Генетический дрейф
| Часть серии о |
| Эволюционная биология |
|---|
 |
Генетический дрейф , также известный как случайный генетический дрейф , аллельный дрейф или эффект Райта . [1] — это изменение частоты существующего варианта гена ( аллеля ) в популяции вследствие случайности. [2]
Генетический дрейф может привести к полному исчезновению вариантов генов и тем самым к уменьшению генетической изменчивости . [3] Это также может привести к тому, что изначально редкие аллели станут гораздо более частыми и даже фиксированными.
Когда существует мало копий аллели, эффект генетического дрейфа более заметен, а когда существует много копий, эффект менее заметен (из-за закона больших чисел ). В середине 20-го века разгорелись энергичные дебаты по поводу относительной важности естественного отбора и нейтральных процессов, включая генетический дрейф. Рональд Фишер , который объяснил естественный отбор, используя менделевскую генетику , [4] придерживались мнения, что генетический дрейф играет незначительную роль в эволюции , и эта точка зрения оставалась доминирующей в течение нескольких десятилетий. В 1968 году популяционный генетик Мотоо Кимура возобновил дебаты своей нейтральной теорией молекулярной эволюции , которая утверждает, что большинство случаев распространения генетических изменений по популяции (хотя и не обязательно изменений фенотипов ) вызваны генетическим дрейфом, действующим на нейтральные мутации . [5] [6] В 1990-х годах была предложена конструктивная нейтральная эволюция , целью которой является объяснение того, как сложные системы возникают посредством нейтральных переходов. [7] [8]
Аналогия с шариками в банке [ править ]
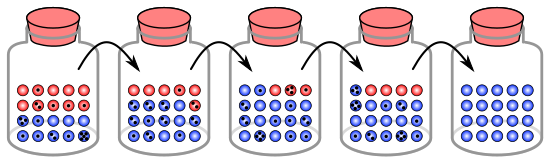
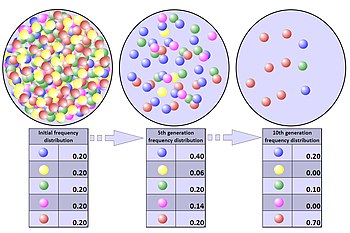
Процесс генетического дрейфа можно проиллюстрировать, используя 20 шариков в банке, представляющих 20 организмов в популяции. [9] Считайте эту банку с шариками стартовой популяцией. Половина шариков в банке красная, а половина — голубая, причем каждый цвет соответствует отдельному аллелю одного гена в популяции. В каждом новом поколении организмы размножаются случайным образом. Чтобы изобразить эту репродукцию, случайным образом выберите шарик из исходной банки и поместите новый шарик того же цвета в новую банку. Это «потомок» оригинального мрамора, а это означает, что оригинальный мрамор остается в банке. Повторяйте этот процесс, пока во второй банке не окажется 20 новых шариков. Во второй банке теперь будет 20 «потомков» или шариков разного цвета. Если во второй банке не содержится ровно 10 красных шариков и 10 синих шариков, то в частотах аллелей произошел случайный сдвиг.
Если этот процесс повторяется несколько раз, количество красных и синих шариков, выбираемых в каждом поколении, будет колебаться. Иногда в банке больше красных шариков, чем в «родительской» банке, а иногда и больше синих. Это колебание аналогично генетическому дрейфу – изменению частоты аллелей в популяции в результате случайного изменения распределения аллелей от одного поколения к другому.
Ни в одном поколении не могло быть выбрано ни одного шарика определенного цвета, то есть у них не было потомства. В этом примере, если красные шарики не выбраны, банка, представляющая новое поколение, будет содержать только синее потомство. Если это произойдет, красный аллель будет навсегда потерян в популяции, а оставшийся синий аллель закрепится: все будущие поколения будут полностью синими. В небольших популяциях фиксация может произойти всего за несколько поколений.
и Вероятность частота аллелей
Механизмы генетического дрейфа можно проиллюстрировать на упрощенном примере. Рассмотрим очень большую колонию бактерий, изолированную в капле раствора. Бактерии генетически идентичны, за исключением одного гена с двумя аллелями, обозначенными A и B , которые являются нейтральными аллелями, что означает, что они не влияют на способность бактерий выживать и размножаться; все бактерии в этой колонии имеют одинаковую вероятность выживания и размножения. Предположим, что половина бактерий имеет аллель А а другая половина — аллель В. , Таким образом, A и B имеют частоту аллелей 1/2.
Затем капля раствора сжимается до тех пор, пока в ней не останется еды, достаточной только для поддержания четырех бактерий. Все остальные бактерии погибают, не размножаясь. Среди четырех выживших существует 16 возможных комбинаций аллелей A и B :
(АААА), (БААА), (АБАА), (ББАА),
(ААБА), (БАБА), (АББА), (БББА),
(АААБ), (БААБ), (АБАБ), (ББАБ),
(ААББ), (БАББ), (АБББ), (ББББ).
Поскольку все бактерии в исходном растворе с одинаковой вероятностью выживут при усадке раствора, четыре выживших представляют собой случайную выборку из исходной колонии. Вероятность того , что каждый из четырех выживших имеет данный аллель, равна 1/2, поэтому вероятность того, что какая-либо конкретная комбинация аллелей произойдет при сокращении раствора, равна

(Первоначальный размер популяции настолько велик, что выборка фактически происходит с заменой). Другими словами, каждая из 16 возможных комбинаций аллелей встречается с одинаковой вероятностью, с вероятностью 1/16.
