Гетеродонтозавр
| Гетеродонтозавр Временной диапазон: ранняя юра ,
| |
|---|---|

| |
| Слепок экземпляра SAM-PK-K1332, Палеонтологический музей Калифорнийского университета. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Семья: | † Гетеродонтозавриды |
| Род: | † Гетеродонтозавр Кромптон и Чариг, 1962 г. |
| Разновидность: | † Х. туки
|
| Биномиальное имя | |
| † Гетеродонтозавр туки Кромптон и Чариг, 1962 г.
| |
| Синонимы | |
| |
Heterodontosaurus) — род динозавров гетеродонтозаврид - , живших в раннем юрском периоде , 200–190 миллионов лет назад. Единственный известный вид , Heterodontosaurus tucki , был назван в 1962 году на основе черепа, обнаруженного в Южной Африке . Название рода означает «разнозубая ящерица» в связи с ее необычным гетеродонтным зубным рядом; видовое название дано в честь Г.К. Така, который поддержал первооткрывателей. С тех пор были найдены и другие экземпляры, в том числе почти полный скелет в 1966 году.
Хотя это был небольшой динозавр, Heterodontosaurus был одним из крупнейших представителей своего семейства , достигая от 1,18 м (3 фута 10 дюймов) до, возможно, 1,75 м (5 футов 9 дюймов) в длину и веся от 2 до 10 кг (4,4 дюйма). и 22,0 фунта). Череп был удлиненным, узким и треугольным, если смотреть сбоку. Передняя часть челюстей была покрыта роговым клювом. У него было три типа зубов; в верхней челюсти за маленькими резцеобразными зубами следовали длинные клыкообразные клыки. Щель отделяла бивни от долотообразных щечных зубов. Тело было коротким с длинным хвостом. Пятипалые передние конечности были длинными и относительно крепкими, тогда как задние конечности были длинными, тонкими и имели четыре пальца.
Heterodontosaurus — одноименный и самый известный представитель семейства Heterodontosauridae. Это семейство считается базовой (или «примитивной») группой отряда птицетазовых динозавров, хотя их ближайшее родство внутри группы обсуждается. Несмотря на большие бивни, гетеродонтозавр считается травоядным или, по крайней мере, всеядным . Хотя раньше считалось, что он способен передвигаться на четвероногих ногах , сейчас считается, что он был двуногим . Замена зубов носила спорадический и не постоянный характер, в отличие от своих родственников. По крайней мере четыре других рода гетеродонтозаврид известны из тех же геологических формаций, что и Heterodontosaurus .
История открытия
[ редактировать ]
Голотип Heterodontosaurus tucki (SAM-PK- K337 ) был обнаружен во время британско-южноафриканской экспедиции в Южную Африку и Басутоленд (прежнее название Лесото ) в 1961–1962 годах. Сегодня он находится в Южноафриканском музее Изико . Он был раскопан на горе на высоте около 1890 м (6201 фут), в местности под названием Тюининдини, в округе Транскей (иногда называемом Гершель ) в Капской провинции Южной Африки. Образец представляет собой раздробленный, но почти целый череп; связанные посткраниальные останки, упомянутые в первоначальном описании, не удалось обнаружить в 2011 году. Животное было научно описано и названо в 1962 году палеонтологами Альфредом Уолтером Кромптоном и Аланом Дж. Чаригом . Название рода относится к зубам различной формы, а видовое название дано в честь Джорджа К. Така, директора Austin Motor Company , который поддерживал экспедицию. К моменту публикации экземпляр не был полностью подготовлен, поэтому были описаны только передние части черепа и нижняя челюсть, причем авторы признали, что их описание было предварительным и служило главным образом для названия животного. Это открытие считалось важным, поскольку немногие ранние птицетазовые В то время были известны динозавры. Подготовка образца, т. е. освобождение костей от матрицы породы, занимала очень много времени, так как они были покрыты тонким, очень твердым железистым слоем, содержащим гематит . Его можно было удалить только алмазной пилой , которая повредила образец. [ 1 ] [ 2 ] [ 3 ] [ 4 ]

В 1966 году второй экземпляр гетеродонтозавра (SAM-PK-K1332) был обнаружен в местности Войизане, в формации Эллиот группы Стормберг скальных образований , на высоте 1770 м (5807 футов) над уровнем моря, на горе Кроммеспруит . Этот экземпляр включал в себя как череп, так и скелет, сохранившиеся в сочленении (т.е. кости сохранились в естественном положении по отношению друг к другу), с небольшим смещением и искажением костей. Посткраниальный скелет был кратко описан палеонтологами Альбертом Санта Лукой, Кромптоном и Чаригом в 1976 году. Кости его передних конечностей ранее обсуждались и фигурировали в статье палеонтологов Питера Гальтона и Роберта Т. Баккера в 1974 году, поскольку этот экземпляр считался важным в установив, что Dinosauria была монофилетической природной группой, тогда как большинство ученых того времени, включая ученых, описавших Heterodontosaurus , считали, что два основных отряда Saurischia и Ornithischia не связаны напрямую. [ 5 ] [ 6 ] Скелет был полностью описан в 1980 году. [ 7 ] SAM-PK-K1332 — наиболее полный скелет гетеродонтозаврид, описанный на сегодняшний день. [ 8 ] Хотя более подробное описание черепа гетеродонтозавра было обещано давно, оно так и не было опубликовано после смерти Чарига в 1997 году. [ 9 ] Лишь в 2011 году череп был полностью описан палеонтологом Дэвидом Б. Норманом и его коллегами. [ 1 ]

