжения, а окружной рост происходит в результате напряжения сдвига. [9] Напряжение сдвига пропорционально скорости внутри сосуда, а также перепаду давления между двумя фиксированными точками на стенке сосуда. [5] Считается, что точный механизм ремоделирования сосудов заключается в высоком напряжении на внутренней стенке сосуда, которое может вызвать рост, ведущий к равномерному сжимающему и растягивающему напряжению по обе стороны стенки сосуда. [9] В общем, обнаружено [ кем? ] что окружное остаточное напряжение является сжимающим и растягивающим, что указывает на то, что внутренние слои эндотелиальной трубки растут больше, чем наружные слои. [15]
Механотрансдукция и генетическая регуляция
[ редактировать ]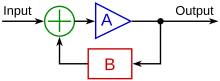
Механизм, с помощью которого разные типы паттернов кровотока и другие физические сигналы по-разному влияют на ремоделирование сосудов у эмбриона, называется механотрансдукцией. Турбулентный поток , который является обычным явлением в развивающейся сосудистой сети, играет роль в формировании сердечных клапанов, которые предотвращают обратный поток, связанный с турбулентностью. [16] Также было показано, что гетерогенные структуры кровотока в крупных сосудах могут создавать асимметрию , возможно, за счет преимущественной активации генов, таких как PITX2, на одной стороне сосуда или, возможно, путем индукции растяжения по окружности с одной стороны, способствуя регрессии с другой стороны. [6] [17] Ламинарный поток также имеет генетические эффекты, такие как снижение апоптоза , ингибирование пролиферации , выравнивание клеток в направлении потока и регулирование многих клеточных сигнальных факторов. [7] Механотрансдукция может действовать посредством петель положительной или отрицательной обратной связи , которые могут активировать или подавлять определенные гены в ответ на физический стресс или нагрузку, оказываемую на сосуд.
Клетка «считывает» структуру потока через интегрина , рецепторы которые обеспечивают механическую связь между внеклеточным матриксом и актиновым цитоскелетом . Этот механизм определяет, как клетка будет реагировать на структуру потока, и может опосредовать клеточную адгезию , что особенно важно для прорастания новых сосудов. [2] В процессе механотрансдукции стресс сдвига может регулировать экспрессию множества различных генов. Следующие примеры были изучены в контексте ремоделирования сосудов с помощью биомеханики:
- Эндотелиальная синтаза оксида азота (eNOS) способствует однонаправленному потоку крови в начале сердечных сокращений и активируется напряжением сдвига. [18]
- Тромбоцитарный фактор роста (PDGF), трансформирующий фактор роста бета (TGFβ) и Круппель-подобный фактор 2 (Klf-2) индуцируются сдвиговым стрессом и могут оказывать повышающее регулирующее воздействие на гены, отвечающие за реакцию эндотелия на турбулентный поток. [7]
- Сдвиговое напряжение вызывает фосфорилирование рецепторов VEGF, которые отвечают за развитие сосудов, особенно за прорастание новых сосудов. [2] [7]
- Гипоксия может вызвать экспрессию фактора 1, индуцируемого гипоксией (HIF-1) или VEGF, чтобы инициировать рост новых ростков в областях эмбриона, лишенных кислорода. [2]
- PDGF-β, VEGFR-2 и connexion43 активируются за счет аномальных паттернов кровотока. [2]
- Напряжение сдвига активирует NF-κB , что индуцирует матриксные металлопротеиназы , вызывающие расширение кровеносных сосудов. [19]
Различные модели потока и их продолжительность могут вызывать очень разные реакции, основанные на генах, регулируемых сдвиговым стрессом. [7] За процесс ремоделирования сосудов эмбриона ответственны как генетическая регуляция, так и физические силы, однако эти факторы редко изучаются совместно. [2] [7]
in vivo Исследование
[ редактировать ]Основная трудность в изучении эмбрионального ремоделирования сосудов in vivo заключалась в том, чтобы отделить эффекты физических сигналов от доставки питательных веществ, кислорода и других сигнальных факторов, которые могут влиять на ремоделирование сосудов. [7] Предыдущая работа включала контроль вязкости крови на ранних стадиях сердечно-сосудистых заболеваний, например, предотвращение проникновения эритроцитов в плазму крови , тем самым снижая вязкость и связанные с ней напряжения сдвига. [18] Крахмал также можно вводить в кровоток для увеличения вязкости и напряжения сдвига. [18] Исследования показали, что ремоделирование сосудов у эмбриона протекает без присутствия эритроцитов , отвечающих за доставку кислорода. [18] Следовательно, ремоделирование сосудов не зависит от присутствия кислорода и фактически происходит до того, как перфузируемые ткани потребуют доставки кислорода. [7] Однако до сих пор неизвестно, могут ли другие питательные вещества или генетические факторы оказывать стимулирующее воздействие на ремоделирование сосудов. [18]