Подсчет комбинаций с одинаковым количеством А и Б дает следующую таблицу:
| А | Б | Комбинации | Вероятность |
| 4 | 0 | 1 | 1/16 |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 0 | 4 | 1 | 1/16 |
Как показано в таблице, общее количество комбинаций, имеющих одинаковое количество аллелей А и аллелей В, равно шести, а вероятность этой комбинации равна 6/16. Общее количество остальных комбинаций равно десяти, поэтому вероятность неодинакового количества аллелей А и В равна 10/16. Таким образом, хотя исходная колония начиналась с равным числом аллелей А и В , вполне возможно, что число аллелей в оставшейся популяции из четырех членов не будет равным. Ситуация равных чисел на самом деле менее вероятна, чем неравных чисел. В последнем случае генетический дрейф произошел из-за того, что частоты аллелей в популяции изменились из-за случайной выборки. В этом примере популяция сократилась до четырех случайных выживших — явление, известное как « узкое место в популяции» .
Вероятности количества выживших копий аллели A (или B ) (приведенные в последнем столбце приведенной выше таблицы) можно рассчитать непосредственно из биномиального распределения , где вероятность «успеха» (вероятность присутствия данной аллели ) равна 1/2 (т. е. вероятность того, что в комбинации имеется k копий аллелей A (или B )) определяется выражением:

где n=4 – количество выживших бактерий.
Математические модели [ править ]
Математические модели генетического дрейфа могут быть разработаны с использованием либо процессов ветвления , либо уравнения диффузии, описывающего изменения частоты аллелей в идеализированной популяции . [10]
Модель Райта-Фишера [ править ]
Рассмотрим ген с двумя аллелями A или B. : При диплоидии популяции, состоящие из N особей, имеют по 2 N копий каждого гена. У человека может быть две копии одной и той же аллели или две разные аллели. Частоте одного аллеля присвоено p , а другого q . Модель Райта-Фишера (названная в честь Сьюэлла Райта и Рональда Фишера ) предполагает, что поколения не перекрываются (например, однолетние растения имеют ровно одно поколение в год) и что каждая копия гена, обнаруженная в новом поколении, рисуется независимо и случайным образом. от всех копий гена в старом поколении. Тогда формула для расчета вероятности получения k копий аллели, которая имела частоту p в последнем поколении, выглядит следующим образом: [11] [12]

где символ « ! » означает функцию факториала . Это выражение также можно сформулировать с использованием биномиального коэффициента ,

Модель Морана [ править ]
Модель Морана предполагает перекрывающиеся поколения. На каждом временном этапе одна особь выбирается для размножения, а другая – для смерти. Таким образом, на каждом временном этапе количество копий данной аллели может увеличиваться на одну, уменьшаться на одну или оставаться прежним. Это означает, что перехода трехдиагональна матрица , а это означает, что математические решения проще для модели Морана, чем для модели Райта-Фишера. С другой стороны, компьютерное моделирование обычно легче выполнять с использованием модели Райта-Фишера, поскольку необходимо рассчитывать меньше временных шагов. В модели Морана требуется N для прохождения одного поколения временных шагов, где N — эффективный размер популяции . В модели Райта-Фишера требуется только один. [13]
На практике модели Морана и Райта-Фишера дают качественно схожие результаты, но генетический дрейф в модели Морана происходит в два раза быстрее.
Другие модели дрифта [ править ]
Если дисперсия в количестве потомков намного больше, чем дисперсия, определяемая биномиальным распределением, предполагаемым моделью Райта-Фишера, то при той же общей скорости генетического дрейфа (эффективный размер дисперсии), генетический дрейф является менее мощной силой. по сравнению с выбором. [14] Даже для той же дисперсии, если более высокие моменты распределения числа потомков превышают моменты биномиального распределения, то сила генетического дрейфа снова существенно ослабляется. [15]
кроме выборки Случайные эффекты , ошибки
Случайные изменения частот аллелей также могут быть вызваны эффектами, отличными от ошибки выборки , например случайными изменениями давления отбора. [16]
Одним из важных альтернативных источников стохастичности , возможно, более важных, чем генетический дрейф, является генетический драфт . [17] Генетический драфт — это воздействие на локус путем отбора связанных локусов. Математические свойства генетического дрейфа отличаются от свойств генетического дрейфа. [18] Направление случайного изменения частоты аллелей автокоррелирует между поколениями. [2]
Дрейф и фиксация [ править ]
Принцип Харди-Вайнберга гласит, что в достаточно больших популяциях частоты аллелей остаются постоянными от одного поколения к другому, если равновесие не нарушается миграцией , генетическими мутациями или отбором . [19]
Однако в конечных популяциях новые аллели не получаются в результате случайной выборки аллелей, передаваемых следующему поколению, но выборка может привести к исчезновению существующей аллели. Поскольку случайная выборка может удалить, но не заменить аллель, а также поскольку случайное снижение или увеличение частоты аллелей влияет на ожидаемое распределение аллелей для следующего поколения, генетический дрейф со временем приводит популяцию к генетическому единообразию. Когда аллель достигает частоты 1 (100%), ее называют «фиксированной» в популяции, а когда аллель достигает частоты 0 (0%), она теряется. Меньшие популяции достигают фиксации быстрее, тогда как в пределе бесконечной популяции фиксация не достигается. Как только аллель становится фиксированной, генетический дрейф прекращается, и частота аллелей не может измениться, если в популяцию не будет введен новый аллель посредством мутации или потока генов . устраняет генетические вариации . Таким образом, даже несмотря на то, что генетический дрейф является случайным и бесцельным процессом, он со временем [20]
Скорость изменения частоты аллелей из за дрейфа -

Если предположить, что генетический дрейф является единственной эволюционной силой, действующей на аллель, то после t поколений во многих реплицируемых популяциях, начиная с частот аллелей p и q , дисперсия частот аллелей между этими популяциями равна

Время фиксации или потери [ править ]
Если предположить, что генетический дрейф — единственная эволюционная сила, действующая на аллель, то в любой момент времени вероятность того, что аллель в конечном итоге закрепится в популяции, — это просто ее частота в популяции в данный момент. [22] Например, если частота p для аллели A составляет 75%, а частота q для аллели B составляет 25%, то при неограниченном времени вероятность A в конечном итоге закрепится в популяции равна 75%, а вероятность того, что B зафиксируется, равна 25%.
