Эволюция рептилий
| Часть серии о |
| Эволюционная биология |
|---|
 |
Рептилии возникли около 320 миллионов лет назад. [1] в каменноугольный период. Рептилии в традиционном смысле этого слова определяются как животные, имеющие чешую или щитки , откладывающие наземные яйца с твердой скорлупой и обладающие экзотермическим метаболизмом. Определенная таким образом, группа является парафилетической , исключая эндотермических животных, таких как птицы, которые произошли от ранних традиционно определяемых рептилий. Определение в соответствии с филогенетической номенклатурой , которая отвергает парафилетические группы, включает птиц, исключая млекопитающих и их синапсидных предков. В таком определении Reptilia идентична Sauropsida .
Хотя сегодня немногие рептилии являются высшими хищниками , в прошлом существовало множество примеров высших рептилий. Рептилии имеют чрезвычайно разнообразную эволюционную историю, которая привела к биологическим успехам, например , динозавры , птерозавры , плезиозавры , мозазавры и ихтиозавры .
Первые рептилии [ править ]
Ранние рептилии [ править ]


Зарождение рептилий лежит около 320–310 миллионов лет назад, в болотах позднего каменноугольного периода, когда первые рептилии произошли от продвинутых Хосеми лабиринтодонтов . [2] Самое старое известное животное, которое, возможно, было амниотой , рептилией, а не амфибией , — Казинерия. [3] [4] (хотя также утверждается, что это темноспондильная амфибия). [5]
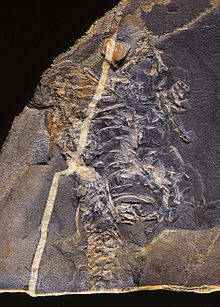
Серия следов из ископаемых слоев Новой Шотландии , датированная 315 миллионами лет назад, показывает типичные пальцы рептилий и отпечатки чешуи. [6] Следы приписывают Хилономусу , древнейшей из известных рептилий. [7] Это было маленькое, похожее на ящерицу животное длиной от 20 до 30 см (8–12 дюймов) с многочисленными острыми зубами, указывающими на насекомоядную диету. [8]
Другие примеры включают Вестлотиану (иногда считающуюся стеблевым амниотом, а не настоящим амниотом ). [9] и Палеотирис , оба одинакового телосложения и, предположительно, схожего образа жизни. Одной из самых известных ранних рептилий является Mesosaurus , род из ранней перми , который вернулся в воду и питался рыбой.
Самые ранние рептилии были в значительной степени затмены более крупными лабиринтодонтными амфибиями, такими как кохлеозавр , и оставались небольшой, незаметной частью фауны до окончания небольшого ледникового периода в конце каменноугольного периода.
Анапсиды, синапсиды, диапсиды и зауропсиды [ править ]

Традиционно предполагалось, что первые рептилии были анапсидами , имевшими твердый череп с отверстиями только для носа, глаз, спинного мозга и т. д.; [10] открытия синапсидоподобных отверстий в крыше черепа нескольких представителей Parareptilia , включая лантанозухоидов , миллереттид , болозавридов , некоторых никторолерид , некоторых проколофоноидов и, по крайней мере, некоторых мезозавров. [11] [12] [13] сделало это более двусмысленным, и в настоящее время неясно, имел ли предковая рептилия череп, похожий на анапсид или на синапсид. [13] Очень скоро после появления первых рептилий они разделились на две ветви. [14] Одна ветвь, Synapsida (включая современных млекопитающих), имела по одному отверстию в крыше черепа позади каждого глаза. Другая ветвь, Sauropsida , сама делится на две основные группы. В одном из них, упомянутом выше Parareptilia, были таксоны с анапсидоподобным черепом, а также таксоны с одним отверстием позади каждого глаза (см. выше). У представителей другой группы, Diapsida , в черепе позади каждого глаза было отверстие, а также второе отверстие, расположенное выше на черепе. Функция отверстий как в синапсидах, так и в диапсидах заключалась в том, чтобы облегчить череп и дать пространство для движения челюстных мышц, что позволяет осуществлять более сильный укус. [10]
Традиционно считалось, что черепахи являются выжившими анапсидами на основании строения их черепа. [15] Обоснование этой классификации оспаривалось: некоторые утверждали, что черепахи — это диапсиды, которые вернулись в это примитивное состояние, чтобы улучшить свою броню (см. Парарептилии ). [2] Более поздние морфологические филогенетические исследования с учетом этого поместили черепах прочно в состав Diapsida. [16] Все молекулярные исследования убедительно подтвердили размещение черепах в диапсидах, чаще всего как сестринскую группу современных архозавров . [17] [18] [19] [20]
Эволюция млекопитающих [ править ]
Пеликозавры |
Основная кладограмма происхождения млекопитающих.Важными событиями при переходе от рептилий к млекопитающим была эволюция теплокровности , прикуса зубов коренных , среднего уха с тремя косточками , волос и молочных желез . К концу триаса существовало много видов, похожих на современных млекопитающих, а к средней юре линии, ведущие к трем современным группам млекопитающих — однопроходным , сумчатым и плацентарным — разошлись.
