Хористодера
| Хористодера | |
|---|---|
 | |
| Скелет Philydrosaurus proselius выставлен в Национальном музее естествознания , Тайвань. | |
 | |
| Скелет икехозавра на выставке в Национальном музее естествознания, Тайвань. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | неодиапсид |
| Заказ: | † Хористодера Коуп , 1876 г. |
| Подгруппы | |
| |
Хористодера (от греческого χωριστός chōristos + δέρη dérē , «отдельная шея»). [2] ) — вымерший отряд полуводных (от 168 до 20 или, возможно , диапсидных рептилий , существовавший от средней юры или, возможно , триаса до миоцена 11,6 миллионов лет назад). Хористодеры морфологически разнообразны, наиболее известными представителями являются крокодилоподобные неохористодеры , такие как Champsosaurus . Другие хористодеры имели морфологию ящерицы или длинной шеи. Choristoderes, по-видимому, были распространены в Северном полушарии и были обнаружены в Северной Америке , Азии и Европе , а также, возможно, в Северной Африке . Хористодеры обычно считаются производными неодиапсидов , которые являются близкими родственниками или членами Sauria .
История открытия
[ редактировать ]
в 1876 году первоначально как подотряд Rhynchocephalia Дринкером Choristodera был выведен Эдвардом Коупом содержания Champsosaurus , который был описан из позднемеловых слоев Монтаны в той же статье. для Коупом [3] [4] Год спустя, в 1877 году, симоедозавр был описан Полем Жерве из верхнепалеоценовых отложений в Серне , недалеко от Реймса , Франция. Они оставались единственными признанными хористодерами более века, пока в конце 20 века не были описаны новые таксоны. [5] Начиная с конца 1970-х годов, дополнительные таксоны были описаны советско-монгольскими группами из нижнемеловых отложений Монголии. В исследованиях 1989–1991 годов Сьюзен Э. Эванс описала новый материал Cteniogenys из средней юры Великобритании. Этот род был впервые описан Чарльзом У. Гилмором в 1928 году из поздней юры на западе Соединенных Штатов и ранее был загадочным. Исследования показали, что это небольшая ящерицеобразная хористодера, отличающаяся от ранее известных крокодилоподобных форм. [6]
Описание
[ редактировать ]
Хористодеры существенно различаются по размеру: самые маленькие роды, такие как Cteniogenys и Lazarussuchus, имели длину всего около 30 см (12 дюймов), а самый крупный известный хористодер, Kosmodraco dakotensis , по оценкам, имел общую длину около 5 м (16 футов). . [5] [1] Неохористодеры , такие как Champsosaurus, являются самой известной группой Choristodera. Они напоминали современных крокодилов , особенно гавиалов . Череп . этих животных имеет длинную тонкую морду с маленькими острыми зубами конической формы Другие хористодеры называются «ненеохористодерами», и они в основном представляют собой небольшие ящерицеподобные формы, хотя Шокава , Хурендухозавр и Гифалозавр обладают длинными шеями, похожими на плезиозавров . Группа «неохористодер» является парафилетической (не содержит всех потомков общего предка), поскольку форма тела, напоминающая ящерицу, представляет наследственную морфологию группы. [5]
Скелетная анатомия
[ редактировать ]
По мнению Мацумото и коллег (2019), хористодер объединяет наличие 9 синапоморфий (общих признаков, характерных для группы), в том числе срединного контакта удлиненных предлобных костей черепа, отделяющего носовые кости от лобных , спинных фланец верхней челюсти изогнут медиально (к средней линии тела), теменное отверстие отсутствует, чешуйчатые кости расширены позади ( кзади ) затылочного мыщелка , зубы конические, субтекодонтные ( расположены в неглубоких лунках) зубные кости тонкие, с удлиненными бороздками, идущими вдоль лабиальной (наружной) поверхности кости, имеются дополнительные крестцовые позвонки , на позвонках имеются расширенные «позвоночные столики», поверхности обоих концов центров позвонков плоские ( амфиплатян). [7] Все известные хористодеры обладают или предположительно обладают новой костью черепа, не встречающейся у других рептилий, называемой «неоморфной костью» или неоморфом, которая является компонентом дерматокраниума . [8] [9] Наследственный череп хористодер имел удлиненные верхние и нижние височные отверстия (отверстия черепа за глазницей), они значительно расширены у неохористодер, особенно у хампсозавра , что придает черепу сердцевидный (сердечкообразный) вид, если смотреть сверху. . [8] У многих «неохористодер» нижние височные окна вторично закрываются. [10] Хористодеры обладали гастралиями (реберными костями, расположенными в брюшной полости), как у туатары и крокодилов . [11]
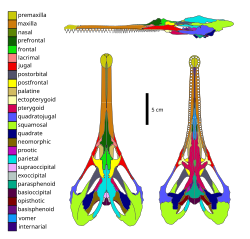 Схема черепа Champsosaurus lindoei , неохористодерана
Схема черепа Champsosaurus lindoei , неохористодерана Череп Lazarussuchus , аллохористодера
Череп Lazarussuchus , аллохористодера


Внутренняя анатомия черепа