Ожидаемое количество поколений, в течение которых произойдет фиксация, пропорционально размеру популяции, так что, по прогнозам, фиксация будет происходить гораздо быстрее в меньших популяциях. [23] Обычно для определения этих вероятностей используется эффективный размер популяции, который меньше общей численности населения. Эффективная популяция ( N e ) учитывает такие факторы, как уровень инбридинга , стадию жизненного цикла, на которой популяция является наименьшей, и тот факт, что некоторые нейтральные гены генетически связаны с другими, находящимися в стадии отбора. [14] Эффективный размер популяции может не быть одинаковым для каждого гена в одной и той же популяции. [24]
Одна перспективная формула, используемая для приблизительного расчета ожидаемого времени до того, как нейтральный аллель зафиксируется в результате генетического дрейфа, в соответствии с моделью Райта-Фишера:

где T — количество поколений, N e — эффективный размер популяции, а p — начальная частота данного аллеля. Результатом является количество поколений, которое, как ожидается , пройдет до того, как произойдет фиксация данного аллеля в популяции с заданным размером ( N e ) и частотой аллели ( p ). [25]
Ожидаемое время потери нейтрального аллеля в результате генетического дрейфа можно рассчитать как [11]

Когда мутация возникает только один раз в популяции, достаточно большой, чтобы начальная частота была незначительной, формулы можно упростить до [26]

для среднего числа поколений, ожидаемых до фиксации нейтральной мутации, и

для среднего числа ожидаемых поколений до потери нейтральной мутации в популяции фактического размера N. [27]
Время до потери как с дрейфом, так и с мутацией
Приведенные выше формулы применимы к аллелю, который уже присутствует в популяции и не подвержен ни мутациям, ни естественному отбору. Если аллель теряется в результате мутации гораздо чаще, чем приобретается в результате мутации, то мутация, а также дрейф могут влиять на время потери. Если аллель, склонная к мутационной потере, начинается как фиксированная в популяции и теряется в результате мутации со скоростью m на репликацию, то ожидаемое время в поколениях до ее потери в гаплоидной популяции определяется выражением
![{\displaystyle {\bar {T}}_{\text{lost}}\approx {\begin{cases}{\dfrac {1}{m}},{\text{ if }}mN_{e}\ll 1\\[8pt]{\dfrac {\ln {(mN_{e})}+\gamma }{m}}{\text{ if }}mN_{e}\gg 1\end{cases}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/0dca9b7dc746a7b2a8d8a63770ac53782d1639e3)
где — постоянная Эйлера . [28] Первое приближение представляет собой время ожидания до тех пор, пока не появится первый мутант, обреченный на потерю, причем потеря затем происходит относительно быстро из-за генетического дрейфа, требуя времени. 1 / м ≫ Н е . Второе приближение представляет время, необходимое для детерминированной потери из-за накопления мутаций. В обоих случаях время фиксации определяется мутацией через член 1 / м , и на него меньше влияет эффективная численность популяции .

Против естественного отбора [ править ]
В естественных популяциях генетический дрейф и естественный отбор не действуют изолированно; оба явления всегда действуют вместе с мутацией и миграцией. Нейтральная эволюция является продуктом как мутации, так и дрейфа, а не только дрейфа. Точно так же, даже когда отбор подавляет генетический дрейф, он может воздействовать только на вариации, которые обеспечивает мутация.
В то время как естественный отбор имеет направление, ведущее эволюцию к наследственной адаптации к текущей среде, генетический дрейф не имеет направления и направляется только математикой случая . [29] В результате дрейф влияет на частоты генотипов внутри популяции без учета их фенотипических эффектов. Напротив, отбор благоприятствует распространению аллелей, фенотипические эффекты которых увеличивают выживаемость и/или воспроизводство их носителей, снижает частоты аллелей, вызывающих неблагоприятные признаки, и игнорирует нейтральные. [30]
Закон больших чисел предсказывает, что когда абсолютное число копий аллели невелико (например, в небольших популяциях ), величина дрейфа частот аллелей за поколение больше. Величина дрейфа достаточно велика, чтобы подавить отбор на любой частоте аллелей, когда коэффициент отбора меньше 1, разделенного на эффективный размер популяции. Таким образом, неадаптивная эволюция, возникающая в результате мутаций и генетического дрейфа, считается последовательным механизмом эволюционных изменений, прежде всего в небольших изолированных популяциях. [31] Математика генетического дрейфа зависит от эффективного размера популяции, но неясно, как это связано с фактическим количеством особей в популяции. [17] Генетическая связь с другими генами, находящимися в процессе селекции, может уменьшить эффективный размер популяции, присущий нейтральному аллелю. При более высокой скорости рекомбинации сцепление уменьшается, а вместе с ним и локальное влияние на эффективный размер популяции. [32] [33] Этот эффект виден в молекулярных данных как корреляция между скоростью локальной рекомбинации и генетическим разнообразием . [34] и отрицательная корреляция между плотностью генов и разнообразием некодирующих участков ДНК . [35] Стохастичность, связанная со связью с другими генами, находящимися в стадии отбора, — это не то же самое, что ошибка выборки, и иногда ее называют генетическим драфтом , чтобы отличить ее от генетического дрейфа. [17]
Низкая частота аллелей делает аллели более уязвимыми к случайному уничтожению, даже преодолевая влияние естественного отбора. Например, хотя невыгодные мутации обычно быстро устраняются в популяции, новые полезные мутации почти так же уязвимы к утрате в результате генетического дрейфа, как и нейтральные мутации. Генетический дрейф не будет иметь никакого эффекта до тех пор, пока частота аллелей полезной мутации не достигнет определенного порога. [30]
в численности населения Узкое место
Узкое место в популяции — это когда популяция сокращается до значительно меньшего размера за короткий период времени из-за какого-то случайного события окружающей среды. В условиях настоящего узкого места в популяции шансы на выживание любого члена популяции чисто случайны и не улучшаются каким-либо конкретным генетическим преимуществом. Узкое место может привести к радикальным изменениям частот аллелей, совершенно независимо от отбора. [36]
Воздействие узкого места в численности населения может сохраняться, даже если узкое место вызвано разовым событием, например, природной катастрофой.Интересным примером узкого места, вызывающего необычное генетическое распределение, является относительно высокая доля людей с полной из палочек цветовой слепотой ( ахроматопсия ) на атолле Пингелап в Микронезии . [37] После узкого места инбридинг увеличивается. Это увеличивает ущерб, наносимый рецессивными вредными мутациями, в процессе, известном как инбредная депрессия . Отбираются худшие из этих мутаций, что приводит к потере других аллелей, генетически связанных с ними, в процессе фонового отбора . [2] В случае рецессивных вредных мутаций этот отбор может быть усилен вследствие «узкого места» из-за генетической чистки . Это приводит к дальнейшей потере генетического разнообразия. Кроме того, устойчивое сокращение численности популяции увеличивает вероятность дальнейших колебаний аллелей из-за дрейфа в будущих поколениях.