Восстание динозавров [ править ]
Пермские рептилии [ править ]
Ближе к концу каменноугольного периода, когда еще существовали наземные рептилиоморфы- лабиринтодонты, из синапсид развились первые полностью наземные крупные позвоночные, пеликозавры , такие как эдафозавр . В среднепермский период климат стал более сухим, что привело к смене фауны: на смену примитивным пеликозаврам пришли более развитые терапсиды . [21]
Анапсидные рептилии, чьи массивные крыши черепа не имели заглазничных отверстий, существовали и процветали на протяжении всей перми. Парейазавры и в достигли гигантских размеров в поздней перми конце концов исчезли в конце периода. [21]
В конце этого периода диапсидные рептилии разделились на две основные линии: архозавров (предков крокодилов и динозавров) и лепидозавров (предшественников современных туатаров , ящериц и змей). Обе группы оставались похожими на ящериц, были относительно небольшими и незаметными в течение пермского периода.
Мезозойская эра, «Эра рептилий» [ править ]
В конце пермского периода произошло самое крупное известное массовое вымирание (см. Пермско-триасовое вымирание ). Большая часть ранней анапсидной/синапсидной мегафауны исчезла, уступив место архозавроморфным диапсидам. Архозавры характеризовались удлиненными задними ногами и прямой позой, ранние формы чем-то напоминали длинноногих крокодилов. Архозавры стали доминирующей группой в триасовый период, развившись в хорошо известных динозавров и птерозавров , а также псевдозухов . Мезозойскую эпоху часто называют «Эрой рептилий» — фраза, придуманная палеонтологом начала XIX века Гидеоном Мантеллом , который признал динозавров и предков крокодилов доминирующими наземными позвоночными. Некоторые из динозавров были крупнейшими наземными животными, когда-либо жившими на Земле, тогда как некоторые из более мелких теропод дали начало первым птицам . [21]
Родственной группой Archosauromorpha является Lepidosauromorpha , в которую входят чешуйчатые и ринхоцефалы , а также их ископаемые родственники. Lepidosauromorpha содержала по крайней мере одну основную группу мезозойских морских рептилий: мозазавров , появившихся в меловой период. Филогенетическое размещение других основных групп ископаемых морских рептилий — зауроптеригов и ихтиозавров , которые развились в раннем триасе и в среднем триасе соответственно — является более спорным. Разные авторы связывали эти группы либо с лепидозавроморфами, либо с лепидозавроморфами. [22] или архозавроморфам, [23] [24] [25] а ихтиозавры также считались диапсидами, которые не принадлежали к наименее инклюзивной кладе, содержащей лепидозавроморфы и архозавроморфы. [26]
Терапсиды подвергались растущему давлению со стороны динозавров юрского периода ; млекопитающие и тритилодонтиды к концу периода были единственными выжившими представителями этой линии.
Эволюция птиц [ править ]

Основными моментами перехода от пресмыкающегося к птице являются эволюция от чешуи к перьям , эволюция клюва ( хотя у других организмов развивалась независимо), галлофикация костей, развитие полета и теплокровности .
