Парарептилии
| Парарептилии | |
|---|---|

| |
| Коллаж из пяти окаменелостей парарептилий. По часовой стрелке сверху: Mesosaurus tenuidens ( мезозавр ), Delorhynchus cifellii (вероятный аклеисторинид ), Scutosaurus karpinski ( парейазавр ), Nyctiphruretus acudens ( никтифруретид ), Hypsognathus fenneri ( проколофонид ); | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | † Парарептилии Олсон , 1947 год. |
| Заказы | |
Parareptilia «почти-рептилии») — вымерший подкласс или клада базальных ( зауропсидов / рептилий , обычно считающийся родственным таксоном Eureptilia . (группа, которая, вероятно, содержит всех ныне живущих рептилий и птиц) Парарептилии впервые возникли ближе к концу каменноугольного периода и достигли наибольшего разнообразия в пермский период . Некоторые экологические инновации были впервые осуществлены парарептилиями среди рептилий. К ним относятся первые рептилии, вернувшиеся в морские экосистемы ( мезозавры ), первые двуногие рептилии ( болозавриды, такие как эвдибамус ), первые рептилии с развитой слуховой системой ( никтеролетериды и др.), а также первые крупные травоядные рептилии ( парейазавры ). Единственными парарептилиями, дожившими до триасового периода, были проколофоноиды , группа мелких универсальных, всеядных и травоядных животных. Самое большое семейство проколофоноидов, проколофониды , повторно диверсифицировалось в триасе, но впоследствии пришло в упадок и к концу периода вымерло. [ 2 ] [ 3 ]
По сравнению с большинством эврептилий, парарептилии сохранили довольно «примитивные» характеристики, такие как крепкое тело с низкой посадкой и большие надвисочные кости в задней части черепа. В то время как все эврептилии, кроме самых ранних, были диапсидами с двумя отверстиями в задней части черепа, парарептилии, как правило, были более консервативными в отношении степени височных окон . В современном использовании Parareptilia впервые использовалась как кладистически правильная альтернатива Anapsida , термину, который исторически относился к рептилиям с твердым черепом без отверстий за глазами. [ 4 ] Тем не менее, не все парарептилии имеют «анапсидные» черепа, а у некоторых есть большие отверстия в задней части черепа. У них также было несколько уникальных приспособлений, таких как большая ямка на верхней челюсти , широкий префронтально - небный контакт и отсутствие супрагленоидного отверстия лопатки . [ 4 ] [ 5 ]
Как и многие другие так называемые «анапсиды», парарептилии исторически были недостаточно изучены. Интерес к их взаимоотношениям возобновился в 1990-х годах, когда в нескольких исследованиях было установлено, что Testudines ( черепахи и их родственники) были членами Parareptilia. [ 4 ] Хотя это позволяет предположить, что парарептилии все-таки не вымерли, происхождение черепах до сих пор активно обсуждается. Многие другие морфологические или генетические анализы находят больше подтверждений существования черепах среди диапсидных эврептилий, таких как зауроптериги или архозавроморфы , а не среди парарептилий. [ 6 ] [ 7 ] [ 8 ] [ 3 ]
Описание
[ редактировать ]Череп
[ редактировать ]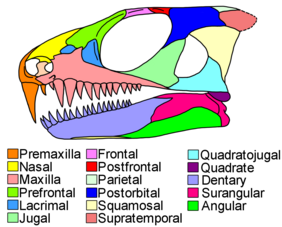
Черепа парарептилий были разнообразны: от мезозавров с удлиненными мордами, заполненными сотнями тонких зубов, до курносых, инкрустированных бугорками черепов парейазавров . Зубы парарептилий весьма различались по форме и функциям у разных видов. Однако на одном черепе они были относительно однородными. В то время как у большинства синапсидов и многих ранних эврептилий в передней половине черепа была псовая область с увеличенными клыкообразными зубами, очень немногие парарептилии обладали собачьими зубами. [ 5 ]