Измерение профилей параболической скорости в сосудах живого эмбриона показывает, что стенки сосудов подвергаются ламинарному и сдвиговому напряжению, которое может иметь биологически активный эффект. [14] Сдвиговое напряжение на сосудистую сеть эмбрионов мышей и кур колеблется в пределах 1–5 дин/см2. [14] Это можно измерить, разрезав срезы кровеносных сосудов и наблюдая за углом отверстия, которое изгибается для снятия остаточного напряжения . [15] или путем измерения гематокрита , присутствующего в кровеносных сосудах, и расчета кажущейся вязкости жидкости. [7]
Из-за трудностей, связанных с визуализацией развития живых эмбрионов и точным измерением небольших значений вязкости, давления, скорости и направления потока, повышенное значение придается разработке точной модели этого процесса. Таким образом, может быть найден эффективный метод изучения этих эффектов in vitro . [ по мнению кого? ]
Моделирование
[ редактировать ]Был предложен ряд моделей для описания влияния жидкости на ремоделирование сосудов у эмбриона. Один момент, который часто упускают из виду [ по мнению кого? ] в этих аналогиях заключается тот факт, что процесс происходит внутри живой системы; Тупиковый конец может отрываться и вновь прикрепляться в другом месте, ветви закрываются и открываются в местах соединения или образуют клапаны, а сосуды чрезвычайно деформируются, способны быстро приспосабливаться к новым условиям и образовывать новые пути. Теоретически формирование сосудистого дерева можно рассматривать с точки зрения теории перколяции . Сеть трубок возникает случайным образом и в конечном итоге образует путь между двумя отдельными и несоединенными точками. Как только некоторое критическое количество отрастающих трубок мигрирует в ранее незанятую область, между этими двумя точками может быть установлен путь, называемый фракталом . [8] Фракталы являются биологически полезными конструкциями, поскольку они основаны на бесконечном увеличении площади поверхности , что с биологической точки зрения приводит к значительному увеличению эффективности транспортировки питательных веществ и отходов. [8] Фрактальный путь гибок; если одно соединение разрывается, создается другое для восстановления пути. [8] Это полезная иллюстрация того, как формируется сосудистое дерево, хотя ее нельзя использовать в качестве модели.Модель агрегации, ограниченной диффузией, дала смоделированные результаты, которые наиболее близки по сравнению с сосудистыми деревьями in vivo . Эта модель предполагает, что рост сосудов происходит по градиенту напряжения сдвига на стенке сосуда, что приводит к увеличению радиусов сосудов. [20] Агрегация, ограниченная диффузией, предполагает, что агрегат растет за счет слияния случайных блуждающих объектов, которые сами движутся по градиенту давления. [5] Случайное блуждание — это просто вероятностная версия уравнения диффузии. [5] Таким образом, при применении этой модели к сосудистому дереву мелкие резистентные сосуды должны быть заменены крупными проводящими сосудами, чтобы сбалансировать давление во всей системе. [5] Эта модель дает структуру, которая более случайна на кончиках, чем на основных линиях, что связано с тем фактом, что формулы Лапласа устойчивы, когда скорость отрицательна по отношению к градиенту давления. [5] В крупных линиях это всегда так, но в маленьких ростках скорость колеблется около 0, что приводит к нестабильному, случайному поведению. [5]
Еще одним важным компонентом процесса ремоделирования является отсоединение разветвленных сосудов, которые затем мигрируют в дистальные области, чтобы гомогенно снабжать кровью. [5] Было обнаружено, что морфогенез ветвления следует модели диэлектрического разрушения , при которой только сосуды с достаточным кровотоком будут расширяться, а другие закрываются. [5] В тех местах внутри сосуда, где две трубки отделяются от одной, одно плечо разделения, скорее всего, закроется, отсоединится и переместится в сторону венозной линии, где оно снова прикрепится. Результатом закрытия ответвления является то, что поток увеличивается и становится менее турбулентным в основной линии, при этом кровь также начинает течь к участкам, которых не хватает. [5] Какая ветвь закроется, зависит от скорости потока, направления и угла разветвления; как правило, угол разветвления 75° или более приводит к необходимости закрытия меньшего ответвления. [5]
