Дрепанозавр
| Дрепанозавры Временной диапазон: поздний триас ,
| |
|---|---|

| |
| Ископаемый экземпляр Drepanosaurus unguicaudatus | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | неодиапсид |
| Клэйд : | † Дрепанозавроморфа Ренесто и др. , 2010 г. |
| Подгруппы | |
| |
Дрепанозавры (члены клады Drepanosauromorpha ) — группа вымерших рептилий , живших между карнийским и ретским этапами позднего триасового периода , примерно между 230 и 210 миллионами лет назад. [ 1 ] Различные виды дрепанозавров характеризовались птичьим черепом, бочкообразным телом и узким горизонтально хвостом. У ряда дрепанозавров были специализированные хватательные конечности и часто цепкие хвосты, подобные хвостам хамелеонов . Обычно считается, что дрепанозавры вели древесный образ жизни (жили на деревьях). [ 2 ] и, вероятно, насекомоядные . [ 3 ] Некоторые исследования поочередно предполагают роющий (роющий) и водный образ жизни некоторых представителей. [ 4 ] Окаменелости дрепанозавров были найдены в Северной Америке ( Аризона , Нью-Мексико , Нью-Джерси , Юта ) и Европе ( Англия и северная Италия ). семейства Название взято от одноименного рода Drepanosaurus , что означает «серповидная ящерица», отсылка к их сильно изогнутым когтям.
Некоторые исследования включали дрепанозавров в группу Avicephala , в которую также входят планирующие Weigeltisauridae , но тесное родство между этими двумя группами подвергалось сомнению другими авторами. Их филогенетическое положение оспаривается: некоторые исследования считают их представителями Archosauromorpha (и, таким образом, более тесно связанными с современными птицами и крокодилами, чем с ящерицами), в то время как другие исследования считают их базальными неодиапсидами , не связанными ни с одним современным видом. рептилии. [ 2 ]
Описание
[ редактировать ]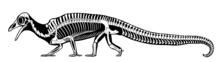
Дрепанозавры отличаются характерными треугольными черепами, напоминающими черепа птиц . Некоторые дрепанозавры, такие как Avicranium , имели заостренные беззубые птичьи клювы . Это сходство с птицами, возможно, привело к ошибочному приписыванию того, что могло быть черепом дрепанозавра, потенциальной «первой птице» Протоавису . [ 5 ]

Дрепанозавры отличались набором причудливых, почти хамелеоноподобных скелетных черт. Над плечами у большинства видов находился специальный «горб», образовавшийся в результате сращения позвонков, который, возможно, использовался для развитого прикрепления мышц к шее и позволял быстро совершать удары головой вперед (возможно, для ловли насекомых). У многих были руки с двумя пальцами вместо оставшихся трех, что было приспособлением для хватания ветвей. Некоторые особи мегаланкозавра (возможно, только самцы или самки) имели противоположные пальцы на каждой ноге, как у приматов, которые, возможно, использовались представителями одного пола для дополнительного захвата во время спаривания. У большинства видов были широкие цепкие хвосты, иногда с большим «когтем» на конце, опять же для облегчения лазания. Эти хвосты, высокие и плоские, как у тритонов и крокодилов , привели некоторых исследователей к выводу, что они были водными, а не древесными. В 2004 году Сентер отверг эту идею, а Кольбер и Олсен в своем описании Гипуронектора заявляют, что, хотя другие дрепанозавры, вероятно, вели древесный образ жизни, Гипуронектор был уникально приспособлен к водной жизни. [ 6 ] Хвост представителей этого рода был чрезвычайно глубоким и не цепким: он гораздо больше напоминал плавник, чем у представителей более эксклюзивной группы Drepanosauridae . [ 7 ] Воздушное передвижение приписывают как минимум двум родам дрепанозавров: Megalancosaurus и Hypuronector . Первое было первоначально предложено Рубеном и соавт. 