Термогенез
Эта статья нуждается в дополнительных цитатах для проверки . ( февраль 2007 г. ) |
Термогенез — процесс производства тепла в организме . Он встречается у всех теплокровных животных, а также у некоторых видов термогенных растений, таких как восточная скунсовая капуста , лилия Вуду ( Sauromatum venosum ) и гигантские кувшинки рода Виктория . Омела сосновая карликовая, Arceuthobium americanum , быстро рассеивает свои семена посредством термогенеза. [1]
Типы
[ редактировать ]В зависимости от того, инициируются ли они посредством локомоции и преднамеренного движения мышц , термогенные процессы можно классифицировать как один из следующих:
- Термогенез физической активности ( EAT )
- Термогенез без физических упражнений ( NEAT ), энергия, расходуемая на все, кроме сна, еды или спортивных упражнений. [2]
- Диетический термогенез ( ДИТ )
Дрожь
[ редактировать ]Одним из способов повышения температуры является дрожь . Он производит тепло, потому что преобразование химической энергии АТФ в кинетическую энергию приводит к тому, что почти вся энергия проявляется в виде тепла. Дрожь — это процесс, при котором температура тела млекопитающих, находящихся в спячке (таких как некоторые летучие мыши и суслики), повышается, когда эти животные выходят из спячки.
не дрожит
[ редактировать ]Этот раздел может быть слишком техническим для понимания большинства читателей . ( декабрь 2018 г. ) |
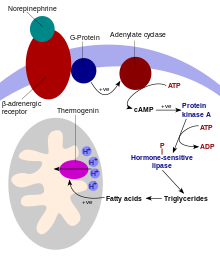
Несократительный термогенез происходит в бурой жировой ткани (буром жире). [3] присутствует почти у всех эвтериан ( свиньи являются единственным известным в настоящее время исключением). [4] [5] ). [6] Бурая жировая ткань содержит уникальный разобщающий белок ( термогенин , также известный как разобщающий белок 1), который позволяет отделить синтез АТФ от производства протонов ( H + ), что позволяет митохондриям сжигать жирные кислоты и кислород для выработки тепла. [7] Атомная структура человеческого разобщающего белка 1 UCP1 была расшифрована с помощью криогенной электронной микроскопии. Структура имеет типичную складку члена семейства SLC25 . [8] [9] UCP1 фиксируется в открытом цитоплазматическом состоянии гуанозинтрифосфатом в зависимости от pH, предотвращая утечку протонов. [10]
В этом процессе такие вещества, как свободные жирные кислоты (полученные из триацилглицеринов ), снимают пуриновое (АДФ, ВВП и др.) ингибирование термогенина, что вызывает приток H + в матрикс митохондрии и минуя канал АТФ-синтазы . Это разъединяет окислительное фосфорилирование , и энергия движущей силы протонов рассеивается в виде тепла, а не производит АТФ из АДФ, который будет хранить химическую энергию для использования организмом. Термогенез также может быть вызван утечкой натриево-калиевого насоса и Ca 2+ насос. [11] Термогенезу способствуют бесполезные циклы , такие как одновременное возникновение липогенеза и липолиза. [12] или гликолиз и глюконеогенез . В более широком контексте на бесполезные циклы могут влиять циклы активности/отдыха, такие как цикл Саммерматтера . [13]
Ацетилхолин стимулирует мышцы, повышая скорость метаболизма . [14]
Низкие потребности термогенеза означают, что свободные жирные кислоты по большей части используют липолиз как метод производства энергии.