У многих амниот имеется ряд маленьких ямок, идущих вдоль костей по краю рта, но у парарептилий ямок всего несколько, причем одна особенно большая ямка вблизи передней части верхней челюсти . [ 4 ] [ 9 ] [ 7 ] Остальная часть черепа у большинства групп парарептилий часто имела сильную текстуру с ямками, гребнями и складками, иногда достигающими кульминации в виде сложных выступов или шипов. Верхняя челюсть обычно низкая, а предлобная и слезная кости перед глазом довольно крупные. У всех парарептилий, кроме мезозавров, префронтальная кость имеет пластинчатую внутреннюю ветвь, которая образует широкий контакт с небной костью неба . [ 4 ] [ 7 ] [ 5 ] На пересечении префронтальной, небной и слезной костей имеется заметное отверстие — орбитоназальное отверстие. Небо парарептилий также имеет беззубые и редуцированные эктоптеригоидные кости - состояние, доведенное до крайности у мезозавров, которые полностью потеряли эктоптеригоид. [ 4 ] [ 5 ]
У большинства парарептилий были большие орбиты (глазницы), значительно длиннее (спереди назад), чем область черепа за глазами. [ 5 ] Скуловая . кость, образующая нижний и задний край глазницы, имеет очень тонкий подглазничный отросток (передняя ветвь), подвисочный отросток обычно отсутствует (нижняя задняя ветвь) и толстый дорсальный отросток (верхняя задняя ветвь) Чешуйчатая кости, лежащие позади скуловой кости , и квадратноскуловая довольно крупные и вставлены сзади для размещения внутренних ушей. [ 4 ] [ 5 ] Традиционно считалось, что парарептилии имеют череп анапсидного типа с скуловой, чешуйчатой и квадратноскуловой костями, прочно сшитыми вместе без каких-либо промежутков или щелей между ними. Этот принцип все еще справедлив для некоторых подгрупп, таких как парейазавры. Однако известно, что все большее число таксонов парарептилий имело подвисочное окно — большое отверстие или выемку, расположенную среди костей позади глаза. У некоторых таксонов края таких отверстий могут включать дополнительные кости, такие как верхнечелюстная или заглазничная . [ 10 ] [ 11 ] При осмотре сверху задний край черепа прямой или имеет широкий срединный выступ. [ 5 ] Задний край черепа изнутри наружу образован тремя парами костей: затеменной , табличной и надвисочной . У парарептилий особенно большие надвисочные кости, которые часто простираются дальше назад, чем таблитчатые. [ 12 ]
Если не считать длинных и тонких челюстей мезозавров, челюсти большинства парарептилий были короткими и толстыми. Челюстной сустав образован суставным ( в нижней челюсти) и квадратным (в верхней челюсти). У многих парарептилий челюстной сустав на черепе смещен вперед за заднюю часть черепной коробки . [ 4 ] [ 7 ] Челюстные мышцы прикрепляются к венечному отростку — треугольной шпоре в задней половине челюсти. И зубная кость, и заднее межчелюстное отверстие (отверстие на внутренней поверхности челюсти) доходят до венечного отростка. [ 4 ] [ 5 ] Надугольная . кость, образующая верхнезаднюю часть челюсти, узкая, пластинчатая [ 13 ]
Посткраниальный скелет
[ редактировать ]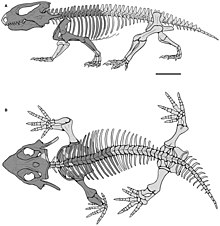
Существовали некоторые различия в форме тела парарептилий: первые члены группы имели внешний вид, похожий на ящерицу , с тонкими конечностями и длинными хвостами. Наиболее успешные и разнообразные группы парарептилий, парейазавры и проколофониды , имели массивное тело с редуцированными хвостами и толстыми конечностями с короткими пальцами. Эта общая форма тела характерна для других «котилозавров», таких как капториниды , диадектоморфы и сеймуриаморфы . [ 3 ] Другой общей «котилозавровой» особенностью парарептилий является «опухший» вид их позвонков , которые имеют широкие и выпуклые верхние поверхности. [ 12 ]