Генетическая изменчивость популяции может быть значительно уменьшена с помощью узкого места, и даже полезные адаптации могут быть навсегда устранены. [38] Утрата изменчивости делает выжившую популяцию уязвимой для любого нового давления отбора, такого как болезни, климатические изменения или смена доступного источника пищи, поскольку адаптация в ответ на изменения окружающей среды требует достаточной генетической изменчивости в популяции для того, чтобы произошел естественный отбор. [39] [40]
В недавнем прошлом было много известных случаев демографического затруднения. До прибытия прерии европейцев Северной Америки были средой обитания миллионов прерийных кур . Только в Иллинойсе их численность упала с примерно 100 миллионов птиц в 1900 году до примерно 50 птиц в 1990-х годах. Сокращение популяции произошло в результате охоты и разрушения среды обитания , но следствием этого стала потеря большей части генетического разнообразия вида. Анализ ДНК , сравнивающий птиц середины века с птицами 1990-х годов, документирует резкое снижение генетической изменчивости всего за последние несколько десятилетий. В настоящее время большие прерийные куры демонстрируют низкий репродуктивный успех . [41]
Однако генетические потери, вызванные узким местом и генетическим дрейфом, могут повысить приспособленность, как в случае с эрлихиями . [42]
Чрезмерная охота также привела к серьезному сокращению популяции северного морского слона в 19 веке. В результате этого снижение генетической изменчивости можно сделать, сравнив его с таковым у южного морского слона , на которого охотились не так агрессивно. [43]
Эффект Основателя [ править ]
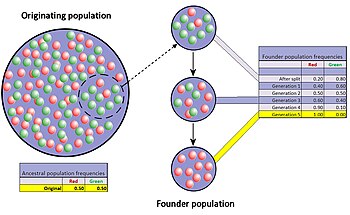
Эффект основателя — это особый случай узкого места в популяции, возникающий, когда небольшая группа в популяции отделяется от исходной популяции и образует новую. Ожидается, что случайная выборка аллелей в только что сформировавшейся новой колонии будет сильно искажать исходную популяцию, по крайней мере, в некоторых отношениях. [44] Возможно даже, что число аллелей некоторых генов в исходной популяции превышает число копий генов у основателей, что делает полное представление невозможным. Когда вновь образованная колония небольшая, ее основатели могут сильно повлиять на генетический состав популяции в далеком будущем.
Хорошо документированный пример можно найти в миграции амишей в Пенсильванию в 1744 году. Два члена новой колонии имели общий рецессивный аллель синдрома Эллиса-Ван Кревельда . Члены колонии и их потомки, как правило, являются религиозной изоляцией и остаются относительно изолированными. В результате инбридинга на протяжении многих поколений синдром Эллиса-Ван Кревельда в настоящее время гораздо более распространен среди амишей, чем среди населения в целом. [30] [45]
Разница в частотах генов между исходной популяцией и колонией может также привести к значительному расхождению этих двух групп на протяжении многих поколений. По мере увеличения разницы, или генетического расстояния , две разделенные популяции могут стать разными как генетически, так и фенетически , хотя этому расхождению способствуют не только генетический дрейф, но и естественный отбор, поток генов и мутации. Возможность относительно быстрых изменений частоты генов в колонии побудила большинство ученых считать эффект основателя (и, как следствие, генетический дрейф) важной движущей силой эволюции новых видов . Сьюэлл Райт был первым, кто придал такое значение случайному дрейфу и небольшим, недавно изолированным популяциям в своей смещающегося баланса . теории видообразования [46] Вслед за Райтом Эрнст Майр создал множество убедительных моделей, чтобы показать, что снижение генетической изменчивости и небольшой размер популяции после эффекта основателя были критически важны для развития новых видов. [47] Однако сегодня эта точка зрения пользуется гораздо меньшей поддержкой, поскольку гипотеза неоднократно проверялась посредством экспериментальных исследований, и результаты были в лучшем случае двусмысленными. [48]
История [ править ]
Роль случайного шанса в эволюции была впервые описана Арендом Л. Хагедорном и Анной Корнелией Хагедорн-Ворстёвель Ла Бранд в 1921 году. [49] Они подчеркнули, что случайная выживаемость играет ключевую роль в потере изменчивости популяций. Фишер (1922) ответил на это первой, хотя и незначительно неверной, математической трактовкой «эффекта Хагедорна». [50] Примечательно, что он ожидал, что многие естественные популяции были слишком велики (N ~ 10 000), чтобы последствия дрейфа были существенными, а дрейф мыслей окажет незначительное влияние на эволюционный процесс. Исправленная математическая трактовка и термин «генетический дрейф» были позже придуманы основателем популяционной генетики Райтом Сьюэллом . Впервые он использовал