Гомеостаз
| Часть серии о |
| Биология |
|---|
В биологии гомеостаз , ( британский также гомеостаз ; / h ɒ m i oʊ ˈ s t eɪ s ɪ s условий , -m i ə - / ) — это состояние устойчивых внутренних физических и химических поддерживаемых живыми системами . [1] Это состояние оптимального функционирования организма, которое включает в себя множество переменных, таких как температура тела и баланс жидкости , которые удерживаются в определенных заранее установленных пределах (гомеостатическом диапазоне). Другие переменные включают pH внеклеточной жидкости , концентрацию натрия , калия и кальция ионов , а также уровень сахара в крови , и их необходимо регулировать, несмотря на изменения в окружающей среде, диете или уровне активности. Каждая из этих переменных контролируется одним или несколькими регуляторами или гомеостатическими механизмами, которые вместе поддерживают жизнь.
Гомеостаз обеспечивается естественным сопротивлением изменениям, когда они уже находятся в оптимальных условиях. [2] и равновесие поддерживается многими регуляторными механизмами; считается, что это центральная мотивация всех органических действий. Все механизмы гомеостатического контроля имеют по крайней мере три взаимозависимых компонента регулируемой переменной: рецептор, центр управления и эффектор. [3] Рецептор — это чувствительный компонент, который отслеживает и реагирует на изменения в окружающей среде, как внешние, так и внутренние. Рецепторы включают терморецепторы и механорецепторы . Центры управления включают дыхательный центр и ренин-ангиотензиновую систему . Эффектор — это цель, на которую воздействуют, чтобы вернуть изменение в нормальное состояние. На клеточном уровне эффекторы включают ядерные рецепторы , которые вызывают изменения в экспрессии генов посредством повышающей или понижающей регуляции и действуют по механизмам отрицательной обратной связи . Примером этого является контроль желчных кислот в печени . [4]
Некоторые центры, такие как ренин-ангиотензиновая система , контролируют более чем одну переменную. Когда рецептор ощущает стимул, он реагирует, отправляя потенциалы действия в центр управления. Центр управления устанавливает диапазон поддержания — допустимые верхний и нижний пределы — для конкретной переменной, например температуры. Центр управления реагирует на сигнал, определяя соответствующий ответ и отправляя сигналы эффектору , которым может быть одна или несколько мышц, орган или железа . Когда сигнал получен и на него воздействуют, рецептору предоставляется отрицательная обратная связь, которая устраняет необходимость в дальнейшей передаче сигналов. [5]
Каннабиноидный рецептор типа 1 (CB1), расположенный в пресинаптическом нейроне , представляет собой рецептор , который может остановить высвобождение стрессового нейромедиатора в постсинаптический нейрон; он активируется эндоканнабиноидами (ЭК), такими как анандамид ( N -арахидоноилэтаноламид; AEA) и 2-арахидоноилглицерин (2-AG), посредством ретроградного сигнального процесса, в котором эти соединения синтезируются и высвобождаются из постсинаптических нейронов и возвращаются обратно в пресинаптический терминал для связывания с рецептором CB1 для модуляции высвобождения нейротрансмиттера для достижения гомеостаза. [6]
Полиненасыщенные жирные кислоты (ПНЖК) представляют собой липидные производные омега-3 (докозагексаеновая кислота, DHA и эйкозапентаеновая кислота, EPA ) или омега-6 (арахидоновая кислота, ARA ), синтезируются из мембранных фосфолипидов и используются в качестве предшественника эндоканнабиноидов ( ЭК) опосредуют значительные эффекты в тонкой настройке гомеостаза организма. [7]
Этимология
[ редактировать ]Слово гомеостаз ( / ˌ h oʊ m i oʊ ˈ s t eɪ s ɪ s / [8] [9] ) использует сочетание форм гомео- и -стазиса , нео -латыни от греческого : ὅμοιος homoios , «похожий» и στάσις stasis , «стоя на месте», что дает идею «оставаться прежним».
История
[ редактировать ]Понятие регуляции внутренней среды было описано французским физиологом Клодом Бернаром в 1849 году, а слово гомеостаз было введено Уолтером Брэдфордом Кэнноном в 1926 году. [10] [11] В 1932 году Джозеф Баркрофт , британский физиолог, первым сказал, что для более высокой функции мозга требуется наиболее стабильная внутренняя среда. Таким образом, для Баркрофта гомеостаз не только организовывался мозгом — гомеостаз служил мозгу. [12] Гомеостаз — почти исключительно биологический термин, относящийся к концепциям, описанным Бернаром и Кэнноном, относительно постоянства внутренней среды, в которой живут и выживают клетки организма. [10] [11] [13] Термин «кибернетика» применяется к технологическим системам управления , таким как термостаты , которые функционируют как гомеостатические механизмы, но часто определяются гораздо шире, чем биологический термин гомеостаз. [5] [14] [15] [16]
Обзор
[ редактировать ]Метаболические процессы всех организмов могут протекать только в очень специфических физических и химических средах. Условия варьируются в зависимости от каждого организма и от того, происходят ли химические процессы внутри клетки или в интерстициальной жидкости, омывающей клетки. Наиболее известными гомеостатическими механизмами у человека и других млекопитающих являются регуляторы, которые поддерживают постоянный состав внеклеточной жидкости (или «внутренней среды»), особенно в отношении температуры , pH , осмоляльности и концентрации натрия , калия , глюкоза , углекислый газ и кислород . Однако множество других гомеостатических механизмов, охватывающих многие аспекты физиологии человека , контролируют другие объекты в организме. Если уровни переменных выше или ниже необходимых, к ним часто добавляются префиксы гипер- и гипо- соответственно, например, гипертермия и гипотермия или гипертония и гипотония . [ нужна ссылка ]

Если сущность находится под гомеостатическим контролем, это не означает, что ее ценность обязательно абсолютно стабильна в отношении здоровья. Например, внутренняя температура тела регулируется гомеостатическим механизмом с помощью датчиков температуры, расположенных, среди прочего, в гипоталамусе головного мозга . [17] Однако заданное значение регулятора регулярно сбрасывается. [18] Например, внутренняя температура тела у людей меняется в течение дня (т.е. имеет циркадный ритм ), при этом самые низкие температуры наблюдаются ночью, а самые высокие - во второй половине дня. Другие нормальные колебания температуры включают те, которые связаны с менструальным циклом . [19] [20] Во время инфекций заданное значение регулятора температуры сбрасывается, что приводит к повышению температуры. [17] [21] [22] Организмы способны в некоторой степени приспосабливаться к различным условиям, таким как изменения температуры или уровня кислорода на высоте, посредством процесса акклиматизации .
Гомеостаз не управляет всеми видами деятельности в организме. [23] [24] Например, сигнал (будь то через нейроны или гормоны ) от датчика к эффектору неизбежно сильно варьируется, чтобы передать информацию о направлении и величине ошибки, обнаруженной датчиком. [25] [26] [27] Точно так же реакция эффектора должна быть легко регулируемой, чтобы обратить вспять ошибку – на самом деле она должна быть почти пропорциональна (но в противоположном направлении) ошибке, которая угрожает внутренней среде. [15] [16] Например, артериальное давление у млекопитающих гомеостатически контролируется и измеряется рецепторами растяжения в стенках дуги аорты и сонных синусах в начале внутренних сонных артерий . [17] Датчики отправляют сообщения через сенсорные нервы в продолговатый мозг мозга, указывая, снизилось или повысилось кровяное давление и насколько. Затем продолговатый мозг передает сигналы по двигательным или эфферентным нервам, принадлежащим вегетативной нервной системе , к широкому кругу эффекторных органов, активность которых впоследствии изменяется, чтобы обратить вспять ошибку в кровяном давлении. Одним из эффекторных органов является сердце, частота которого увеличивается ( тахикардия ), когда артериальное давление падает, или замедляется ( брадикардия ), когда давление поднимается выше заданного значения. [17] Таким образом, частота сердечных сокращений (для которой в организме нет датчика) не контролируется гомеостатически, а является одним из эффекторных ответов на ошибки артериального давления. Другой пример – скорость потоотделения . Это один из эффекторов гомеостатического контроля температуры тела, и поэтому он сильно варьируется примерно пропорционально тепловой нагрузке, которая угрожает дестабилизировать внутреннюю температуру тела, для чего в гипоталамусе мозга есть датчик. [ нужна ссылка ]
Управление переменными
[ редактировать ]Температура ядра
[ редактировать ]
Млекопитающие регулируют свою внутреннюю температуру, используя сигналы терморецепторов в гипоталамусе , мозге, [17] [28] спинной мозг , внутренние органы и крупные вены. [29] [30] Помимо внутренней регуляции температуры, может вступить в действие процесс, называемый аллостазом , который корректирует поведение, чтобы адаптироваться к экстремально жарким или холодным условиям (и к другим проблемам). [31] Эти корректировки могут включать поиск тени и снижение активности, поиск более теплых условий и увеличение активности или скопление в кучу. [32] Поведенческая терморегуляция имеет приоритет над физиологической терморегуляцией, поскольку необходимые изменения могут произойти быстрее, а способность физиологической терморегуляции реагировать на экстремальные температуры ограничена. [33]
Когда внутренняя температура падает, кровоснабжение кожи снижается из-за интенсивной вазоконстрикции . [17] Приток крови к конечностям (имеющим большую площадь поверхности) аналогичным образом уменьшается и возвращается в туловище через глубокие вены, лежащие рядом с артериями (образующие вены-соединяющиеся вены ). [28] [32] [34] Это действует как противоточная система обмена , которая закорачивает тепло артериальной крови непосредственно в венозную кровь, возвращающуюся в туловище, вызывая минимальные потери тепла от конечностей в холодную погоду. [28] [32] [35] Подкожные вены конечностей туго сужены, [17] не только уменьшая теплопотери от этого источника, но и нагнетая венозную кровь в противоточную систему в глубину конечностей.