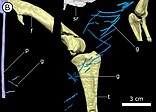
Другие экземпляры, относящиеся к Heterodontosaurus, включают переднюю часть черепа молодого человека (SAM-PK-K10487), фрагментированную верхнюю челюсть (SAM-PK-K1326), левую верхнюю челюсть с зубами и прилегающими костями (SAM-PK-K1334), все которые были собраны в местонахождении Войизане во время экспедиций 1966–1967 гг., хотя первый был идентифицирован только как принадлежащий к этому роду. в 2008 году. Частичная морда (NM QR 1788), найденная в 1975 году на ферме Тушиелау к югу от Войизане, считалась принадлежащей Massospondylus до 2011 года, когда она была реклассифицирована как Heterodontosaurus . Палеонтолог Роберт Брум обнаружил частичный череп, возможно, в формации Кларенс в Южной Африке, который был продан Американскому музею естественной истории в 1913 году как часть коллекции, почти полностью состоящей из синапсид окаменелостей . Этот экземпляр (AMNH 24000) был впервые идентифицирован как принадлежащий полувзрослому гетеродонтозавру Серено, который сообщил об этом в монографии 2012 года о Heterodontosauridae, первой всеобъемлющей обзорной статье. о семье. [ 3 ] [ 10 ] В этом обзоре частичный посткраниальный скелет (SAM-PK-K1328) из Войизане также классифицирован как Heterodontosaurus . Однако в 2014 году Гальтон предположил, что он может принадлежать к родственному роду Pegomastax , который Серено назвал на основе части черепа из того же места. [ 8 ] В 2005 году новый экземпляр гетеродонтозавра (AM 4766) был найден в русле реки недалеко от Грэхэмстауна в Восточно-Капской провинции ; он был очень целым, но камни вокруг него было слишком трудно полностью удалить. Поэтому в 2016 году образец был отсканирован в Европейской установке синхротронного излучения , чтобы помочь выявить скелет и помочь в исследовании его анатомии и образа жизни, некоторые из которых были опубликованы в 2021 году. [ 11 ] [ 12 ] [ 13 ]
В 1970 году палеонтолог Ричард А. Талборн предположил, что Heterodontosaurus был младшим синонимом рода Lycorhinus , который был назван в 1924 году вместе с видом L. angustidens , также по экземпляру, обнаруженному в Южной Африке. Он реклассифицировал типовой вид как член более старого рода, как новую комбинацию Lycorhinus tucki , которую он считал отдельной из-за небольших различий в ее зубах и стратиграфии. [ 14 ] Он повторил это утверждение в 1974 году при описании третьего вида Lycorhinus , Lycorhinus consors , после критики синонимии со стороны Гальтона в 1973 году. [ 15 ] В 1974 году Чариг и Кромптон согласились, что Heterodontosaurus и Lycorhinus принадлежали к одному семейству Heterodontosauridae, но не согласились с тем, что они достаточно похожи, чтобы их можно было считать родственными. Они также отметили, что фрагментарный характер и плохая сохранность экземпляра голотипа Lycorhinus angustidens не позволили полностью правильно сравнить его с H. tucki . [ 16 ] Несмотря на разногласия, ни одна из сторон не исследовала голотип L. angustidens из первых рук, но после этого палеонтолог Джеймс А. Хопсон также выступил в защиту разделения родов Heterodontosaurus в 1975 году и переместил L. consors в отдельный род Abrictosaurus . [ 17 ]
Описание
[ редактировать ]
Гетеродонтозавр был небольшим динозавром. Самый полный скелет, SAM-PK-K1332, принадлежал животному длиной около 1,18 м (3 фута 10 дюймов). В отдельных исследованиях его вес оценивался по разным оценкам в 1,8 кг (4,0 фунта), 2,59 кг (5,7 фунта) и 3,4 кг (7,5 фунта). [ 18 ] [ 8 ] Закрытие позвоночных швов на скелете указывает на то, что экземпляр был взрослым и, вероятно, вполне взрослым. Второй экземпляр, состоящий из неполного черепа, указывает на то, что гетеродонтозавр мог вырасти значительно крупнее — до длины 1,75 м (5 футов 9 дюймов) и с массой тела почти 10 кг (22 фунта). Причина разницы в размерах между двумя экземплярами неясна и может отражать изменчивость внутри одного вида, половой диморфизм или присутствие двух отдельных видов. [ 8 ] [ 19 ] Размер этого динозавра сравнивают с размером индейки . [ 20 ] Heterodontosaurus был одним из крупнейших известных представителей семейства Heterodontosauridae . [ 21 ] североамериканские Fruitadens достигали длины всего от 65 до 75 см (от 26 до 30 дюймов). В семейство входят одни из самых маленьких известных птицетазовых динозавров — например, [ 22 ] [ 23 ]
После описания родственного Tianyulong в 2009 году, который сохранился с сотнями длинных нитевидных покровов (иногда сравниваемых с щетинками ) от шеи до хвоста, Heterodontosaurus также изображался с такими структурами, например, в публикациях палеонтологов Грегори С. Пол и Пол Серено . Серено заявил, что гетеродонтозавр при жизни мог выглядеть как «проворный двуногий дикобраз ». [ 8 ] [ 24 ] Реставрация, опубликованная Серено, также показала гипотетическую структуру , расположенную на морде, над носовой ямкой (впадиной). [ 25 ]
Череп и зубной ряд
[ редактировать ]