термин «дрейф» в 1929 году. [51] хотя в то время он использовал это слово в смысле направленного процесса изменения или естественного отбора. Случайный дрейф, вызванный ошибкой выборки, стал известен как «эффект Сьюэлла-Райта», хотя ему никогда не было вполне комфортно видеть свое имя, данное ему. Райт называл все изменения частоты аллелей либо «устойчивым дрейфом» (например, отбором), либо «случайным дрейфом» (например, ошибкой выборки). [52] «Дрейф» стал использоваться как технический термин исключительно в стохастическом смысле. [53] Сегодня ее обычно определяют еще более узко, с точки зрения ошибки выборки. [54] хотя это узкое определение не является универсальным. [55] [56] Райт писал, что «ограничение «случайного дрейфа» или даже «дрейфа» только одним компонентом, последствиями случайных отборов проб, имеет тенденцию приводить к путанице». [52] Сьюэлл Райт считал процесс случайного генетического дрейфа посредством ошибки выборки эквивалентным процессу инбридинга, но более поздние работы показали, что они различны. [57]
На заре современного эволюционного синтеза ученые начали смешивать новую науку популяционной генетики с Чарльза Дарвина теорией естественного отбора . В рамках этой концепции Райт сосредоточил внимание на влиянии инбридинга на небольшие относительно изолированные популяции. Он представил концепцию адаптивного ландшафта , в котором такие явления, как скрещивание и генетический дрейф в небольших популяциях, могут оттолкнуть их от адаптивных пиков, что, в свою очередь, позволяет естественному отбору подтолкнуть их к новым адаптивным пикам. [58] Райт считал, что меньшие популяции больше подходят для естественного отбора, потому что «инбридинг был достаточно интенсивным, чтобы создавать новые системы взаимодействия посредством случайного дрейфа, но недостаточно интенсивным, чтобы вызвать случайную неадаптивную фиксацию генов». [59]
Взгляды Райта на роль генетического дрейфа в эволюционной схеме практически с самого начала были противоречивыми. Одним из самых громких и влиятельных критиков был коллега Рональд Фишер. Фишер признал, что генетический дрейф сыграл некоторую роль в эволюции, но незначительную. Фишера обвиняли в неправильном понимании взглядов Райта, поскольку в своей критике Фишер, казалось, утверждал, что Райт почти полностью отверг отбор. Для Фишера рассмотрение процесса эволюции как длительного, устойчивого и адаптивного прогресса было единственным способом объяснить постоянно растущую сложность более простых форм. Но дебаты между «постепенными» и теми, кто больше склоняется к модели эволюции Райта, где отбор и дрейф вместе играют важную роль, продолжаются. [60]
В 1968 году Мотоо Кимура возобновил дебаты своей нейтральной теорией молекулярной эволюции , которая утверждает, что большая часть генетических изменений вызвана генетическим дрейфом, действующим на нейтральные мутации. [5] [6]
Роль генетического дрейфа в эволюции посредством ошибки выборки подверглась критике со стороны Джона Х. Гиллеспи. [61] и Уильям Б. Провайн , которые утверждают, что отбор на связанных сайтах является более важной стохастической силой.
См. также [ править ]
- Перипатрическое видообразование
- Антигенный дрейф
- Коалесцентная теория
- Конструктивная нейтральная эволюция
- Генофонд
- Мейотический драйв
Примечания и ссылки [ править ]
- ^ Гулд С.Дж. (2002). «Глава 7, раздел «Синтез как упрочнение» ». Структура эволюционной теории .
- ^ Jump up to: Перейти обратно: а б с Мазель Дж. (октябрь 2011 г.). «Генетический дрейф» . Современная биология . 21 (20). Ячеечный пресс : R837-8. дои : 10.1016/j.cub.2011.08.007 . ПМИД 22032182 .
- ^ Звезда Б, Спенсер Х.Г. (май 2013 г.). «Влияние генетического дрейфа и потока генов на селективное поддержание генетических вариаций» . Генетика . 194 (1): 235–44. дои : 10.1534/genetics.113.149781 . ПМЦ 3632471 . ПМИД 23457235 .
- ^ Миллер 2000 , с. 54
- ^ Jump up to: Перейти обратно: а б Кимура М. (февраль 1968 г.). «Скорость эволюции на молекулярном уровне». Природа . 217 (5129). Издательская группа «Природа»: 624–6. Бибкод : 1968Natur.217..624K . дои : 10.1038/217624a0 . ПМИД 5637732 . S2CID 4161261 .
- ^ Jump up to: Перейти обратно: а б Футуйма 1998 , с. 320
- ^ Штольцфус А (1999). «О возможности конструктивной нейтральной эволюции» . Журнал молекулярной эволюции . 49 (2): 169–181. Бибкод : 1999JMolE..49..169S . дои : 10.1007/PL00006540 . ISSN 0022-2844 . ПМИД 10441669 . S2CID 1743092 . Архивировано из оригинала 30 июля 2022 года . Проверено 20 января 2022 г.
- ^ Муньос-Гомес С.А., Билоликар Г., Видеман Дж.Г., Гейлер-Самеротт К. (апрель 2021 г.). «Конструктивная нейтральная эволюция 20 лет спустя» . Журнал молекулярной эволюции . 89 (3): 172–182. Бибкод : 2021JMolE..89..172M . дои : 10.1007/s00239-021-09996-y . ПМЦ 7982386 . ПМИД 33604782 .
