Мезозавр
| Мезозавры Временной диапазон: Приуралье , | |
|---|---|
 | |
| Мезозавр | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | † Парарептилии |
| Заказ: | † Мезозаврия Сили , 1892 г. |
| Семья: | † Мезозавриды Баур, 1889 г. |
| Роды | |
Мезозавры («средние ящерицы») — группа мелких водных рептилий, живших в раннепермский период ( Приуралье ), примерно 299–270 миллионов лет назад . Мезозавры были первыми известными водными рептилиями , по-видимому, вернувшимися к водному образу жизни от более наземных предков. Непонятно, что и как многие земные черты, проявленные этими предками; недавние исследования не могут с уверенностью установить, были ли первые амниоты полностью наземными животными или только амфибиями. [1] Большинство авторов считают, что мезозавры вели водный образ жизни. [1] [2] хотя взрослые животные могли быть земноводными, а не полностью водными, о чем свидетельствует их умеренная адаптация скелета к полуводному образу жизни. [3] [4] Точно так же их родство сомнительно; они, возможно, были одними из самых базальных зауропсидов [5] [6] или среди наиболее базальных парарептилий (в случае которых парарептилиями были базальные зауропсиды). [7] [8]
Филогения
[ редактировать ]Филогенетическое положение мезозавров имеет важное значение для определения Reptilia. В одном из первых крупных филогенетических исследований амниот (позвоночных, откладывающих яйца на суше) Готье с соавт. (1988) поместили Mesosauridae в группу под названием Parareptilia . [9] Парарептилия означает «на стороне рептилий» и была помещена за пределы клады Рептилии, которая считалась коронной группой . В качестве коронной группы Reptilia включала самого недавнего общего предка двух основных линий ныне живущих рептилий — анапсидов (в частности, черепах ) и диапсидов (всех других ныне живущих рептилий) — и всех потомков этого общего предка. Эта точка зрения на размещение черепах вне диапсидов сейчас устарела, и большинство современных палеонтологов полагают, что Testudines ( черепахи и их союзники) произошли от диапсидных рептилий, потерявших височные окна. Более поздние морфологические филогенетические исследования с учетом этого поместили черепах прочно в диапсиды. [10] [11] [12] [13] и, чаще, как родственный таксон Archosauria (состоящий из крокодилов , динозавров , включая птиц , и их союзников). [14] Более того, Anapsida редко считается действительной кладой в недавних филогенетических анализах. [15] [16] В этом смысле Reptilia была таксоном, основанным на узлах, потому что первый общий предок рептилий должен был быть «узлом» на филогенетическом дереве. В рамках этой филогении многие вымершие формы, традиционно считавшиеся рептилиями, включая мезозавров, были исключены из группы, поскольку они находились за пределами узла. [17]
| Готье и др. , 1988 г. [9] |

Исследование Лорина и Рейша (1995) стало вторым крупным филогенетическим анализом амниот. [18] Как Готье и др. Лаурин и Рейс использовали Reptilia в качестве коронной группы и поместили мезозавров за пределы группы. Их филогения отличалась тем, что парарептилии Gauthier et al. теперь считались близкими родственниками черепах в коронной группе Reptilia. Лаурин и Рейс приняли название Sauropsida как таксон, основанный на узлах, включающий последнего общего предка мезозавров и рептилий. Традиционно амниоты делятся на две группы: линию млекопитающих, называемую Synapsida , и линию рептилий, называемую Reptilia или Sauropsida. Фактически, исследование Готье (1994) определило Sauropsida как всех амниот, более тесно связанных с рептилиями, чем с млекопитающими, а это означало, что Sauropsida был таксоном, основанным на стволе, охватывающим всю рептильную линию или рептильный «ствол» Amniota (Synapsida была ствол млекопитающих). В рамках этой филогении единственная группа, которая не позволяет Sauropsida быть эквивалентной Reptilia, — это мезозавры. [17]
| Лаурин и Рейс (1995) [18] |
Более поздние филогенетические анализы, такие как анализ Modesto (1999), подтверждают данные Gauthier et al. (1988), совместив мезозавров с парарептилиями. [7] Однако эти филогении следуют за Лорином и Рейсом (1995), которые поместили Parareptilia в кронную группу Reptilia, а это означает, что мезозавры снова стали членами Reptilia. Используя основанное на узлах определение Лаурина и Рейша зауропсиды как «последнего общего предка мезозавров, тестудинов и диапсидов, а также всех его потомков», [18] Sauropsida и Reptilia — эквивалентные группы; мезозавры и тестудины более тесно связаны друг с другом, чем любая из групп с диапсидами, [а] это означает, что клада, содержащая тестудины и диапсиды (которая могла бы быть рептилиями коронной группы), также должна содержать мезозавров. Поскольку Reptilia была названа раньше, чем Sauropsida, она чаще всего используется в современных филогенетических анализах. [17]
| Модесто, 1999 г. [7] |