Череп гетеродонтозавра был небольшим, но прочным. Два наиболее полных черепа имели длину 108 мм (4 дюйма) ( образец голотипа SAM-PK-K337) и 121 мм (5 дюймов) (образец SAM-PK-K1332). Череп был удлиненным, узким и треугольным, если смотреть сбоку, самой высокой точкой которого был сагиттальный гребень , откуда череп спускался к кончику морды. Задняя часть черепа имела крючковидную форму, смещенную к квадратной кости . Орбита (глазное отверстие) была большой и круглой, а большая шпорообразная кость, глазная кость , выступала назад в верхнюю часть отверстия. Ниже глазницы скуловая кость образовывала выступающий вбок выступ или роговидную структуру. Скуловая кость также образовывала «лезвие», которое вместе с выступом на крыловидной кости образовывало прорезь для направления движения нижней челюсти. Вентрально предглазничная ямка была ограничена выступающим костным гребнем, к которому должна была прикрепляться мясистая щека животного. [ 1 ] Также было высказано предположение, что гетеродонтозавры и другие базальные (или «примитивные») орхитисхи имели структуры, похожие на губы, как у ящериц (на основании сходства их челюстей), а не соединительную кожу между верхней и нижней челюстями (например, щеки). [ 26 ] Пропорционально большое нижнее височное окно имело яйцевидную форму, было наклонено назад и располагалось позади глазного отверстия. Эллиптическое верхнее височное окно было видно только при взгляде на верхнюю часть черепа. Левое и правое верхние височные окна были разделены сагиттальным гребнем, который обеспечивал боковые поверхности прикрепления челюстной мускулатуры у живого животного. [ 1 ]

Нижняя челюсть сужалась вперед, а зубная кость (основная часть нижней челюсти) была прочной. Передняя часть челюстей была покрыта беззубым ороговевшим клювом (или рамфотекой). Верхний клюв прикрывал переднюю часть предчелюстной кости, а нижний — предчелюстных костей , которые являются соответственно передними костями верхней и нижней челюсти у птицетазовых. Об этом свидетельствуют шероховатые поверхности этих конструкций. Нёбо было узким и сужалось к передней части. Наружные ноздри были небольшими, и верхняя граница этого отверстия, по-видимому, не была полностью перекрыта костью. Если это не произошло из-за перелома, возможно, разрыв образовался из соединительной ткани, а не из кости. В предглазничной ямке, большом углублении между глазным и ноздрейным отверстиями, было два отверстия меньшего размера. Впадина над мордой получила название «носовая ямка» или «борозда». Подобная ямка также наблюдается у Tianyulong , Agilisaurus и Eoraptor , но ее функция неизвестна. [ 22 ] [ 1 ] [ 25 ]
Необычной особенностью черепа были зубы разной формы ( гетеродонтия ), в честь которых назван род, который в основном известен у млекопитающих. У большинства динозавров (да и у большинства рептилий ) в челюстях был один тип зубов, но у гетеродонтозавра их было три. Клювистый кончик рыла был беззуб, тогда как задняя часть предчелюстной кости верхней челюсти имела по три зуба с каждой стороны. Первые два верхних зуба были маленькими и имели конусообразную форму (сравнимы с резцами ), тогда как третий с каждой стороны был значительно увеличен, образуя выступающие, клыки похожие на клыки . Эти первые зубы, вероятно, были частично закрыты верхним клювом. Первые два зуба нижней челюсти также образовывали клыки, но были намного крупнее верхних. [ 22 ] [ 1 ]


Клыки имели мелкие зазубрины по заднему краю, но только нижние были зазубрены спереди. Одиннадцать высоких долотообразных щечных зубов располагались с каждой стороны задних частей верхней челюсти, которые были отделены от клыков большой диастемой (промежутком). Щечные зубы постепенно увеличивались в размерах, причем средние зубы были самыми крупными, а после этого момента уменьшались в размерах. Эти зубы имели толстый слой эмали на внутренней стороне и были приспособлены к износу ( гипсодонтия ), а также имели длинные корни, прочно встроенные в лунки. Бивни нижней челюсти входят в углубление в диастеме верхней челюсти. Щечные зубы нижней челюсти в целом соответствовали зубам верхней челюсти, хотя поверхность эмали у них располагалась наружу. Верхний и нижний ряды зубов были вставлены, что создавало «щечную выемку», также наблюдаемую у других птицетазовых животных. [ 22 ] [ 1 ] Несмотря на разные типы зубов, их гистология и микроструктура эмали не были сложными. Но в то время как эмаль истончалась по направлению к внешней поверхности зубов, одновременно с истончением эмали возникла толстая полоса износостойкого дентина, которая сформировала режущий гребень окклюзионной поверхности, который обычно заполняется эмалью. [ 27 ]
Посткраниальный скелет
[ редактировать ]
Шея состояла из девяти шейных позвонков , которые образовывали S-образную кривую, на что указывает форма тел позвонков на виде сбоку скелета. Тела передних шейных позвонков имеют форму параллелограмма , средних — прямоугольные, задних — трапециевидную форму. [ 22 ] [ 7 ] Туловище было коротким, состояло из 12 спинных и 6 сросшихся крестцовых позвонков. [ 22 ] Хвост был длинным по сравнению с телом; хотя он и не полностью известен, он, вероятно, состоял из 34–37 хвостовых позвонков. Спинной отдел позвоночника укреплен окостеневшими сухожилиями , начиная с четвертого спинного позвонка. Эта особенность присутствует у многих других птицетазовых динозавров и, вероятно, противодействует стрессу, вызванному изгибающими силами, действующими на позвоночник во время двуногого передвижения. В отличие от многих других птицетазовых, хвост Heterodontosaurus не имел окостеневших сухожилий и поэтому, вероятно, был гибким. [ 7 ]