1998 г. на основе птичьего характера и пропорций конечностей. [ 8 ] Хотя это предположение не было полностью исключено, с тех пор оно было в значительной степени отвергнуто из-за мегаланкозавра . неуклюжей, похожей на хамелеона анатомии [ 5 ] Гипуронектор , однако, гораздо скорее является планёром или летателем из-за удлиненных передних конечностей. [ 9 ] Адаптации, связанные с ископаемыми или рытьем, были обнаружены у трех родов дрепанозавров: Skybalonyx , Ancistronychus и Drepanosaurus . В частности, дрепанозавр , возможно, был приспособлен к рытью с помощью крюка и вытягивания, подобно современным муравьедам . Скайбалоникс обладал когтями, похожими по форме на современных кротов и ехидн , оба из которых являются копателями с вращением плечевых суставов. [ 4 ]

Филогенетическое положение дрепанозавров весьма спорно. Различные исследования предположили, что дрепанозавры являются проторозаврами архозавроморфами . [ 10 ] лепидозавроморфы , родственные кунеозавридам , [ 11 ] неящероподобные диапсиды , родственные вейгельтисавридам , [ 6 ] или (совсем недавно) базальные неодиапсиды . [ 12 ]
Ранние исследования
[ редактировать ]Когда дрепанозавр и долаброзавр были впервые обнаружены (в 1980 и 1992 годах соответственно), каждый из них считался ранним лепидозавром , предком современных ящериц. Мегаланкозавр впервые считался текодонтом ( то есть архозаврообразным ) после его открытия в 1980 году, но более поздние исследования определили его как пролацертиформного и, возможно, даже предка птиц, хотя эта последняя гипотеза не была подтверждена последующими исследованиями. [ 12 ]
Дрепанозавры вне Neodiapsida: Avicephala и Simiosauria.
[ редактировать ]В исследовании 2004 года Сентер поместил дрепанозавров вместе с целурозавридами ( вейгельтисавридами ) и лонгисквамами в кладу , которую он назвал Avicephala . Анализ Сентера поместил Avicephala в состав Diapsida , но за пределами Neodiapsida , определяемого Сентером как клада, содержащая «все таксоны, филогенетически отнесенные к Younginiformes и живым диапсидам». [ 6 ] [ 13 ]
Внутри Avicephala Сентер назвал группу Simiosauria («ящерицы-обезьяны») в честь чрезвычайно производных древесных форм. Симиозаврия была определена как «все таксоны, более тесно связанные с Drepanosauridae, чем с Coelurosauravus или Sauria ». Однако Ренесто и его коллеги (см. ниже) обнаружили, что дрепанозавриды обитают в пределах Саурии, что сделало бы кладу Симиозавров устаревшей. Сентер обнаружил, что Гипуронектор , первоначально описанный как дрепанозаврид, на самом деле находится сразу за пределами этой клады, вместе с примитивным дрепанозавром Валлезавром . Он также обнаружил тесную связь между дрепанозавридами Dolabrosaurus и Megalancosaurus . [ 6 ]
Следующая кладограмма была предложена Сентером в его анализе 2004 года: [ 6 ]
| Симиозаврия |
| ||||||||||||||||||
Клада, содержащая дрепанозавридов, Longisquama и Coelurosauravus (а также Wapitisaurus ), также была обнаружена в ходе анализа 2003 года, проведенного Джоном Мерком; однако в анализе Мерка эта клада была вложена в Neodiapsida как таксон Sauria сестринский . [ 14 ]

В исследовании 2006 года Ренесто и Бинелли обнаружили, что когда птерозавр Евдиморфодон был добавлен к исходной матрице Сентера, он оказался членом Avicephala. [ 15 ] Авторы также провели второй анализ, на этот раз на основе набора символов и матрицы, обновленной путем оценки дополнительных символов, которые ранее считались неизвестными, и путем добавления нескольких соответствующих символов. Этот анализ выявил дрепанозавридов как сестринский таксон Eudimorphodon ; клада, содержащая птерозавров и дрепанозавридов, была обнаружена как сестринский таксон Archosauriformes . Longisquama и Coelurosauravus не оказались близкими родственниками дрепанозаврид, а вместо этого были обнаружены как ненеодиапсидные диапсиды, как в анализе Сентера. Однако вполне возможно, что такое расположение может быть результатом плохого изучения Longisquama , а не отражением его истинного филогенетического положения. Авторы отметили, что существуют сходства в строении передних конечностей и плечевых областей Longisquama и всех или некоторых дрепанозавридов (например, длина плечевой кости как Longisquama , так и Vallesaurus равна четвертому пальцу кисти ) . Они подчеркнули, что не могут исключить возможность того, что по крайней мере некоторые сходства конвергентны из-за схожего поведения, и что они не исследовали Лонгисквама из первых рук. Следовательно, дальнейшие исследования дрепанозавридов должны принять во внимание гипотезу о том, что Longisquama может быть дрепанозавридом. [ 15 ]