[ редактировать ]Внутренняя анатомия черепа хористодер известна только у Champsosaurus. Champsosaurus Мозговая коробка плохо . окостеневшая в передней части черепа (в передней части), но хорошо окостеневшая в задней (задней), как и у других диапсид Краниальная эндокаста (пространство, занимаемое мозгом в своде черепа ) пропорционально узкая как в латеральной, так и в дорсовентральной оси, с увеличенным шишковидным телом и обонятельными луковицами . Оптические доли и флоккулы небольшого размера, что указывает в лучшем случае на среднюю способность зрения. Обонятельные камеры носовых ходов и обонятельные ножки черепной коробки достаточно велики, что указывает на то, что хампзозавр , вероятно, обладал хорошими обонятельными способностями (обонянием). В носовых ходах отсутствуют костные раковины . Полукружные каналы внутреннего уха наиболее похожи на таковые у других водных рептилий. Расширение саккулюса указывает на то, что хампзозавр , вероятно, обладал повышенной чувствительностью к низкочастотным звукам и вибрациям. [12]
Зубной ряд
[ редактировать ]У большинства хористодер довольно простые недифференцированные ( гомодонты ) зубы с полосатой эмалью , покрывающей коронку зуба, но не основание. У Neochoristoderes зубы полностью покрыты полосатой эмалью с эмалевой складкой у основания, лабиолингвально сжатыми и крючковатыми, за исключением Ikechosaurus , у которого зубы все еще довольно простые, если не считать начала эмалевой складки. Имплантация зубов субтекодонтная, при этом зубы замещаются путем эрозии ямки на лингвальной (стороне зуба, обращенной к языку) поверхности основания зуба. Среди neochoristoderes наблюдается некоторая дифференциация зубов: передние зубы острее и тоньше задних. Хористодеры сохраняют небные зубы (зубы присутствуют на костях нёба). В отличие от большинства групп диапсидов, у которых небные зубы редуцированы или полностью утрачены, небные зубы у хористодер широко развиты, что указывает на манипуляции с пищей во рту, вероятно, в сочетании с языком. У большинства хористодер имеются продольные ряды небных зубов. крыловидная , небная и сошник , а также ряд на фланце крыловидной кости. У некоторых неохористодер небные зубные ряды преобразованы в зубные батареи на приподнятых платформах. Морфология небных зубов идентична морфологии маргинальных зубов неохористодер, а замена небных зубов почти идентична замене маргинальных зубов. [13]
Кожа
[ редактировать ]

Исключительно сохранившийся экземпляр Monjurosuchus сохранил складчатую кожу, что указывает на то, что при жизни он, вероятно, был тонким и мягким. Сохранившиеся чешуи мелкие, перекрывающиеся, на вентральной нижней стороне тела они меньше, чем на дорсальной поверхности. Вдоль спины (верхней средней линии) тела проходит двойной ряд более крупных яйцевидных чешуек. Окаменелость также сохранила перепончатые лапы . [14] Гифалозавр был покрыт чешуей различной формы, в зависимости от их положения на теле, по крайней мере с одним, а возможно, и с несколькими рядами больших овоидных чешуек, спускающихся по бокам туловища и хвоста. На ступнях имеются следы перепонок, а на хвосте, вероятно, были дополнительные ткани сверху и снизу, что позволяло использовать его в качестве плавника для продвижения гифалозавра по воде. [11] [15] отпечатки кожи у Champsosaurus Известны также , они состоят из мелких (0,6-0,1 мм) пустулатных и ромбовидных чешуек, причем наиболее крупные чешуйки располагаются на боковых сторонах тела, уменьшаются в размерах дорсально, остеодермы отсутствуют. [16] Экземпляр Lazarussuchus из Мената сохраняет некоторые остатки мягких тканей, но не имеет чешуи, что указывает на то, что задняя стопа (pes) не имела перепонок, а над хвостовыми позвонками хвоста имеется темная окрашенная область с зубчатым краем, что наводит на мысль о гребень похож на тот, что встречается у некоторых ныне живущих рептилий, таких как туатара, ящерицы и крокодилы. [17]
Палеобиология
[ редактировать ]Хористодеры встречаются исключительно в пресноводных отложениях, часто связаны с черепахами , рыбами, лягушками , саламандрами и крокодилообразными . Судя по всему, они были почти исключительно обнаружены в теплом умеренном климате , при этом ареал неохористодер простирался до высоких слоев канадской Арктики во время коньякско - сантонских стадий позднего мела (~ 89-83 миллиона лет назад), времени чрезвычайно теплого периода. Из-за морфологического сходства хористодер и крокодилиформ часто предполагалось, что они существуют в конкуренции. Однако «неохористодеры» были меньше взрослых водных крокодилообразных и с большей вероятностью конкурировали с другими таксонами. Для более похожих на крокодила неохористодер, по-видимому, произошла дифференциация ниш : гариалоподобные неохористодеры встречаются в сочетании с тупыми мордами крокодилообразных, но не в ассоциации с узкорылыми формами. [5]
Диета