Считается, что эволюция птиц началась в юрский период, когда самые ранние птицы произошли от динозавров-теропод . Птицы относятся к биологическому классу Aves. Самый ранний известный вид в Авесе — Archeopteryx Lithographica , родом из позднеюрского периода . динозавров Современная филогенетика относит птиц к кладе Theropoda . Согласно нынешнему консенсусу, Aves и Crocodilia являются единственными живыми членами неранговой клады Archosauria .
| Целурозавры |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Упрощенная кладограмма Сентера (2007). [27]
Гибель динозавров [ править ]
Конец мелового периода ознаменовал упадок мегафауны рептилий мезозойской эры. Наряду с огромной вулканической активностью в то время падение метеорита, создавшее границу мела и палеогена считается основной причиной этого массового вымирания . Из крупных морских рептилий остались только морские черепахи , а из динозавров сохранились лишь мелкие пернатые тероподы в виде птиц . Конец «Эры рептилий» привел к «Эре млекопитающих». Несмотря на изменение формулировок, диверсификация рептилий продолжалась на протяжении всего кайнозоя. Сегодня чешуйчатые составляют большинство ныне живущих рептилий (более 90%). [28] На сегодняшний день насчитывается около 9766 видов рептилий. [28] по сравнению с 5400 видами млекопитающих, поэтому количество видов рептилий (без птиц) почти вдвое превышает количество млекопитающих.
Смена ролей [ править ]
После того, как мел-палеогеновое вымирание уничтожило всех нептичьих динозавров ( птиц обычно считают выжившими динозаврами) и несколько групп млекопитающих, плацентарные и сумчатые млекопитающие диверсифицировались во множество новых форм и экологических ниш на протяжении палеогеновой и неогеновой эпох. Некоторые из них достигали огромных размеров и почти такого же разнообразия, как когда-то динозавры. млекопитающих Тем не менее, мегафауна так и не достигла высот небоскребов некоторых зауроподов.
Тем не менее, крупные рептилии по-прежнему составляют важные компоненты мегафауны, такие как гигантские черепахи , [29] [30] крупные крокодилы и, более локально, крупные вараниды .
Четыре отряда Рептилий [ править ]
Testudines[editчерепахи
Тестудины, или черепахи, возможно, произошли от анаспид, но их точное происхождение неизвестно и активно обсуждается. Окаменелости датируются примерно 220 миллионами лет назад и имеют удивительно схожие характеристики. Эти первые черепахи сохраняют то же строение тела, что и все современные черепахи, и в основном являются травоядными, а некоторые питаются исключительно мелкими морскими организмами. Считается, что панцирь торговой марки произошел от продолжений позвоночника и расширенных ребер, которые слились вместе. Это подтверждается окаменелостью Odontochelys semitestacea , у которой неполный панцирь, берущий начало от ребер и позвоночника. У этого вида также были зубы на клюве, что еще больше подтверждает, что он является переходным ископаемым, хотя это утверждение до сих пор остается спорным. Этот панцирь эволюционировал для защиты от хищников, но также значительно замедляет наземные виды. Это привело к исчезновению многих видов в последнее время. Из-за того, что чужеродные виды превосходят их за пищу и неспособности спастись от людей, в этом порядке существует много видов, находящихся под угрозой исчезновения.
Сфенодонтия [ править ]
Сфенодонты возникли в середине триаса и теперь состоят из одного рода, tuatara , который включает в себя два исчезающих вида, обитающих в Новой Зеландии и на некоторых небольших прилегающих к ней островах. Их эволюционная история наполнена множеством видов. [31] Недавние палеогенетические открытия показывают, что туатары склонны к быстрому видообразованию.
Чешуйчатый [ править ]
Последний отряд рептилий — чешуйчатые. [32] различаются по подвижной квадратной кости (обеспечивающей движение верхней челюсти), роговой чешуе и гемипенам . Они происходят из ранней юры и состоят из трех подотрядов Lacertilia (парафилетические), Serpentes и Amphisbaenia . Хотя это самый новый отряд, чешуйчатые содержат больше видов, чем любой другой отряд рептилий. Чешуйчатые — монофилетическая группа, включенная вместе со Sphenodontia (например, tuataras ) в Lepidosauria . Последний надотряд вместе с некоторыми вымершими животными, такими как плезиозавры , составляют Lepidosauromorpha , сестринский инфракласс группы Archosauromorpha , в которую входят крокодилы, черепахи и птицы. Хотя чешуйчатые окаменелости впервые появляются в ранней юре , митохондриальная филогенетика предполагает, что они появились в поздней перми . Большинство эволюционных взаимоотношений внутри чешуйчатых еще не полностью выяснено, при этом наиболее проблематичны отношения змей с другими группами. Судя по морфологическим данным, ящерицы-игуаниды отделились от других чешуйчатых очень рано, но недавно молекулярная филогения , как на основе митохондриальной , так и на основе ядерной ДНК , не подтверждает это раннее расхождение. [33] быстрее, Поскольку у змей молекулярные часы чем у других чешуекрылых. [33] и есть немного ранних окаменелостей змей и предков змей, [34] трудно разрешить взаимоотношения змей с другими группами чешуйчатых.