Скорость метаболизма увеличивается, первоначально за счет несократительного термогенеза . [36] с последующим дрожательным термогенезом , если предшествующие реакции недостаточны для коррекции гипотермии .
обнаруживают повышение внутренней температуры Когда терморецепторы , потовые железы кожи стимулируются через холинергические симпатические нервы, выделяя пот на кожу, который, испаряясь, охлаждает кожу и кровь, текущую через нее. Альтернативным эффектором у многих позвоночных является дыхание, охлаждающее тело также за счет испарения воды, но на этот раз со слизистых оболочек горла и рта. [37]
Глюкоза крови
[ редактировать ]
сахара в крови Уровень регулируется в достаточно узких пределах. [38] У млекопитающих первичными сенсорами этого являются бета-клетки островков поджелудочной железы . [39] [40] Бета-клетки реагируют на повышение уровня сахара в крови секрецией инсулина в кровь и одновременно подавляют соседними альфа-клетками . секрецию глюкагона в кровь [39] Эта комбинация (высокий уровень инсулина в крови и низкий уровень глюкагона) действует на эффекторные ткани, главными из которых являются печень , жировые клетки и мышечные клетки . Печень не может производить глюкозу , вместо этого поглощая ее и превращая в гликоген и триглицериды . Гликоген хранится в печени, а триглицериды секретируются в кровь в виде частиц липопротеинов очень низкой плотности (ЛПОНП), которые поглощаются жировой тканью и откладываются там в виде жиров. Жировые клетки поглощают глюкозу через специальные транспортеры глюкозы ( GLUT4 ), количество которых в клеточной стенке увеличивается под прямым воздействием инсулина, действующего на эти клетки. Глюкоза, которая таким образом поступает в жировые клетки, превращается в триглицериды (по тем же метаболическим путям, которые используются печенью), а затем сохраняется в этих жировых клетках вместе с триглицеридами, полученными из ЛПОНП, которые вырабатываются в печени. Мышечные клетки также поглощают глюкозу через инсулин-чувствительные каналы глюкозы GLUT4 и преобразуют ее в мышечный гликоген. [41]
Падение уровня глюкозы в крови приводит к остановке секреции инсулина и глюкагона выделению из альфа-клеток в кровь. Это подавляет поглощение глюкозы из крови печенью, жировыми клетками и мышцами. Вместо этого печень сильно стимулируется к производству глюкозы из гликогена (посредством гликогенолиза ) и из неуглеводных источников (таких как лактат и дезаминированные аминокислоты ) с использованием процесса, известного как глюконеогенез . [42] Произведенная таким образом глюкоза выбрасывается в кровь, исправляя обнаруженную ошибку ( гипогликемию ). Гликоген, хранящийся в мышцах, остается в мышцах и расщепляется только во время тренировки до глюкозо-6-фосфата , а затем до пирувата, который поступает в цикл лимонной кислоты или превращается в лактат . В кровь возвращаются только лактат и отходы цикла лимонной кислоты. Печень может поглощать только лактат и в процессе энергозатратного глюконеогенеза превращать его обратно в глюкозу. [ нужна ссылка ]
Уровни железа
[ редактировать ]Контроль уровня железа в организме является критически важной частью многих аспектов здоровья и болезней человека. Для человека железо одновременно необходимо организму и потенциально вредно. [43]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2023 г. ) |
Регулирование меди
[ редактировать ]Медь всасывается, транспортируется, распределяется, хранится и выводится из организма в соответствии со сложными гомеостатическими процессами, которые обеспечивают постоянное и достаточное поступление микроэлемента, одновременно избегая его избыточного уровня. Если в течение короткого периода времени поступает недостаточное количество меди, запасы меди в печени истощаются. Если это истощение будет продолжаться, может развиться состояние дефицита меди. Если в организм попадает слишком много меди, это может привести к избытку меди. Оба этих состояния, дефицит и избыток, могут привести к повреждению тканей и заболеваниям. Однако благодаря гомеостатической регуляции организм человека способен сбалансировать широкий диапазон потребления меди для нужд здоровых людей. [44]
Многие аспекты гомеостаза меди известны на молекулярном уровне. Существенность меди обусловлена ее способностью действовать как донор или акцептор электронов, поскольку потоки ее степени окисления между Cu 1+ ( медь ) и Cu 2+ ( медный ). В качестве компонента около дюжины купроферментов медь участвует в ключевых окислительно-восстановительных (т.е. окислительно-восстановительных) реакциях в важнейших метаболических процессах, таких как митохондриальное дыхание, синтез меланина и сшивание коллагена . Медь является неотъемлемой частью антиоксидантного фермента супероксиддисмутазы меди-цинка и играет роль в гомеостазе железа в качестве кофактора церулоплазмина.
Уровни газов крови
[ редактировать ]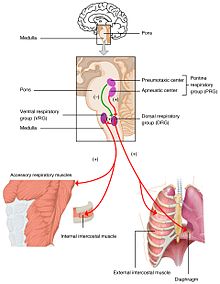
Изменения уровней кислорода, углекислого газа и pH плазмы передаются в дыхательный центр , в ствол мозга , где они регулируются.Парциальное давление кислорода углекислого и газа в артериальной крови контролируется периферическими хеморецепторами ( ПНС ) в сонной артерии и дуге аорты . Изменение парциального давления углекислого газа обнаруживается по изменению pH спинномозговой жидкости центральными хеморецепторами ( ЦНС ) в продолговатом мозге мозга ствола . Информация от этих наборов датчиков отправляется в дыхательный центр, который активирует эффекторные органы – диафрагму и другие дыхательные мышцы . Повышенный уровень углекислого газа в крови или пониженный уровень кислорода приведет к более глубокому дыханию и увеличению частоты дыхания , чтобы вернуть газы крови в равновесие.
Слишком мало углекислого газа и, в меньшей степени, слишком много кислорода в крови могут временно остановить дыхание — состояние, известное как апноэ , которое фридайверы используют, чтобы продлить время пребывания под водой.
Парциальное давление углекислого газа является скорее решающим фактором при мониторинге pH. [45] Однако на большой высоте (выше 2500 м) приоритет имеет контроль парциального давления кислорода, и гипервентиляция поддерживает постоянный уровень кислорода. При более низком уровне углекислого газа, чтобы поддерживать pH на уровне 7,4, почки выделяют ионы водорода в кровь и выделяют бикарбонат в мочу. [46] [47] Это важно для акклиматизации на большой высоте . [48]
Содержание кислорода в крови
[ редактировать ]Почки измеряют содержание кислорода, а не парциальное давление кислорода в артериальной крови. Когда содержание кислорода в крови хронически низкое, чувствительные к кислороду клетки секретируют эритропоэтин (ЭПО). в кровь [49] Эффекторной тканью является красный костный мозг , который производит эритроциты (эритроциты, также называемые эритроцитами ). Увеличение количества эритроцитов приводит к увеличению гематокрита в крови и последующему увеличению гемоглобина , что увеличивает способность переносить кислород. Это механизм, благодаря которому жители высокогорья имеют более высокий гематокрит, чем жители уровня моря, а также почему люди с легочной недостаточностью или шунтами справа налево в сердце (посредством которых венозная кровь минует легкие и поступает непосредственно в системный кровоток). кровообращение) имеют столь же высокий гематокрит. [50] [51]
Независимо от парциального давления кислорода в крови, количество кислорода, которое может быть перенесено, зависит от содержания гемоглобина. Парциального давления кислорода может быть достаточно, например, при анемии , но содержание гемоглобина будет недостаточным, а следовательно, и содержание кислорода. При достаточном поступлении железа, витамина B12 и фолиевой кислоты ЭПО может стимулировать выработку эритроцитов, а содержание гемоглобина и кислорода восстанавливается до нормального уровня. [50] [52]
Артериальное давление
[ редактировать ]Мозг может регулировать кровоток в диапазоне значений артериального давления путем сужения и расширения сосудов артерий. [53]
Рецепторы высокого давления, называемые барорецепторами, в стенках дуги аорты и каротидного синуса (в начале внутренней сонной артерии ) контролируют артериальное давление . [54] Повышение давления обнаруживается, когда стенки артерий растягиваются из-за увеличения объема крови . Это заставляет клетки сердечной мышцы секретировать гормон предсердный натрийуретический пептид в кровь (ANP). Это действует на почки, подавляя секрецию ренина и альдостерона, вызывая выброс натрия и сопутствующей воды в мочу, тем самым уменьшая объем крови. [55] Затем эта информация передается через афферентные нервные волокна к одиночному ядру продолговатого мозга . [56] Отсюда стимулируются двигательные нервы, относящиеся к вегетативной нервной системе, которые влияют на деятельность главным образом сердца и артерий наименьшего диаметра, называемых артериолами . Артериолы являются основными сосудами сопротивления артериального дерева , и небольшие изменения в диаметре вызывают большие изменения сопротивления потоку через них. Когда артериальное кровяное давление повышается, артериолы стимулируются к расширению, что облегчает выход крови из артерий, тем самым сдувая их и возвращая кровяное давление обратно к нормальному. В то же время сердце стимулируется через холинергические парасимпатические нервы , чтобы оно сокращалось медленнее (так называемая брадикардия ), обеспечивая уменьшение притока крови в артерии, тем самым усиливая снижение давления и исправляя исходную ошибку.