[ редактировать ]Предполагается, что неохористодераны были рыбоядными . [16] В частности, считается, что шампзозавр питался, как современные гавиалы, поворачивая голову в сторону, чтобы ловить отдельных рыб с косяков, в то время как симоедозавр , как полагают, был более универсальным, способным ловить как водную, так и наземную добычу. [18] Было высказано предположение, что Cteniogenys и Lazarussuchus питались беспозвоночными. [6] В сохранившемся содержимом кишечника экземпляра Monjurosuchus обнаружены фрагменты кутикулы членистоногих . [14] Еще один экземпляр Monjurosuchus был найден с сохранившимися черепами семи молодых особей в брюшной полости. Было предложено представить это как свидетельство каннибализма . [19] Однако это предложение подверглось критике со стороны других авторов, которые предполагают, что они, скорее всего, представляют собой эмбрионы на поздних стадиях. [20] У экземпляра гифалозавра был обнаружен небольшой реберный скелет в брюшной полости, что позволяет предположить, что он по крайней мере время от времени поедал добычу позвоночных. [11]
Воспроизведение
[ редактировать ]Образец Hyphalosaurus baitaigouensis был обнаружен с 18 полностью развитыми эмбрионами в теле матери, что позволяет предположить, что они были живородящими . [21] но другой экземпляр показывает, что Hyphalosaurus baitaigouensis также обладал яйцами с мягкой скорлупой, похожими на яйца лепидозавров . [22] Возможное объяснение этого состоит в том, что гифалозавр был яйцеживородящим , причем яйца с тонкой скорлупой вылуплялись сразу после того, как они были отложены, предположительно на суше. [23] хотя также было высказано предположение, что этот вид использовал как живородящие, так и яйцеродящие способы размножения. [20] Эмбрион икехозавра был обнаружен сохранившимся внутри слабо минерализованного яйца с пергаментной скорлупой, что позволяет предположить, что икехозавр был яйцекладущим и откладывал яйца на суше. [20] Monjuruosuchus были живородящими. Предполагается, что [20] У Champsosaurus было высказано предположение, что взрослые самки могли выползать на берег и откладывать яйца на суше, тогда как самцы и молодые особи, по-видимому, неспособны на это, основываясь на предположительно половом диморфном сращении крестцовых позвонков и наличии более крепких костей конечностей у Champsosaurus. предполагаемые самки. [24] Скелет Филидрозавра был обнаружен вместе с связанными с ним молодыми особями после вылупления, что позволяет предположить, что они занимались родительской заботой после вылупления . [23]
Треки
[ редактировать ]Следы из раннего мела ( альба ) Южной Кореи, получившие ихнотаксона название Novapes ulsanensis , были отнесены к choristoderans на основании сходства сохранившихся пятипалых следов с морфологией стопы Monjurosuchus . В следах сохранились следы перепонок между пальцами. Основываясь на расстоянии между отпечатками, авторы исследования предположили, что хористодеры могут « высоко ходить », как современные крокодилы. [25] Следы, приписываемые неохористодерам, получившим название Champsosaurichnus parfeti, также были обнаружены в позднемеловой формации Ларами в Соединенных Штатах, хотя присутствуют только два отпечатка, и невозможно различить кисть (передняя часть стопы) и стопу (задняя часть стопы). [26]
Классификация и филогения
[ редактировать ]
Внутренняя систематика
[ редактировать ]Исторически внутренняя филогенетика Choristoderes была неясной: неохористодеры были обнаружены как хорошо поддерживаемая клада, но взаимоотношения «ненеохористодер» плохо выяснены. [7] Однако в течение 2010-х годов «ненеохористодеры» из раннего мела Азии (за исключением Heishanosaurus ) наряду с Lazarussuchus из кайнозоя Европы были обнаружены (при слабой поддержке) как принадлежащие к монофилетической кладе, которые были неофициально названы «Allochoristoderes» Донга и его коллег в 2020 году, характеризующиеся общей чертой полностью закрытых нижних височных окон, при этом Cteniogenys из средней-поздней юры Европы и Северной Америки последовательно обнаруживаются как самые базальные хористодеры. [10] Длинношеие «неохористодеры» Шокава и Гифалозавр часто обнаруживались как клада, которую Гао и Фокс в 2005 году назвали Hyphalosauridae. [27] Находка более полного материала ранее фрагментарного хурендухозавра показывает, что он также имел длинную шею и также был обнаружен в составе клады. [28]
Филогения на основе анализа Донга и коллег (2020): [10]
| Хористодера |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Отношения с другими рептилиями
[ редактировать ]Хористодеры повсеместно признаны членами Neodiapsida , но их точное место в кладе неясно из-за сочетания примитивных и производных особенностей, а также длинной призрачной линии (отсутствия летописи окаменелостей) после их отделения от других рептилий. [29] Первоначально его поместили в ринхоцефалию, но позже Коуп предложил поместить его в лацертиллу из-за формы шейных позвонков. Луи Долло в 1891 году вернул хористодеру к ринхоцефалиям, но в 1893 году предположил близкое родство с парейазавром . Альфред Ромер в публикациях 1956 и 1968 годов поместил Choristodera в парафилетическую или полифилетическую группу « Eosuchia », описав их как «ответвление основной группы эозухий», классификация, которая получила широкое признание. Однако использование компьютерной кладистики в 1980-х годах продемонстрировало немонофилию «Eosuchia», что снова сделало классификацию хористодер неопределённой. [30] Последующие исследования либо предполагали отнесение к архозавроморфам , лепидозавроморфам или членам Diapsida incertae sedis . В анализе отношений неодиапсидов, проведенном Мартином Эскуррой в 2016 году , они были обнаружены как члены продвинутой группы неодиапсидов Sauria в политомии с Lepidosauromorpha и Archosauromorpha, причем то, что они были самыми ранними расходящимися членами любой группы, также вполне вероятно. [29] Положение базальных архозавроморфов подтверждается последовательностью окостенения их эмбрионов. [20]