- ^ «Ошибка выборки и эволюция» . Понимание эволюции . Калифорнийский университет в Беркли . Архивировано из оригинала 8 декабря 2015 года . Проверено 1 декабря 2015 г.
- ^ Валь Л.М. (август 2011 г.). «Фиксация, когда N и s меняются: классические подходы дают новые элегантные результаты» . Генетика . 188 (4). Американское генетическое общество : 783–5. дои : 10.1534/genetics.111.131748 . ПМК 3176088 . ПМИД 21828279 .
- ^ Jump up to: Перейти обратно: а б Хартл и Кларк 2007 , с. 112
- ^ Тиан 2008 , с. 11
- ^ Моран П.А. (1958). «Случайные процессы в генетике». Математические труды Кембриджского философского общества . 54 (1): 60–71. Бибкод : 1958PCPS...54...60M . дои : 10.1017/S0305004100033193 . S2CID 85823386 .
- ^ Jump up to: Перейти обратно: а б Чарльзуорт Б. (март 2009 г.). «Фундаментальные концепции генетики: эффективный размер популяции и закономерности молекулярной эволюции и изменчивости». Обзоры природы. Генетика . 10 (3). Издательская группа Nature : 195–205. дои : 10.1038/nrg2526 . ПМИД 19204717 . S2CID 205484393 .
- ^ Дер Р., Эпштейн К.Л. , Плоткин Дж.Б. (сентябрь 2011 г.). «Обобщенные популяционные модели и природа генетического дрейфа». Теоретическая популяционная биология . 80 (2). Эльзевир : 80–99. дои : 10.1016/j.tpb.2011.06.004 . ПМИД 21718713 .
- ^ Ли и Граур 1991 , стр. 28.
- ^ Jump up to: Перейти обратно: а б с Гиллеспи Дж. Х. (ноябрь 2001 г.). «Влияет ли размер популяции вида на его эволюцию?» . Эволюция; Международный журнал органической эволюции . 55 (11). Джон Уайли и сыновья от Общества изучения эволюции : 2161–9. дои : 10.1111/j.0014-3820.2001.tb00732.x . ПМИД 11794777 . S2CID 221735887 .
- ^ Нехер Р.А., Шрайман Б.И. (август 2011 г.). «Генетическая тяга и квазинейтральность в больших факультативно-сексуальных популяциях» . Генетика . 188 (4). Американское генетическое общество: 975–96. arXiv : 1108.1635 . дои : 10.1534/genetics.111.128876 . ПМК 3176096 . ПМИД 21625002 .
- ^ Юэнс 2004
- ^ Ли и Граур 1991 , стр. 29.
- ^ Бартон и др. 2007 , с. 417
- ^ Футуйма 1998 , с. 300
- ^ Отто С.П. , Уитлок MC (июнь 1997 г.). «Вероятность фиксации в популяциях меняющегося размера» (PDF) . Генетика . 146 (2). Американское генетическое общество: 723–33. дои : 10.1093/генетика/146.2.723 . ПМК 1208011 . ПМИД 9178020 . Архивировано (PDF) из оригинала 19 марта 2015 г.
- ^ Каттер А.Д., Чой Дж.Ю. (август 2010 г.). «Естественный отбор формирует нуклеотидный полиморфизм в геноме нематоды Caenorhabditis briggsae» . Геномные исследования . 20 (8). Лабораторное издательство Колд-Спринг-Харбор : 1103–11. дои : 10.1101/гр.104331.109 . ПМК 2909573 . ПМИД 20508143 .
- ^ Хедрик 2005 , с. 315
- ^ Ли и Граур 1991 , стр. 33.
- ^ Кимура и Ота 1971
- ^ Мэйсел Дж. , Кинг О.Д., Моган Х. (январь 2007 г.). «Потеря адаптивной пластичности в течение длительного периода застоя окружающей среды» . Американский натуралист . 169 (1). Издательство Чикагского университета от имени Американского общества натуралистов : 38–46. дои : 10.1086/510212 . ПМК 1766558 . ПМИД 17206583 .
- ^ «Естественный отбор: как работает эволюция» . Экшнбионаука . Вашингтон, округ Колумбия: Американский институт биологических наук . Архивировано из оригинала 6 января 2010 года . Проверено 24 ноября 2009 г. Интервью с Дугласом Дж. Футуймой . См. ответ на вопрос: Является ли естественный отбор единственным механизмом эволюции?
- ^ Jump up to: Перейти обратно: а б с Кавалли-Сфорца, Меноцци и Пьяцца, 1996 г.
- ^ Циммер 2001
- ^ Голдинг 1994 , с. 46
- ^ Чарльзуорт Б., Морган М.Т., Чарльзуорт Д. (август 1993 г.). «Влияние вредных мутаций на нейтральные молекулярные вариации» (PDF) . Генетика . 134 (4). Американское генетическое общество: 1289–303. дои : 10.1093/генетика/134.4.1289 . ПМК 1205596 . ПМИД 8375663 . Архивировано (PDF) из оригинала 12 марта 2020 г. Проверено 9 декабря 2015 г.
- ^ Прегрейвс, округ Колумбия (сентябрь 2005 г.). «Рекомбинация усиливает адаптацию белков у Drosophila melanogaster» . Современная биология . 15 (18). Cell Press: 1651–6. дои : 10.1016/j.cub.2005.07.065 . ПМИД 16169487 . S2CID 15120927 .