Низкое давление в артериях вызывает противоположный рефлекс сужения артериол и ускорение сердечного ритма ( тахикардия ). Если падение артериального давления очень быстрое или чрезмерное, продолговатый мозг стимулирует мозговое вещество надпочечников через «преганглионарные» симпатические нервы выделять адреналин в кровь (адреналин). Этот гормон усиливает тахикардию и вызывает тяжелую вазоконстрикцию артериол всех органов, кроме основного органа (особенно сердца, легких и мозга). Эти реакции обычно очень эффективно корректируют низкое артериальное давление ( гипотонию ).
Уровень кальция
[ редактировать ]
Ионизированный кальций в плазме (Ca 2+ ) концентрация очень жестко контролируется парой гомеостатических механизмов. [57] Сенсор первого расположен в паращитовидных железах , где главные клетки воспринимают Ca 2+ уровне посредством специализированных рецепторов кальция в их мембранах. Сенсорами второго являются парафолликулярные клетки щитовидной железы . Главные клетки паращитовидной железы секретируют паратиреоидный гормон (ПТГ) в ответ на падение уровня ионизированного кальция в плазме; парафолликулярные клетки щитовидной железы секретируют кальцитонин в ответ на повышение уровня ионизированного кальция в плазме.
Эффекторными двенадцатиперстная органами первого гомеостатического механизма являются кости , почки и, через гормон, выделяемый почками в кровь в ответ на высокие уровни ПТГ в крови, и тощая кишка . Паратиреоидный гормон (в высоких концентрациях в крови) вызывает резорбцию кости , высвобождая кальций в плазму. Это очень быстрое действие, которое может исправить угрожающую гипокальциемию в течение нескольких минут. Высокие концентрации ПТГ вызывают выведение ионов фосфата с мочой. Поскольку фосфаты соединяются с ионами кальция с образованием нерастворимых солей (см. также костные минералы ), снижение уровня фосфатов в крови приводит к высвобождению свободных ионов кальция в пул ионизированного кальция плазмы. ПТГ оказывает второе действие на почки. Он стимулирует выработку и выброс почками кальцитриола в кровь. Этот стероидный гормон действует на эпителиальные клетки верхних отделов тонкой кишки, увеличивая их способность поглощать кальций из содержимого кишечника в кровь. [58]
Второй гомеостатический механизм с датчиками в щитовидной железе высвобождает кальцитонин в кровь, когда уровень ионизированного кальция в крови повышается. Этот гормон действует в первую очередь на кости, вызывая быстрое удаление кальция из крови и отложение его в нерастворимой форме в костях. [59]
Два гомеостатических механизма, действующие через ПТГ, с одной стороны, и кальцитонин, с другой, могут очень быстро исправить любую надвигающуюся ошибку в уровне ионизированного кальция в плазме, либо удаляя кальций из крови и откладывая его в скелете, либо удаляя из него кальций. . Скелет . действует как чрезвычайно большой депо кальция (около 1 кг) по сравнению с депо кальция в плазме (около 180 мг) Долгосрочная регуляция происходит за счет абсорбции или потери кальция из кишечника.
Другим примером являются наиболее хорошо изученные эндоканнабиноиды , такие как анандамид ( N -арахидоноилэтаноламид; AEA) и 2-арахидоноилглицерин (2-AG), синтез которых происходит под действием ряда внутриклеточных ферментов, активируемых в ответ на повышение внутриклеточного уровня кальция. обеспечить гомеостаз и предотвращение развития опухоли посредством предполагаемых защитных механизмов, которые предотвращают клеток рост и миграцию путем активации CB1 и/или CB2 и прилегающих рецепторов . [60]
Концентрация натрия
[ редактировать ]Гомеостатический механизм, контролирующий концентрацию натрия в плазме, гораздо более сложен, чем большинство других гомеостатических механизмов, описанных на этой странице.
Датчик расположен в юкстагломерулярном аппарате почек, который удивительно косвенно определяет концентрацию натрия в плазме. Вместо того, чтобы измерять его непосредственно в крови, протекающей мимо юкстагломерулярных клеток , эти клетки реагируют на концентрацию натрия в жидкости почечных канальцев после того, как она уже претерпела определенную модификацию в проксимальных извитых канальцах и петле Генле . [61] Эти клетки также реагируют на скорость кровотока через юкстагломерулярный аппарат, которая в нормальных условиях прямо пропорциональна артериальному давлению , что делает эту ткань вспомогательным датчиком артериального давления.
В ответ на снижение концентрации натрия в плазме или падение артериального давления юкстагломерулярные клетки выделяют ренин . в кровь [61] [62] [63] Ренин — это фермент, который отщепляет декапептид (короткую белковую цепь длиной 10 аминокислот) от α-2-глобулина плазмы, называемого ангиотензиногеном . Этот декапептид известен как ангиотензин I. [61] Он не имеет известной биологической активности. Однако, когда кровь циркулирует через легкие, эндотелиальный фермент легочных капилляров, называемый ангиотензинпревращающим ферментом (АПФ), отщепляет еще две аминокислоты от ангиотензина I с образованием октапептида, известного как ангиотензин II . Ангиотензин II — гормон, который действует на кору надпочечников выброс в кровь стероидного гормона альдостерона , вызывая . Ангиотензин II также действует на гладкие мышцы стенок артериол, вызывая сужение этих сосудов малого диаметра, тем самым ограничивая отток крови из артериального дерева, вызывая повышение артериального давления. Таким образом, это усиливает меры, описанные выше (под заголовком «Артериальное кровяное давление»), которые защищают артериальное кровяное давление от изменений, особенно гипотонии .
Стимулированный ангиотензином II альдостерон, высвобождаемый из клубочковой зоны надпочечников, оказывает влияние, в частности, на эпителиальные клетки дистальных извитых канальцев и собирательных трубочек почек. Здесь он вызывает реабсорбцию ионов натрия из почечной канальцевой жидкости в обмен на ионы калия, которые секретируются из плазмы крови в канальцевую жидкость и выходят из организма через мочу. [61] [64] Реабсорбция ионов натрия из жидкости почечных канальцев останавливает дальнейшую потерю ионов натрия из организма и, следовательно, предотвращает ухудшение гипонатриемии . Гипонатриемию можно скорректировать только потреблением соли в рационе. Однако неясно, может ли «солевой голод» быть инициирован гипонатриемией и по какому механизму это может происходить.
Когда концентрация ионов натрия в плазме превышает норму ( гипернатриемия ), высвобождение ренина из юкстагломерулярного аппарата прекращается, прекращается выработка ангиотензина II и, как следствие, высвобождение альдостерона в кровь. Почки реагируют выделением ионов натрия в мочу, тем самым нормализуя концентрацию ионов натрия в плазме. Низкие уровни ангиотензина II в крови снижают артериальное давление как неизбежный сопутствующий ответ.
Реабсорбция ионов натрия из канальцевой жидкости в результате высокого уровня альдостерона в крови сама по себе не приводит к возврату воды почечных канальцев в кровь из дистальных извитых канальцев или собирательных трубочек . Это связано с тем, что натрий реабсорбируется в обмен на калий и, следовательно, вызывает лишь умеренное изменение осмотического градиента между кровью и канальцевой жидкостью. Кроме того, эпителий дистальных извитых канальцев и собирательных трубочек непроницаем для воды в отсутствие антидиуретического гормона (АДГ) в крови. АДГ является частью контроля баланса жидкости . Его уровни в крови варьируются в зависимости от осмоляльности плазмы, которая измеряется в гипоталамусе головного мозга. Действие альдостерона на почечные канальцы предотвращает потерю натрия во внеклеточную жидкость (ECF). Таким образом, осмоляльность ECF не меняется, а значит, и концентрация АДГ в плазме не изменяется. Однако низкие уровни альдостерона вызывают потерю ионов натрия из ECF, что потенциально может вызвать изменение внеклеточной осмоляльности и, следовательно, уровней АДГ в крови.