Филогенетический анализ 2017 года, проведенный Лорином (который ранее опубликовал исследование 1995 года) и Пиньейро, выявил мезозавров как базальных представителей Sauropsida/Reptilia и больше не присутствует в Parareptilia, при этом Parareptilia была переопределена как включающая бывших представителей Procolophonomorpha (которые оказались парафилетическими). , Millerosauria , Pareiasauria и Pantestudines , причем последние две оказались родственными группами друг для друга. Также было обнаружено, что Parareptilia фактически гнездится внутри Diapsida как сестринская группа Neodiapsida . [19]
| Лорин и Ананас , [19] |
В 2012 году было обнаружено, что у мезозавра есть отверстия в задней части черепа, называемые нижними височными окнами . Раньше считалось, что эта характеристика присутствует только у синапсидов и диапсидов. [20] Это подтвердило предыдущие результаты немецкого палеонтолога Фридриха фон Хюне, опубликованные уже в 1941 году. [21] Состояние черепа мезозавра наиболее похоже на состояние черепа синапсид, поскольку у обоих отсутствуют верхние височные отверстия диапсид. Нижние височные окна пока известны только у мезозавров , но могут присутствовать у всех мезозавров. Наличие или отсутствие височных окон является важным фактором в филогении мезозавров и других амниот, поскольку три основные группы амниот - Synapsida, Diapsida и Anapsida - были названы в честь количества отверстий в их черепе; Diapsida означает «две арки» в отношении двух перекладин, закрывающих верхнее и нижнее окна, Synapsida означает «сросшаяся арка» в отношении единственной перемычки в нижней части черепа, закрывающей одно отверстие, а Anapsida означает «нет арки». «имеется в виду черепа без каких-либо перемычек или окон. Мезозавров традиционно относили к анапсидам, поскольку считалось, что у них отсутствовали височные окна. Однако появление фенестров у амниот считалось весьма изменчивым признаком внутри группы за много лет до их открытия у Мезозавр ; многие анапсиды, такие как Candelaria , Bolosaurus и лантанозухоиды , обладают нижними височными окнами.
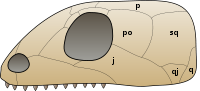

Филогенетическое положение мезозавров влияет на современное понимание того, как у амниот возникли височные окна. Если филогения Лорина и Рейша (1995) верна в том смысле, что мезозавры являются базальными зауропсидами, то нижнее височное окно может быть примитивной особенностью амниот, присутствующей у самого недавнего общего предка амниот. Синапсиды сохранили бы свои окна, как и зауропсиды, за исключением черепах и большинства парарептилий. Другая возможность, согласно филогении Лорина и Рейша, заключается в том, что нижние височные окна развились независимо у мезозавров, синапсидов, диапсид и некоторых парарептилий, и что отсутствие окон является примитивной особенностью у амниот. Если вместо этого мезозавры являются членами Parareptilia, наличие височных окон, вероятно, не является примитивной особенностью у амниот, а нижние височные отверстия у мезозавров могут быть характерны для линии базальных парарептилий, которая также включает несущих фенестры лантанозухоидов и болозавров . [20]
Биология
[ редактировать ]Долгое время считалось, что это прибрежные формы, обитавшие, вероятно, на относительно мелководье. [22] но недавние исследования показывают, что, по крайней мере, выходцы из Уругвая населяли гиперсоленую среду, а не прибрежную морскую среду. [23] Недавно описанные эмбрионы показывают, что пахиостоз ребер (которые были толще и плотнее, чем у наземных четвероногих) развился еще до вылупления, что позволяет предположить, что мезозавры были способны плавать при рождении или вскоре после этого. По-видимому, они были не очень быстрыми пловцами: оптимальная скорость плавания составляла от 0,15 до 0,86 м/с, но это должно было быть несколько быстрее, чем скорость их основной добычи, пигоцефаломорфных ракообразных. [24] Их репродуктивный способ несколько неопределенен, поскольку связь между взрослыми особями и возможными эмбрионами в утробе матери предполагает живорождение, как и у многих водных рептилий, но также было обнаружено потенциально изолированное яйцо. [25]
доказательства хищничества как пигоцефаломорфных ракообразных, так и представителей их собственного вида Недавно были установлены . Считается, что мезозавры в целом были адаптированы к гиперсоленой среде обитания. [26]
Исследование пропорций туловища и хвоста позвоночного столба мезозавра предполагает, что, хотя молодые особи могли вести полностью водный образ жизни, взрослые особи могли проводить некоторое время на суше; Это еще раз подтверждается тем, что взрослые животные редко встречаются в водной среде, а некоторые фекалии имеют признаки высыхания. Однако трудно сказать, насколько они были наземными, поскольку в том же исследовании говорится, что добыча пищи на суше была затруднена из-за их специализации на водной жизни. [4]
Примечания
[ редактировать ]- ^ Исследования с помощью молекулярной филогенетики , изучающие гены и белки живых организмов, позволяют предположить, что тестудины (черепахи) являются диапсидами. Эти исследования показывают, что мезозавры не образуют кладу с черепахами, исключающую диапсиды, но ископаемые данные все же позволяют предположить, что мезозавры образуют группу с парарептилиями. В большинстве последних исследований Reptilia не используется в качестве кронной группы и до сих пор содержит мезозавров и парарептилий.