Лопатка у была покрыта дополнительным элементом, надлопаткой, которая у динозавров известна только Parksosaurus . В области груди Heterodontosaurus обладал хорошо развитой парой стернальных пластинок , напоминавших таковые у теропод, но отличавшихся от гораздо более простых стернальных пластинок других птицетазовых. Грудинные пластины были соединены с грудной клеткой элементами, известными как грудинные ребра. В отличие от других птицетазовых это соединение было подвижным, что позволяло телу расширяться при дыхании. Гетеродонтозавр — единственное известное птицетазовое животное, обладавшее гастралиями (костными элементами в коже между пластинками грудины и лобком таза). Гастралии располагались в два продольных ряда, каждый из которых содержал около девяти элементов. [ 13 ] Таз был длинным и узким, а лобковая кость напоминала таковую у более продвинутых птицетазовых животных. [ 28 ] [ 1 ] [ 29 ]

Передние конечности были крепко построены. [ 2 ] и пропорционально длинный, составляющий 70% длины задних конечностей. Радиус ( предплечья составлял 70% длины плечевой кости кости предплечья). [ 22 ] Рука была большой, по длине приближалась к плечевой кости, и имела пять пальцев, приспособленных для хватания. [ 22 ] [ 2 ] Второй палец был самым длинным, за ним следовали третий и первый пальцы ( большой палец ). [ 22 ] Первые три пальца заканчивались большими и сильными когтями. Четвертый и пятый пальцы сильно редуцированы и, возможно, рудиментарны . Фаланговая формула , определяющая количество костей пальцев в каждом пальце, начиная с первого, была 2-3-4-3-2. [ 22 ]
Задние конечности были длинными, тонкими и заканчивались четырьмя пальцами, первый из которых ( большой палец ) не касался земли. Уникально для птицетазовых животных несколько костей голени и стопы были слиты: большеберцовая и малоберцовая кости срослись с верхними костями предплюсны ( таранной и пяточной ), образуя большеберцовую кость , тогда как нижние кости предплюсны срослись с плюсневыми костями , образуя предплюсне-плюсневую кость . [ 22 ] Это созвездие можно встретить и у современных птиц, у которых оно развилось независимо . [ 2 ] Большеберцовая кость была примерно на 30% длиннее бедренной кости . [ 22 ] Ногтевые кости пальцев ног были когтеобразными, а не копытными, как у более развитых птицетазовых животных. [ 8 ]
Классификация
[ редактировать ]
Когда он был описан в 1962 году, Heterodontosaurus был классифицирован как примитивный представитель Ornithischia, одного из двух основных отрядов Dinosauria (второй — Saurischia). Авторы обнаружили, что он наиболее похож на малоизвестные роды Geranosaurus и Lycorhinus , второй из которых до тех пор считался терапсидным стволовым млекопитающим из-за его зубного ряда. Они отметили некоторое сходство с орнитоподами и условно поместили новый род в эту группу. [ 4 ] Палеонтологи Альфред Ромер и Оскар Кун независимо друг от друга назвали семейство Heterodontosauridae в 1966 году семейством птицетазовых динозавров, включая Heterodontosaurus и Lycorhinus . [ 30 ] [ 31 ] [ 32 ] Вместо этого Тулборн считал этих животных гипсилофодонтидами , а не отдельным семейством. [ 14 ] Баккер и Гальтон признали, что Heterodontosaurus важен для эволюции птицетазовых динозавров, поскольку рисунок его рук был общим с примитивными ящеротазами и, следовательно, был примитивным или базальным для обеих групп. [ 6 ] Это оспаривалось некоторыми учеными, которые полагали, что эти две группы вместо этого развились независимо от предков « текодонтийных » архозавров , и что их сходство произошло из-за конвергентной эволюции. Некоторые авторы также предположили связь, например потомок/предок, между гетеродонтозавридами и фаброзавридами , которые оба являются примитивными птицетазами, а также с примитивными цератопсами , такими как пситтакозавры , хотя природа этих отношений обсуждалась. [ 7 ]

К 1980-м годам большинство исследователей считали гетеродонтозаврид отдельным семейством примитивных птицетазовых динозавров, но с неопределенным положением по отношению к другим группам внутри отряда. К началу 21 века преобладали теории о том, что это семейство было сестринской группой либо Marginocephalia (в которую входят пахицефалозавриды и цератопсы), либо Cerapoda (первая группа плюс орнитоподы), либо одной из самых базальных радиаций птицетазовых. , до раскола Genasauria ( в который входят производные птицетазовые). [ 1 ] Heterodontosauridae были определены Серено как клада в 1998 и 2005 годах, и у этой группы общие черты черепа, такие как три или меньше зубов в каждой предчелюстной кости, клыковидные зубы, за которыми следует диастема, и скуловой рог под глазом. [ 33 ] В 2006 году палеонтолог Сюй Син и его коллеги назвали кладу Heterodontosauriformes , в которую вошли Heterodontosauridae и Marginocephalia, поскольку некоторые особенности, ранее известные только у гетеродонтозавров, были также замечены в базальном цератопсовом роде Yinlong . [ 34 ]