Таким образом, с помощью комбинированных моделей диффузионно-ограниченной агрегации и диэлектрического распада можно описать несколько важных параметров ремоделирования сосудов: вероятность закрытия ветки (пластичность расщепления сосуда), повторного соединения сосуда с венозной линией (пластичность возобновления роста проростков), сопротивление усадке кончиков прорастающих клеток (баланс между внешним сжатием и внутренним напряжением сдвига) и соотношение внешнего роста тканей и внутреннего расширения сосудов. Однако эта модель не учитывает диффузию кислорода или сигнальные факторы, которые могут играть роль в ремоделировании эмбриональных сосудов. [5] Эти модели последовательно воспроизводят большинство аспектов сосудистой сети, наблюдаемых in vivo в нескольких различных специализированных случаях. [5]
Применение для изучения прогрессирования заболевания
[ редактировать ]
Ремоделирование сосудов в неэмбриональных тканях считается симптомом прогрессирования заболевания. Сердечно-сосудистые заболевания остаются одной из наиболее частых причин смерти во всем мире [22] и часто связано с закупоркой или стенозом кровеносных сосудов, что может иметь драматические биомеханические последствия. При остром и хроническом ремоделировании увеличение напряжения сдвига из-за уменьшения диаметра заблокированного сосуда может вызвать вазодилатацию, тем самым восстанавливая типичные уровни напряжения сдвига. [6] [23] Однако расширение также приводит к увеличению кровотока через сосуд, что может привести к гиперемии , повлиять на физиологические регуляторные функции ниже пораженного сосуда и оказать повышенное давление на атеросклеротические бляшки, что может привести к их разрыву. [6] Закупорку кровеносных сосудов в настоящее время лечат путем хирургической установки стентов , чтобы заставить сосуды открыться и восстановить нормальный кровоток. Понимая влияние повышенного напряжения сдвига на гомеостатические регуляторы, можно разработать альтернативные, менее инвазивные методы лечения закупорки сосудов.
Рост опухолей часто приводит к реактивации роста кровеносных сосудов и ремоделированию сосудов с целью перфузии новой ткани кровью и поддержания ее пролиферации. [2] Было показано, что рост опухоли является самоорганизующимся и ведет себя более похоже на эмбриональные ткани, чем на ткани взрослого человека. [24] Также рассматривается рост сосудов и динамика кровотока в опухолях. [ кем? ] для повторения роста сосудов у развивающихся эмбрионов. [2] В этом смысле ремоделирование эмбриональных сосудов можно рассматривать как модель тех же путей, которые активируются при росте опухоли, и более глубокое понимание этих путей может привести к разработке новых терапевтических средств, которые могут ингибировать образование опухолей. [ оригинальное исследование? ]
И наоборот, ангиогенез и ремоделирование сосудов являются важным аспектом заживления ран и долгосрочной стабильности тканевых трансплантатов. [2] Когда кровоток нарушается, ангиогенез обеспечивает прорастание сосудов, которые мигрируют в лишенные ткани и восстанавливают перфузию. Таким образом, изучение ремоделирования сосудов может также дать важное представление о разработке новых методов улучшения заживления ран и интеграции тканей трансплантатов за счет снижения частоты отторжения. [ по мнению кого? ]
Ссылки
[ редактировать ]- ^ Уоллс, Джонатон Р.; Култас, Ли; Россант, Джанет; Хенкельман, Р. Марк (6 августа 2008 г.). «Трехмерный анализ развития сосудов в мышином эмбрионе» . ПЛОС ОДИН . 3 (8): е2853. Бибкод : 2008PLoSO...3.2853W . дои : 10.1371/journal.pone.0002853 . ISSN 1932-6203 . ПМЦ 2478714 . ПМИД 18682734 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа Джонс, Элизабет А.В.; ле Нобль, Фердинанд; Эйхманн, Энн (2006). «Что определяет структуру кровеносных сосудов? Генетическая предварительная спецификация против гемодинамики». Физиология . 21 (6): 388–395. дои : 10.1152/физиол.00020.2006 . ISSN 1548-9213 . ПМИД 17119151 .