Эволюционная история
[ редактировать ]
Судя по их примитивному филогенетическому положению, хористодеры должны были отделиться от всех других известных групп рептилий до конца пермского периода , более 250 миллионов лет назад. [5] В 2015 году Райнер Р. Шох сообщил о новом небольшом (длиной ~ 20 см) диапсиде из среднего триаса ( ладинского периода ) Нижнего Кейпера в Южной Германии , известном как по краниальному, так и по посткраниальному материалу, который, как он утверждал, представляет собой старейшую из известных хористодер. [31] Исторически предполагалось, что пахистрофей из позднего триаса ( рета ) Британии был хористодером. [32] но позже было показано, что он принадлежит к группе морских рептилий Thalattosauria . [33] Самый древний однозначный хористодер - это небольшой ящерицеобразный Cteniogenys, самые древние известные останки которого известны из поздней средней юры ( бат ~ 168-166 миллионов лет назад) формаций Форест Мрамор и Килмалуаг в Британии, а останки также известны из Верхних Юрская формация Алькобаса в Португалии и формация Моррисон в США, с широко схожими остатками, также известными из позднесреднеюрской ( келловейской ) формации Балабансай в Кыргызстане в Центральной Азии . [5] [34] батская итатская свита Западной Сибири, [35] а также, возможно, формация Ануал батского возраста в Марокко, Северная Африка. [36]
Хористодеры претерпели значительную эволюционную радиацию в Азии в раннем меловом периоде , что представляет собой высшую точку разнообразия хористодер, включая первые находки гавиалоподобных Neochoristodera, которые, по-видимому, развились в условиях регионального отсутствия водных неозуховых крокодилиформ. [5] Частичная бедренная кость неопределенной хористодеры известна из члена желтой кошки формации Кедровых гор в Северной Америке. [37] По-видимому, они отсутствуют в тщательно отобранных европейских местонахождениях берриасского возраста группы Пурбек в Великобритании и барремского возраста формации Ла Уэргина в Испании. [5] хотя есть сведения о небольшом таксоне, похожем на Cteniogenys, из беррианского возраста Костный пласт Анжак-Шаранта во Франции. [38] Во второй половине позднего мела ( кампан — маастрихт ) неохористодер Champsosaurus встречается в Юте, Вайоминге, Монтане, Северной Дакоте, Альберте и Саскачеване, которые находились вдоль западного побережья Западного Внутреннего морского пути на острове Ларамидия . [5] Неопределенные остатки neochoristoderes также известны из канадской Высокой Арктики, датируемые ранним поздним мелом ( коньяк - турон ). [39] и из формации Навесинк в Нью-Джерси из позднего мела (маастрихт), которая образовала отдельный остров Аппалачи . [40] Позвонки из сеномана Германии [41] кампанского возраста и Грюнбахская формация в Австрии. [42] указывают на присутствие хористодер в Европе в этот период времени. Единственная находка хористодер из Азии в позднем мелу — это единственный позвонок из турона Японии. [43] Фрагментарные останки, обнаруженные в кампанских Олдмана и Парка динозавров формациях в Альберте , Канада, также, возможно, предполагают присутствие небольших тел «неохористодер» в Северной Америке в позднем меловом периоде.
Чампзозавр пережил вымирание K-Pg и вместе с другими неохористодерами симоедозаврами присутствует в Европе, Азии и Северной Америке во время палеоцена, однако они вымерли в раннем эоцене . Их вымирание совпадает с крупным круговоротом фауны, связанным с повышенными температурами . [5] Мелкие «неохористодеры», отсутствующие в летописи окаменелостей после раннего мела (за исключением возможных североамериканских останков), вновь появляются в виде ящерицеподобных Lazarussuchus из позднего палеоцена Франции. [17] Европейский эндемик Lazarussuchus - последний известный хористодер, переживший вымирание неохористодер в начале эоцена, причем самые молодые известные останки - это останки L. dvoraki из раннего миоцена Чешской Республики. [5] [44] а также, насколько это возможно, неопределенные останки Lazarussuchus, обнаруженные в позднем миоцене (~ 11,6 миллионов лет назад) на юге Германии. [45]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Браунштейн, CD (2022). «Высокое морфологическое разнообразие в причудливой палеоценовой фауне хищных пресноводных рептилий» . BMC Экология и эволюция . 22 (1): 34. дои : 10.1186/s12862-022-01985-z . ПМЦ 8935759 . ПМИД 35313822 .
