Мезозавр
| Мезозавр Временной диапазон: | |
|---|---|
 | |
| Голотип Mesosaurus tenuidens (экз. MNHN 1865-77) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | † Парарептилии |
| Заказ: | † Мезозаврия |
| Семья: | † Мезозавриды |
| Род: | † Мезозавры Жерве , 1865 г. [1] |
| Разновидность: | † М. тонкий |
| Биномиальное имя | |
| † Мезозавр tenuidens Жерве, 1865 г. [1] | |
| Синонимы | |
Список | |
Мезозавр что означает «средняя ящерица») — вымерший род рептилий ( из ранней перми южной Африки и Южной Америки . Наряду с родами Brazilosaurus и Stereosternum он входит в семейство Mesosauridae и отряд Mesosauria . Долгое время мезозавр считался одной из первых морских рептилий , хотя новые данные позволяют предположить, что, по крайней мере, уругвайские особи населяли гиперсоленый водоем, а не типичную морскую среду. [3] В любом случае у него было множество приспособлений к полностью водному образу жизни. Обычно считается, что это был анапсид , хотя Фридрих фон Хюне считал его синапсидом . [4] Недавнее исследование филогении Mesosauridae определило эту группу либо как самую базальную кладу внутри Parareptilia , либо как самую базальную кладу внутри Sauropsida (причем последняя является менее поддерживаемой позицией). [5] несмотря на то, что череп мезозавра обладал «синапсидным состоянием» с одним височным окном . [6] [7]
Открытие и присвоение имени
[ редактировать ]Голотип , вероятно , , M. tenuidens MNHN 1865-77, получил прозвище « Гриква Мезозавр », и он был найден в хижине Гриква в Южной Африке в Кимберли, Северный Кейп, около 1830 года и использовался в качестве крышки от кастрюли. [1] [8] Обстоятельства его открытия и то, как он был отобран у предыдущих владельцев в Южной Африке, неизвестны, но известно то, что экземпляр в конце концов появился в коллекции французского палеонтолога Поля Жерве в 1860-х годах, и он обозначил его как голотип новый род и вид он назвал Mesosaurus tenuidens в 1865 году. [1]
С тех пор останки мезозавра также были обнаружены в Южной Америке и впервые были идентифицированы в 1908 году как принадлежащие ко второму виду, M. brasiliensis , Дж. Х. МакГрегором. [9] Более поздние исследования показали, что M. brasiliensis был тем же животным, что и M. tenuidens , который и по сей день остается единственным действующим видом мезозавра .
два других вида мезозавридов С тех пор были описаны : Stereosternum. [10] и бразилозавр , [11] которые также считаются синонимами Mesosaurus tenuidens согласно Piñeiro et al. (2021). [2]
Описание
[ редактировать ]У мезозавра был длинный череп, который был больше, чем у Stereosternum , и имел более длинные зубы. Зубы расположены под углом наружу, особенно на кончиках челюстей. [12]

Кости посткраниального скелета толстые, подвергшиеся пахиостозу . Мезозавр необычен среди рептилий тем, что у него есть клейтрум , обычно встречающийся у более примитивных костных рыб и четвероногих. Головка межключицы мезозавра треугольная, в отличие от голов других ранних рептилий, имеющих ромбовидную форму. [13]

Палеобиология
[ редактировать ]Мезозавр был одной из первых рептилий, которые, как известно, вернулись в воду после того, как ранние четвероногие вышли на сушу в позднем девоне или позже в палеозое. [14] Его длина составляла около 1 метра (3,3 фута), с перепончатыми лапами, обтекаемым телом и длинным хвостом, который, возможно, поддерживал плавник. Вероятно, он передвигался по воде с помощью своих длинных задних ног и гибкого хвоста. Его тело также было гибким и могло легко двигаться вбок, но у него были сильно утолщенные ребра , которые не позволяли ему скручивать тело. [15]