Концентрация калия
[ редактировать ]Высокие концентрации калия в плазме вызывают деполяризацию мембран клеток клубочковой зоны наружного слоя коры надпочечников . [65] Это вызывает выброс альдостерона в кровь.
Альдостерон действует преимущественно на дистальные извитые канальцы и собирательные трубочки почек, стимулируя выведение ионов калия с мочой. [61] Однако это происходит за счет активации базолатерального Na. + /К + насосы эпителиальных клеток канальцев. Эти натрий/калиевые обменники перекачивают три иона натрия из клетки в интерстициальную жидкость и два иона калия в клетку из интерстициальной жидкости. Это создает градиент концентрации ионов , который приводит к реабсорбции натрия (Na + ) ионы из канальцевой жидкости в кровь и секретирование калия (K + ) ионы из крови в мочу (просвет собирательных трубочек). [66] [67]
Баланс жидкости
[ редактировать ]Общее количество воды в организме должно поддерживаться в равновесии. Баланс жидкости предполагает поддержание стабилизации объема жидкости, а также поддержание стабильного уровня электролитов во внеклеточной жидкости. Баланс жидкости поддерживается процессом осморегуляции и поведением. Осмотическое давление регистрируется осморецепторами в ядре гипоталамуса срединном преоптическом . плазмы, Измерение осмоляльности позволяющее оценить содержание воды в организме, основано на том факте, что потери воды из организма (через неизбежную потерю воды через кожу , которая не является полностью водонепроницаемой и поэтому всегда слегка влажной, водяной пар в выдыхаемый воздух , потливость , рвота , нормальный кал и особенно диарея ) гипотоничны , то есть они менее соленые, чем жидкости организма (сравните, например, вкус слюны со вкусом слез. Последние имеют почти одинаковую соль). содержание в виде внеклеточной жидкости, тогда как первая гипотонична по отношению к плазме. Слюна не имеет соленого вкуса, тогда как слезы явно соленые). Почти все нормальные и аномальные потери Поэтому вода в организме приводит к тому, что внеклеточная жидкость становится гипертонической . И наоборот, чрезмерное потребление жидкости разбавляет внеклеточную жидкость, заставляя гипоталамус регистрировать гипотонической гипонатриемии состояния .
Когда гипоталамус обнаруживает гипертоническую внеклеточную среду, он вызывает секрецию антидиуретического гормона (АДГ), называемого вазопрессином , который действует на эффекторный орган, которым в данном случае является почка . Действие вазопрессина на почечные канальцы заключается в реабсорбции воды из дистальных извитых канальцев и собирательных трубочек , предотвращая тем самым усиление потери воды с мочой. Гипоталамус одновременно стимулирует близлежащий центр жажды, вызывая почти непреодолимое (если гипертонус достаточно сильный) желание выпить воды. Прекращение мочеиспускания предотвращает гиповолемии и гипертонуса ухудшение ; употребление воды исправляет дефект.
Гипоосмоляльность приводит к очень низким уровням АДГ в плазме. Это приводит к ингибированию реабсорбции воды из почечных канальцев, что приводит к выделению больших объемов очень разбавленной мочи и избавлению организма от избыточной воды.
Потеря воды с мочой, когда гомеостат воды в организме не поврежден, представляет собой компенсаторную потерю воды, корректирующую любой избыток воды в организме. Однако, поскольку почки не могут вырабатывать воду, рефлекс жажды является важнейшим вторым эффекторным механизмом гомеостата воды в организме, корректирующим любой дефицит воды в организме.
рН крови
[ редактировать ]
рН плазмы может быть изменен респираторными изменениями парциального давления углекислого газа; или изменены метаболическими изменениями соотношения угольной кислоты и ионов бикарбоната . Бикарбонатная буферная система регулирует соотношение угольной кислоты к бикарбонату равным 1:20, при этом соотношении pH крови составляет 7,4 (как поясняется уравнением Хендерсона-Хассельбаха ). Изменение pH плазмы приводит к кислотно-щелочному дисбалансу .В кислотно-основном гомеостазе существуют два механизма, которые могут помочь регулировать pH. Дыхательная компенсация — механизм дыхательного центра , регулирующий парциальное давление углекислого газа путем изменения частоты и глубины дыхания, чтобы вернуть pH в норму. Парциальное давление углекислого газа также определяет концентрацию углекислоты, при этом в игру также может вступить бикарбонатная буферная система. Почечная компенсация может помочь бикарбонатной буферной системе.Датчик концентрации бикарбоната в плазме достоверно неизвестен. Весьма вероятно, что клетки почечных канальцев дистальных извитых канальцев сами чувствительны к pH плазмы. [ нужна ссылка ] быстро превращается в водород и бикарбонат В результате метаболизма этих клеток образуется углекислый газ, который под действием карбоангидразы . [68] Когда pH ECF падает (становится более кислым), клетки почечных канальцев выделяют ионы водорода в канальцевую жидкость, которые покидают организм через мочу. Ионы бикарбоната одновременно секретируются в кровь, что снижает содержание угольной кислоты и, следовательно, повышает pH плазмы. [68] Обратное происходит, когда рН плазмы повышается выше нормы: ионы бикарбоната выводятся с мочой, а ионы водорода – в плазму.
Когда ионы водорода выделяются в мочу, а бикарбонат - в кровь, последний соединяется с избытком ионов водорода в плазме, стимулирующим почки на выполнение этой операции. Результатом реакции в плазме является образование угольной кислоты, которая находится в равновесии с парциальным давлением диоксида углерода в плазме. Это строго регулируется, чтобы гарантировать отсутствие чрезмерного накопления углекислоты или бикарбоната. Таким образом, общий эффект заключается в том, что ионы водорода теряются с мочой, когда pH плазмы падает. Сопутствующее повышение содержания бикарбоната в плазме поглощает повышенное количество ионов водорода (вызванное падением pH плазмы), и образующийся в результате избыток углекислоты утилизируется в легких в виде углекислого газа. Это восстанавливает нормальное соотношение между бикарбонатом и парциальным давлением углекислого газа и, следовательно, pH плазмы.Обратное происходит, когда высокий pH плазмы стимулирует почки выделять ионы водорода в кровь и выделять бикарбонат в мочу. Ионы водорода соединяются с избытком ионов бикарбоната в плазме, снова образуя избыток углекислоты, которую можно выдыхать в виде углекислого газа в легких, сохраняя концентрацию ионов бикарбоната в плазме, парциальное давление углекислого газа и, следовательно, , pH плазмы постоянный.
Цереброспинальная жидкость
[ редактировать ]Цереброспинальная жидкость (СМЖ) позволяет регулировать распределение веществ между клетками головного мозга, [69] и нейроэндокринные факторы, небольшие изменения которых могут вызвать проблемы или повреждения нервной системы. Например, высокая глицина концентрация нарушает контроль температуры и артериального давления , а высокий pH спинномозговой жидкости вызывает головокружение и обморок . [70]
нейротрансмиссия
[ редактировать ]Тормозные нейроны ЦНС играют гомеостатическую роль в балансе активности нейронов между возбуждением и торможением. Тормозные нейроны, используя ГАМК , производят компенсирующие изменения в нейронных сетях, предотвращая неконтролируемый уровень возбуждения. [71] Дисбаланс между возбуждением и торможением, как полагают, причастен к ряду нервно-психических расстройств . [72]
Нейроэндокринная система
[ редактировать ]Нейроэндокринная система — это механизм, с помощью которого гипоталамус поддерживает гомеостаз, регулируя обмен веществ , размножение, поведение в еде и питье, использование энергии, осмолярность и кровяное давление.
Регуляция обмена веществ осуществляется посредством связей гипоталамуса с другими железами. [73] Три эндокринные железы гипоталамо -гипофизарно-гонадной оси (ось HPG) часто работают вместе и выполняют важные регуляторные функции. Двумя другими регуляторными эндокринными осями являются ось гипоталамус-гипофиз-надпочечники (ось HPA) и ось гипоталамус-гипофиз-щитовидная железа (ось HPT).
Печень также выполняет множество регуляторных функций обмена веществ. Важной функцией является производство и контроль желчных кислот . Слишком большое количество желчной кислоты может быть токсичным для клеток, и ее синтез может ингибироваться за счет активации FXR рецептора ядерного . [4]
Генная регуляция
[ редактировать ]На клеточном уровне гомеостаз осуществляется несколькими механизмами, включая регуляцию транскрипции , которая может изменять активность генов в ответ на изменения.