- ^ Сьюс, Х.-Д. (2019). «Архозавроморфы: Хористодера». Возникновение рептилий: 320 миллионов лет эволюции . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса. стр. 167–168. ISBN 978-1-4214-2867-3 .
- ^ Коуп ЭД (1876 г.). «О некоторых вымерших рептилиях и батрахиях из русл рек Джудит и Фокс-Хиллз в Монтане» . Труды Академии естественных наук Филадельфии . 1876 : 340–359.
- ^ Коуп, ЭД (1884). «Хористодера» . Американский натуралист . 18 : 815–817.
- ^ Перейти обратно: а б с д и ж г час я дж к Мацумото Р., Эванс С.Э. (2010). «Хористодеры и пресноводные комплексы Лавразии» . Журнал иберийской геологии . 36 (2): 253–274. Бибкод : 2010JIbG...36..253M . дои : 10.5209/rev_jige.2010.v36.n2.11 .
- ^ Перейти обратно: а б Эванс С.Е., Хехт МК (1993). «История вымершей группы рептилий, Choristodera: долголетие, Lazarus-Taxa и летопись окаменелостей». В Хехт М., Макинтайр Р.Дж., Клегг М.Т. (ред.). Эволюционная биология . Бостон, Массачусетс: Springer US. стр. 323–338. дои : 10.1007/978-1-4615-2878-4_8 . ISBN 978-1-4615-2878-4 .
- ^ Перейти обратно: а б Мацумото Р., Донг Л., Ван Й., Эванс С.Э. (2019). «Первая запись почти полной хористодеры (Reptilia: Diapsida) из верхней юры провинции Хэбэй, Китайская Народная Республика» (PDF) . Журнал систематической палеонтологии . 17 (12): 1031–1048. Бибкод : 2019JSPal..17.1031M . дои : 10.1080/14772019.2018.1494220 . S2CID 92421503 .
- ^ Перейти обратно: а б Даджен Т.В., Мэддин ХК, Эванс Д.С., Мэллон Дж.К. (апрель 2020 г.). «Компьютерный томографический анализ черепа Champsosaurus lindoei и последствия неоморфного окостенения хористодера» . Журнал анатомии . 236 (4): 630–659. дои : 10.1111/joa.13134 . ПМК 7083570 . ПМИД 31905243 .
- ^ Цинь, Ваньин; Йи, Хунъюй; Гао, Кэцинь (2 июля 2022 г.). «Неоморфное окостенение, соединяющее черепную коробку, чешуйчатую кость и квадратную кость у хористодерных рептилий: данные микроКТ» . Ископаемый рекорд . 25 (1): 1–10. Бибкод : 2022FossR..25....1Q . дои : 10.3897/fr.25.79595 . ISSN 2193-0074 .
- ^ Перейти обратно: а б с Донг, Липин; Мацумото, Рёко; Кусухаши, Нао; Ван, Юаньцин; Ван, Юань; Эванс, Сьюзен Э. (2 августа 2020 г.). «Новая хористодера (Reptilia: Choristodera) из апт-альбского угольного месторождения в Китае» . Журнал систематической палеонтологии . 18 (15): 1223–1242. Бибкод : 2020JSPal..18.1223D . дои : 10.1080/14772019.2020.1749147 . ISSN 1477-2019 . S2CID 219047160 .
- ^ Перейти обратно: а б с Гао К.К., Ксепка Д.Т. (июнь 2008 г.). «Остеология и таксономическая ревизия Hyphalosaurus (Diapsida: Choristodera) из нижнего мела провинции Ляонин, Китай» . Журнал анатомии . 212 (6): 747–68. дои : 10.1111/j.1469-7580.2008.00907.x . ПМЦ 2423398 . ПМИД 18510504 .
- ^ Даджен, Томас В.; Мэддин, Хиллари С.; Эванс, Дэвид С.; Мэллон, Джордан К. (28 апреля 2020 г.). «Внутренняя черепная анатомия Champsosaurus (Choristodera: Champsosauridae): значение для нейросенсорной функции» . Научные отчеты . 10 (1): 7122. Бибкод : 2020NatSR..10.7122D . дои : 10.1038/s41598-020-63956-y . ISSN 2045-2322 . ПМЦ 7188685 . ПМИД 32346021 .
- ^ Мацумото Р., Эванс С.Э. (март 2016 г.). «Морфология и функция небного зубного ряда Choristodera» . Журнал анатомии . 228 (3): 414–29. дои : 10.1111/joa.12414 . ПМК 5341546 . ПМИД 26573112 .
- ^ Перейти обратно: а б Кецин Г., Эванс С., Цян Дж., Норелл М., Шуань Дж. (25 сентября 2000 г.). «Исключительный ископаемый материал полуводной рептилии из Китая: разрешение загадки». Журнал палеонтологии позвоночных . 20 (3): 417–421. doi : 10.1671/0272-4634(2000)020[0417:EFMOAS]2.0.CO;2 . S2CID 86064702 .
