Диапы
| Диапсидные рептилии Временной диапазон: Пенсильвания – настоящее время , | |
|---|---|
 | |
| Схема черепа ареосцелидии Petrolacosauruskansensis | |
 | |
| Нильский крокодил ( Crocodylus niloticus ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Римская ткань |
| Клэйд : | Диапсида Осборн , 1903 год. |
| Подгруппы | |
| |
Диапсиды («две арки») — клада зауропсидов , отличающаяся от более примитивных эврептилий наличием двух отверстий, известных как височные окна , на каждой стороне черепа . Самые ранние традиционно идентифицированные диапсиды, ареосцелидии , появились около трехсот миллионов лет назад, в конце каменноугольного периода. [1] Все диапсиды, кроме наиболее примитивных в кладе Araeoscelidia, часто относят к кладе Neodiapsida . Диапсиды чрезвычайно разнообразны и включают птиц и все современные группы рептилий, включая черепах , которые исторически считались вне группы. [2] Все современные рептилии и птицы относятся к субкладу неодиапсидов Sauria . Хотя некоторые диапсиды потеряли либо одно отверстие (ящерицы), либо оба отверстия (змеи и черепахи), либо имеют сильно перестроенный череп (современные птицы), их по-прежнему классифицируют как диапсиды на основании их происхождения. не менее 17 084 видов диапсидных животных: 9 159 видов птиц, Сохранилось [3] и 7925 змей, ящериц, туатара , черепах и крокодилов. [4]
Характеристики
[ редактировать ]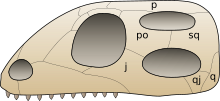
Название «Диапсида» означает «две арки», и диапсиды традиционно классифицируются на основе двух наследственных отверстий черепа ( височных окон ) сзади над и под глазом. Такое расположение позволяет прикрепить более крупные и сильные мышцы челюсти и позволяет челюсти открываться более широко. Более неясной наследственной характеристикой является относительно длинная кость нижней части руки ( лучевая кость ) по сравнению с костью верхнего плеча ( плечевая кость ).
Базальные неящероподобные неодиаспиды изначально напоминали ящериц, но базальные неящероподобные неодиапсиды включают водные/амфибийные таксоны ( клаудиозавр и некоторые тангасавриды ). [5] ящерица планирующая Weigeltisauridae , [6] а также триасовых хамелеоноподобных дрепанозавров . [7]
Классификация
[ редактировать ]Первоначально диапсиды были классифицированы как один из четырех подклассов класса Reptilia , каждый из которых основывался на количестве и расположении отверстий в черепе. Другими тремя подклассами были Synapsida (одно отверстие в нижней части черепа, для «рептилий, похожих на млекопитающих»), Anapsida (без отверстия в черепе, включая черепах и их родственников) и Euryapsida (одно отверстие в верхней части черепа, включая многие доисторические подклассы). морские рептилии). С появлением филогенетической номенклатуры эта система классификации была сильно модифицирована. Сегодня синапсиды часто не считаются настоящими рептилиями, в то время как Euryapsida оказались неестественным скоплением диапсидов, потерявших одно из отверстий черепа. Генетические исследования и открытие триасовых Pappochelys показали, что то же самое происходит и с черепахами, которые на самом деле представляют собой сильно модифицированные диапсиды. В филогенетических системах к этой группе относят и птиц (потомков традиционных диапсидных рептилий).
Некоторые современные исследования взаимоотношений рептилий предпочитают использовать название «диапсид» для обозначения коронной группы всех современных диапсидных рептилий, но не их вымерших родственников. Однако многие исследователи также отдают предпочтение более традиционному определению, включающему доисторических ареосцелид . В 1991 году Лорин определил Diapsida как кладу , «самого недавнего общего предка ареосцелидий, лепидозавров и архозавров , а также всех их потомков». [8]
определение кладе Neodiapsida дал Филогенетическое Лаурин в 1991 году. Он определил ее как ветвящуюся кладу , содержащую всех животных, более тесно связанных с « Younginiformes » (позже, более конкретно, измененными на Youngina capensis ), чем с Petrolacosaurus (представляющим Araeoscelidia ). . [9] Самые ранние известные неодиапсиды, такие как Оровенатор, известны из ранней перми, около 290 миллионов лет назад. [10]
Все генетические исследования подтвердили гипотезу о том, что черепахи являются диапсидными рептилиями; некоторые поместили черепах внутри архозавроморф, [11] [12] или, что чаще, как сестринскую группу ныне живущим архозаврам. [13] [14] [15] [16]
Современные рептилии и птицы отнесены к субкладу неодиапсидов Sauria , определяемому как последний общий предок Lepidosauria (который включает ящериц, змей и туатару) и Archosauria (который включает, среди прочего, крокодилов и динозавров, включая птиц). [17]
Кладистический анализ Лорина и Пиньейро (2017) обнаруживает Parareptilia как часть Diapsida, при этом парейазавры, черепахи, миллереттиды и проколофоноиды обнаруживаются как более производные, чем базальная диапсида Younginia . [18] Исследование Дэвида П. Форда и Роджера Б. Дж. Бенсона, проведенное в 2020 году, также выявило, что Parareptilia глубоко укоренена в Diapsida как сестринская группа Neodiapsida . Однако это исключает мезозавров , которые оказались базальными среди зауропсидов. [19] Другие недавние исследования показали, что более традиционное расположение парарептилий находится за пределами Диапсиды. [17]
Положение высокоразвитых мезозойских групп морских рептилий Thalattosauria , Ichthyosauromorpha и Sauropterygia внутри Neodiapsida неясно, и они могут находиться в пределах Sauria. [17]
Отношения
[ редактировать ]Ниже приведены кладограммы, показывающие взаимоотношения основных групп диапсид.