- ^ Тома, Р. (1893). Исследования гистогенеза и гистомеханики сосудистой системы 1186 г. Штутгарт, Германия: Фердинанд Энке
- ^ Jump up to: а б с Флери, В. (2000). Ветвящийся морфогенез в модели реакции-диффузии. Физический обзор E 61: 4156 – 4160. ПМИД 11088210
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В Нгуен, Ти-Хань; Эйхман, Энн; Ле Нобль, Фердинанд; Флери, Винсент (14 июня 2006 г.). «Динамика морфогенеза ветвления сосудов: влияние кровотока и тканевого кровотока» (PDF) . Физический обзор E . 73 (6): 061907. Бибкод : 2006PhRvE..73f1907N . дои : 10.1103/PhysRevE.73.061907 . ISSN 1539-3755 . ПМИД 16906864 .
- ^ Jump up to: а б с д и Коллер, Акос; Кейли, Габор (1996). «Регуляция сосудистой резистентности в здоровье и заболеваниях, зависимая от напряжения сдвига: роль эндотелия». Эндотелий . 4 (4): 247–272. дои : 10.3109/10623329609024701 . ISSN 1062-3329 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Джонс, Элизабет А.В. (2010). «Механотрансдукция и динамика жидкости в крови в развивающихся кровеносных сосудах» . Канадский журнал химической инженерии . 88 (2): 136–143. doi : 10.1002/cjce.20290 . ISSN 0008-4034 .
- ^ Jump up to: а б с д и ж г Форгакс Г. и Ньюман С.А. (2005). Биологическая физика развивающегося эмбриона. Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0-521-78337-8
- ^ Jump up to: а б с д и ж г час я Табер, Ларри А. (2001). «Биомеханика сердечно-сосудистого развития». Ежегодный обзор биомедицинской инженерии . 3 (1): 1–25. doi : 10.1146/annurev.bioeng.3.1.1 . ISSN 1523-9829 . ПМИД 11447055 .
- ^ Jump up to: а б с д и ле Нобль, Фердинанд; Мойон, Дельфина; Пардано, Люк; Юань, Ли; Джонов, Валентин; Маттейсен, Роберт; Бреант, Кристиана; Флери, Винсент; Эйхманн, Анна (15 января 2004 г.). «Поток регулирует артериально-венозную дифференцировку в желточном мешке эмбриона курицы». Разработка . 131 (2): 361–375. дои : 10.1242/dev.00929 . ISSN 1477-9129 . ПМИД 14681188 .
- ^ Мойон, Д. и др . (сентябрь 2001 г.). Пластичность эндотелиальных клеток при артериально-венозной дифференцировке эмбриона птиц. Развитие 128: 3359 – 3370. PMID 11546752
- ^ Вакимото, Кодзи; и др. (2000). «Направленное разрушение гена-обменника Na+/Ca2+ приводит к апоптозу кардиомиоцитов и нарушениям сердечного ритма» . Журнал биологической химии . 275 (47): 36991–36998. дои : 10.1074/jbc.M004035200 . ПМИД 10967099 .