Энергетический баланс
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( ноябрь 2017 г. ) |
Количество энергии, получаемой с пищей , должно соответствовать количеству потребляемой энергии. Для достижения энергетического гомеостаза аппетит регулируется двумя гормонами: грелином и лептином . Грелин стимулирует голод и прием пищи, а лептин сигнализирует о сытости (сытости).
Обзор мер по изменению веса, включая диету , физические упражнения и переедание, проведенный в 2019 году, показал, что гомеостаз массы тела не может точно исправить «энергетические ошибки», потерю или набор калорий, в краткосрочной перспективе. [74]
Клиническое значение
[ редактировать ]Многие заболевания являются результатом гомеостатического сбоя. Почти любой гомеостатический компонент может работать со сбоями либо в результате наследственного дефекта , врожденной ошибки метаболизма , либо приобретенного заболевания. Некоторые гомеостатические механизмы имеют встроенную избыточность, которая гарантирует, что жизнь не будет подвергаться немедленной угрозе в случае неисправности какого-либо компонента; но иногда гомеостатический сбой может привести к серьезному заболеванию, которое, если не лечить, может привести к летальному исходу. Хорошо известный пример гомеостатической недостаточности показан при сахарном диабете 1 типа . Здесь регуляция уровня сахара в крови не может функционировать, поскольку бета-клетки островков поджелудочной железы разрушаются и не могут производить необходимый инсулин . Уровень сахара в крови повышается в состоянии, известном как гипергликемия . [ нужна ссылка ]
Гомеостат ионизированного кальция в плазме может быть нарушен постоянной, неизменной, избыточной продукцией паратиреоидного гормона аденомой паращитовидной железы, что приводит к типичным признакам гиперпаратиреоза , а именно высокому уровню ионизированного кальция в плазме. 2+ уровней и резорбции кости, что может привести к спонтанным переломам. Аномально высокие концентрации ионизированного кальция в плазме вызывают конформационные изменения во многих белках клеточной поверхности (особенно в ионных каналах и рецепторах гормонов или нейромедиаторов). [75] вызывая вялость, мышечную слабость, анорексию, запоры и лабильные эмоции. [76]
Гомеостат воды в организме может быть нарушен из-за неспособности секретировать АДГ в ответ даже на нормальные ежедневные потери воды через выдыхаемый воздух, фекалии и незаметное потоотделение . При нулевом сигнале АДГ в крови почки производят огромные неизменные объемы очень разбавленной мочи, что приводит к обезвоживанию и смерти, если не лечить.
По мере старения организмов эффективность их систем управления снижается. Неэффективность постепенно приводит к нестабильной внутренней среде, что увеличивает риск заболеваний и приводит к физическим изменениям, связанным со старением. [5]
Различные хронические заболевания удерживаются под контролем с помощью гомеостатической компенсации, которая маскирует проблему, компенсируя ее (восполняя) другим способом. Однако компенсаторные механизмы со временем изнашиваются или нарушаются под действием нового осложняющего фактора (например, появления сопутствующей острой вирусной инфекции), который заставляет организм пройти через новый каскад событий. Такая декомпенсация демаскирует основное заболевание, ухудшая его симптомы. Общие примеры включают декомпенсированную сердечную недостаточность , почечную недостаточность и печеночную недостаточность . [ нужна ссылка ]
Биосфера
[ редактировать ]В Геи гипотезе Джеймс Лавлок [77] заявил, что вся масса живого вещества на Земле (или на любой планете с жизнью) функционирует как огромный гомеостатический суперорганизм , который активно изменяет свою планетарную среду, чтобы создать условия окружающей среды, необходимые для его собственного выживания. С этой точки зрения вся планета поддерживает несколько гомеостазов (основным из которых является температурный гомеостаз). Существует ли такая система на Земле, остается открытым спором. Однако общепринятыми являются некоторые относительно простые гомеостатические механизмы. Например, иногда утверждается, что когда уровень углекислого газа в атмосфере повышается, некоторые растения могут лучше расти и, таким образом, удалять больше углекислого газа из атмосферы. Однако потепление усугубило засухи, сделав воду фактическим ограничивающим фактором на суше. Утверждается, что когда солнечного света много, а температура атмосферы повышается, фитопланктон поверхностных вод океана, действуя как глобальный солнечный свет и, следовательно, датчики тепла, может процветать и производить больше диметилсульфид (ДМС). Молекулы DMS действуют как ядра конденсации облаков , которые производят больше облаков и, таким образом, увеличивают альбедо атмосферы , что приводит к снижению температуры атмосферы. Однако повышение температуры моря привело к расслоению океанов, отделив теплые, освещенные солнцем воды от прохладных, богатых питательными веществами вод. Таким образом, питательные вещества стали ограничивающим фактором, а уровень планктона фактически упал за последние 50 лет, а не повысился. По мере того, как ученые узнают больше о Земле, обнаруживается огромное количество петель положительной и отрицательной обратной связи, которые вместе поддерживают метастабильное состояние, иногда в очень широком диапазоне условий окружающей среды.
Прогнозирующий
[ редактировать ]Прогнозирующий гомеостаз — это упреждающий ответ на ожидаемую угрозу в будущем, например, стимуляцию секреции инсулина гормонами кишечника, которые попадают в кровь в ответ на прием пищи. [39] Эта секреция инсулина происходит до того, как уровень сахара в крови повысится, что снижает уровень сахара в крови в ожидании большого притока в кровь глюкозы в результате переваривания углеводов в кишечнике. [78] Такие упреждающие реакции представляют собой системы с разомкнутым контуром, которые, по сути, основаны на «догадках» и не являются самокорректирующими. [79] Упреждающие реакции всегда требуют замкнутой системы отрицательной обратной связи для исправления «перелетов» и «недолетов», к которым склонны упреждающие системы.
Другие поля
[ редактировать ]Этот термин стал использоваться и в других областях, например:
Риск
[ редактировать ]Актуарий . может ссылаться на гомеостаз риска , когда (например) люди, имеющие антиблокировочную систему тормозов, имеют не лучшие показатели безопасности, чем те, у кого нет антиблокировочной системы тормозов, поскольку первые бессознательно компенсируют более безопасный автомобиль за счет менее безопасных привычек вождения До появления антиблокировочной системы тормозов некоторые маневры включали незначительные заносы, вызывающие страх и избегание. Теперь антиблокировочная система сдвигает границы такой обратной связи, и модели поведения выходят за рамки карательных мер. Было также высказано предположение, что экологические кризисы являются примером гомеостаза риска, при котором определенное поведение продолжается до тех пор, пока не возникнут доказанные опасные или драматические последствия. [80] [ самостоятельно опубликованный источник? ]
Стресс
[ редактировать ]Социологи и психологи могут ссылаться на гомеостаз стресса , тенденцию населения или отдельного человека оставаться на определенном уровне стресса , часто вызывающую искусственный стресс, если «естественного» уровня стресса недостаточно. [81] [ самостоятельно опубликованный источник? ]
Жан-Франсуа Лиотар , теоретик постмодерна, применил этот термин к социальным «центрам власти», которые он описывает в «Состоянии постмодерна» как «управляемые принципом гомеостаза», например, к научной иерархии, которая иногда игнорирует радикальное новое открытие на долгие годы, потому что оно дестабилизирует ранее принятые нормы.
Технология
[ редактировать ]Известные технологические гомеостатические механизмы включают:
- Термостат работает путем включения и выключения обогревателей или кондиционеров в ответ на выходной сигнал датчика температуры.
- Круиз-контроль регулирует дроссельную заслонку автомобиля в зависимости от изменения скорости. [82] [83]
- Автопилот . управляет рулевым управлением самолета или корабля в ответ на отклонение от заранее установленного компасного направления или маршрута [84]
- Системы управления технологическими процессами на химическом или нефтеперерабатывающем заводе поддерживают уровни жидкости, давление, температуру, химический состав и т. д. путем управления нагревателями, насосами и клапанами. [85]
- Центробежный регулятор , парового двигателя разработанный Джеймсом Уаттом в 1788 году, уменьшает дроссельную заслонку в ответ на увеличение скорости двигателя или открывает клапан, если скорость падает ниже заданной скорости. [86] [87]
Общество и культура
[ редактировать ]Использование суверенной власти, кодексов поведения, религиозных и культурных практик и других динамических процессов в обществе можно охарактеризовать как часть развитой гомеостатической системы упорядочения жизни и поддержания общего равновесия, которая защищает безопасность целого от внутренних и внешние дисбалансы или опасности. [88] [89] Можно сказать, что здоровые гражданские культуры достигли оптимального гомеостатического баланса между множеством противоречивых проблем, таких как противоречие между уважением прав личности и заботой об общественном благе, [90] или между эффективностью правительства и реагированием на интересы граждан. [91] [92]
См. также
[ редактировать ]- Апоптоз - запрограммированная гибель клеток в многоклеточных организмах.