- ^ Ван, Мяоян; Син, Лида; Ню, Кеченг; Лян, Цинцин; Эванс, Сьюзен Э. (апрель 2023 г.). «Новый экземпляр раннемелового длинношеего хористодера Hyphalosaurus из Ляонина, Китай, с исключительно сохранившимся покровом» . Меловые исследования . 144 : 105451. Бибкод : 2023CrRes.14405451W . дои : 10.1016/j.cretres.2022.105451 .
- ^ Перейти обратно: а б Эриксон Б.Р. (июнь 1985 г.). «Аспекты некоторых анатомических структур Champsosaurus (Reptilia: Eosuchia)». Журнал палеонтологии позвоночных . 5 (2): 111–127. Бибкод : 1985JVPal...5..111E . дои : 10.1080/02724634.1985.10011849 .
- ^ Перейти обратно: а б Мацумото Р., Баффетаут Э., Эскуилли Ф., Хервет С., Эванс С.Э. (2013). «Новый материал хористодеры Lazarussuchus (Diapsida, Choristodera) из палеоцена Франции». Журнал палеонтологии позвоночных . 33 (2): 319–339. Бибкод : 2013JVPal..33..319M . дои : 10.1080/02724634.2012.716274 . S2CID 129438118 .
- ^ Мацумото, Рёко; Фудзивара, Синъити; Эванс, Сьюзен Э. (2021). «Пищевое поведение и функциональная морфология шеи длиннорылой водной ископаемой рептилии Champsosaurus (Reptilia: Diapsida) в сравнении с современным крокодилом Gavialisangeticus» . Журнал анатомии . 240 (5): 893–913. дои : 10.1111/joa.13600 . ISSN 1469-7580 . ПМЦ 9005684 . ПМИД 34865223 . S2CID 244917425 .
- ^ Ван, Сяолинь; Мяо, Десуи; Чжан, Югуан (1 февраля 2005 г.). «Каннибализм у полуводных рептилий раннего мела Китая» . Китайский научный бюллетень . 50 (3): 282–284. Бибкод : 2005ЧСБу..50..282Вт . дои : 10.1007/BF02897540 . ISSN 1861-9541 . S2CID 95194181 .
- ^ Перейти обратно: а б с д и Цзян, Баоюй; Он, Имин; Эльслер, Армин; Ван, Шэнъюй; Китинг, Джозеф Н.; Сон, Джуньи; Кернс, Стюарт Л.; Бентон, Майкл Дж. (12 июня 2023 г.). «Длительная задержка эмбриона и живорождение у первых амниот» . Экология и эволюция природы . 7 (7): 1131–1140. Бибкод : 2023NatEE...7.1131J . дои : 10.1038/s41559-023-02074-0 . ISSN 2397-334X . ПМЦ 10333127 . ПМИД 37308704 .
- ^ Цзи Ц, Ву XC, Ченг Юнь (апрель 2010 г.). «Меловые хористодерные рептилии родили живых детенышей». Die Naturwissenschaften . 97 (4): 423–8. Бибкод : 2010NW.....97..423J . дои : 10.1007/s00114-010-0654-2 . ПМИД 20179895 . S2CID 8719805 .
- ^ Хоу Л.Х., Ли П.П., Ксепка Д.Т., Гао К.К., Норелл М.А. (апрель 2010 г.). «Последствия яиц с гибкой скорлупой у хористодерных рептилий мелового периода» . Слушания. Биологические науки . 277 (1685): 1235–9. дои : 10.1098/рспб.2009.2035 . ПМЦ 2842823 . ПМИД 20018793 .
- ^ Перейти обратно: а б Люй Дж, Кобаяши Ю, Диминг Д.С., Лю Ю (20 октября 2014 г.). «Послеродовой родительский уход за меловым диапсидом из северо-восточного Китая» (PDF) . Геонаучный журнал . 19 (2): 273–280. дои : 10.1007/s12303-014-0047-1 . S2CID 49570935 .
- ^ Кацура Ю. (2007). «Слияние крестцов и анатомии у Champsosaurus (Diapsida, Choristodera)». Историческая биология . 19 (3): 263–271. Бибкод : 2007HBio...19..263K . дои : 10.1080/08912960701374659 . S2CID 84966652 .
- ^ Ли Й.Н., Конг Д.И., Юнг Ш.Х. (сентябрь 2020 г.). «Первый возможный путь хористодер из формации Тэгу нижнего мела в Южной Корее и его влияние на передвижение хористодер» . Научные отчеты . 10 (1): 14442. Бибкод : 2020NatSR..1014442L . дои : 10.1038/s41598-020-71384-1 . ПМЦ 7468130 . ПМИД 32879388 .