У парарептилий отсутствовало супрагленоидное отверстие на лопатке , отверстие, которое также отсутствует у варанопидов и неодиапсидов. [ 5 ] [ 11 ] У большинства из них была довольно короткая и толстая плечевая кость , расширенная около локтя. В отличие от ранних эврептилий, наружная часть нижней плечевой кости имела как небольшой супинаторный отросток, так и эктепикондилярное отверстие и бороздку. [ 4 ] Локтевая кость обычно имеет плохо развитый локтевой отросток , что является еще одной чертой в отличие от самых ранних эврептилий. [ 4 ] [ 5 ]
У большинства парарептилий подвздошная кость имела веерообразную форму и была ориентирована вертикально (а не горизонтально), что было необычной чертой среди ранних амниот. [ 4 ] [ 9 ] [ 7 ] Крестцовые ребра, соединяющие позвоночник с подвздошной костью, обычно были тонкими или веерообразными, с большими промежутками между ними. [ 4 ] Задние конечности обычно были ненамного длиннее передних, имели толстые лодыжки, как у рептилий, и короткие пальцы ног. Есть некоторые исключения, такие как Eudibamus , раннепермский болозаврид с очень удлиненными задними конечностями. [ 14 ]
История классификации
[ редактировать ]Название Parareptilia было придумано Олсоном в 1947 году для обозначения вымершей группы палеозойских рептилий, в отличие от остальных рептилий или Eureptilia («настоящие рептилии»). [ 15 ] Термин Олсена обычно игнорировался, и различные таксоны, позже известные как парарептилии, обычно не помещались в отдельные группы друг с другом. Многие из них были классифицированы как «котилозавры» ( таксон мусорной корзины , состоящий из массивных «примитивных» рептилий или рептилиеподобных четвероногих) или « анапсиды » (рептилии без височных окон , такие как современные черепахи).
Использование слова Parareptilia было возрождено кладистическими исследованиями для обозначения тех традиционных «анапсидов», которые, как считалось, не имели отношения к черепахам. Готье и др. (1988) предоставили первые филогенетические определения названий многих амниот таксонов и утверждали, что капториниды и черепахи были сестринскими группами, составляющими кладу Anapsida (в гораздо более ограниченном контексте, чем обычно применяется). Необходимо было найти название для клады различных ранних ныряющих рептилий пермского и триасового периода , которые больше не входят в состав анапсидов. Термин Олсена «парарептилии» был выбран для обозначения этой клады, хотя его нестабильность в их анализе означала, что Готье и др. (1988) не были достаточно уверены в том, чтобы сделать Parareptilia формальным таксоном. Их кладограмма выглядит следующим образом: [ 16 ]
| Амниота |
| ||||||||||||



Лорин и Рейс (1995) обнаружили несколько иную топологию, в которой Reptilia делится на Parareptilia и Eureptilia. Они утверждали, что Testudines (черепахи) были членами Parareptilia; Фактически, они явно определили Parareptilia как «Testudines и все амниоты, более тесно связанные с ними, чем с диапсидами». Captorhinidae были перенесены в Eureptilia, тогда как Parareptilia включала черепах наряду со многими таксонами, названными так Gauthier et al. (1988). Было одно важное исключение: мезозавры были помещены за пределы обеих групп как сестринский таксон коронной группы Reptilia. Мезозавров по-прежнему считали зауропсидами, поскольку они были ближе к рептилиям, чем к синапсидам. Традиционная группа «Анапсида» была отвергнута как парафилетическая совокупность. Кладограмма Laurin & Reisz (1995) представлена ниже: [ 4 ]
| Амниота |
| ||||||||||||||||||||||||||||||