У мезозавра был небольшой череп с длинными челюстями . Ноздри располагались вверху, что позволяло существу дышать, при этом только верхняя часть его головы выходила на поверхность, как у современного крокодила . что зубы служили фильтрующим устройством для фильтрационного питания планктонных Первоначально считалось , организмов. [15] Однако эта идея основывалась на предположении, что зубы мезозавра были многочисленными и близко расположенными в челюстях. Недавно исследованные останки мезозавра показывают, что у него было меньше зубов и что зубной ряд был пригоден для ловли мелкой нектонной добычи, такой как ракообразные. [12]
Пахиостоз, наблюдаемый в костях мезозавра , возможно, позволил ему достичь нейтральной плавучести в верхних нескольких метрах толщи воды . Дополнительный вес мог стабилизировать животное на поверхности воды. Альтернативно, это могло бы придать мезозавру больший импульс при скольжении под водой. Хотя многие особенности предполагают полностью водный образ жизни, [16] Мезозавр , возможно, был способен перемещаться по суше на короткие периоды времени. Его локти и лодыжки были ограничены в движениях, из-за чего ходьба казалась невозможной. Более вероятно, что если бы мезозавр вышел на сушу, он бы продвигался вперед так же, как живые самки морских черепах, гнездящихся на пляжах. [13]
явно амниотного Ископаемые эмбрионы мезозавра ) типа на поздней стадии развития (т.е. зародыши были обнаружены в Уругвае и Бразилии. Эти окаменелости являются самым ранним свидетельством зародышей амниот, хотя предполагается, что амниоты имели свою типичную репродуктивную стратегию с момента их первого появления в позднем карбоне . До их описания самые старые известные зародыши амниот происходили из триаса . Один изолированный свернутый плод под названием FC-DPV 2504 не окружен известковой яичной скорлупой, что позволяет предположить, что железы в яйцеводе мезозавра и , вероятно, всех палеозойских амниот не были способны секретировать карбонат кальция , в отличие от постпалеозойских архозавров . Это могло бы объяснить нехватку окаменелостей яиц в палеозойской летописи окаменелостей амниотов. Один экземпляр мезозавра под названием MCN-PV 2214 представляет собой взрослую особь среднего размера с маленькой особью в грудной клетке , которая интерпретируется как плод « внутриутробно », что даже позволяет предположить, что мезозавр , как и многие другие морские рептилии , родил живо. Если эта интерпретация верна, этот образец будет представлять собой самый ранний известный пример Живорождение в летописи окаменелостей. Изолированный плод FC-DPV 2504, однако, скорее указывает на яйцеживородящую стратегию размножения у мезозавра . [17]
Исследование пропорций позвоночного столба показало, что, хотя молодые мезозавры могли вести полностью водный образ жизни, взрослые животные некоторое время проводили на суше. Это подтверждается редкостью взрослых животных в водных условиях и наличием у копролита высыхающих трещин. Однако насколько наземными были эти животные, сказать сложно, поскольку их пахиостоз и другие приспособления к водному образу жизни затруднили бы добычу пищи на суше. [18]
Распределение
[ редактировать ]Мезозавр сыграл важную роль в предоставлении доказательств теории дрейфа континентов , поскольку его останки были найдены в южной Африке, формации Уайтхилл и восточной части Южной Америки ( формация Мело , Уругвай и формация Ирати , Бразилия), двух широко разделенных регионах. [19] [20] Поскольку мезозавр был прибрежным животным и, следовательно, с меньшей вероятностью пересек Атлантический океан , такое распределение указывало на то, что два континента раньше были соединены вместе.
Галерея
[ редактировать ] Ископаемое из Бразилии
Ископаемое из Бразилии Двойное ископаемое в Луизиане
Двойное ископаемое в Луизиане Реставрация
Реставрация Из Бразилии, в Музее естественной истории в Вене.