Ссылки
[ редактировать ]- ^ Jump up to: а б Кановиль, А. и М. Лорен. 2010. Эволюция микроанатомии плечевой кости и образа жизни амниот, а также некоторые комментарии к палеобиологическим выводам. Биологический журнал Линнеевского общества 100:384–406.
- ^ Модесто, С. 2006. Черепной скелет раннепермской водной рептилии Mesosaurus tenuidens : значение для взаимоотношений и палеобиологии. Зоологический журнал Линнеевского общества 146:345-368.
- ^ Пинейро, Г. 2008. Мезозавры и другие окаменелости позднего палеозоя; стр. 179–205 в Д. Перере (ред.), Fósiles de Uruguay. ДИРАК, Монтевидео.
- ^ Jump up to: а б Пабло Нуньес Демарко и др. Был ли мезозавр полностью водной рептилией? Передний. Экол. Evol, опубликовано в Интернете 27 июля 2018 г.; дои: 10.3389/fevo.2018.00109
- ^ Лорин, М. и Р.Р. Рейс. 1995. Переоценка ранней филогении амниот. Зоологический журнал Линнеевского общества 113:165-223.
- ^ Лорен, Мишель; Пинейро, Грасиела Х. (2017). «Переоценка таксономического положения мезозавров и удивительная филогения ранних амниот» . Границы в науках о Земле . 5 : 88. Бибкод : 2017FrEaS...5...88L . дои : 10.3389/feart.2017.00088 . hdl : 20.500.12008/33548 .
- ^ Jump up to: а б с Модесто, СП (1999). «Наблюдения за строением раннепермской рептилии Stereosternum tumidum Cope». Палеонтология Африканская . 35 : 7–19.
- ^ Макдугалл, MJ; Модесто, СП; Броклхерст, Н.; Верьер, А.; Рейс, РР; Фрёбиш, Дж. (2018). «Ответ: переоценка таксономического положения мезозавров и удивительная филогения ранних амниот». Передний. Науки о Земле. 6:99 дои: 10.3389/feart.2018.00099
- ^ Jump up to: а б Готье, Дж.А.; Клюге, АГ; Роу, Т. (1988). «Ранняя эволюция амниоты». В Бентоне, MJ (ред.). Филогения и классификация четвероногих . Том. 1. Оксфорд: Кларендон Пресс. стр. 103–155. ISBN 978-0198577058 .
- ^ деБрага, М.; Риппель, О. (1997). «Филогения рептилий и взаимоотношения черепах» (PDF) . Зоологический журнал Линнеевского общества . 120 (3): 281–354. дои : 10.1111/j.1096-3642.1997.tb01280.x .
- ^ РУТА, МАРЧЕЛЛО; СИСНЕРОС, ХУАН К.; ЛИБРЕХТ, ТОРСТЕН; ЦУДЖИ, ЛИНДА А.; МЮЛЛЕР, ЙОХАННЕС (27 апреля 2011 г.). «Амниоты через крупные биологические кризисы: смена фауны среди парарептилий и массовое вымирание в конце пермского периода» . Палеонтология . 54 (5): 1117–1137. дои : 10.1111/j.1475-4983.2011.01051.x . ISSN 0031-0239 . S2CID 83693335 .
- ^ Борсук-Бялыницкая, Магдалена; Эванс, Сьюзен Э. (2009). «Длинношеий архозавроморф из раннего триаса Польши». Палеонтологическая Полоника . 65 : 203–234.
