Диапаз
| Диапсид -рептилии Временный диапазон: Пенсильванский - присутствует ,
| |
|---|---|

| |
| Схема черепа Araeoscelidian Petrolacosaurus kansensis | |

| |
| Нил Крокодил ( Crocodylus niloticus ) | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Рептилия |
| Клада : | Ромерид |
| Клада : | Диапсида Осборн , 1903 |
| Подгруппы | |
| |
Диапсиды («две арки») представляют собой кладу сауропсидов , отличающиеся от более примитивных эвриптилей в присутствии двух отверстий, известных как височные фенестра , на каждой стороне их черепа . Самые ранние традиционно идентифицированные диапсиды, ареоселидианцы , появились около трехсот миллионов лет назад в течение позднего каменноугольного периода. [ 1 ] Все диапсиды, кроме наиболее примитивных в кладской ареоселидии, часто помещаются в клад -неодиапсиду . Диапсиды чрезвычайно разнообразны и включают птиц и все современные группы рептилий, в том числе черепахи , которые исторически считались вне группы. [ 2 ] Все современные рептилии и птицы размещаются в подкладе Neodiapsid Sauria . Несмотря на то, что некоторые диапедиды потеряли одну дыру (ящерицы), либо обоих отверстий (змей и черепах), либо имеют сильно реструктурированный череп (современные птицы), они все еще классифицируются как диапсиды, основанные на их происхождении. По меньшей мере 17 084 вида животных диапсидов существуют: 9 159 птиц, [ 3 ] и 7 925 змей, ящериц, туатары , черепах и крокодилов. [ 4 ]
Характеристики
[ редактировать ]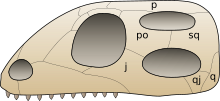
Название Diapsida означает «две арки», а диапсиды традиционно классифицируются на основе двух их предков ( височные фенестра ) выше и под глазом. Такое расположение позволяет прикреплять более сильные, более сильные мышцы челюсти и позволяет челюсти открываться более широко. Более неясной наследственной характеристикой является относительно длинная нижняя кость руки ( радиус ) по сравнению с костью верхней руки ( плечевая кость ).
Базально-нон-саурийские неодиазпиды были похожими на ящерицу, но базальные не-саурские неодиапсиды включают водные/амфибийные таксоны ( Claudiosaurus и некоторые тангасавриды ) [ 5 ] скользящая ящерица -Weigeltisauridae , [ 6 ] а также триасы, похожие на хамелеон, дропанозавры . [ 7 ]
Классификация
[ редактировать ]Диапсиды первоначально были классифицированы как один из четырех подклассов классовой рептилии , которые были основаны на количестве и расположении отверстий в черепе. Другие три подклассы были Synapsida (одно открытие на черепе, для «рептилий, похожих на млекопитающих»), Anapsida черепа, включая черепах и их родственников) и Euryapdid (без открытия морские рептилии). С появлением филогенетической номенклатуры эта система классификации была сильно модифицирована. Сегодня синапсиды часто не считаются истинными рептилиями, в то время как Euryapsida было обнаружено неестественной сборкой диапсидов, которые потеряли одно из их отверстий черепа. Генетические исследования и обнаружение триасовых паппушели показали, что это также имеет место в черепах, которые на самом деле являются сильно модифицированными диапсидами. В филогенетических системах птицы (потомки традиционных рептилий диапсидов) также считаются членами этой группы.
Некоторые современные исследования отношений рептилий предпочитают использовать название «Диапсид», чтобы обозначить группу короны всех современных рептилий диапсидов, но не их вымершие родственники. Тем не менее, многие исследователи также предпочитают более традиционное определение, которое включает доисторические ареоселидианцы . В 1991 году Лаурин определил Diapsida как кладу «самого последнего общего предка ареоселидийцев, лепидозавров и архозавров и всех его потомков». [ 8 ]
Кладе неодиапсида дали филогенетическое определение Лаурином в 1991 году. Он определил ее как кладку , основанную на ветвях , содержащую всех животных, более тесно связанных с « молодежными » (позже, более конкретно, подвергалась получению до Youngina Capensis ), чем с Петролакозавром (представление Araeoscelidia ) Полем [ 9 ] Самые ранние известные неодиапсиды, такие как Оренатор, известны с раннего перми, около 290 миллионов лет назад. [ 10 ]
Все генетические исследования подтвердили гипотезу о том, что черепахи являются рептилиями диапсидов; Некоторые разместили черепах в архозавройморфе, [ 11 ] [ 12 ] Или, чаще, как сестринская группа для существующих архозавров. [ 13 ] [ 14 ] [ 15 ] [ 16 ]
Современные рептилии и птицы размещаются в подкладе Neodiapsid Sauria , определяемой как последний общий предок Лепидозаурии (в том числе ящерицы, змеи и туатара), а также архозавриат (в том числе крокодильцы и динозавры, включая птиц). [ 17 ]
Кладистический анализ Лаурина и Пиньейро (2017) восстанавливает парарептилию в рамках Diapsida, с Pareiasaurs, Turtles, Millerettids и проколофоноидами, восстановленными как более полученные, чем базальная диапсида . [ 18 ] Исследование 2020 года, проведенное Дэвидом П. Фордом и Роджером Б.Дж. Бенсоном, также обнаружило парарептилию, как глубоко вложенные в Диапсиду, как сестринскую группу в неодиапсиду . Тем не менее, это исключает мезозавров , которые, как было признано базальными среди сауропсидов. [ 19 ] Другие недавние исследования показали, что более традиционное расположение парарептилов находится за пределами Diapsida. [ 17 ]
Положение высокоэтапных мезозойских морских групп рептилий Thalattosauria , Ichthyosauromorpha и Sauropterygia в Neodiapsida, неопределенно, и они могут лежать в Саурии. [ 17 ]
Отношения
[ редактировать ]Ниже приведены кладограммы, показывающие отношения основных групп диапсидов.