- ^ Локли, Мартин Г.; Хант, Адриан П. (14 сентября 1995 г.). «Следы цератопсид и связанная с ними ихнофауна из формации Ларами (верхний мел: маастрихт) в Колорадо» . Журнал палеонтологии позвоночных . 15 (3): 592–614. Бибкод : 1995JVPal..15..592L . дои : 10.1080/02724634.1995.10011251 . ISSN 0272-4634 .
- ^ Гао, Кэ-Цинь; Фокс, Ричард К. (16 ноября 2005 г.). «Новая хористодера (Reptilia: Diapsida) из нижнего мела западной провинции Ляонин, Китай, и филогенетические связи Monjurouchidae» . Зоологический журнал Линнеевского общества . 145 (3): 427–444. дои : 10.1111/j.1096-3642.2005.00191.x . ISSN 1096-3642 .
- ^ Мацумото, Рёко; Цогтбаатар, Хишигджав; Исигаки, Синобу; Цогтбаатар, Чинзориг; Энхтайван, Зориг; Эванс, Сьюзен (2019). «Выявление пропорций тела загадочного хористодера Хурендухозавра из Монголии» . Acta Palaeontologica Polonica . 64 . дои : 10.4202/app.00561.2018 . S2CID 133925482 .
- ^ Перейти обратно: а б Эскурра, доктор медицины (28 апреля 2016 г.). «Филогенетические взаимоотношения базальных архозавроморфов с акцентом на систематике протерозуховых архозавроформ» . ПерДж . 4 : е1778. дои : 10.7717/peerj.1778 . ISSN 2167-8359 . ПМЦ 4860341 . ПМИД 27162705 .
- ^ Гао, Кэцинь; Фокс, Ричард К. (декабрь 1998 г.). «Новые хористодеры (Reptilia: Diapsida) из верхнего мела и палеоцена, Альберты и Саскачевана, Канада, и филогенетические отношения Choristodera» . Зоологический журнал Линнеевского общества . 124 (4): 303–353. дои : 10.1111/j.1096-3642.1998.tb00580.x .
- ^ Шох, Р.Р. (2015). «Рептилии» (PDF) . Леттенкейпер: окно во времена до динозавров (на немецком языке). Государственный музей естественной истории Штутгарта. стр. 231–264.
- ^ Сторрс, Г.В.; Гауэр, ди-джей (1 ноября 1993 г.). «Самая ранняя хористодера (Diapsida) и пробелы в летописи окаменелостей полуводных рептилий» . Журнал Геологического общества . 150 (6): 1103–1107. Бибкод : 1993JGSoc.150.1103S . дои : 10.1144/gsjgs.150.6.1103 . ISSN 0016-7649 . S2CID 86088809 .
- ^ Куинн, Джейкоб Г.; Мато-Рэйвен, Евангелия Р.; Уайтсайд, Дэвид И.; Маршалл, Джон Э.А.; Хатчинсон, Дебора Дж.; Бентон, Майкл Дж. (4 июня 2024 г.). «Отношения и палеоэкология Pachystropheus rhaeticus, загадочной последней морской рептилии триаса (Diapsida: Thalattosauria)» . Журнал палеонтологии позвоночных . дои : 10.1080/02724634.2024.2350408 . ISSN 0272-4634 .
- ^ Аверьянов Александр О.; Мартин, Томас; Эванс, Сьюзен Э.; Бакиров, Айзек А. (1 января 2006 г.). «Первые юрские хористодеры Азии» . Naturwissenschaften . 93 (1): 46–50. Бибкод : 2006NW.....93...46A . дои : 10.1007/s00114-005-0061-2 . ISSN 1432-1904 . ПМИД 16344982 . S2CID 7111980 .
- ^ «Среднеюрский комплекс позвоночных Березовского угольного разреза в Западной Сибири (Россия)». Глобальная геология . 19 (4): 187–204. 2016. doi : 10.3969/j.issn.1673-9736.2016.04.01 .
- ^ Хаддуми Х., Аллен Р., Меслоу С., Метайс Г., Монбарон М., Понс Д. и др. (2016). «Гуэльб-эль-Ахмар (бат, Ануальная синклиналь, восточное Марокко): первая континентальная флора и фауна, включая млекопитающих из средней юры Африки» (PDF) . Исследования Гондваны . 29 (1): 290–319. Бибкод : 2016GondR..29..290H . дои : 10.1016/j.gr.2014.12.004 .
- ^ Бритт, Брукс Б.; Шитц, Родни Д.; Бринкман, Дональд Б.; Эберт, Дэвид А. (11 декабря 2006 г.). «Барремский неохористодер из формации Кедр-Маунтин, штат Юта, США» Журнал палеонтологии позвоночных . 26 (4): 1005–1008. doi : 10.1671/0272-4634(2006)26[1005:abnftc]2.0.co;2 . ISSN 0272-4634 . S2CID 86258448 .