Из Бразилии, в Музее естественной истории в Вене.![Ранняя реконструкция скелета M. brasiliensis, показывающая множество мелких зубов в челюстях (MacGregor, 1908).[9]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e7/Mesosaurus.png/120px-Mesosaurus.png) Ранняя реконструкция скелета M. brasiliensis, показывающая множество мелких зубов в челюстях (MacGregor, 1908). [9]
Ранняя реконструкция скелета M. brasiliensis, показывающая множество мелких зубов в челюстях (MacGregor, 1908). [9] Ископаемое невылупившейся молоди или плода Mesosaurus tenuidens (FC-DPV 2504) из Уругвая
Ископаемое невылупившейся молоди или плода Mesosaurus tenuidens (FC-DPV 2504) из Уругвая Скелетные формы в белесых выветриваемых сланцах формации Уайтхилл, Китмансхуп , Намибия.
Скелетные формы в белесых выветриваемых сланцах формации Уайтхилл, Китмансхуп , Намибия.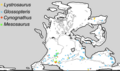 Распространение четырех пермских и триасовых групп окаменелостей, используемых в качестве биогеографических свидетельств дрейфа континентов и образования сухопутных мостов. Местоположение останков мезозавра показано зелеными квадратами.
Распространение четырех пермских и триасовых групп окаменелостей, используемых в качестве биогеографических свидетельств дрейфа континентов и образования сухопутных мостов. Местоположение останков мезозавра показано зелеными квадратами.







Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Жерве, П. (1865). Описание Mesosaurus tenuidens . Ископаемая рептилия из Южной Африки. Академия наук и литературы Монпелье. Мемуары секции наук 6 (2): 169-175
- ^ Перейти обратно: а б с д Пинейро, Г.; Фериголо, Дж.; Монес, А.; Нуньес Демарко, П. (2021). «Переоценка таксономии мезозавров: Stereosternum и Brazilosaurus действительны ли таксоны ?» . Бразильский журнал палеонтологии . 24 (3): 205–235. дои : 10.4072/rbp.2021.3.04 . hdl : 11336/175571 . S2CID 244188443 .
- ^ Пинейро, Г .; Рамос, А.; Госо, К.; Скарабино, Ф.; Лаурин, М. (2012). «Необычные условия окружающей среды сохранили пермского мезозавра Консерват-Лагерштетте из Уругвая» . Acta Palaeontologica Polonica . 57 (2): 299–318. дои : 10.4202/app.2010.0113 .
- ^ Хюне, Ф. фон (1940). «Остеология и систематическое положение мезозавра ». Палеонтографический отдел А. 92 : 45-58.
- ^ Лорин, Майкл; Пиньейро, Грасиела (2 ноября 2017 г.). «Переоценка таксономического положения мезозавров и удивительная филогения ранних амниот» . Границы в науках о Земле . 5 : 88. Бибкод : 2017FrEaS...5...88L . дои : 10.3389/feart.2017.00088 . hdl : 20.500.12008/33548 .
- ^ Пинейро, Грасиела (2008). «Мезозавры и другие окаменелости позднего палеозоя». В Д. Перере (ред.). Окаменелости Уругвая . ДИРАК, Монтевидео.
- ^ Пинейро, Г.; Фериголо, Дж.; Рамос, А.; Лаурин, М. (2012). «Краниальная морфология раннепермских мезозавридов Mesosaurus tenuidens и переоценка эволюции нижних височных окон». Comptes Рендус Палевол . 11 (5): 379–391. Бибкод : 2012CRPal..11..379P . дои : 10.1016/j.crpv.2012.02.001 .
- ^ Хелм, Чарльз и Бенуа, Жюльен. (2019). Геомифология Южной Африки. Исследовательские ворота 36.
- ^ Перейти обратно: а б МакГрегор, Дж. Х. (1908) Mesosaurus brasiliensis nov. сп. IN: Уайт, IC (1908) Комиссия по исследованиям угольных шахт Бразилии - Заключительный отчет; (Двуязычный отчет, португальский и английский), Imprensa Nacional, Рио-де-Жанейро, Бразилия, 617 стр.: Часть II, стр. 301–336.
- ^ Коуп, ЭД (1885). Вклад в палеонтологию позвоночных Бразилии. Труды Американского философского общества 25, 7-15.
- ^ Т. Шикама и Х. Одзаки (1966). «О скелете рептилии из палеозойской формации Сан-Паулу, Бразилия». Труды и труды Палеонтологического общества Японии . Новая серия. 64 : 351–358.