Кладограмма после Bickelmann et al. , 2009 [ 20 ] и Reis et al. , 2011: [ 21 ]
| ||||
В кладограмме Ли (2013) ниже использовалась комбинация генетических (молекулярных) и ископаемых (морфологических) данных. [ 22 ]
| Диапсида |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Эта вторая кладограмма основана на исследовании Pritchard и Nesbitt 2017 года. [ 23 ]
| Neodiapsida |
| |||||||||||||||||||||||||||||||||

Следующая кладограмма была обнаружена Simões et al . (2022): [ 17 ]
| Neoreptilia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Диапсида» . ucmp.berkeley.edu .
- ^ Schoch, Rainer R.; Sues, Hans-Dieter (2016). «Происхождение диапсидов черепах». Зоология . 119 (3): 159–161. Bibcode : 2016zool..119..159s . doi : 10.1016/j.zool.2016.01.004 . PMID 26934902 .
- ^ Барроукло, Джордж Ф.; Кракрафт, Джоэл; Клика, Джон; и Зинк, Роберт М. (23 ноября 2016 г.). Грин, Энди Дж. (Ред.). "Сколько там видов птиц и почему это имеет значение?" Полем Plos один . 11 (11): E0166307. Bibcode : 2016ploso..1166307b . doi : 10.1371/journal.pone.0166307 . PMC 5120813 . PMID 27880775 .
- ^ Ридер, Тод В.; Таунсенд, Тед М.; Малкахи, Даниэль Г.; Нунан, Брайс П.; Вуд, Перри Л. младший; Сайты, Джек В. младший; и Wiens, John J. (2015). Уилф, Питер (ред.). «Интегрированные анализы разрешают конфликты из -за филогения рептилий сквамата и выявляют неожиданные размещения для ископаемых таксонов» . Plos один . 10 (3): E0118199. BIBCODE : 2015PLOSO..1018199R . doi : 10.1371/journal.pone.0118199 . PMC 4372529 . PMID 25803280 .
- ^ Нуньес Демарко, Пол; Менхель, Мельтт; Лаурин, Мишель; Piñeiro, Graciela (2018-07-27). "Был ли Месосасурус полностью водной рептилий?" Полем Границы экологии и эволюции 6 : 109. doi : 10.3389/ fevo.2018.0 HDL : 20.500.12008/ 3 ISSN 2296-7
- ^ Причард, Адам С.; Суды, Ганс-Дитер; Скотт, Дайан; Рейс, Роберт Р. (2021-05-20). «Остеология, отношения и функциональная морфология Weigeltisaurus jaekeli (Diapsida, Weigeltisauridae) на основе полного скелета из верхнего пермского купфершифера Германии» . ПЕРЕЙ . 9 : E11413. doi : 10.7717/peerj.11413 . ISSN 2167-8359 . PMC 8141288 . PMID 34055483 .
- ^ Причард, Адам С.; Nesbitt, Sterling J. (октябрь 2017 г.). «Подобный птице череп в триасовой рептилии диапсида увеличивает гетерогенность морфологического и филогенетического излучения диапейда» . Королевское общество открыто наука . 4 (10): 170499. Bibcode : 2017rsos .... 470499p . doi : 10.1098/rsos.170499 . ISSN 2054-5703 . PMC 5666248 . PMID 29134065 .
- ^ Benton, MJ, Donoghue, PC, Asher, RJ, Friedman, M., Near, TJ, & Vinther, J. (2015). «Ограничения на шкалу эволюционной истории животных». Palaeontologia Electronica , 18.1.1fc; 1-106; palaeo-electronica.org/content/fc-1
- ^ Рейс, Роберт Р.; Модесто, Шон П.; Скотт, Дайан М. (22 декабря 2011 г.). «Новая ранняя пермская рептилия и ее значение в ранней эволюции диапсидов» . Труды Королевского общества B: Биологические науки . 278 (1725): 3731–3737. doi : 10.1098/rspb.2011.0439 . PMC 3203498 . PMID 21525061 .