Дрепанозавры как родственники кунеозаврид
[ редактировать ]В анализе 2004 года, проведенном Йоханнесом Мюллером, дрепанозавриды также оказались неящероподобными неодиапсидами; однако в этом анализе не было обнаружено, что Drepanosauridae тесно связаны с Coelurosauravus , а скорее были обнаружены как сестринский таксон Kuehneosauridae . [ 16 ] В исследовании 2009 года Сьюзен Э. Эванс провела филогенетический анализ с использованием модифицированной версии матрицы Мюллера. Эванс также обнаружил Drepanosauridae как сестринский таксон клады, содержащей базальный кунеозаврид Pamelina и остальную часть Kuehneosauridae; однако, в отличие от анализа Мюллера, дрепанозавриды и кунеозавриды были обнаружены как нелепидозавровые лепидозавроморфы . Эванс отметил, что эти две семьи имеют мало общих синапоморфий , а Мюллер сослался только на две. Один из них — усиление угла наклона зигапофизов задних спинных позвонков ; Эванс отметил, что этот признак также присутствует в скелетах ящериц, принадлежащих к современному роду Draco , «и, вероятно, является функциональным (и, следовательно, потенциально конвергентным)». , другая синапоморфия, закрытое щитовидное окно в тазу , «может быть изменчивым у британских кунеозавров и остается неизвестным у Памелины По словам Эванса ». Автор также отметил, что существует много различий между черепами дрепанозавридов и кунеозавридов и что единственные признаки черепа, общие для членов обоих семейств, — это примитивные признаки неодиапсидов и, следовательно, не могут использоваться для подтверждения тесного родства между двумя кладами. [ 11 ]
Дрепанозавры как архозавроморфы и отказ от Авицефал
[ редактировать ]В 1998 году Дилкс утверждал, что дрепанозавры были близкими родственниками танистрофеидов , и его филогенетический анализ использовался многими другими авторами. [ 10 ] Готтманн-Кесада и Сандер (2009) включили одного представителя Drepanosauridae, Megalancosaurus , в свой анализ взаимоотношений архозавроморфов; Было обнаружено, что это один из самых основных известных представителей Archosauromorpha и родственный таксон Protorosaurus . [ 17 ]
В более позднем исследовании Renesto et al. [ 1 ] продемонстрировал, что кладограмма Сентера 2004 года была основана на плохо определенных символах и данных. Таким образом, полученная в результате филогения была очень необычной по сравнению с любыми другими предыдущими исследованиями дрепанозавров или родственных им таксонов. Новая кладограмма, предложенная в этом последнем исследовании, отказалась как от Avicephala (поскольку она была полифилетической ), так и от Simiosauria . Определение Симиозаврии, данное Сентером, включало Sauria как внешний спецификатор, в результате чего клада стала устаревшей в исследовании Ренесто и др. (где дрепанозавры гнездились внутри Sauria). Вместо этого Ренесто и его коллеги определили новую кладу, Drepanosauromorpha, как наименее инклюзивную кладу, содержащую Hypuronector limnaios и Megalancosaurus preonensis . Был создан более инклюзивный таксон, Elyurosauria («ящерица со свернутым хвостом»), чтобы включить всех дрепанозавров со свернутыми хвостами. Таким образом, Vallesaurus является более производным, чем Hypuronector (как показывает его морфология). Drepanosaurus и Megalancosaurus также были помещены в новый таксон Megalancosaurinae.