- ^ Нордборг М, Ху Т., Ишино Ю, Джавери Дж, Тумаджян С, Чжэн Х, Баккер Е, Калабрезе П, Гладстон Дж, Гоял Р, Якобссон М, Ким С, Морозов Ю, Падхукасахасрам Б, Планнол В, Розенберг Н.А., Шах С , Уолл Дж.Д., Ван Дж., Чжао К., Калбфляйш Т., Шульц В., Крейтман М., Бергельсон Дж. (июль 2005 г.). «Схема полиморфизма Arabidopsis thaliana» . ПЛОС Биология . 3 (7). Публичная научная библиотека : e196. дои : 10.1371/journal.pbio.0030196 . ПМЦ 1135296 . ПМИД 15907155 .

- ^ Робинсон Р., изд. (2003). «Узкое место народонаселения» . Генетика . Том. 3. Нью-Йорк: Справочник Macmillan USA . ISBN 0-02-865609-1 . LCCN 2002003560 . OCLC 614996575 . Проверено 14 декабря 2015 г.
- ^ Хасселс И.Е., Мортон Н.Е. (май 1972 г.). «Атоллы Пингелап и Мокил: ахроматопсия» . Американский журнал генетики человека . 24 (3): 304–309. ПМЦ 1762260 . ПМИД 4555088 .
- ^ Футуйма 1998 , стр. 303–304.
- ^ О'Корри-Кроу Дж. (март 2008 г.). «Изменение климата и молекулярная экология арктических морских млекопитающих» . Экологические приложения . 18 (2 доп.). Экологическое общество Америки : S56-76. дои : 10.1890/06-0795.1 . ПМИД 18494363 .
- ^ Корнюэ Ж.М., Луикарт Дж. (декабрь 1996 г.). «Описание и анализ мощности двух тестов для выявления недавних узких мест в популяции на основе данных о частоте аллелей» . Генетика . 144 (4). Американское генетическое общество: 2001–14. doi : 10.1093/генетика/144.4.2001 . ПМЦ 1207747 . ПМИД 8978083 .
- ^ Садава и др. 2008 , гл. 1, 21–33, 52–57
- ^ Дейл С., Моран Н.А. (август 2006 г.). «Молекулярные взаимодействия между бактериальными симбионтами и их хозяевами» . Клетка . 126 (3): 453–65. дои : 10.1016/j.cell.2006.07.014 . ПМИД 16901780 . S2CID 15985536 .
- ^ «Узкие места и эффекты основателя» . Понимание эволюции . Калифорнийский университет, Беркли. Архивировано из оригинала 4 декабря 2015 года . Проверено 14 декабря 2015 г.
- ^ Кэмпбелл 1996 , с. 423
- ^ «Генетический дрейф и эффект основателя» . Библиотека эволюции (веб-ресурс). Эволюция. Бостон, Массачусетс: Образовательный фонд WGBH ; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595 . Архивировано из оригинала 14 марта 2009 года . Проверено 7 апреля 2009 г.
- ^ Вольф, Броди и Уэйд, 2000 г.
- ^ Привет, Fitch и Аяла, 2005 г.
- ^ Ховард и Берлохер, 1998 г.
- ^ Хагедорн А.Л., Хагедорн-Форстёвель Ла Бранд AC (1921). Относительная ценность процессов, вызывающих эволюцию . Гаага: Мартинус Нийхофф. Архивировано из оригинала 28 марта 2019 года . Проверено 28 марта 2019 г.
- ^ Фишер Р.А. (1922). «О соотношении доминирования». Труды Королевского общества Эдинбурга . 42 : 321–341. дои : 10.1017/s0370164600023993 .
- ^ Райт С. (ноябрь – декабрь 1929 г.). «Эволюция доминирования». Американский натуралист . 63 (689). Чикаго, Иллинойс: Издательство Чикагского университета от имени Американского общества натуралистов: 556–561. дои : 10.1086/280290 . ISSN 0003-0147 . JSTOR 2456825 . S2CID 85301374 .
- ^ Jump up to: Перейти обратно: а б Райт С. (1955). «Классификация факторов эволюции». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 20 . Лабораторный пресс Колд-Спринг-Харбор: 16–24D. дои : 10.1101/SQB.1955.020.01.004 . ПМИД 13433551 . Симпозиум: «Популяционная генетика: природа и причины генетической изменчивости популяций».
- ^ Стивенсон 1991
- ^ Фриман и Херрон, 2007 г.
- ^ Мазель Дж. (август 2012 г.). «Переосмысление Харди-Вайнберга и генетического дрейфа в бакалавриате по биологии». Биоэссе . 34 (8). Джон Уайли и сыновья: 701–710. doi : 10.1002/bies.201100178 . ПМИД 22576789 . S2CID 28513167 .
- ^ Линч 2007
- ^ Кроу Дж. Ф. (март 2010 г.). «Райт и Фишер об инбридинге и случайном дрейфе» . Генетика . 184 (3). Американское генетическое общество: 609–611. дои : 10.1534/genetics.109.110023 . ПМЦ 2845331 . ПМИД 20332416 .
- ^ Ларсон 2004 , стр. 221–243.
- ^ Стивенсон 1991 : Цитата, приписываемая Уильяму Б. Провайну в книге «Истоки теоретической популяционной генетики» (1971), стр. 162; Чикаго: Издательство Чикагского университета.
- ↑ Аверс 1989.
- ^ Гиллеспи Дж. Х. (июнь 2000 г.). «Генетический дрейф в бесконечной популяции. Модель псевдоавтостопа» . Генетика . 155 (2). Американское генетическое общество: 909–919. дои : 10.1093/генетика/155.2.909 . ПМК 1461093 . ПМИД 10835409 .
Библиография [ править ]
- Аверс CJ (1989). Процесс и закономерности в эволюции . Нью-Йорк: Издательство Оксфордского университета . ISBN 0-19-505275-7 . LCCN 88005368 . OCLC 17677554 .