Крокодил [ править ]
Первые организмы, показавшие сходные характеристики с крокодилами. [35] были Круротарси , появившиеся в раннем триасе 250 миллионов лет назад. 220 миллионов лет назад это быстро привело к возникновению клады Евзухий, которая в конечном итоге привела к отряду Крокодилов, первый из которых возник около 85 миллионов лет назад в позднем меловом периоде. Самые ранние ископаемые свидетельства существования евзухий относятся к роду Isisfordia . Ранние виды питались в основном рыбой и растительностью. Они жили на суше, у большинства были длинные ноги (по сравнению с современными крокодилами), а многие были двуногими. По мере увеличения диверсификации появилось множество высших хищников, все из которых сейчас вымерли. Современные крокодилы возникли благодаря специфическим эволюционным признакам. Полная потеря прямохождения была заменена в целом низкой четвероногой позицией на легкий и менее заметный вход в водоемы. Форма черепа/челюсти изменилась, чтобы обеспечить больший захват, а также направленные вверх ноздри и глаза. Мимикрия очевидна, поскольку спины всех крокодилов напоминают какое-то плавающее бревно, а их общая коричневая и зеленая цветовая гамма имитирует мох или дерево. Их хвост также принял форму весла, чтобы увеличить скорость плавания. Единственные оставшиеся группы [36] К этому отряду относятся аллигаторы , кайманы , крокодилы , гавиалы и ложные гавиалы .
Ссылки [ править ]
- ^ «Эволюция рептилий» . Свободные тексты по биологии . Свободные тексты. 15 августа 2021 г. Проверено 13 декабря 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Лорин, М.; Рейс, Р.Р. (1995). «Переоценка ранней филогении амниот». Зоологический журнал Линнеевского общества . 113 (2): 165–223. дои : 10.1111/j.1096-3642.1995.tb00932.x . S2CID 29661691 . ( абстрактный )
- ^ Патон, РЛ; Патон, ТР; Клак, Дж.А. (1999). «Скелет, похожий на амниоту, из раннего карбона Шотландии». Природа . 398 (6727): 508–513. Бибкод : 1999Natur.398..508P . дои : 10.1038/19071 . S2CID 204992355 .
- ^ Монастерский, Р. (1999) Из болот, Как ранние позвоночные животные закрепились на суше всеми 10 пальцами ног. Новости науки, Vol. 155, № 21, с. 328. статья
- ^ Глава 6: «Прогулка с ранними четвероногими: эволюция посткраниального скелета и филогенетическое сходство Temnospondyli (Vertebrata: Tetrapoda)». В : Кэт Поли (2006). « Посткраниальный скелет темноспондилов (Tetrapoda: temnospondyli) ». Кандидатская диссертация. Университет Ла Троб, Мельбурн.
- ^ Фалькон-Ланг, HJ; Бентон, MJ; Стимсон, М. (2007). «Экология ранних рептилий, выявленная по тропам Нижней Пенсильвании». Журнал Геологического общества . 164 (6): 1113–1118. дои : 10.1144/0016-76492007-015 . S2CID 140568921 .
- ^ «Самые ранние свидетельства существования рептилий» . Sflorg.com. 17 октября 2007 г. Архивировано из оригинала 16 июля 2011 г. Проверено 16 марта 2010 г.
- ^ Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 62. ИСБН 1-84028-152-9 .