- Церебральная ауторегуляция - процесс у млекопитающих, целью которого является поддержание адекватного и стабильного мозгового кровотока.
- Хронобиология - Область биологии.
- Энантиостаз
- Геофизиология - научная гипотеза о
- Гликобиология - исследование сахаридов.
- Гомеорезис
- Гомеостатическая пластичность - способность нейронов регулировать собственную возбудимость относительно сетевой активности.
- Гормезис - Характеристика биологических процессов.
- Принцип Ле Шателье - принцип прогнозирования влияния изменения условий на химическое равновесие.
- Закон Ленца – Электромагнитное сопротивление изменениям
- Метаболостаз
- Осмос - миграция молекул в область с более низкой концентрацией растворенных веществ.
- Протеостаз - биологические пути внутри клеток, которые контролируют биогенез, сворачивание, транспортировку и деградацию белков, присутствующих внутри и снаружи клетки.
- Старение – ухудшение функций с возрастом.
- Устойчивое состояние – когда концентрации молекул в живой клетке или органе остаются постоянными.
- Системная биология - Вычислительное и математическое моделирование сложных биологических систем.
- Vis medicatrix naturae - латинская фраза, подтверждающая способность организма к самоисцелению.
Ссылки
[ редактировать ]- ^ Гордон., Беттс Дж. Анатомия и физиология . ДеСэ, Питер., Джонсон, Эдди., Джонсон, Джоди Э., Король, Оксана., Круз, Дин Х., По, Брэндон. Хьюстон, Техас. п. 9. ISBN 978-1-947172-04-3 . OCLC 1001472383 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мартин, Элизабет (2008). Биологический словарь (6-е изд.). Оксфорд: Издательство Оксфордского университета. стр. 315–316. ISBN 978-0-19-920462-5 .
- ^ Биология онлайн (27 октября 2019 г.). «Гомеостаз» . Биология онлайн . Архивировано из оригинала 12 августа 2020 года . Проверено 27 октября 2019 г.
- ^ Перейти обратно: а б Калаани, штат Нью-Йорк; Мангельсдорф, диджей (2006). «LXRS и FXR: инь и янь холестерина и жирового обмена». Ежегодный обзор физиологии . 68 : 159–91. doi : 10.1146/annurev.phyol.68.033104.152158 . ПМИД 16460270 .
- ^ Перейти обратно: а б с Мариб Э.Н., Хён К.Н. (2009). Основы анатомии и физиологии человека (9-е изд.). Сан-Франциско: Пирсон/Бенджамин Каммингс. ISBN 978-0-321-51342-7 .
- ^ Ловинджер, Дэвид М. (2008), «Пресинаптическая модуляция эндоканнабиноидами», в Зюдхофе, Томас К.; Старк, Клаус (ред.), Фармакология высвобождения нейротрансмиттеров , Справочник по экспериментальной фармакологии, том. 184, Springer Berlin Heidelberg, стр. 435–477, doi : 10.1007/978-3-540-74805-2_14 , ISBN. 978-3-540-74805-2 , PMID 18064422
- ^ Фрейтас, Геркулес Резенде; Исаак, Алинни Розендо; Мальчер-Лопес, Ренато; Диас, Бруно Лоуренсо; Тревензоли, Исида Хара; Рейс, Рикардо Аугусто Де Мело (26 ноября 2018 г.). «Полиненасыщенные жирные кислоты и эндоканнабиноиды в здоровье и болезни». Пищевая неврология . 21 (10): 695–714. дои : 10.1080/1028415X.2017.1347373 . ISSN 1028-415X . ПМИД 28686542 . S2CID 40659630 .
- ^ «Гомеостаз» . Словарь Merriam-Webster.com .
- ^ «Гомеостаз» . Dictionary.com Полный (онлайн). nd
- ^ Перейти обратно: а б Кэннон, ВБ (1932). Мудрость тела . Нью-Йорк: WW Нортон. стр. 177–201.
- ^ Перейти обратно: а б Кэннон, WB (1926). «Физиологическая регуляция нормальных состояний: некоторые попытки постулатов, касающихся биологической гомеостатики». В А. Петтите (ред.). Друзьям Чарльза Рича, его коллегам, его ученикам (на французском языке). Париж: Les Éditions Médicales. п. 91.
- ^ Смит, Джерард П. (2008). «Непризнанный вклад Павлова и Баркрофта в теорию гомеостаза Кэннона». Аппетит . 51 (3): 428–432. дои : 10.1016/j.appet.2008.07.003 . ПМИД 18675307 . S2CID 43088475 .
- ^ Зорея, Аарон (2014). Стероиды (проблемы здравоохранения и медицины сегодня) . Вестпорт, Коннектикут: Greenwood Press. п. 10. ISBN 978-1-4408-0299-7 .
- ^ Риггс, DS (1970). Теория управления и механизмы физиологической обратной связи . Балтимор: Уильямс и Уилкинс.
- ^ Перейти обратно: а б Холл, Джон (2011). Учебник Гайтона и Холла по медицинской физиологии (12-е изд.). Филадельфия, Пенсильвания: Сондерс/бихер. стр. 4–9. ISBN 978-1-4160-4574-8 .
- ^ Перейти обратно: а б Милсум, Дж. Х. (1966). Анализ систем биологического контроля . Нью-Йорк: МакГроу-Хилл.
- ^ Перейти обратно: а б с д и ж г Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Принципы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Издательство. стр. 315–316 , 475, 657–658. ISBN 978-0-06-350729-6 .
- ^ Ханская академия. «Гомеостаз» . Ханская академия . Архивировано из оригинала 20 октября 2019 года . Проверено 13 июля 2018 г.
- ^ Сведан, Надя Габриэле (2001). Женская спортивная медицина и реабилитация . Липпинкотт Уильямс и Уилкинс. п. 149. ИСБН 978-0-8342-1731-7 . Архивировано из оригинала 10 мая 2020 года . Проверено 11 октября 2019 г.
- ^ Вешлер, Тони (2002). Возьмите на себя ответственность за свою фертильность . Нью-Йорк: ХарперКоллинз. стр. 52 , 316, 361–362. ISBN 978-0-06-093764-5 .
- ^ Клюге, Мэтью Дж. (2015). Лихорадка: ее биология, эволюция и функции . Издательство Принстонского университета. п. 57. ИСБН 978-1-4008-6983-1 .
- ^ Гармель, Гас М. (2012). «Лихорадка у взрослых» . В Махадеване, СВ; Гармель, Гас М. (ред.). Введение в неотложную клиническую медицину (2-е изд.). Кембридж: Издательство Кембриджского университета. п. 375. ИСБН 978-0-521-74776-9 . Архивировано из оригинала 30 декабря 2019 года . Проверено 11 октября 2019 г.
- ^ Уэст, Брюс Дж (2006). Где медицина ошиблась: заново открывая путь к сложности . Исследования нелинейных явлений в науках о жизни. Том. 11. Нью-Джерси: World Scientific. дои : 10.1142/6175 . ISBN 978-981-256-883-0 . Архивировано из оригинала 6 марта 2022 года . Проверено 23 января 2019 г.
- ^ Лонго, Джузеппе; Монтевиль, Маэль (2014). Перспективы организмов . Конспект лекций по морфогенезу. Спрингер. дои : 10.1007/978-3-642-35938-5 . ISBN 978-3-642-35937-8 . S2CID 27653540 .
- ^ Шеннон, Клод Э.; Уивер, Уоррен (1963). Математическая теория связи (4-е печатное изд.). Урбана: Издательство Университета Иллинойса. ISBN 978-0-252-72548-7 .
- ^ Ракер, Р. (1987). Инструменты разума: математика информации . Хармондсворт: Книги Пингвинов. стр. 25–30.
- ^ Коэслаг, Йохан Х.; Сондерс, Питер Т.; Весселс, Джабус А. (1999). «Хромогранины и контррегуляторные гормоны: имеют ли они гомеостатический смысл?» . Журнал физиологии . 517 (3): 643–649. дои : 10.1111/j.1469-7793.1999.0643s.x . ПМК 2269385 . ПМИД 10358106 .
- ^ Перейти обратно: а б с Уильямс, Питер Л.; Уорик, Роджер; Дайсон, Мэри; Баннистер, Лоуренс Х. (1989). Анатомия Грея (двадцать седьмое изд.). Эдинбург: Черчилль Ливингстон. стр. 691–692, 791, 10011–10012. ISBN 0-443-04177-6 .
- ^ Тэнси, Этейн А.; Джонсон, Кристофер Д. (2015). «Последние достижения в терморегуляции» . Достижения в области физиологического образования . 39 (3): 139–148. дои : 10.1152/advan.00126.2014 . ISSN 1043-4046 . ПМИД 26330029 . S2CID 11553866 . Архивировано из оригинала 10 мая 2020 года . Проверено 26 января 2019 г.