полный список генов человека и мыши, регулирующих холодовой термогенез (CIT) у живых животных ( in vivo ) или образцов тканей ( ex vivo ). Был собран [15] и доступен в CITGeneDB. [16]
Эволюционная история
[ редактировать ]У птиц и эвтерий
[ редактировать ]Биологические процессы, обеспечивающие термогенез у животных, не произошли от одного общего предка . [17] Скорее, птичьи (птицы) и эвтерийные (плацентарные млекопитающие) линии развили способность осуществлять термогенез независимо посредством отдельных эволюционных процессов. [17] Тот факт, что один и тот же эволюционный признак развился независимо в двух разных линиях после их последнего известного общего предка, означает, что термогенные процессы классифицируются как пример конвергентной эволюции . Однако, хотя обе клады способны осуществлять термогенез, задействованные биологические процессы различны. Причина, по которой и птицы, и плацентарные особи развили способность осуществлять термогенез, является предметом постоянного изучения биологами-эволюционистами , и были предложены два конкурирующих объяснения, объясняющих, почему этот признак проявляется в обеих линиях. [17]
Одним из объяснений конвергенции является модель «аэробной способности». Эта теория предполагает, что естественный отбор отдавал предпочтение особям с более высокой скоростью метаболизма в состоянии покоя , и что по мере увеличения метаболической способности птиц и плаценты у них развивалась способность к эндотермическому термогенезу. [18] Исследователи связали высокий уровень потребления кислорода с высокой скоростью метаболизма в состоянии покоя, предполагая, что эти два явления напрямую коррелируют. Вместо того, чтобы животные развивали способность поддерживать высокие и стабильные температуры тела только для того, чтобы иметь возможность терморегуляции без помощи окружающей среды, эта теория предполагает, что термогенез на самом деле является побочным продуктом естественного отбора для более высоких аэробных и метаболических способностей. [19] Эти более высокие метаболические способности изначально могли возникнуть по той простой причине, что животные, способные усваивать больше кислорода в течение более длительных периодов времени, были бы лучше приспособлены, например, для того, чтобы убегать от хищников или собирать пищу. [19] Эта модель, объясняющая развитие термогенеза, более старая и более широко принята среди биологов-эволюционистов, изучающих термогенез.
Второе объяснение – модель «родительской заботы». Эта теория предполагает, что конвергентная эволюция термогенеза у птиц и плаценты основана на общих поведенческих чертах . В частности, и птицы, и эвтерии обеспечивают высокий уровень родительской заботы о молодом потомстве. Предполагается, что такой высокий уровень ухода дает новорожденным или вылупившимся животным возможность быстрее взрослеть, поскольку им приходится тратить меньше энергии на удовлетворение своих потребностей в пище, жилье и температуре. [17] Таким образом, модель «родительской заботы» предполагает, что у родителей была выбрана более высокая аэробная способность как средство удовлетворения потребностей их потомства . [18] Хотя модель «родительской заботы» действительно отличается от модели «аэробных способностей», она имеет некоторое сходство в том, что оба объяснения повышения термогенеза основаны на естественном отборе, отдающем предпочтение людям с более высокими аэробными способностями по той или иной причине. Основное различие между двумя теориями заключается в том, что модель «родительской заботы» предполагает, что определенная биологическая функция (забота о детях) привела к избирательному давлению для более высоких скоростей метаболизма.