- Бартон Н.Х. , Бриггс Д.Е. , Эйзен Дж.А. , Гольдштейн Д.Б. , Патель Н.Х. (2007). Эволюция . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор . ISBN 978-0-87969-684-9 . LCCN 2007010767 . ОСЛК 86090399 .
- Кэмпбелл Н.А. (1996). Биология . Серия Бенджамина/Каммингса по наукам о жизни (4-е изд.). Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании ISBN 0-8053-1940-9 . LCCN 95045572 . OCLC 33333455 .
- Кавалли-Сфорца Л.Л. , Меноцци П., Пьяцца А (1996). История и география человеческих генов (сокращенное издание в мягкой обложке). Принстон, Нью-Джерси: Издательство Принстонского университета . ISBN 0-691-02905-9 . ОСЛК 35527063 .
- Юэнс В.Дж. (2004). Математическая популяционная генетика I. Теоретическое введение . Междисциплинарная прикладная математика. Том. 27 (2-е изд.). Нью-Йорк: Springer-Verlag . ISBN 0-387-20191-2 . LCCN 2003065728 . OCLC 53231891 .
- Фриман С., Херрон Дж.К. (2007). Эволюционный анализ (4-е изд.). Река Аппер-Сэддл, Нью-Джерси: Пирсон Прентис Холл . ISBN 978-0-13-227584-2 . LCCN 2006034384 . ОСЛК 73502978 .
- Футуйма Д (1998). Эволюционная биология (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates . ISBN 0-87893-189-9 . LCCN 97037947 . ОСЛК 37560100 .
- Голдинг Б., изд. (1994). Ненейтральная эволюция: теории и молекулярные данные . Нью-Йорк: Чепмен и Холл . ISBN 0-412-05391-8 . LCCN 93047006 . ОСЛК 29638235 . «Материалы семинара, спонсируемого Канадским институтом перспективных исследований ».
- Хартл Д.Л., Кларк АГ (2007). Принципы популяционной генетики (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-308-2 . LCCN 2006036153 . OCLC 75087956 .
- Хедрик П.В. (2005). Генетика популяций (3-е изд.). Бостон, Массачусетс: Издательство Jones and Bartlett . ISBN 0-7637-4772-6 . LCCN 2004056666 . OCLC 56194719 .
- Эй Джей , Фитч В.М. , Аяла Ф.Дж. , ред. (2005). Систематика и происхождение видов: К 100-летию Эрнста Майра . Вашингтон, округ Колумбия: Издательство национальных академий . ISBN 978-0-309-09536-5 . LCCN 2005017917 . OCLC 70745851 .
- Ховард DJ, Берлохер С.Х., ред. (1998). Бесконечные формы: виды и видообразование . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-510901-6 . LCCN 97031461 . OCLC 37545522 .
- Кимура М. , Охта Т. (1971). Теоретические аспекты популяционной генетики . Монографии по популяционной биологии. Том. 4. Принстон, Нью-Джерси: Издательство Принстонского университета . стр. 1–219. ISBN 0-691-08096-8 . LCCN 75155963 . OCLC 299867647 . ПМИД 5162676 .
- Ларсон Э.Дж. (2004). Эволюция: замечательная история научной теории . Современные библиотечные хроники. Том. 17. Нью-Йорк: Современная библиотека . ISBN 0-679-64288-9 . LCCN 2003064888 . OCLC 53483597 .
- Ли В.Х. , Граур Д. (1991). Основы молекулярной эволюции . Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-452-9 . LCCN 90043581 . OCLC 22113526 .
- Линч М. (2007). Истоки архитектуры генома . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-484-3 . LCCN 2007000012 . OCLC 77574049 .
- Миллер Дж . (2000). Брачный разум: как сексуальный выбор повлиял на эволюцию человеческой природы . Нью-Йорк: Даблдей . ISBN 0-385-49516-1 . LCCN 00022673 . OCLC 43648482 .
- Садава Д., Хеллер Х.К. , Орианс Г.Х. , Первс В.К., Хиллис Д.М. (2008). Жизнь: наука биология . Том. II: Эволюция, разнообразие и экология (8-е изд.). Сандерленд, Массачусетс; Гордонсвилл, Вирджиния: Sinauer Associates; WH Фриман и компания . ISBN 978-0-7167-7674-1 . LCCN 2006031320 . OCLC 71632224 .
- Стивенсон Дж. К. (1991). Словарь понятий физической антропологии . Справочные источники по социальным и гуманитарным наукам. Том. 10. Вестпорт, Коннектикут: Greenwood Press . ISBN 0-313-24756-0 . LCCN 90022815 . OCLC 22732327 .
- Тянь Дж. П. (2008). Эволюционные алгебры и их приложения . Конспект лекций по математике. Том. 1921. Берлин; Нью-Йорк: Спрингер. дои : 10.1007/978-3-540-74284-5 . ISBN 978-3-540-74283-8 . LCCN 2007933498 . OCLC 173807298 . Збл 1136.17001 .
- Вольф Дж.Б., Броди Э.Д., Уэйд М.Дж. , ред. (2000). Эпистаз и эволюционный процесс . Оксфорд, Великобритания; Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-512806-0 . LCCN 99046515 . OCLC 42603105 .
- Циммер С (2001). Эволюция: Триумф идеи . Введение Стивена Джея Гулда ; предисловие Ричарда Хаттона (1-е изд.). Нью-Йорк: ХарперКоллинз . ISBN 0-06-019906-7 . LCCN 2001024077 . OCLC 46359440 .
Внешние ссылки [ править ]
- Шихи, Боб. «Программа моделирования популяционной генетики» . Рэдфорд, Вирджиния: Рэдфордский университет . Проверено 21 декабря 2015 г.
- Граймс, Билл. «Моделирование генетического дрейфа» . Тусон, Аризона: Университет Аризоны . Проверено 25 августа 2016 г.