- ^ Бушманн, И. и др . (апрель 2010 г.). Пульсирующий сдвиг и Gja5 модулируют идентичность артерий и события ремоделирования во время артериогенеза, управляемого потоком. Развитие 137: 2187–2196. ПМИД 20530546
- ^ Jump up to: а б с Джонс, EAV; Барон, Миннесота; Фрейзер, SE; Дикинсон, Мэн (2004). «Измерение гемодинамических изменений во время развития млекопитающих» . Американский журнал физиологии. Физиология сердца и кровообращения . 287 (4): H1561–H1569. дои : 10.1152/ajpheart.00081.2004 . ISSN 0363-6135 . ПМИД 15155254 .
- ^ Jump up to: а б Чуонг, СиДжей и Фунг, ЮК (1986). Об остаточном напряжении в артериях. Журнал биомеханики 108: 189 – 192. ПМИД 3079517
- ^ Хоув, Джей Р.; Кестер, Рейнхард В.; Форухар, Ариан С.; Асеведо-Болтон, Габриэль; Фрейзер, Скотт Э.; Гариб, Мортеза (2003). «Силы внутрисердечной жидкости являются важным эпигенетическим фактором эмбрионального кардиогенеза». Природа . 421 (6919): 172–177. Бибкод : 2003Natur.421..172H . дои : 10.1038/nature01282 . ISSN 0028-0836 . ПМИД 12520305 .
- ^ Яширо, Кента; Ширатори, Хидетака; Хамада, Хироши (2007). «Гемодинамика, определяемая генетической программой, управляет асимметричным развитием дуги аорты». Природа . 450 (7167): 285–288. Бибкод : 2007Natur.450..285Y . дои : 10.1038/nature06254 . ISSN 0028-0836 . ПМИД 17994097 .
- ^ Jump up to: а б с д и Лючитти, Дженнифер Л.; и др. (15 сентября 2007 г.). «Сосудистое ремоделирование желточного мешка мыши требует гемодинамической силы» . Разработка . 134 (18): 3317–3326. дои : 10.1242/dev.02883 . ISSN 1477-9129 . ПМК 4260474 . ПМИД 17720695 .
- ^ Кастье, Ив; Рамкелавон, Бхама; Риу, Стефани; Тедги, Ален; Леу, Стефани (2009). «Роль NF-κB в ремоделировании сосудов, индуцированном потоком». Антиоксиданты и окислительно-восстановительная сигнализация . 11 (7): 1641–1649. дои : 10.1089/ars.2008.2393 . ISSN 1523-0864 . ПМИД 19320561 .
- ^ Флери, Винсент; Шварц, Лоран (1999). «Агрегация, ограниченная диффузией из-за напряжения сдвига, как простая модель васкулогенеза». Фракталы . 07 (1): 33–39. дои : 10.1142/S0218348X99000050 . ISSN 0218-348X .
- ^ Ясуока, Хиронао; Кодама, Риеко; Цудзимото, Масахико; Ёсидомэ, Кацухидэ; Акамацу, Хироки; Накахара, Масааки; Инагаки, Мичиган; Санке, Токио; Накамура, Ясуши (2009). «Экспрессия нейропилина-2 при раке молочной железы: корреляция с метастазами в лимфатические узлы, плохой прогноз и регуляция экспрессии CXCR4» . БМК Рак . 9 (1): 220. дои : 10.1186/1471-2407-9-220 . ISSN 1471-2407 . ПМЦ 2719661 . ПМИД 19580679 .
- ^ Мендис, С. и др . (2011). Глобальный атлас по профилактике сердечно-сосудистых заболеваний и борьбе с ними. Всемирная организация здравоохранения: Женева. ISBN 978-92-4-156437-3
- ^ Кастье, Ив; Брандес, Ральф П.; Леше, Гай; Тедги, Ален; Леу, Стефани (16 сентября 2005 г.). «p47phox-зависимая НАДФН-оксидаза регулирует ремоделирование сосудов, индуцированное потоком». Исследование кровообращения . 97 (6): 533–540. дои : 10.1161/01.RES.0000181759.63239.21 . ISSN 0009-7330 . ПМИД 16109921 .
- ^ Дорманн С. и Дойч А. (2002). Моделирование самоорганизованного аваскулярного роста опухоли с помощью гибридного клеточного автомата. В Silico Biology 2: 393–406. ПМИД 12542422