Многие роды были отнесены к Heterodontosauridae с момента создания этого семейства, однако Heterodontosaurus остается наиболее полностью известным родом и служил основным ориентиром для этой группы в палеонтологической литературе. [ 8 ] [ 3 ] Кладограмма ниже показывает взаимосвязи внутри Heterodontosauridae и соответствует анализу Серено, 2012: [ 35 ]
| Гетеродонтозавриды |
| ||||||||||||||||||||||||

Гетеродонтозавриды существовали от позднего триаса до раннего мела и существовали не менее 100 миллионов лет. Они известны из Африки, Евразии и Америки, но большинство из них обнаружено на юге Африки. разделились на две основные линии Гетеродонтозавриды, по-видимому, к ранней юре ; один с зубами с низкой коронкой и один с зубами с высокой коронкой (включая Heterodontosaurus ). Члены этих групп разделены биогеографически : группа с низкой короной была обнаружена в районах, которые когда-то были частью Лавразии (северный массив), а группа с высокой короной - в районах, которые были частью Гондваны (южный массив). В 2012 году Серено выделил представителей последней группы в подсемейство отдельное Heterodontosaurinae . Гетеродонтозавр, по-видимому, является наиболее производным гетеродонтозавром из-за деталей его зубов, таких как очень тонкая эмаль, расположенная асимметрично. Уникальные особенности зубов и челюстей гетеродонтозаврин, по-видимому, являются специализацией для эффективной переработки растительного материала, а их уровень сложности сравним с уровнем более поздних птицетазовых животных. [ 35 ]
В 2017 году сходство между скелетами Heterodontosaurus и раннего теропода Eoraptor было использовано палеонтологом Мэтью Г. Бароном и его коллегами, чтобы предположить, что птицетазовых следует сгруппировать с тероподами в группу под названием Ornithoscelida . Традиционно тероподов объединяли с зауроподоморфами в группу Saurischia. [ 36 ] В 2020 году палеонтолог Поль-Эмиль Дьедонне и его коллеги предположили, что представители Heterodontosauridae были базальными маргиноцефалами, не образующими свою собственную естественную группу, а вместо этого постепенно приводящими к пахицефалозаврам, и, следовательно, были базальными членами этой группы. Эта гипотеза позволит уменьшить призрачную линию пахицефалозавров и вернуть происхождение орнитопод к ранней юре. Подсемейство Heterodontosaurinae считалось действительной кладой внутри Pachycephalosauria, содержащей Heterodontosaurus , Abrictosaurus и Lycorhinus . [ 37 ]
Палеобиология
[ редактировать ]Диета и функция бивней
[ редактировать ]
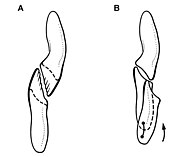
Гетеродонтозавр обычно считается травоядным динозавром. [ 38 ] В 1974 году Талборн предположил, что бивни динозавра не играли важной роли в питании; скорее, их использовали бы в бою с сородичами, для демонстрации, в качестве визуальной угрозы или для активной защиты. Подобные функции наблюдаются у увеличенных бивней современных мунтжаков и шевротанов , но изогнутые бивни бородавочников (используемые для рытья) различны. [ 15 ]
Несколько более поздних исследований выявили возможность того, что динозавр был всеяден и использовал свои бивни для убийства добычи во время периодической охоты. [ 1 ] [ 10 ] [ 39 ] В 2000 году Пол Барретт предположил, что форма предчелюстных зубов и мелкие зазубрины бивней напоминают хищных животных, намекая на факультативное хищничество. Напротив, у мунтжака нет зазубрин на бивнях. [ 39 ] В 2008 году Батлер и его коллеги утверждали, что увеличенные бивни сформировались на ранних стадиях развития особи и, следовательно, не могут представлять собой половой диморфизм. Таким образом, бой с сородичами является маловероятной функцией, поскольку увеличенные бивни можно было бы ожидать только у самцов, если бы они были инструментом для боя. Вместо этого более вероятны функции питания или защиты. [ 10 ] Также было высказано предположение, что гетеродонтозавр мог использовать свои скуловые выступы для нанесения ударов во время боя, и что глазная кость могла защищать глаза от таких атак. [ 40 ] В 2011 году Норман и его коллеги обратили внимание на руки, которые относительно длинные и оснащены большими загнутыми когтями. Эти особенности в сочетании с длинными задними конечностями, позволяющими быстро бегать, сделали животное способным схватывать мелкую добычу. Будучи всеядным, гетеродонтозавр имел бы значительное преимущество отбора в засушливый сезон, когда растительности было мало. [ 1 ]