в середине-конце 1990-х годов, Напротив, в нескольких исследованиях, проведенных Оливье Риппелем и Майклом де Брагой утверждалось, что черепахи на самом деле были лепидозавроморфов, диапсидами родственными зауроптеригам . [ 17 ] [ 18 ] [ 9 ] [ 6 ] [ 7 ] Сродство черепах с диапсидом подтверждается молекулярной филогенией . [ 19 ] [ 20 ] [ 21 ] [ 22 ] Первый полногеномный филогенетический анализ был проведен Wang et al. (2013). Используя черновые геномы Chelonia mydas и Pelodiscus sinensis, команда использовала самый большой набор данных о черепахах на сегодняшний день в своем анализе и пришла к выводу, что черепахи, вероятно, являются сестринской группой крокодилов и птиц (Archosauria). [ 8 ] Такое расположение внутри диапсидов предполагает, что линия черепах утратила характеристики диапсидного черепа, поскольку черепахи обладают анапсидным черепом. Это сделало бы парарептилий полностью вымершей группой с чертами черепа, напоминающими черепах в результате конвергентной эволюции . Поскольку черепахи расположены вне парарептилий, Цудзи и Мюллер (2009) переопределили Parareptilia как «наиболее инклюзивную кладу, содержащую Milleretta Rubidgei и Procolophon trigoniceps , но не Captorhinus aguti ». [ 3 ]
Кладограмма ниже соответствует анализу М.С. Ли, проведенному в 2013 году. [ 23 ]
| Амниота |
| ||||||||||||



Кладограмма ниже соответствует анализу Li et al . (2018). [ 24 ]
|
† Парарептилии | |||||||||||||||||||||||||||||||||||||
| Эврептилия |
| ||||||||||||||||||||||||||||||||||||



Исследование Дэвида П. Форда и Роджера Б. Дж. Бенсона, проведенное в 2020 году, показало, что Parareptilia была вложена в состав Diapsida как сестринская группа Neodiapsida , при этом клада, содержащая Neodiapsida и Parareptilia, получила название Neoreptilia, что предполагает, что парарептилии изначально были диапсидами. Это исключило мезозавров , которые снова оказались базальными среди зауропсидов. [ 11 ] Некоторые исследования показали, что Parareptilia являются парафилетическими, причем некоторые парарептилии более тесно связаны с диапсидами, чем с другими парарептилиями, согласно Simões et al. (2022) использовали Neoreptilia для клады, содержащей Procolophonomorpha + Neodiapsida. [ 25 ]
Эволюционная история
[ редактировать ]Самыми старыми известными парарептилиями являются болозавр Erpetonyx и аклеисторинид Carbonodraco из позднего карбона ( московско - гжельского периода ) Северной Америки, который представляет собой единственных известных парарептилий каменноугольного периода, что указывает на то, что первоначальная диверсификация группы произошла в позднем карбоне. [ 1 ] Многочисленные линии парарептилий появились в ранней перми, и группа достигла космополитического распространения. Разнообразие парарептилий сократилось к концу перми, и проколофоноиды , впервые появившиеся в поздней перми, были единственной группой парарептилий, пережившей пермско-триасовое вымирание . Разнообразие проколофонид резко сократилось, начиная со среднего триаса, а к концу триаса группа вымерла. [ 26 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Манн А., МакДэниел Э.Дж., МакКолвилл Э.Р., Мэддин Х.К. « Carbonodraco lundi gen et sp. nov., старейшая парарептилия из Линтона, штат Огайо, и новые знания о ранней радиации рептилий» . Королевское общество открытой науки . 6 (11): 191191. Бибкод : 2019RSOS.... 691191M. дои : 10.1098/rsos.191191 . ПМК 6894558 . ПМИД 31827854 .
- ^ Бота-Бринк Дж., Смит Р.М. (1 сентября 2012 г.). «Палеобиология триасовых проколофонид, выведенная по микроструктуре костей». Comptes Рендус Палевол . 11 (6): 419–433. дои : 10.1016/j.crpv.2012.03.002 . ISSN 1631-0683 .