- ^ Рейс, Роберт Р.; Модесто, Шон П.; Скотт, Дайан М. (2011-12-22). «Новая ранняя пермская рептилия и ее значение в ранней эволюции диапсидов» . Труды Королевского общества B: Биологические науки . 278 (1725): 3731–3737. doi : 10.1098/rspb.2011.0439 . ISSN 0962-8452 . PMC 3203498 . PMID 21525061 .
- ^ Ли, MSY (2013). «Происхождение черепах: понимание филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. doi : 10.1111/jeb.12268 . PMID 24256520 . S2CID 2106400 .
- ^ Маннен, Хидюки; Ли, Стивен С. -Л. (Октябрь 1999). «Молекулярные доказательства клады черепах». Молекулярная филогенетика и эволюция . 13 (1): 144–148. doi : 10.1006/mpev.1999.0640 . PMID 10508547 .
- ^ Zardoya, R.; Мейер, А. (1998). «Полный митохондриальный геном предполагает сродство диапсидов черепах» . Proc Natl Acad Sci USA . 95 (24): 14226–14231. Bibcode : 1998pnas ... 9514226Z . doi : 10.1073/pnas.95.24.14226 . ISSN 0027-8424 . PMC 24355 . PMID 9826682 .
- ^ Iwabe, n.; Hara, Y.; Kumazawa, Y.; Шибамото, К.; Saito, Y.; Miyata, T.; Като, К. (2004-12-29). «Сестринская групповая связь черепах с птичьей кладом, выявленной ядерными ДНК-кодированными белками» . Молекулярная биология и эволюция . 22 (4): 810–813. doi : 10.1093/molbev/msi075 . PMID 15625185 .
- ^ Роос, Джонас; Аггарвал, Рамеш К.; Янке, Аксель (ноябрь 2007 г.). «Расширенные митогеномные филогенетические анализы дают новое понимание эволюции крокодилиана и их выживаемости границы мела -третья». Молекулярная филогенетика и эволюция . 45 (2): 663–673. doi : 10.1016/j.ympev.2007.06.018 . PMID 17719245 .
- ^ Кацу, у.; Браун, Эль; Guillette, LJ Jr.; Игучи Т. (2010-03-17). «От рептильной филогеномики до геномов рептилий: анализ протоонкогенов C-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. doi : 10.1159/000297715 . PMID 20234127 . S2CID 12116018 .
- ^ Jump up to: а беременный в дюймовый Simões, Tiago R.; Kammerer, Christian F.; Caldwell, Michael W.; и Пирс, Стефани Э. (2022). «Последовательные климатические кризисы в глубоком прошлом привело к ранней эволюции и радиации рептилий» . Наука достижения . 8 (33): EABQ1898. Бибкод : 2022Scia .... 8.1898s . doi : 10.1126/sciadv.abq1898 . PMC 9390993 . PMID 35984885 .
- ^ Лаурин, Мишель; Пиньейро, Грасиэла Х. (2017). «Переоценка таксономического положения мезозавров и удивительная филогения ранних амниотов» (PDF) . Границы в науке Земли . 5 : 88. Bibcode : 2017freas ... 5 ... 88L . doi : 10.3389/feart.2017.00088 . S2CID 32426159 .
- ^ Ford DP, Benson RB (январь 2020 г.). «Филогения ранних амниотов и сродство парарептилий и варанопиды» . Природа экология и эволюция . 4 (1): 57–65. doi : 10.1038/s41559-019-1047-3 . PMID 31900445 . S2CID 209673326 .
- ^ Бикельманн, Констанц; Мюллер, Йоханнес; и Рейс, Роберт Р. (2009). «Загадочный диапсид Acerosodontosaurus piveteaui (Reptilia: Neodiapsida) из верхнего пермца Мадагаскара и парафилию молодых рептилий». Канадский журнал наук о Земле . 49 (9): 651–661. Bibcode : 2009cajes..46..651s . doi : 10.1139/e09-038 .
- ^ Рейс, Роберт Р.; Модесто, Шон П.; и Скотт, Дайан М. (2011). «Новая ранняя пермская рептилия и ее значение в ранней эволюции диапсидов» . Труды Королевского общества б . 278 (1725): 3731–7. doi : 10.1098/rspb.2011.0439 . PMC 3203498 . PMID 21525061 .
- ^ Ли, MSY (2013). «Происхождение черепах: понимание филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. doi : 10.1111/jeb.12268 . PMID 24256520 . S2CID 2106400 .
- ^ Причард, Адам С.; Nesbitt, Sterling J. (2017-10-11). «Подобный птице череп в триасовой рептилии диапсида увеличивает гетерогенность морфологического и филогенетического излучения диапейда» . Королевское общество открыто наука . 4 (10): 170499. Bibcode : 2017rsos .... 470499p . doi : 10.1098/rsos.170499 . ISSN 2054-5703 . PMC 5666248 . PMID 29134065 .
Внешние ссылки
[ редактировать ] Данные, связанные с диапсидом в Wikispecies
Данные, связанные с диапсидом в Wikispecies - Диапсида . Мишель Лаурин и Жак А. Готье. Веб -проект Tree of Life . 22 июня 2000 г.





Основные существующие рептилий клады |
|---|