В 2012 году Серено указал на несколько особенностей черепа и зубных рядов, которые предполагают чисто или, по крайней мере, преимущественно травоядную диету. К ним относятся роговой клюв и специальные щечные зубы (подходящие для срезания растительности), а также мясистые щеки, которые помогали удерживать пищу во рту во время жевания . Мышцы челюсти увеличены, челюстной сустав расположен ниже уровня зубов. Такое глубокое положение челюстного сустава позволяло равномерно распределять прикус по зубному ряду, в отличие от ножницеобразного прикуса, наблюдаемого у хищных динозавров. Наконец, размеры и положение бивней у отдельных представителей Heterodontosauridae весьма различны; Таким образом, специфическая функция питания кажется маловероятной. Серено предположил, что гетеродонтозавриды можно сравнить с современными пекариями , которые обладают похожими бивнями и питаются разнообразным растительным материалом, таким как корни, клубни, фрукты, семена и трава. [ 38 ] Батлер и его коллеги предположили, что аппарат питания Heterodontosaurus был специализирован для переработки жесткого растительного материала, и что поздно выжившие члены семейства ( Fruitadens , Tianyulong и Echinodon ), вероятно, демонстрировали более общий рацион, включающий как растения, так и беспозвоночных . Гетеродонтозавр характеризовался сильным укусом при небольших углах раскрытия зева, но более поздние представители были приспособлены к более быстрому укусу и более широкому раскрытию зева. [ 41 ] Исследование механики челюстей птицетазовых, проведенное в 2016 году, показало, что относительная сила укуса Heterodontosaurus была сопоставима с таковой у более продвинутого Scelidosaurus . Исследование показало, что бивни могли играть роль в питании, задевая нижнюю часть клюва во время сбора растительности. [ 42 ]
Замена зуба и эстивация
[ редактировать ]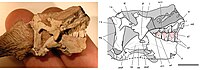
ли гетеродонтозавр Много споров вызвал вопрос о том, демонстрировал и в какой степени непрерывную замену зубов , которая типична для других динозавров и рептилий. В 1974 и 1978 годах Тулборн обнаружил, что черепа, известные на тот момент, не имели никаких признаков непрерывной замены зубов: щечные зубы известных черепов изношены равномерно, что указывает на то, что они образовались одновременно. Недавно прорезавшиеся зубы отсутствуют. Дополнительные доказательства были получены на основе стираемых фасеток зубов, которые образовались в результате контакта зубов нижнего и верхнего зубных рядов. Фасеты износа сливались друг с другом, образуя сплошную поверхность вдоль всего зубного ряда. Эта поверхность указывает на то, что процесс пищеварения осуществлялся движениями челюстей вперед и назад, а не простыми вертикальными движениями, как это было у родственных динозавров, таких как Фаброзавр . Движения вперед и назад возможны только в том случае, если зубы изношены равномерно, что еще раз усиливает аргументы в пользу отсутствия постоянной замены зубов. В то же время Тулборн подчеркнул, что регулярная замена зубов необходима этим животным, поскольку предполагаемая диета, состоящая из жесткого растительного материала, привела бы к быстрой стираемость зубов. Эти наблюдения привели Талборна к выводу, что гетеродонтозавр, должно быть, регулярно заменял весь свой набор зубов одновременно. Такая полная замена могла быть возможна только в периоды эстивации , когда животное не питалось. Летний период также соответствует предполагаемой среде обитания животных, которая напоминала пустыню, включая жаркие засушливые сезоны, когда еды было мало. [ 15 ] [ 43 ] [ 44 ]
Всесторонний анализ, проведенный в 1980 году Хопсоном, поставил под сомнение идеи Талборна. Хопсон показал, что рисунок фасеток стирания на зубах на самом деле указывает на вертикальные и боковые движения челюсти, а не вперед и назад. Кроме того, Хопсон продемонстрировал вариабельность степени износа зубов, что указывает на постоянную замену зубов. Он признал, что рентгеновские снимки наиболее полного экземпляра показали, что у этого человека действительно отсутствовали непрорезавшиеся сменные зубы. По мнению Хопсона, это указывало на то, что только у молодых особей постоянно менялись зубы, и что этот процесс прекращался по достижении взрослой жизни. Гипотеза эстивации Талборна была отвергнута Хопсоном из-за отсутствия доказательств. [ 44 ]
В 2006 году Батлер и его коллеги провели компьютерную томографию черепа подростка SAM-PK-K10487. К удивлению этих исследователей, сменные зубы, которые еще не прорезались, присутствовали даже на этой ранней онтогенетической стадии. Несмотря на эти результаты, авторы утверждали, что замена зубов должна была произойти, поскольку у молодых особей была та же морфология зубов, что и у взрослых особей — эта морфология изменилась бы, если бы зуб просто постоянно рос. В заключение Батлер и его коллеги предположили, что замена зубов у гетеродонтозавра , должно быть, была более спорадической, чем у родственных динозавров. [ 10 ] Непрорезавшиеся сменные зубы у гетеродонтозавра не были обнаружены до 2011 года, когда Норман и его коллеги описали верхнюю челюсть экземпляра SAM-PK-K1334. Еще один череп молодого человека (AMNH 24000), описанный Серено в 2012 году, также имел непрорезавшиеся сменные зубы. Как показали эти открытия, смена зубов у гетеродонтозавров носила эпизодический характер, а не непрерывный, как у других гетеродонтозаврид. Непрорезавшиеся зубы имеют треугольную форму при виде сбоку, что является типичной морфологией зубов у базальных птицетазовых. Таким образом, характерная долотообразная форма полностью прорезавшихся зубов возникла в результате контакта зубов между зубными рядами верхней и нижней челюстей. [ 1 ] [ 2 ] [ 38 ]
Передвижение, обмен веществ и дыхание.