Несмотря на то, что оба объяснения основаны на сходных объяснениях процесса, благодаря которому организмы приобрели способность осуществлять недрожательный термогенез, ни одно из этих объяснений не обеспечило достаточно широкого консенсуса, чтобы считаться полностью авторитетным в отношении конвергентной эволюции NST у птиц и млекопитающих, и ученые продолжают провести исследования, подтверждающие обе позиции. [19] [17] [18]
Несократительный термогенез
[ редактировать ]Термогенез бурой жировой ткани (БЖТ) является одной из двух известных форм несократительного термогенеза (НСТ). Этот тип тепловыделения встречается только у плаценты, но не у птиц или других термогенных организмов. BAT NST возникает, когда разобщающий белок 1 (UCP1) осуществляет окислительное фосфорилирование в телах плаценты, что приводит к выработке тепла (Berg et al., 2006, стр. 1178). [20] Этот процесс обычно начинается у плаценты только после того, как они подверглись воздействию низких температур в течение длительного периода времени, после чего этот процесс позволяет организму поддерживать высокую и стабильную температуру, не полагаясь на механизмы терморегуляции окружающей среды (такие как солнечный свет/тень). ). Поскольку эвтерии — единственная клада, хранящая бурую жировую ткань, ученые ранее считали, что UCP1 эволюционировал вместе с бурой жировой тканью. Однако недавние исследования показали, что UCP1 также можно обнаружить у животных, не являющихся животными, например, у рыб, птиц и рептилий. [21] Это открытие означает, что UCP1, вероятно, существовал у общего предка до возникновения эвтерийской линии. Однако после этого эволюционного раскола UCP1 развился независимо у плакучих в результате процесса, который, по мнению ученых, был вызван не естественным отбором, а скорее нейтральными процессами, такими как генетический дрейф . [21]
Эволюция несократительного термогенеза скелетных мышц
[ редактировать ]Вторая форма НСТ встречается в скелетных мышцах. В то время как плацента использует для термогенеза как BAT, так и NST скелетных мышц, птицы используют только последнюю форму. Также было показано, что этот процесс в редких случаях происходит у рыб . [17] При NST скелетных мышц ионы кальция скользят по мышечным клеткам, генерируя тепло. [17] Хотя первоначально считалось, что BAT NST является единственным процессом, с помощью которого животные могут поддерживать эндотермию, теперь ученые подозревают, что NST скелетных мышц была исходной формой этого процесса и что BAT NST развился позже. [17] Хотя ученые когда-то также считали, что только птицы поддерживают температуру своего тела с помощью NST скелетных мышц, исследования конца 2010-х годов показали, что млекопитающие и другие плацентарные животные также используют этот процесс, когда у них нет достаточных запасов бурой жировой ткани в организме. [22]
NST скелетных мышц также может быть использован для поддержания температуры тела у гетеротермных млекопитающих во время состояний оцепенения или гибернации . [17] Учитывая, что ранние плафоны и рептилии, которые позже развились в птичьи линии, были либо гетеротермными, либо экзотермными, считается, что обе формы NST не развились полностью до вымирания K-pg примерно 66 миллионов лет назад. [23] Однако по некоторым оценкам эволюция этих персонажей произошла раньше, примерно 100 млн лет назад. [24] Наиболее вероятно, что процесс развития способности к термогенезу в том виде, в котором он существует в настоящее время, был процессом, который начался до исчезновения K-pg и завершился значительно позже. Тот факт, что NST скелетных мышц распространен среди плаценты в периоды оцепенения и гибернации, еще раз подтверждает теорию о том, что эта форма термогенеза старше, чем BAT NST. Это связано с тем, что ранние эвтерианцы не обладали способностью к недрожательному термогенезу в том виде, в котором он существует в настоящее время, поэтому они чаще использовали оцепенение и спячку в качестве средств терморегуляции, полагаясь на системы, которые теоретически возникли до BAT NST. Однако среди биологов-эволюционистов до сих пор нет единого мнения ни о порядке развития этих двух процессов, ни о точных временных рамках их эволюции.
Регулирование
[ редактировать ]Несократительный термогенез регулируется главным образом гормонами щитовидной железы и симпатической нервной системой . Некоторые гормоны, такие как норадреналин и лептин , могут стимулировать термогенез, активируя симпатическую нервную систему. Повышение уровня инсулина после еды может быть причиной термогенеза, вызванного диетой ( термический эффект пищи ).Прогестерон также повышает температуру тела .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ деБрюн Р.А., Паэткау М., Росс К.А., Годфри Д.В., Фридман Ч.Р. (февраль 2015 г.). «Распространение семян карликовой омелы, вызванное термогенезом» . Природные коммуникации . 6 (1): 6262. Бибкод : 2015NatCo...6.6262D . дои : 10.1038/ncomms7262 . ПМК 4347025 . ПМИД 25662062 .