- ^ Стэндринг, Сьюзен (7 августа 2015 г.). Анатомия Грея: анатомические основы клинической практики . Стэндринг, Сьюзен (41-е изд.). [Филадельфия]. стр. 141, 151–152. ISBN 978-0-7020-6851-5 . OCLC 920806541 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Первс, Дейл (2011). Нейронаука (5-е изд.). Сандерленд, Массачусетс: Синауэр. п. 458. ИСБН 978-0-87893-695-3 .
- ^ Перейти обратно: а б с Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Издательская компания Бенджамина/Каммингса. стр. 897–898. ISBN 978-0-8053-1800-5 .
- ^ Флорис, AD (январь 2011 г.). «Функциональная архитектура поведенческой терморегуляции». Европейский журнал прикладной физиологии . 111 (1): 1–8. дои : 10.1007/s00421-010-1602-8 . ПМИД 20711785 . S2CID 9109352 .
- ^ Гилрой, Энн М.; Макферсон, Брайан Р.; Росс, Лоуренс М. (2008). Атлас анатомии . Штутгарт: Медицинское издательство Thieme. стр. 318, 349. ISBN. 978-1-60406-062-1 .
- ^ Шмидт-Нильсен К (1981). «Противоточные системы у животных». Научный американец . 244 (5): 118–28. Бибкод : 1981SciAm.244e.118S . doi : 10.1038/scientificamerican0581-118 . ПМИД 7233149 .
- ^ Стюарт, ИК (2011). Физиология человека (Двенадцатое изд.). Нью-Йорк: МакГроу-Хилл. п. 667.
- ^ Робертшоу, Дэвид (август 2006 г.). «Механизмы контроля респираторной потери тепла через испарение у тяжело дышащих животных» . Журнал прикладной физиологии . 101 (2): 664–668. doi : 10.1152/japplphysicalol.01380.2005 . ISSN 8750-7587 . ПМИД 16675613 .
- ^ Багаван, Невада (2002). Медицинская биохимия (4-е изд.). Академическая пресса . п. 499. ИСБН 978-0-12-095440-7 . Архивировано из оригинала 6 марта 2022 года . Проверено 21 октября 2020 г.
- ^ Перейти обратно: а б с Коэслаг, Йохан Х.; Сондерс, Питер Т.; Тербланш, Элмари (2003). «Тематический обзор: переоценка гомеостата глюкозы в крови, которая всесторонне объясняет комплекс диабета 2 типа и синдрома X» . Журнал физиологии . 549 (Часть 2): 333–346. дои : 10.1113/jphysicalol.2002.037895 . ПМК 2342944 . ПМИД 12717005 .
- ^ Страйер, Люберт (1995). Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 164, 773–774. ISBN 0-7167-2009-4 .
- ^ Рихтер, Эрик А.; Харгривз, Марк (июль 2013 г.). «Упражнения, GLUT4 и усвоение глюкозы скелетными мышцами» . Физиологические обзоры . 93 (3): 993–1017. doi : 10.1152/physrev.00038.2012 . ISSN 0031-9333 . ПМИД 23899560 .
- ^ Аронофф, Стивен Л.; Берковиц, Кэти; Шрейнер, Барб; Хочу, Лаура (1 июля 2004 г.). «Метаболизм и регуляция глюкозы: помимо инсулина и глюкагона» . Диабетический спектр . 17 (3): 183–190. дои : 10.2337/диаспект.17.3.183 . ISSN 1040-9165 . Архивировано из оригинала 3 января 2020 года . Проверено 19 июля 2018 г.
- ^ «Современное понимание гомеостаза железа» . Academic.oup.com . Проверено 4 марта 2023 г.
- ^ Беркхед, Джейсон Л.; Гоголин Рейнольдс, Кэтрин А.; Абдель-Гани, Салах Э.; Коху, Кристофер М.; Пилон, Маринус (июнь 2009 г.). «Медный гомеостаз» . Новый фитолог . 182 (4): 799–816. дои : 10.1111/j.1469-8137.2009.02846.x . ISSN 0028-646X . ПМИД 19402880 .
- ^ Спайер, К.М.; Гурин А.В. (12 сентября 2009 г.). «Хемосенсорные пути в стволе мозга, контролирующие кардиореспираторную активность» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1529): 2603–10. дои : 10.1098/rstb.2009.0082 . ПМК 2865116 . ПМИД 19651660 .
- ^ Пикок, Эндрю Дж. (17 октября 1998 г.). «Кислород на большой высоте» . Британский медицинский журнал . 317 (7165): 1063–1066. дои : 10.1136/bmj.317.7165.1063 . ПМЦ 1114067 . ПМИД 9774298 .
- ^ Янг, Эндрю Дж; Ривз, Джон Т. (2002). «Адаптация человека к большой высоте» (PDF) . Медицинские аспекты суровых условий окружающей среды . Том. 2. Институт Бордена, Вашингтон, округ Колумбия: Управление главного хирурга армии США. CiteSeerX 10.1.1.175.3270 . Архивировано из оригинала (PDF) 16 сентября 2012 года . Проверено 5 января 2009 г.
- ^ Харрис, Н. Стюарт; Нельсон, Сара В. (16 апреля 2008 г.). «Высотная болезнь – церебральные синдромы» . Специальности электронной медицины > Неотложная медицина > Экология . Архивировано из оригинала 12 июня 2016 года . Проверено 30 июня 2016 г.
- ^ Альбертс, Брюс (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк [ua]: Гирлянда. стр. 1292–1293. ISBN 978-0-8153-4072-0 .
- ^ Перейти обратно: а б Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Издательство. стр. 444–445 . ISBN 978-0-06-350729-6 .
- ^ Фишер Дж.В., Кури С., Дьюси Т., Мендель С. (1996). «Продукция эритропоэтина интерстициальными клетками гипоксических почек обезьян». Британский журнал гематологии . 95 (1): 27–32. дои : 10.1046/j.1365-2141.1996.d01-1864.x . ПМИД 8857934 . S2CID 38309595 .
- ^ Йелькманн В. (2007). «Эритропоэтин после столетия исследований: моложе, чем когда-либо». Европейский журнал гематологии . 78 (3): 183–205. дои : 10.1111/j.1600-0609.2007.00818.x . ПМИД 17253966 . S2CID 37331032 .
- ^ «Обзор черепно-мозговых травм у взрослых: пакет для самообучения» (PDF) . Региональное здравоохранение Орландо. 2004. Архивировано из оригинала (PDF) 27 февраля 2008 года.
- ^ Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Издательство Оксфордского университета. п. 4. ISBN 978-0-19-856878-0 .
- ^ Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (5-е изд.). Харпер и Роу. п. 430 . ISBN 978-0-06-350729-6 .
- ^ Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Издательство Оксфордского университета. стр. 299–302. ISBN 978-0-19-856878-0 .
- ^ Брини, Мариса; Оттолини, Денис; Кали, Тито; Карафоли, Эрнесто (2013). «Кальций в здоровье и болезни». В Сигеле, Астрид; Хельмут, Роланд КО (ред.). Взаимосвязь между ионами незаменимых металлов и заболеваниями человека . Ионы металлов в науках о жизни. Том. 13. Спрингер. стр. 81–137. дои : 10.1007/978-94-007-7500-8_4 . ISBN 978-94-007-7499-5 . ПМИД 24470090 .
- ^ Страйер, Люберт (1995). «Витамин D получается из холестерина под действием света, расщепляющего кольца». В: Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. п. 707. ИСБН 0-7167-2009-4 .
- ^ Фельзенфельд, AJ; Левин, бакалавр наук (23 марта 2015 г.). «Кальцитонин, забытый гормон: заслуживает ли он того, чтобы его забыли?» . Клинический журнал почек . 8 (2): 180–187. дои : 10.1093/ckj/sfv011 . ISSN 2048-8505 . ПМК 4370311 . ПМИД 25815174 . Архивировано из оригинала 6 марта 2022 года . Проверено 18 июня 2021 г.
- ^ Аяканну, Тангесверан; Тейлор, Энтони Х.; Марсило, Тимоти Х.; Уиллетс, Джонатон М.; Конье, Джастин С. (2013). «Эндоканнабиноидная система и рак, зависимый от половых стероидов» . Международный журнал эндокринологии . 2013 : 259676. doi : 10.1155/2013/259676 . ISSN 1687-8337 . ПМЦ 3863507 . ПМИД 24369462 .
- ^ Перейти обратно: а б с д и Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Издательство. стр. 420–421 . ISBN 978-0-06-350729-6 .
- ^ Престон, Ричард А.; Матерсон, Би Джей; Реда, диджей; Уильямс, Д.В.; Гамбургер, Р.Дж.; Кушман, туалет; Андерсон, Р.Дж. (1998). «Статья JAMA, январь 2012 г.» . ДЖАМА . 280 (13): 1168–72. дои : 10.1001/jama.280.13.1168 . ПМИД 9777817 .