[ редактировать ]
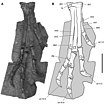

Хотя большинство исследователей сейчас считают гетеродонтозавра двуногим , бегуном [ 45 ] некоторые более ранние исследования предполагали частичное или полностью четвероногое передвижение. В 1980 году Санта-Лука описал несколько особенностей передней конечности, которые также присутствуют у современных четвероногих животных и подразумевают сильную мускулатуру рук: к ним относятся большой локтевой отросток (костный выступ, образующий самую верхнюю часть локтевой кости), увеличивающий плечо рычага предплечье. Медиальный надмыщелок плечевой кости был увеличен, обеспечивая места прикрепления сильных мышц- сгибателей предплечья. Более того, выступы на когтях могли увеличить толчок руки вперед во время ходьбы. По словам Санта-Луки, гетеродонтозавр был четвероногим, когда двигался медленно, но мог переключиться на гораздо более быстрый бег на двух ногах. [ 7 ] Палеонтологи Тереза Марьянска и Гальшка Осмольска поддержали гипотезу Санта-Луки в 1985 году; более того, они отметили, что у наиболее известного экземпляра дорсальный шип был сильно согнут вниз. [ 46 ] В 1987 году Грегори С. Пол предположил, что гетеродонтозавр мог быть обязательно четвероногим и что эти животные могли скакать галопом для быстрого передвижения. [ 47 ] Дэвид Вейшампель и Лоуренс Уитмер в 1990 году, а также Норман и его коллеги в 2004 году высказались в пользу исключительно двуногого передвижения, основанного на морфологии когтей и плечевого пояса . [ 22 ] [ 48 ] Анатомические данные, предложенные Санта-Лукой, были идентифицированы как приспособления для добывания пищи; крепкие и сильные руки могли использоваться для выкапывания корней и взлома гнезд насекомых. [ 22 ]

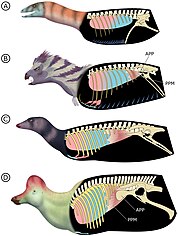
Большинство исследований рассматривают динозавров как эндотермических (теплокровных) животных с повышенным метаболизмом , сравнимым с метаболизмом современных млекопитающих и птиц. В исследовании 2009 года Герман Понцер и его коллеги рассчитали аэробную выносливость различных динозавров. Даже при умеренной скорости бега Heterodontosaurus превысил бы максимальные аэробные возможности, возможные для эктотермного (хладнокровного) животного, что указывает на эндотермию этого рода. [ 49 ]
Динозавры, вероятно, обладали системой воздушных мешков , как у современных птиц, которая обеспечивала вентиляцию неподвижных легких. Поток воздуха создавался за счет сокращения грудной клетки, что обеспечивалось подвижностью грудинных ребер и наличием гастралий. Расширения воздушных мешков также проникали в кости, образуя углубления и камеры — состояние, известное как посткраниальная скелетная пневмонития. У птицетазовых, за исключением Heterodontosaurus , отсутствовали подвижные грудинные ребра и гастралии, а у всех птицетазовых (включая Heterodontosaurus ) отсутствовала посткраниальная пневматическость скелета. Вместо этого у птицетазовых было заметное переднее расширение лобковой кости, передний лобковый отросток (APP), который отсутствовал у других динозавров. Основываясь на синхротронных данных хорошо сохранившегося экземпляра гетеродонтозавра (AM 4766), Виктор Радермахер и его коллеги в 2021 году предположили, что дыхательная система птицетазовых резко отличается от системы дыхания других динозавров и что гетеродонтозавр представляет собой промежуточную стадию. По мнению этих авторов, птицетазовые утратили способность сокращать грудную клетку для дыхания и вместо этого полагались на мышцу, которая непосредственно вентилировала легкие, которую они назвали полово-перитонеальная мышца . APP таза обеспечило бы место прикрепления этой мышцы. У гетеродонтозавра был зарождающийся APP, а его гастралии были уменьшены по сравнению с динозаврами, не являющимися птицетазовыми, что позволяет предположить, что таз уже участвовал в дыхании, а сокращение грудной клетки стало менее важным. [ 13 ]
Рост и предполагаемый половой диморфизм
[ редактировать ]
Онтогенез , , или развитие особи от ювенильного состояния к взрослому, у Heterodontosaurus плохо изучен поскольку молодые особи редки. Как показывает череп молоди SAM-PK-K10487, по мере роста животного глазницы становились пропорционально меньше, а морда становилась длиннее и содержала дополнительные зубы. Подобные изменения были зарегистрированы и у некоторых других динозавров. Однако морфология зубов не изменилась с возрастом, что указывает на то, что рацион молоди был таким же, как и у взрослых. Предполагаемая длина черепа молоди составляла 45 мм (2 дюйма). Если предположить, что пропорции тела такие же, как у взрослых особей, длина тела этого подростка должна была составлять 450 мм (18 дюймов). Действительно, особь, вероятно, была бы меньше, поскольку у молодых животных обычно голова пропорционально крупнее. [ 10 ]