- ^ Jump up to: а б с д Цуджи Л.А., Мюллер Дж. (2009). «Собираем историю Parareptilia: филогения, разнообразие и новое определение клады» . Ископаемый рекорд . 12 (1): 71–81. дои : 10.1002/mmng.200800011 . ISSN 1860-1014 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Лаурин М., Рейш Р.Р. (1 февраля 1995 г.). «Переоценка ранней филогении амниот» (PDF) . Зоологический журнал Линнеевского общества . 113 (2): 165–223. дои : 10.1111/j.1096-3642.1995.tb00932.x . ISSN 0024-4082 .
- ^ Jump up to: а б с д и ж г час я дж Мюллер Дж., Цудзи Л.А. (сентябрь 2007 г.). «Слух, соответствующий импедансу, у палеозойских рептилий: свидетельства развитого сенсорного восприятия на ранней стадии эволюции амниот» . ПЛОС ОДИН . 2 (9): е889. Бибкод : 2007PLoSO...2..889M . дои : 10.1371/journal.pone.0000889 . ЧВК 1964539 . ПМИД 17849018 .
- ^ Jump up to: а б Риппель О., деБрага М. (декабрь 1996 г.). «Черепахи как диапсидные рептилии» (PDF) . Природа . 384 (6608): 453–455. Бибкод : 1996Natur.384..453R . дои : 10.1038/384453a0 . ISSN 1476-4687 . S2CID 4264378 .
- ^ Jump up to: а б с д и ж деБрага М., Риппель О. (1 июля 1997 г.). «Филогения рептилий и взаимоотношения черепах» . Зоологический журнал Линнеевского общества . 120 (3): 281–354. дои : 10.1111/j.1096-3642.1997.tb01280.x . ISSN 0024-4082 .
- ^ Jump up to: а б Ван З., Паскуаль-Аная Дж., Задисса А., Ли В., Ниимура И., Хуан З. и др. (июнь 2013 г.). «Проекты геномов черепах с мягким панцирем и зеленой морской черепахи дают представление о развитии и эволюции строения тела, специфичного для черепах» . Природная генетика . 45 (6): 701–706. дои : 10.1038/ng.2615 . ПМК 4000948 . ПМИД 23624526 .
- ^ Jump up to: а б с Дебрага М., Рейш Р.Р. (19 сентября 1996 г.). «Раннепермская рептилия Acleistorhinus pteroticus и ее филогенетическое положение» (PDF) . Журнал палеонтологии позвоночных . 16 (3): 384–395. дои : 10.1080/02724634.1996.10011328 . ISSN 0272-4634 .
- ^ МакДугалл М.Дж., Рейс Р.Р. «Первая запись о никтифруретидном парарептилии из ранней перми Северной Америки с обсуждением височных фенестраций парарептилий». Зоологический журнал Линнеевского общества . 172 (3): 616–630. дои : 10.1111/zoj.12180 .
- ^ Jump up to: а б с Форд Д.П., Бенсон Р.Б. (январь 2020 г.). «Филогения ранних амниот и сходство Parareptilia и Varanopidae» . Экология и эволюция природы . 4 (1): 57–65. дои : 10.1038/s41559-019-1047-3 . ПМИД 31900445 . S2CID 209673326 .
- ^ Jump up to: а б Цуджи Л.А., Мюллер Дж., Рейс Р.Р. (1 января 2012 г.). «Анатомия Emeroleter levis и филогения парарептилий никтеролетера» (PDF) . Журнал палеонтологии позвоночных . 32 (1): 45–67. дои : 10.1080/02724634.2012.626004 . ISSN 0272-4634 . S2CID 55268829 .
- ^ Хариди Ю., Макдугалл М.Дж., Рейс Р.Р. (13 декабря 2017 г.). «Нижняя челюсть раннепермской парарептилии Delorhynchus, первое свидетельство наличия у рептилии множественных зубчатых венчиков». Зоологический журнал Линнеевского общества . 184 (3): 791–803. doi : 10.1093/zoolinnean/zlx085 . ISSN 0024-4082 .
- ^ Берман Д.С., Рейш Р.Р., Скотт Д., Хенрици А.С., Сумида С.С., Мартенс Т. (ноябрь 2000 г.). «Раннепермская двуногая рептилия». Наука . 290 (5493): 969–972. Бибкод : 2000Sci...290..969B . дои : 10.1126/science.290.5493.969 . ПМИД 11062126 .