- ^ Уильямс Г.Х., Длуги Р.Г. (2008). «Глава 336: Заболевания коры надпочечников». Лоскальцо Дж., Фаучи А.С., Браунвальд Э., Каспер Д.Л., Хаузер С.Л., Лонго Д.Л. (ред.). Принципы внутренней медицины Харрисона . Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-146633-2 .
- ^ Бауэр Дж. Х., Гаунтнер WC (март 1979 г.). «Влияние хлорида калия на активность ренина плазмы и альдостерона плазмы во время ограничения натрия у нормального человека» . Почки Int . 15 (3): 286–93. дои : 10.1038/ki.1979.37 . ПМИД 513492 .
- ^ Ху С., Русин К.Г., Тан З., Гуальярдо Н.А., Барретт П.К. (июнь 2012 г.). «Клетки клубочковой зоны коры надпочечников мыши являются собственными электрическими генераторами» . Джей Клин Инвест . 122 (6): 2046–2053. дои : 10.1172/JCI61996 . ПМЦ 3966877 . ПМИД 22546854 .
- ^ Палмер, LG; Фриндт, Г (2000). «Секреция альдостерона и калия корковыми собирательными трубочками» . Почки Интернешнл . 57 (4): 1324–8. дои : 10.1046/j.1523-1755.2000.00970.x . ПМИД 10760062 .
- ^ Линас С.Л., Петерсон Л.Н., Андерсон Р.Дж., Айзенбри Г.А., Саймон Ф.Р., Берл Т. (июнь 1979 г.). «Механизм сохранения калия почками у крыс» . Почки Интернешнл . 15 (6): 601–11. дои : 10.1038/ki.1979.79 . ПМИД 222934 .
- ^ Перейти обратно: а б Тортора, Джерард Дж.; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Издательство. стр. 581–582 , 675–676. ISBN 978-0-06-350729-6 .
- ^ Сакка, Л.; Колл, Г.; Чазал, Дж. (декабрь 2011 г.). «Анатомия и физиология спинномозговой жидкости» . Европейские анналы оториноларингологии, болезней головы и шеи . 128 (6): 309–316. дои : 10.1016/j.anorl.2011.03.002 . ПМИД 22100360 .
- ^ Саладин, Кеннет (2012). Анатомия и физиология (6-е изд.). МакГроу Хилл. стр. 519–20.
- ^ Флорес, CE; Мендес, П. (2014). «Формирование торможения: структурная пластичность ГАМКергических синапсов, зависящая от активности» . Границы клеточной нейронауки . 8 : 327. дои : 10.3389/fncel.2014.00327 . ПМК 4209871 . ПМИД 25386117 .
- ^ Хм, Джи Вон (13 ноября 2017 г.). «Роль глиальных клеток в формировании тормозных синапсов и нервных цепей» . Границы молекулярной нейронауки . 10 :381. doi : 10.3389/fnmol.2017.00381 . ПМК 5694142 . ПМИД 29180953 .
- ^ Тони, Р. (2004). «Нейроэндокринная система: организация и гомеостатическая роль». Журнал эндокринологических исследований . 27 (6 дополнений): 35–47. ПМИД 15481802 .
- ^ Левицкий Д.А.; Сьюэлл, А; Чжун, Ю; Барре, Л; Шоен, С; Агаронник, Н; Леклер, JL; Чжо, Вт; Пакановский, К. (1 февраля 2019 г.). «Количественная оценка неточности потребления энергии людьми для компенсации навязанных энергетических ошибок: проблема физиологического контроля потребления пищи человеком». Аппетит . 133 : 337–343. дои : 10.1016/j.appet.2018.11.017 . ПМИД 30476522 . S2CID 53712116 .
- ^ Армстронг CM, Кота Дж (март 1999 г.). «Кальциевый блок Na + каналов и его влияние на скорость закрытия» . Proceedings of the National Academy of Sciences of the United States of America . 96 (7): 4154–7. Bibcode : 1999PNAS...96.4154A . doi : 10.1073/pnas.96.7.4154 22436 ПМЦ .
- ^ Харрисон, TR Принципы внутренней медицины (третье изд.). Нью-Йорк: Книжная компания McGraw-Hill. стр. 170, 571–579.
- ^ Лавлок, Джеймс (1991). Исцеление Гайи: Практическая медицина для планеты . Нью-Йорк: Книги Гармонии. ISBN 978-0-517-57848-3 .
- ^ Борон, Уолтер Ф.; Булпаеп, Эмиль Л. (2009). Медицинская физиология: клеточный и молекулярный подход (2-е Международное изд.). Филадельфия, Пенсильвания: Сондерс/Эльзевир. ISBN 978-1-4160-3115-4 .
- ^ Коэслаг, Дж. Х.; Сондерс, ПТ; Весселс, Дж. А. (1997). «Гомеостаз глюкозы с бесконечным приростом: дальнейшие уроки из притчи о мире ромашек?». Журнал эндокринологии . 134 (2): 187–192. дои : 10.1677/joe.0.1540187 . ПМИД 9291828 .
- ^ Спенсер, Лачи (2015). Флотация: руководство по сенсорной депривации, релаксации и изоляционным резервуарам . Лулу.com. п. 29. ISBN 978-1-329-17375-0 . [ самостоятельный источник ]
- ^ Спенсер, Лачи (29 мая 2015 г.). Флотация: руководство по сенсорной депривации, релаксации и изоляционным резервуарам . Лулу.com. ISBN 978-1-329-17375-0 . Архивировано из оригинала 3 января 2020 года . Проверено 11 октября 2019 г. [ самостоятельный источник ]
- ^ «Американ Моторс 1966» . Автомобильная жизнь . 12 : 46. 1965. Архивировано из оригинала 2 января 2020 года . Проверено 9 марта 2015 г.
- ^ Ницца, Карим (15 января 2001 г.). «Как работают системы круиз-контроля» . Как все работает. Архивировано из оригинала 6 марта 2015 года . Проверено 9 марта 2015 г.
- ^ Харрис, Уильям (10 октября 2007 г.). «Как работает автопилот» . HowStuffWorks.com . Архивировано из оригинала 15 апреля 2018 года . Проверено 14 апреля 2018 г.
- ^ Уайт, Дуглас (3 октября 2005 г.). «Передовая технология автоматизации снижает затраты на электроэнергию на нефтеперерабатывающем заводе» . Нефтегазовый журнал . Архивировано из оригинала 13 июля 2018 года . Проверено 13 июля 2018 г.
- ^ Максвелл, Джеймс Клерк (1868). «О губернаторах». Труды Лондонского королевского общества . 16 : 270–283. дои : 10.1098/rspl.1867.0055 . JSTOR 112510 . S2CID 51751195 .
- ^ Беннетт, Стюарт (1992). История техники управления, 1930-1955 гг . ИЭПП. п. п. 48 . ISBN 978-0-86341-299-8 .
- ^ Дамасио, Антонио (2018). Странный порядок мышления: жизнь, чувства и создание культур (под ред. электронной книги). Нью-Йорк: Книги Пантеона. п. 27. ISBN 978-0-307-90876-6 .
- ^ Воган-Уильямс, Ник (2017). «Карл Шмитт, Джорджио Агамбен и «номос» современной политической жизни». В Прозоров, Сергей; Рентеа, Симона (ред.). Справочник Рутледжа по биополитике . Лондон: Рутледж-Тейлор и Фрэнсис. п. 146. ИСБН 978-1-315-61275-1 .
- ^ Лим, Тэ Соп; Ан, Сохун (2015). «Диалектика культуры и динамическое балансирование между индивидуальностью и коллективностью». Журнал Азиатско-Тихоокеанской коммуникации . 25 (1): 63–77. дои : 10.1075/japc.25.1.04lim .
- ^ Уикхэм, Кэрри Розефски (2020). «Переосмысление истоков гражданской культуры и почему это важно для изучения арабского мира» . Правительство и оппозиция . 55 (1): 1–20. дои : 10.1017/gov.2019.12 . S2CID 151139202 .
- ^ Павоне, Таммазо (2014). «Политическая культура и демократический гомеостаз: критический обзор книги Габриэля Алмонда и Сидни Вербы «Гражданская культура» (PDF) . Принстонский университет. п. 2 . Проверено 30 июня 2022 г.
Дальнейшее чтение
[ редактировать ]- Клаузен, MJ; Поульсен, Х. (2013). «Гомеостаз натрия и калия в клетке». В Банки, Люсия (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 41–67. дои : 10.1007/978-94-007-5561-1_3 . ISBN 978-94-007-5560-4 . ПМИД 23595670 . электронная книга ISBN 978-94-007-5561-1 ISSN 1559-0836 электронный- ISSN 1868-0402
Внешние ссылки
[ редактировать ]- Гомеостаз. Архивировано 15 августа 2017 года в Wayback Machine.
- Уолтер Брэдфорд Кэннон, Гомеостаз (1932)