Альтернативная кладограмма, представленная Renesto et al. (2010). [ 1 ]
| Дрепанозавроморфа |
| ||||||||||||||||||
Ренесто и др. (2010) использовали модифицированные версии матриц из более раннего анализа Лорина (1991). [ 18 ] и Дилкс (1998) [ 10 ] с целью определить филогенетическое положение Drepanosauromorpha внутри Diapsida. Анализы с использованием матрицы Лорина выявили дрепанозавров либо как сестринскую группу клады, содержащей Prolacerta , Trilophosaurus и Hyperodapedon ( Archosauromorpha ), либо в неразрешенной политомии с Archosauromorpha и Lepidosauromorpha . Анализы с использованием матрицы Дилкса выявили дрепанозавров либо как архозавроморфов- проторозавров и сестринского таксона клады, содержащей Tanystropeus , Langobardisaurus и Macrocnemus , либо в неразрешенной политомии с Lepidosauromorpha, Choristodera и несколькими кладами архозавроморфов. Ренесто и др. (2010) пришли к выводу, что синапоморфии авицефал, предложенные Сентером (2004), представляют собой просто эволюционные конвергенции, вызванные общим образом жизни, разделяемым дрепанозавридами, целурозавридами и лонгисквамами . Авторы не исключили, что дрепанозавры и лонгисквамы действительно могут быть близкими родственниками. [ 1 ]
Филогенетическое исследование, опубликованное Buffa et al. (2024) не обнаружили «авицефаланов» как близкородственных. Филогенетический анализ авторов выявил Drepanosauromorpha как аллокотозавровых архозавроморфов, в частности, как группу трилофозавридов сестринскую . [ 2 ]
Дрепанозавры как базальные диапсиды
[ редактировать ]В 2017 году Притчард и Несбитт применили филогенетический анализ при описании Avicranium , нового рода дрепанозавров. Это исследование показало, что Drepanosauromorpha была одной из самых ранних расходящихся групп диапсид в анализе, даже более базальной, чем вейгелтисавриды (такие как Coelurosauravus ), тангасавриды (такие как Hovasaurus ) и юнгиниды (такие как Youngina ). Однако они не были столь же базальными, как Petrolacosaurus , один из самых ранних и примитивных известных диапсидов. Хотя дрепанозавры известны только из позднего триаса , эта новая находка предполагает, что первые представители линии дрепанозавроморфов, возможно, развились гораздо раньше, в перми (около 260 миллионов лет назад). [ 12 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Ренесто, Южная Каролина; Шпильманн, ДА; Лукас, SG (29 мая 2009 г.). «Самая старая запись о дрепанозавридах (Reptilia, Diapsida) из позднего триаса (Адаманский карьер Пласериас, Аризона, США) и стратиграфического диапазона дрепанозаврид» . Новогодний ежегодник геологии и палеонтологии - Трактаты . 252 (3): 315–325. дои : 10.1127/0077-7749/2009/0252-0315 . ISSN 0077-7749 .
- ^ Jump up to: а б с Буффа, Валентин; Фрей, Эберхард; Стейер, Дж. Себастьян; Лорен, Мишель (11 мая 2024 г.). « Двухперые «птицы»: Avicranium renestoi и парафилия птицеголовых рептилий (Diapsida: «Avicephala») » Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae050 . ISSN 0024-4082 .
- ^ Кастиелло, Марко; Ренесто, Сильвио; Беннетт, С. Кристофер (16 ноября 2016 г.). «Роль передней конечности в захвате добычи у позднетриасовой рептилии Megalancosaurus (Diapsida, Drepanosauromorpha)» . Историческая биология . 28 (8): 1090–1100. дои : 10.1080/08912963.2015.1107552 . ISSN 0891-2963 .
- ^ Jump up to: а б Дженкинс, XA; Причард, AC; Марш, AD; Клигман, БТ; Сидор, Калифорния; Рид, Кентукки (08 октября 2020 г.). «Использование ручной морфологии ногтей для прогнозирования использования субстрата Drepanosauromorpha и описания нового вида» . Журнал палеонтологии позвоночных . 40 (5): e1810058. дои : 10.1080/02724634.2020.1810058 . ISSN 0272-4634 . S2CID 225136171 .
- ^ Jump up to: а б Ренесто, С. (31 июля 2000 г.). «Птичья голова на теле хамелеона: Новые экземпляры загадочной диапсидной рептилии Megalancosaurus из позднего триаса Северной Италии» . Итальянская литература по палеонтологии и стратиграфии . 106 (2): 157–179. дои : 10.13130/2039-4942/5396 .
- ^ Jump up to: а б с д и Сентер, П. (2004). «Филогения Drepanosauridae (Reptilia: Diapsida)» . Журнал систематической палеонтологии . 2 (3): 257–268. дои : 10.1017/s1477201904001427 . ISSN 1477-2019 . S2CID 83840423 .
- ^ Кольбер, Э.Х.; Олсен, ЧП (22 июня 2001 г.). «Новая и необычная водная рептилия из формации Локатонг в Нью-Джерси (поздний триас, супергруппа Ньюарка)» . Американский музей Novitates (3334): 1–24. doi : 10.1206/0003-0082(2001)334<0001:anauar>2.0.co;2 . ISSN 0003-0082 . S2CID 17294610 .