- ^ Эванс, Сьюзен Э. (2009). «Ранняя рептилия-кунеозаврида (Reptilia: Diapsida) из раннего триаса Польши». Польская Палеонтология . 65 : 145–178.
- ^ Филд, Дэниел Дж.; Готье, Жак А.; Кинг, Бенджамин Л.; Пизани, Давиде; Лайсон, Тайлер Р.; Петерсон, Кевин Дж. (5 мая 2014 г.). «На пути к согласованности в филогении рептилий: микроРНК поддерживают близость архозавров, а не лепидозавров к черепахам» . Эволюция и развитие . 16 (4): 189–196. дои : 10.1111/ede.12081 . ISSN 1520-541X . ПМЦ 4215941 . ПМИД 24798503 .
- ^ Модесто, Шон П.; Андерсон, Джейсон С. (1 октября 2004 г.). Лутзони, Франсуа (ред.). «Филогенетическое определение рептилий» . Систематическая биология . 53 (5): 815–821. дои : 10.1080/10635150490503026 . ISSN 1076-836X . ПМИД 15545258 .
- ^ Цудзи, Линда А.; Мюллер, Йоханнес (2009). «Собираем историю Parareptilia: филогения, разнообразие и новое определение клады» . Ископаемый рекорд . 12 (1): 71–81. дои : 10.1002/mmng.200800011 .
- ^ Jump up to: а б с Модесто, СП; Андерсон, Дж. С. (2004). «Филогенетическое определение рептилий» . Систематическая биология . 53 (5): 815–821. дои : 10.1080/10635150490503026 . ПМИД 15545258 .
- ^ Jump up to: а б с Лорин, М.; Рейс, Р.Р. (1995). «Переоценка ранней филогении амниот» (PDF) . Зоологический журнал Линнеевского общества . 113 (2): 165–223. дои : 10.1111/j.1096-3642.1995.tb00932.x .
- ^ Jump up to: а б Лорен, Мишель; Пинейро, Грасиела Х. (2017). «Переоценка таксономического положения мезозавров и удивительная филогения ранних амниот» . Границы в науках о Земле . 5 : 88. Бибкод : 2017FrEaS...5...88L . дои : 10.3389/feart.2017.00088 . hdl : 20.500.12008/33548 . ISSN 2296-6463 .
- ^ Jump up to: а б Пинейро, Г.; Фериголо, Дж.; Рамос, А.; Лаурин, М. (2012). «Краниальная морфология раннепермских мезозавридов Mesosaurus tenuidens и переоценка эволюции нижних височных окон». Comptes Рендус Палевол . 11 (5): 379–391. дои : 10.1016/j.crpv.2012.02.001 .
- ^ Хюне, Ф. фон (1941). «Остеология и систематическое положение мезозавра» . Палеонтографический отдел А. 92 : 43–58.
- ^ Оелофсен, Б. и округ Колумбия Араужо. 1983. Палеоэкологические последствия распространения мезозаврид в пермском море Ирати (бассейн Параны), Южная Америка. Бразильский журнал геонаук 13:1–6.
- ^ Пинейро, Г.; Рамос, А.; Госо, К.; Скарабино, Ф.; Лаурин, М. (2012). «Необычные условия окружающей среды сохранили пермский мезозавр Консерват-Лагерштетте из Уругвая» (PDF) . Acta Palaeontologica Polonica . 57 (2): 299–318. дои : 10.4202/app.2010.0113 . S2CID 55671441 .
- ^ Вилламил, JN; Демарко, Пенсильвания; Менегель, М.; Бланко, RE; Джонс, В.; Риндеркнехт, АС; Лорин, М.; Пинейро, Г. (2015). «Оценки оптимальной скорости плавания раннепермского мезозавра Mesosaurus tenuidens (Gervais 1867) из Уругвая». Историческая биология . 28 (7): 963–971. дои : 10.1080/08912963.2015.1075018 . S2CID 85846750 .
- ^ Пинейро, Г.; Фериголо, Дж.; Менегель, М.; Лаурин, М. (2012). «Самые старые известные амниотические эмбрионы предполагают живорождение у мезозавров». Историческая биология . 24 (6): 620–630. дои : 10.1080/08912963.2012.662230 . S2CID 59475679 .
- ^ Ривалдо Р. Да Силва; Хорхе Фериголо; Петр Байдек; Грасиела Х. Ананас (2017). «Питание мезозаврид» Границы в науках о Земле. 5: Статья 23. doi:10.3389/feart.2017.00023.