- ^ Рута, М.; Коутс, Мичиган; Квик, DLJ (2003). «Возвращение к ранним взаимоотношениям четвероногих» (PDF) . Биологические обзоры . 78 (2): 251–345. дои : 10.1017/S1464793102006103 . ПМИД 12803423 . S2CID 31298396 . Архивировано из оригинала (PDF) 22 мая 2008 г. Проверено 23 января 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б Ромер, А.С. и Т.С. Парсонс. 1977. Тело позвоночного. 5-е изд. Сондерс, Филадельфия. (6-е изд. 1985 г.)
- ^ Хуан С. Сиснерос; Росс Дамиани; Цезарь Шульц; Атила да Роза; Сибеле Шванке; Леопольдо В. Нето; Педро Л. П. Аурелио (2004). «Проколофоноидная рептилия с височными отверстиями из среднего триаса Бразилии» . Труды Королевского общества B: Биологические науки . 271 (1547): 1541–1546. дои : 10.1098/rspb.2004.2748 . ПМК 1691751 . ПМИД 15306328 .
- ^ Линда А. Цудзи; Йоханнес Мюллер (2009). «Собираем историю Parareptilia: филогения, разнообразие и новое определение клады» . Ископаемый рекорд . 12 (1): 71–81. дои : 10.1002/mmng.200800011 .
- ↑ Перейти обратно: Перейти обратно: а б Грасиела Пиньейро; Хорхе Фериголо; Алехандро Рамос; Мишель Лорен (2012). «Краниальная морфология раннепермских мезозавридов Mesosaurus tenuidens и переоценка эволюции нижних височных окон». Comptes Рендус Палевол . 11 (5): 379–391. дои : 10.1016/j.crpv.2012.02.001 .
- ^ Ван Тунинен, М.; Хэдли, Э.А. (2004). «Ошибка в оценке скорости и времени, полученная на основе записей окаменелостей ранних амниот и птичьих молекулярных часов» (PDF) . Журнал молекулярной биологии . 59 (2): 267–276. Бибкод : 2004JMolE..59..267В . дои : 10.1007/s00239-004-2624-9 . PMID 15486700 . S2CID 25065918 . Архивировано из оригинала (PDF) 28 июня 2011 г.
- ^ Бентон, MJ (2000). Палеонтология позвоночных (2-е изд.). Blackwell Science Ltd. Лондон: ISBN 0-632-05614-2 . , 3-е изд. 2004 г. ISBN 0-632-05637-1
- ^ Риппель О, ДеБрага М (1996). «Черепахи как диапсидные рептилии» (PDF) . Природа . 384 (6608): 453–5. Бибкод : 1996Natur.384..453R . дои : 10.1038/384453a0 . S2CID 4264378 .
- ^ Зардоя, Р.; Мейер, А. (1998). «Полный митохондриальный геном предполагает сходство черепах с диапсидом» (PDF) . Proc Natl Acad Sci США . 95 (24): 14226–14231. Бибкод : 1998PNAS...9514226Z . дои : 10.1073/pnas.95.24.14226 . ISSN 0027-8424 . ПМК 24355 . ПМИД 9826682 .
- ^ Ивабе, Н.; Хара, Ю.; Кумазава, Ю.; Сибамото, К.; Сайто, Ю.; Мията, Т.; Като, К. (29 декабря 2004 г.). «Сестринская групповая связь черепах с кладой птиц и крокодилов, выявленная с помощью белков, кодируемых ядерной ДНК» . Молекулярная биология и эволюция . 22 (4): 810–813. дои : 10.1093/molbev/msi075 . ПМИД 15625185 .
- ^ Роос, Йонас; Аггарвал, Рамеш К.; Янке, Аксель (ноябрь 2007 г.). «Расширенный митогеномный филогенетический анализ дает новое представление об эволюции крокодилов и их выживании на границе мелового и третичного периодов». Молекулярная филогенетика и эволюция . 45 (2): 663–673. дои : 10.1016/j.ympev.2007.06.018 . ПМИД 17719245 .
- ^ Гийетт, Ю.; Браун, Эль; Гийетт, Эл-Джей-младший; Игучи, Т. (17 марта 2010 г.). «От филогеномики рептилий к геномам рептилий: анализ протоонкогенов c-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. дои : 10.1159/000297715 . ПМИД 20234127 . S2CID 12116018 .