- ^ Левин Дж. А. (декабрь 2002 г.). «Термогенез без физических упражнений (NEAT)». Лучшие практики и исследования. Клиническая эндокринология и обмен веществ . 16 (4): 679–702. дои : 10.1053/beem.2002.0227 . ПМИД 12468415 .
- ^ Фокс С.И. (2011). Физиология человека (Двенадцатое изд.). МакГроу Хилл. п. 667.
- ^ Берг Ф., Густавсон У., Андерссон Л. (август 2006 г.). «Ген разобщающего белка 1 (UCP1) нарушен в линии свиней: генетическое объяснение плохой терморегуляции у поросят» . ПЛОС Генетика . 2 (8): е129. дои : 10.1371/journal.pgen.0020129 . ПМК 1550502 . ПМИД 16933999 .
- ^ Хоу Л, Ху С.И., Ван С. (апрель 2017 г.). «У свиньи нет бурой жировой ткани» . Журнал ФАСЭБ . 31 (С1). doi : 10.1096/fasebj.31.1_supplement.lb695 .
- ^ Хейворд Дж. С., Лиссон, Пенсильвания (1992). «Эволюция бурого жира: его отсутствие у сумчатых и однопроходных». Канадский журнал зоологии . 70 (1): 171–179. дои : 10.1139/z92-025 .
- ^ Кэннон Б, Недергаард Дж (январь 2004 г.). «Бурая жировая ткань: функция и физиологическое значение». Физиологические обзоры . 84 (1): 277–359. doi : 10.1152/physrev.00015.2003 . ПМИД 14715917 . S2CID 14289041 .
- ^ Рупрехт Дж. Дж., Кунджи Э. Р. (март 2020 г.). «Семейство митохондриальных носителей SLC25: структура и механизм» . Тенденции биохимических наук . 45 (3): 244–258. дои : 10.1016/j.tibs.2019.11.001 . ПМЦ 7611774 . ПМИД 31787485 .
- ^ Кунджи Э.Р., Кинг М.С., Рупрехт Дж.Дж., Тангаратнараджа К. (сентябрь 2020 г.). «Семейство носителей SLC25: важные транспортные белки в митохондриальной физиологии и патологии» . Физиология . 35 (5): 302–327. дои : 10.1152/физиол.00009.2020 . ПМЦ 7611780 . ПМИД 32783608 .
- ^ Джонс С.А., Гогои П., Рупрехт Дж.Дж., Кинг М.С., Ли Ю., Зёгг Т. и др. (июнь 2023 г.). «Структурные основы ингибирования пуриновыми нуклеотидами человеческого разобщающего белка 1» . Достижения науки . 9 (22): eadh4251. Бибкод : 2023SciA....9H4251J . дои : 10.1126/sciadv.adh4251 . ПМЦ 10413660 . ПМИД 37256948 . S2CID 259002752 .
- ^ Морриссетт Дж. М., Франк Дж. П., Блок BA (март 2003 г.). «Характеристика изоформ рианодинового рецептора и Ca2+-АТФазы в органе термогенного нагревателя голубого марлина (Makaira nigricans)» . Журнал экспериментальной биологии . 206 (Часть 5): 805–812. дои : 10.1242/jeb.00158 . ПМИД 12547935 .
- ^ Солинас Г., Саммерматтер С., Майниери Д., Гублер М., Пирола Л., Виманн М.П. и др. (ноябрь 2004 г.). «Прямое влияние лептина на термогенез скелетных мышц опосредовано циклическим циклом субстратов между липогенезом de novo и окислением липидов» (PDF) . Письма ФЭБС . 577 (3): 539–544. дои : 10.1016/j.febslet.2004.10.066 . ПМИД 15556643 . S2CID 18266296 .
- ^ Саммерматтер С., Хандшин С. (ноябрь 2012 г.). «PGC-1α и упражнения по контролю массы тела» . Международный журнал ожирения . 36 (11): 1428–1435. дои : 10.1038/ijo.2012.12 . ПМИД 22290535 .