Кладограмма по Bickelmann et al. , 2009 г. [20] и Рейс и др. , 2011: [21]
| ||||||||||||||||||||||
В приведенной ниже кладограмме Ли (2013) использовалась комбинация генетических (молекулярных) и ископаемых (морфологических) данных. [22]
| Диапсида |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Эта вторая кладограмма основана на исследовании Притчарда и Несбитта, проведенного в 2017 году. [23]
| неодиапсид |
| |||||||||||||||||||||||||||||||||

Следующая кладограмма была найдена Simões et al . (2022): [17]
| Неорептилия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Диапсида» . ucmp.berkeley.edu .
- ^ Шох, Райнер Р.; Сьюс, Ханс-Дитер (2016). «Диапсидное происхождение черепах». Зоология . 119 (3): 159–161. Бибкод : 2016Zool..119..159S . дои : 10.1016/j.zool.2016.01.004 . ПМИД 26934902 .
- ^ Барроуклаф, Джордж Ф.; Кракрафт, Джоэл; Клика, Джон; и Зинк, Роберт М. (23 ноября 2016 г.). Грин, Энди Дж. (ред.). «Сколько существует видов птиц и почему это важно?» . ПЛОС ОДИН . 11 (11): e0166307. Бибкод : 2016PLoSO..1166307B . дои : 10.1371/journal.pone.0166307 . ПМК 5120813 . ПМИД 27880775 .
- ^ Ридер, Тод В.; Таунсенд, Тед М.; Малкахи, Дэниел Г.; Нунан, Брайс П.; Вуд, Перри Л. младший; Сайты, Джек В. младший; и Винс, Джон Дж. (2015). Уилф, Питер (ред.). «Комплексный анализ разрешает конфликты по поводу филогении чешуйчатых рептилий и выявляет неожиданные места размещения ископаемых таксонов» . ПЛОС ОДИН . 10 (3): e0118199. Бибкод : 2015PLoSO..1018199R . дои : 10.1371/journal.pone.0118199 . ПМЦ 4372529 . ПМИД 25803280 .
- ^ Нуньес Демарко, Пабло; Менегель, Мелитта; Лорен, Мишель; Пиньейру, Грасиела (27 июля 2018 г.). «Был ли мезозавр полностью водной рептилией?» . Границы экологии и эволюции . 6 : 109. дои : 10.3389/fevo.2018.00109 . hdl : 20.500.12008/30631 . ISSN 2296-701X .
- ^ Причард, Адам К.; Сьюс, Ханс-Дитер; Скотт, Дайан; Рейс, Роберт Р. (20 мая 2021 г.). «Остеология, взаимоотношения и функциональная морфология Weigeltisaurus jaekeli (Diapsida, Weigeltisauridae) на основе полного скелета из верхнепермского купфершифера в Германии» . ПерДж . 9 : е11413. дои : 10.7717/peerj.11413 . ISSN 2167-8359 . ПМЦ 8141288 . ПМИД 34055483 .
- ^ Причард, Адам К.; Несбитт, Стерлинг Дж. (октябрь 2017 г.). «Птичье череп у триасовой диапсидной рептилии увеличивает гетерогенность морфологической и филогенетической радиации диапсид» . Королевское общество открытой науки . 4 (10): 170499. Бибкод : 2017RSOS....470499P . дои : 10.1098/rsos.170499 . ISSN 2054-5703 . ПМК 5666248 . ПМИД 29134065 .
- ^ Бентон, М.Дж., Донохью, ПК, Ашер, Р.Дж., Фридман, М., Ниар, Т.Дж., и Винтер, Дж. (2015). «Ограничения на временные рамки истории эволюции животных». Палеонтология Электроника , 18.1.1FC; 1-106; palaeo-electronica.org/content/fc-1
- ^ Рейс, Роберт Р.; Модесто, Шон П.; Скотт, Дайан М. (22 декабря 2011 г.). «Новая раннепермская рептилия и ее значение в ранней эволюции диапсид» . Труды Королевского общества B: Биологические науки . 278 (1725): 3731–3737. дои : 10.1098/rspb.2011.0439 . ПМЦ 3203498 . ПМИД 21525061 .
- ^ Рейс, Роберт Р.; Модесто, Шон П.; Скотт, Дайан М. (22 декабря 2011 г.). «Новая раннепермская рептилия и ее значение в эволюции ранних диапсид» . Труды Королевского общества B: Биологические науки . 278 (1725): 3731–3737. дои : 10.1098/rspb.2011.0439 . ISSN 0962-8452 . ПМЦ 3203498 . ПМИД 21525061 .