- ↑ Перейти обратно: Перейти обратно: а б с Кольбер Э.Х. и Моралес М. (2001): «Эволюция позвоночных животных Кольбера: история позвоночных животных во времени» . 4-е издание. John Wiley & Sons, Inc., Нью-Йорк — ISBN 978-0-471-38461-8 .
- ^ Готье Ж.А. (1994): Разнообразие амниот . В: Д. Р. Протеро и Р. М. Шох (ред.) Основные особенности эволюции позвоночных: 129–159. Ноксвилл, Теннесси: Палеонтологическое общество.
- ^ Джон В. Мерк (1997). «Филогенетический анализ эвриапсидных рептилий». Журнал палеонтологии позвоночных . 17 (Дополнение к 3): 65А. дои : 10.1080/02724634.1997.10011028 .
- ^ Шон Модесто; Роберт Рейс; Дайан Скотт (2011). «Неодиапсидная рептилия из нижней перми Оклахомы». Программа и тезисы 71-го ежегодного собрания Общества палеонтологии позвоночных : 160.
- ^ «GEOL 331/BSCI 333 Палеонтология четвероногих» . Геол.umd.edu .
- ^ Рёсуке Мотани; Начио Минура; Тацуро Андо (1998). «Отношения ихтиозавров, освещенные новыми примитивными скелетами из Японии». Природа . 393 (6682): 255–257. Бибкод : 1998Natur.393..255M . дои : 10.1038/30473 . S2CID 4416186 .
- ^ Фил Сентер (2007). «Новый взгляд на филогению целурозаврии (Dinosauria: Theropoda)». Журнал систематической палеонтологии . 5 (4): 429–463. дои : 10.1017/S1477201907002143 . S2CID 83726237 .
- ↑ Перейти обратно: Перейти обратно: а б «База данных рептилий» . Проверено 1 февраля 2012 года .
- ^ Хансен, DM; Донлан, CJ; Гриффитс, CJ; Кэмпбелл, Кей Джей (апрель 2010 г.). «Экологическая история и скрытый природоохранный потенциал: крупные и гигантские черепахи как модель для замены таксонов» (PDF) . Экография . 33 (2). Уайли : 272–284. дои : 10.1111/j.1600-0587.2010.06305.x . Архивировано из оригинала (PDF) 24 июля 2011 г. Проверено 26 февраля 2011 г.
- ^ Сион, Алабама; Тонни, EP; Сойбельзон, Л. (2003). «Разорванный зигзаг: позднекайнозойское вымирание крупных млекопитающих и черепах в Южной Америке» . Преподобный Муз. Аргентино Сиенс. Нат . НС 5 (1): 1–19. дои : 10.22179/REVMACN.5.26 . ISSN 1514-5158 .
- ^ «Обнаружена древняя ящерицеобразная рептилия» . Архивировано из оригинала 25 сентября 2017 г. Проверено 18 апреля 2015 г.
- ^ Ридер, Тод В.; Таунсенд, Тед М.; Малкахи, Дэниел Г.; Нунан, Брайс П.; Вуд, Перри Л.; Сайты, Джек В.; Винс, Джон Дж. (2015). «Комплексный анализ разрешает конфликты по поводу филогении чешуйчатых рептилий и выявляет неожиданные места размещения ископаемых таксонов» . ПЛОС ОДИН . 10 (3): e0118199. Бибкод : 2015PLoSO..1018199R . дои : 10.1371/journal.pone.0118199 . ПМЦ 4372529 . ПМИД 25803280 .
- ↑ Перейти обратно: Перейти обратно: а б Кумадзава, Ёсинори (2007). «Митохондриальные геномы основных семейств ящериц предполагают их филогенетические связи и древнюю радиацию». Джин . 388 (1–2): 19–26. дои : 10.1016/j.gene.2006.09.026 . ПМИД 17118581 .
- ^ «Ящерицы и змеи живы!» . Американский музей естественной истории. Архивировано из оригинала 28 января 2008 г. Проверено 25 декабря 2007 г.
- ^ «Крокодилы: естественная история и охрана — крокодилы, кайманы, аллигаторы, гавиалы» . Крокодил.com . Проверено 12 апреля 2022 г.
- ^ «Аллигатор против крокодила – объяснение различий (с видео и изображениями)» . Диффен.com .
Основные существующие рептилий клады |
|---|
| Базы данных органов управления : Национальные |
|---|