- ^ Олсон ЕС (1947). Рой СК (ред.). «Семейство Diadectidae и его влияние на классификацию рептилий» . Геология . Филдиана. 11 (1). Чикаго: Чикагский музей естественной истории: 1–53 . дои : 10.5962/bhl.title.3579 . Проверено 15 января 2015 г.
- ^ Готье Дж.А., Клюге А.Г., Роу Т. (1988). «Ранняя эволюция амниоты». В Бентоне М.Дж. (ред.). Филогения и классификация четвероногих, Том 1: Земноводные, Рептилии, Птицы. Специальный том Ассоциации систематики . Том. 35А. Кларендон Пресс. стр. 103–155.
- ^ Риппель О (1994). «Остеология Simosaurus gaillardoti и взаимоотношения зауроптеригий стволовой группы» . Филдиана Геология . 28 (1462): и-85.
- ^ Риппель О (1995). «Исследования формирования скелета у рептилий: последствия для взаимоотношений черепах». Зоология . 98 : 298–308.
- ^ Зардоя Р., Мейер А. (ноябрь 1998 г.). «Полный митохондриальный геном предполагает сходство черепах с диапсидом» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (24): 14226–14231. Бибкод : 1998PNAS...9514226Z . дои : 10.1073/pnas.95.24.14226 . ПМК 24355 . ПМИД 9826682 .
- ^ Ивабе Н., Хара Ю., Кумазава Ю., Сибамото К., Сайто Ю., Мията Т., Като К. (апрель 2005 г.). «Сестринская групповая связь черепах с кладой птиц и крокодилов, выявленная с помощью белков, кодируемых ядерной ДНК» . Молекулярная биология и эволюция . 22 (4): 810–813. дои : 10.1093/molbev/msi075 . ПМИД 15625185 .
- ^ Роос Дж., Аггарвал Р.К., Янке А. (ноябрь 2007 г.). «Расширенный митогеномно-филогенетический анализ дает новое представление об эволюции крокодилов и их выживании на границе мелового и третичного периодов». Молекулярная филогенетика и эволюция . 45 (2): 663–673. дои : 10.1016/j.ympev.2007.06.018 . ПМИД 17719245 .
- ^ Катсу Ю., Браун Э.Л., Гийетт Л.Дж., Игучи Т. (2009). «От филогеномики рептилий к геномам рептилий: анализ протоонкогенов c-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. дои : 10.1159/000297715 . ПМИД 20234127 . S2CID 12116018 .
- ^ Ли М.С. (декабрь 2013 г.). «Происхождение черепах: идеи филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. дои : 10.1111/jeb.12268 . ПМИД 24256520 . S2CID 2106400 .
- ^ Ли, Чун; Фрейзер, Николас К.; Риппель, Оливье; Ву, Сяо-Чун (август 2018 г.). «Триасовая стволовая черепаха с беззубым клювом» . Природа . 560 (7719): 476–479. дои : 10.1038/s41586-018-0419-1 . ISSN 0028-0836 . ПМИД 30135526 . S2CID 52067286 .
- ^ Симойнс, ТР; Каммерер, CF; Колдуэлл, штат Вашингтон; Пирс, ЮВ (2022 г.). «Последовательные климатические кризисы в глубоком прошлом привели к ранней эволюции и радиации рептилий» . Достижения науки . 8 (33): eabq1898. дои : 10.1126/sciadv.abq1898 . ПМЦ 9390993 . ПМИД 35984885 .
- ^ Макдугалл М.Дж., Броклхерст Н., Фребиш Дж. (март 2019 г.). «Видовое богатство и неравенство парарептилий в период массового вымирания в конце пермского периода» . Слушания. Биологические науки . 286 (1899): 20182572. doi : 10.1098/rspb.2018.2572 . ПМК 6452079 . ПМИД 30890099 .