- ^ Ронан Аллен, Ромен Вулло, Ли Розада, Жереми Анкетен, Рено Бурже и др.. Палеобиоразнообразие позвоночных раннего мела (берриаса) Анжак-Шаранта Лагерштетт (юго-запад Франции): последствия для круговорота континентальной фауны на границе J/K . Geodiversitas, Национальный музей естественной истории Парижа, В печати. ffhal-03264773f
- ^ Вандермарк, Дебора; Тардуно, Джон А.; Бринкман, Дональд Б. (май 2007 г.). «Ископаемая популяция шампсозавров из высоких широт Арктики: последствия для палеотемператур позднего мела» . Палеогеография, Палеоклиматология, Палеоэкология . 248 (1–2): 49–59. Бибкод : 2007PPP...248...49В . дои : 10.1016/j.palaeo.2006.11.008 .
- ^ Даджен, Томас В.; Лэндри, Зоя; Каллахан, Уэйн Р.; Мелинг, Карл М.; Балванц, Стивен (2021). «Аппалачская популяция неохористодер (Diapsida, Choristodera) выявлена с использованием ископаемых свидетельств и моделирования экологической ниши» . Палеонтология . 64 (5): 629–643. Бибкод : 2021Palgy..64..629D . дои : 10.1111/пала.12545 . ISSN 1475-4983 . S2CID 237761128 .
- ^ Рейсс С., Шеер У., Сакс С., Кир Б.П. (13 декабря 2018 г.). «Заполнение биостратиграфического пробела: Первый хористодер из нижнего - среднего мелового интервала Европы». Меловые исследования . 96 : 135–141. дои : 10.1016/j.cretres.2018.12.009 . ISSN 0195-6671 . S2CID 134904339 .
- ^ Баффето, Эрик (1989). «Первые свидетельства существования Choristodera (Reptilia, Diapsida) в верхнем мелу Европы: позвонки шампсозавра из отложений Госау (Кампан) в Нижней Австрии» [Первые свидетельства существования хористодер (Reptilia, Diapsida) в верхнем мелу Европы: позвонки шампсозавра из пласты Госау (кампан) Нижней Австрии] (PDF) . Отчеты о заседаниях класса математики и естественных наук Австрийской академии наук (на немецком языке). 197 : 389–394.
- ^ Мацумото, Рёко; Мията, Шинья; Мицузука, Сюнсуке; Эванс, Сьюзен Э. (август 2021 г.) «Первая находка хористодер из верхнего мела Азии, формация Тамагава» . Группа, Япония» . Меловые исследования . 129 : 104999. doi : 10.1016/j.cretres.2021.104999 . ISSN 0195-6671 . S2CID 238680387 .
- ^ Эванс С.Е., Клембара Дж (2005). «Хористодерная рептилия (Reptilia: Diapsida) из нижнего миоцена северо-западной Богемии (Чехия)». Журнал палеонтологии позвоночных . 25 (1): 171–184. doi : 10.1671/0272-4634(2005)025[0171:ACRRDF]2.0.CO;2 . ISSN 0272-4634 . S2CID 84097919 .
- ^ Киршер, У.; Прието, Дж.; Бахтадсе, В.; Азиз, Х. Абдул; Допплер, Г.; Хагмайер, М.; Бёме, М. (1 августа 2016 г.). «Биохронологическая связующая точка основания тортонского яруса в европейских наземных условиях: магнитостратиграфия самых верхних отложений верхней пресноводной молассы Северо-Альпийского предгорья в Баварии (Германия)» (PDF) . Информационные бюллетени по стратиграфии . 49 (3): 445–467. дои : 10.1127/nos/2016/0288 . ISSN 0078-0421 .
Дальнейшее чтение
[ редактировать ]- де Брага М., Риппель О. (1997). «Филогения рептилий и взаимоотношения черепах» . Зоологический журнал Линнеевского общества . 120 (3): 281–354. дои : 10.1111/j.1096-3642.1997.tb01280.x .
- Эриксон Б.Р. (1972). Лепидозавровая рептилия Champsosaurus в Северной Америке . Палеонтология, Монография. Том. 1. Музей науки Миннесоты.
- Мацумото Р., Сузуки С., Цогтбаатар К., Эванс С.Э. (февраль 2009 г.). «Новый материал о загадочной рептилии Хурендухозавре (Diapsida: Choristodera) из Монголии» естественных наук Общество 96 (2): 233–42. Бибкод : 2009NW.....96..233M . дои : 10.1007/s00114-008-0469-6 . ПМИД 19034405 . S2CID 13542692 .
- Ксепка Д., Гао К., Норелл М.А. (2005). «Новая хористодера из мела Монголии» . Американский музей Novitates (3468): 1–22. doi : 10.1206/0003-0082(2005)468<0001:ancftc>2.0.co;2 . HDL : 2246/2778 . S2CID 54179348 .

| Базы данных органов управления : Национальные |
|---|
- Хористодера
- Отряды доисторических четвероногих
- Келловианские первые выступления
- Среднеюрские таксономические отряды
- Позднеюрские таксономические порядки
- Раннемеловые таксономические порядки
- Таксономические порядки позднего мела
- Таксономические порядки палеоцена
- Таксономические порядки эоцена
- Таксономические порядки олигоцена
- Таксономические порядки миоцена
- Аквитанское вымирание
- Третичное вымирание таксонов позвоночных
- Таксоны, названные Эдвардом Дринкером Коупом