- ^ Ли, MSY (2013). «Происхождение черепах: выводы из филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. дои : 10.1111/jeb.12268 . ПМИД 24256520 . S2CID 2106400 .
- ^ Маннен, Хидеюки; Ли, Стивен С.-Л. (октябрь 1999 г.). «Молекулярные доказательства существования клада черепах». Молекулярная филогенетика и эволюция . 13 (1): 144–148. дои : 10.1006/mpev.1999.0640 . PMID 10508547 .
- ^ Зардоя, Р.; Мейер, А. (1998). «Полный митохондриальный геном предполагает сходство черепах с диапсидом» . Proc Natl Acad Sci США . 95 (24): 14226–14231. Бибкод : 1998PNAS...9514226Z . дои : 10.1073/pnas.95.24.14226 . ISSN 0027-8424 . ПМК 24355 . ПМИД 9826682 .
- ^ Ивабе, Н.; Хара, Ю.; Кумазава, Ю.; Сибамото, К.; Сайто, Ю.; Мията, Т.; Като, К. (29 декабря 2004 г.). «Сестринская группа черепах к кладе птиц и крокодилов, выявленная с помощью белков, кодируемых ядерной ДНК» . Молекулярная биология и эволюция . 22 (4): 810–813. дои : 10.1093/molbev/msi075 . ПМИД 15625185 .
- ^ Роос, Йонас; Аггарвал, Рамеш К.; Янке, Аксель (ноябрь 2007 г.). «Расширенный митогеномный филогенетический анализ дает новое понимание эволюции крокодилов и их выживания на границе мелового и третичного периодов». Молекулярная филогенетика и эволюция . 45 (2): 663–673. дои : 10.1016/j.ympev.2007.06.018 . ПМИД 17719245 .
- ^ Катсу, Ю.; Браун, Эль; Гийетт, Эл-Джей-младший; Игучи, Т. (17 марта 2010 г.). «От филогеномики рептилий к геномам рептилий: анализ протоонкогенов c-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. дои : 10.1159/000297715 . ПМИД 20234127 . S2CID 12116018 .
- ^ Перейти обратно: а б с д Симойнс, Тьяго Р.; Каммерер, Кристиан Ф.; Колдуэлл, Майкл В.; и Пирс, Стефани Э. (2022). «Последовательные климатические кризисы в глубоком прошлом привели к ранней эволюции и радиации рептилий» . Достижения науки . 8 (33): eabq1898. Бибкод : 2022SciA....8.1898S . дои : 10.1126/sciadv.abq1898 . ПМЦ 9390993 . ПМИД 35984885 .
- ^ Лорен, Мишель; Пинейро, Грасиела Х. (2017). «Переоценка таксономического положения мезозавров и удивительная филогения ранних амниот» (PDF) . Границы в науках о Земле . 5 : 88. Бибкод : 2017FrEaS...5...88L . дои : 10.3389/feart.2017.00088 . S2CID 32426159 .
- ^ Форд Д.П., Бенсон Р.Б. (январь 2020 г.). «Филогения ранних амниот и сходство Parareptilia и Varanopidae» . Экология и эволюция природы . 4 (1): 57–65. дои : 10.1038/s41559-019-1047-3 . ПМИД 31900445 . S2CID 209673326 .
- ^ Бикельманн, Констанца; Мюллер, Йоханнес; и Рейс, Роберт Р. (2009). «Загадочный диапсид Acerosodontosaurus piveeaui (Reptilia: Neodiapsida) из верхней перми Мадагаскара и парафилия молодых рептилий». Канадский журнал наук о Земле . 49 (9): 651–661. Бибкод : 2009CaJES..46..651S . дои : 10.1139/E09-038 .
- ^ Рейс, Роберт Р.; Модесто, Шон П.; и Скотт, Дайан М. (2011). «Новая раннепермская рептилия и ее значение в эволюции ранних диапсид» . Труды Королевского общества Б. 278 (1725): 3731–7. дои : 10.1098/rspb.2011.0439 . ПМЦ 3203498 . ПМИД 21525061 .
- ^ Ли, MSY (2013). «Происхождение черепах: выводы из филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. дои : 10.1111/jeb.12268 . ПМИД 24256520 . S2CID 2106400 .
- ^ Причард, Адам К.; Несбитт, Стерлинг Дж. (11 октября 2017 г.). «Птичье череп у триасовой диапсидной рептилии увеличивает гетерогенность морфологической и филогенетической радиации диапсид» . Королевское общество открытой науки . 4 (10): 170499. Бибкод : 2017RSOS....470499P . дои : 10.1098/rsos.170499 . ISSN 2054-5703 . ПМК 5666248 . ПМИД 29134065 .
Внешние ссылки
[ редактировать ] Данные, связанные с Диапсидом , на Wikispecies
Данные, связанные с Диапсидом , на Wikispecies - Диапсида . Мишель Лорен и Жак А. Готье. Веб-проект «Древо жизни» . 22 июня 2000 г.





Основные существующие рептилий клады |
|---|