- ^ Перейти обратно: а б Модесто, СП (2006). «Черепной скелет раннепермской водной рептилии Mesosaurus tenuidens : значение для взаимоотношений и палеобиологии» . Зоологический журнал Линнеевского общества . 146 (3): 345–368. дои : 10.1111/j.1096-3642.2006.00205.x .
- ^ Перейти обратно: а б Модесто, СП (2010). «Посткраниальный скелет водной парарептилии Mesosaurus tenuidens из гондваанской перми». Журнал палеонтологии позвоночных . 30 (5): 1378–1395. Бибкод : 2010JVPal..30.1378M . дои : 10.1080/02724634.2010.501443 . S2CID 131561248 .
- ^ Лорен, Мишель (2010). Как позвоночные покинули воду (иллюстрировано под ред.). Издательство Калифорнийского университета. стр. xv + 199. ISBN 978-0-520-26647-6 .
- ^ Перейти обратно: а б Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 65. ИСБН 978-1-84028-152-1 .
- ^ Кановиль, Аврора; Мишель Лорен (2010). «Эволюция микроанатомии плечевой кости и образа жизни амниот и некоторые комментарии к палеобиологическим выводам» . Биологический журнал Линнеевского общества . 100 (2): 384–406. дои : 10.1111/j.1095-8312.2010.01431.x .
- ^ Пинейро, Г.; Фериголо, Дж.; Менегель, М.; Лаурин, М. (2012). «Самые старые известные амниотические эмбрионы предполагают живорождение у мезозавров». Историческая биология . 24 (6): 620–630. Бибкод : 2012HBio...24..620P . дои : 10.1080/08912963.2012.662230 . S2CID 59475679 .
- ^ Пабло Нуньес Демарко и др. Был ли мезозавр полностью водной рептилией? Передний. Экол. Evol, опубликовано в Интернете 27 июля 2018 г.; дои : 10.3389/fevo.2018.00109
- ^ Пинейро, Грасиела (2008). Д. Перера (ред.). Окаменелости Уругвая ДИРАК, Монтевидео.
- ^ Тревик, Стив (2016). «Тектоника плит в биогеографии». Международная географическая энциклопедия: Люди, Земля, окружающая среда и технологии . John Wiley & Sons, Ltd., стр. 1–9. дои : 10.1002/9781118786352.wbieg0638 . ISBN 9781118786352 .
Дальнейшее чтение
[ редактировать ]- Паркер, Стив. Динозавр: полное руководство по динозаврам. Firefly Books Inc, 2003. Стр. 90
- Кэрролл, РЛ (1988). Палеонтология и эволюция позвоночных . WH Фриман и компания. ISBN 9780716718222 .
- ЛеГранд, Гомер Юджин (1988). Дрейфующие континенты и меняющиеся теории: современная революция в геологии и научные изменения (иллюстрированное издание). Издательство Кембриджского университета. стр. 313 . ISBN 978-0-521-31105-2 .
- Маргулис, Линн ; Клиффорд Мэтьюз; Аарон Хазелтон (2000). Эволюция окружающей среды: последствия происхождения и эволюции жизни на планете Земля . Автор Клиффорд Мэтьюз, Аарон Хазелтон (2-е изд.). МТИ Пресс. п. 338. ИСБН 978-0-262-63197-6 .
- Сепкоски, Джек (2002). «Сборник родов ископаемых морских животных» . Бюллетени американской палеонтологии . 363 : 5–560. ISBN 978-0-87710-450-6 .




- Доисторические морские рептилии
- Роды доисторических рептилий
- Пермские рептилии Африки
- Приуральская жизнь
- Окаменелости Намибии
- Окаменелости Южной Африки
- Пермские рептилии Южной Америки
- Пермская Бразилия
- Окаменелости Бразилии
- Пермский Уругвай
- Окаменелости Уругвая
- Бассейн Параны
- Ископаемые таксоны, описанные в 1865 году.
- Таксоны, названные Полем Жерве