- ^ Эванс С.С., Репаски Е.А., Фишер Д.Т. (июнь 2015 г.). «Лихорадка и терморегуляция иммунитета: иммунная система чувствует тепло» . Обзоры природы. Иммунология . 15 (6): 335–349. дои : 10.1038/nri3843 . ПМЦ 4786079 . ПМИД 25976513 .
- ^ Ли Дж, Дэн С.П., Вэй Г, Ю П (январь 2018 г.). «CITGeneDB: обширная база данных генов человека и мыши, усиливающих или подавляющих термогенез, индуцированный холодом, подтвержденная экспериментами по возмущению на мышах» . База данных . 2018 . дои : 10.1093/база данных/bay012 . ПМК 5868181 . ПМИД 29688375 .
- ^ Ли Дж, Дэн С.П., Вэй Г, Ю П (январь 2018 г.). «CITGeneDB: обширная база данных генов человека и мыши, усиливающих или подавляющих термогенез, индуцированный холодом, подтвержденная экспериментами по возмущению на мышах» . База данных . 2018 . дои : 10.1093/база данных/bay012 . ПМК 5868181 . ПМИД 29688375 .
- ^ Jump up to: а б с д и ж г час я Новак Дж., Жиру С., Арнольд В., Руф Т. (09.11.2017). «Мышечный несократительный термогенез и его роль в эволюции эндотермии» . Границы в физиологии . 8 : 889. дои : 10.3389/fphys.2017.00889 . ПМК 5684175 . ПМИД 29170642 .
- ^ Jump up to: а б с Котея П. (март 2000 г.). «Ассимиляция энергии, родительская забота и эволюция эндотермии» . Слушания. Биологические науки . 267 (1442): 479–484. дои : 10.1098/rspb.2000.1025 . ПМК 1690555 . ПМИД 10737405 .
- ^ Jump up to: а б с Беннетт А.Ф., Рубен Дж.А. (ноябрь 1979 г.). «Энотермия и активность у позвоночных». Наука . 206 (4419): 649–654. Бибкод : 1979Sci...206..649B . дои : 10.1126/science.493968 . ПМИД 493968 .
- ^ Берг Ф., Густавсон У., Андерссон Л. (август 2006 г.). Барш Г.С. (ред.). «Ген разобщающего белка 1 (UCP1) нарушен в линии свиней: генетическое объяснение плохой терморегуляции у поросят» . ПЛОС Генетика . 2 (8): е129. дои : 10.1371/journal.pgen.0020129 . ПМК 1550502 . ПМИД 16933999 .
- ^ Jump up to: а б Хьюз Д.А., Ястрох М., Стоункинг М., Клингенспор М. (январь 2009 г.). «Молекулярная эволюция UCP1 и эволюционная история несократительного термогенеза млекопитающих» . Эволюционная биология BMC . 9 (1): 4. Бибкод : 2009BMCEE...9....4H . дои : 10.1186/1471-2148-9-4 . ПМЦ 2627829 . ПМИД 19128480 .
- ^ Бал Н.К., Маурья С.К., Сингх С., Веренс XH, Периасами М. (август 2016 г.). «Повышенная зависимость от мышечного термогенеза при резкой минимизации функции бурой жировой ткани» . Журнал биологической химии . 291 (33): 17247–17257. дои : 10.1074/jbc.M116.728188 . ПМК 5016124 . ПМИД 27298322 .
- ^ Лавгроув Б.Г. (2012). «Единое происхождение гетеротермии у млекопитающих». Жизнь в сезонном мире . Берлин, Гейдельберг: Springer Berlin Heidelberg. стр. 3–11. дои : 10.1007/978-3-642-28678-0_1 . ISBN 978-3-642-28677-3 .
- ^ Лабоча МК, Хейс Дж. П. (2019). «Энотерм». Энциклопедия экологии . Эльзевир. стр. 368–374. дои : 10.1016/b978-0-12-409548-9.11221-7 . ISBN 9780444641304 .
Внешние ссылки
[ редактировать ]- Термогенез Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)