- ^ Шох, Р.Р. (29 ноября 2007 г.). «Остеология малого архозавра Aetosaurus из верхнего триаса Германии» . Новогодний ежегодник геологии и палеонтологии - Трактаты . 246 (1): 1–35. дои : 10.1127/0077-7749/2007/0246-0001 . ISSN 0077-7749 .
- ^ Ренесто, С.; Шпильманн, Дж.А.; Лукас, СГ; Спаньоли, GT (2010). Таксономия и палеобиология позднетриасовых (карнийско-норийских: адаманско-апачских) дрепанозавров (Diapsida: Archosauromorpha: Drepanosauromorpha) . Альбукерке, Нью-Мексико, США: Музей естественной истории и науки Нью-Мексико. OCLC 550640808 .
- ^ Jump up to: а б с Дилкс, DM (1998). «Раннетриасовый ринхозавр Mesosuchus Browni и взаимоотношения базальных архозавроморфных рептилий» . Философские труды Лондонского королевского общества, серия B. 353 (1368): 501–541. дои : 10.1098/rstb.1998.0225 . ПМК 1692244 .
- ^ Jump up to: а б Эванс, SE (2009). «Ранняя рептилия-кунеозаврид из раннего триаса Польши» (PDF) . Польская палеонтология . 65 : 145–178.
- ^ Jump up to: а б с Причард, AC; Несбитт, SJ (01 октября 2017 г.). «Птичий череп у триасовой диапсидной рептилии увеличивает гетерогенность морфологической и филогенетической радиации диапсид» . Королевское общество открытой науки . 4 (10): 170499. Бибкод : 2017RSOS....470499P . дои : 10.1098/rsos.170499 . ISSN 2054-5703 . ПМК 5666248 . ПМИД 29134065 .
- ^ Ренесто, С. (31 марта 1994 г.). « Мегаланкозавр , возможно, древесный архозавроморф (рептилия) из верхнего триаса северной Италии» . Журнал палеонтологии позвоночных . 14 (1): 38–52. дои : 10.1080/02724634.1994.10011537 . ISSN 0272-4634 .
- ^ Мерк, Дж. (2003). «Древесная радиация неящеричных диапсид». Журнал палеонтологии позвоночных . 23 (Дополнение к 3): 78А. дои : 10.1080/02724634.2003.10010538 . S2CID 220410105 .
- ^ Jump up to: а б Ренесто, С.; Бинелли, Г. (31 марта 2006 г.). « Vallesaurus cenensis WILD, 1991, дрепанозаврид (Reptilia, Diapsida) из позднего триаса северной Италии» . Итальянский журнал палеонтологии и стратиграфии . 112 (1): 77–94. дои : 10.13130/2039-4942/5851 .
- ^ Мюллер, Дж. (2004). «Отношения между диапсидными рептилиями и влияние таксонного отбора». В Арратии, Г.; Уилсон, MVH; Клотье, Р. (ред.). Последние достижения в области происхождения и ранней радиации позвоночных . Верлаг Доктор Фридрих Пфейль. стр. 379–408. ISBN 978-3-89937-052-2 .
- ^ Готтманн-Кесада, А.; Сандер, премьер-министр (19 марта 2009 г.). «Переописание раннего архозавроморфа Protorosaurus speneri MEYER, 1832 и его филогенетических связей» . Палеонтографика Абтейлунг А. 287 (4–6): 123–220. дои : 10.1127/пала/287/2009/123 .
- ^ Лаурин, М. (1991). «Остеология нижнепермского эозуха из Техаса и обзор филогении диапсид» . Зоологический журнал Линнеевского общества . 101 (1): 59–95. дои : 10.1111/j.1096-3642.1991.tb00886.x .
Внешние ссылки
[ редактировать ]- Обезьяньи ящерицы триаса — иллюстрированная статья о дрепанозаврах из HMNH.
- Профессор Сильвио Ренесто — Палеонтология позвоночных в Университете Инсубрии: исследования — изображения и обсуждение дрепанозавра .
- Профессор Сильвио Ренесто — Палеонтология позвоночных в Университете Инсубрии: исследование — изображения и обсуждение мегаланкозавра .