В 1974 году Талборн предположил, что большие бивни гетеродонтозаврид представляют собой вторичный половой признак . Согласно этой теории, только взрослые особи мужского пола могли обладать полностью развитыми бивнями; голотип родственного абриктозавра , у которого вообще не было бивней, представлял собой самку. [ 15 ] Эта гипотеза была подвергнута сомнению палеонтологом Ричардом Батлером и его коллегами в 2006 году, которые утверждали, что молодой череп SAM-PK-K10487 обладал бивнями, несмотря на его раннюю стадию развития. В этом состоянии вторичных половых признаков не ожидается. Более того, бивни присутствуют почти во всех известных гетеродонтозавров черепах ; однако наличие полового диморфизма предполагает соотношение 50:50 между особями, имеющими бивни, и теми, у кого бивней нет. Единственным исключением является голотип абриктозавра ; отсутствие бивней у этой особи интерпретируется как специализация именно этого рода. [ 10 ]
Палеосреда
[ редактировать ]
Гетеродонтозавр известен по окаменелостям, найденным в формациях супергруппы Кару , включая формацию Верхний Эллиот и формацию Кларенс, которые датируются геттангийским и синемурийским возрастом нижней юры , около 200–190 миллионов лет назад . Первоначально гетеродонтозавр считалось, что произошел из верхнего триаса . Формация Аппер Эллиот состоит из красного/фиолетового аргиллита и красного/белого песчаника , тогда как более молодая формация Кларенс состоит из белого/кремового песчаника. Формация Кларенс менее богата окаменелостями, чем формация Аппер Эллиот; его отложения также часто образуют скалы, что ограничивает доступ для охотников за ископаемыми. [ 3 ] [ 4 ] Для формации Верхний Эллиот характерны животные, которые кажутся более легкими по телосложению, чем животные формации Нижний Эллиот, что, возможно, было адаптацией к более сухому климату на юге Африки в это время. Обе формации известны обилием окаменелостей позвоночных, в том числе темноспондиловых амфибий , черепах, лепидозавров , этозавров , крокодиломорфов , не являющихся млекопитающими и цинодонтов . [ 4 ] [ 50 ]
Другие динозавры из этих формаций включают геназавра Lesothosaurus , базального зауроподоморфа Massospondylus и теропода Megapnosaurus . Формация Аппер-Эллиот демонстрирует самое большое известное разнообразие гетеродонтозаврид среди всех горных пород; кроме Heterodontosaurus , он содержал Lycorhinus , Abrictosaurus и Pegomastax . Еще один член семейства, Геранозавр , известен из формации Кларанс. Высокое разнообразие гетеродонтозаврид привело исследователей к выводу, что разные виды могли питаться разными источниками пищи, чтобы избежать конкуренции ( разделение ниш ). [ 19 ] [ 41 ] Благодаря своему высокоспециализированному зубному ряду Heterodontosaurus мог специализироваться на жёстком растительном материале, в то время как менее специализированный Abrictosaurus мог преимущественно потреблять более мягкую растительность. [ 19 ] [ 41 ] Положение отдельных экземпляров гетеродонтозаврид в последовательности горных пород плохо известно, что затрудняет определение того, сколько из этих видов действительно были ровесниками и какие виды существовали в разное время. [ 19 ] [ 41 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л Норман, Д.Б.; Кромптон, штат Аризона; Батлер, Р.Дж.; Порро, Л.Б.; Чариг, AJ (2011). «Нижнеюрский птицетазовый динозавр Heterodontosaurus tucki Crompton & Charig, 1962: анатомия черепа, функциональная морфология, таксономия и взаимоотношения» . Зоологический журнал Линнеевского общества : 182–276. дои : 10.1111/j.1096-3642.2011.00697.x .
- ^ Перейти обратно: а б с д и Серен, ПК (2012). стр. 100-1 114–132.
- ^ Перейти обратно: а б с д Серен, ПК (2012). стр. 100-1 4–17.
- ^ Перейти обратно: а б с д Кромптон, штат Аризона; Чариг, AJ (1962). «Новый птицетаз из верхнего триаса Южной Африки». Природа . 196 (4859): 1074–1077. Бибкод : 1962Natur.196.1074C . дои : 10.1038/1961074a0 . S2CID 4198113 .
- ^ Санта-Лука, AP; Кромптон, штат Аризона; Чариг, AJ (1976). «Полный скелет позднетриасового птицетазового Heterodontosaurus tucki ». Природа . 264 (5584): 324–328. Бибкод : 1976Natur.264..324S . дои : 10.1038/264324a0 . S2CID 4283187 .
- ^ Перейти обратно: а б Баккер, RT; Гальтон, премьер-министр (1974). «Монофилия динозавров и новый класс позвоночных». Природа . 248 (5444): 168–172. Бибкод : 1974Natur.248..168B . дои : 10.1038/248168a0 . S2CID 4220935 .
- ^ Перейти обратно: а б с д и Санта-Лука, AP (1980). «Посткраниальный скелет Heterodontosaurus tucki (Reptilia, Ornithischia) из Штормберга в Южной Африке» . Анналы Южноафриканского музея . 79 (7): 159–211. ISSN 0303-2515 . OCLC 11886969 .
- ^ Перейти обратно: а б с д и ж г Гальтон, премьер-министр (2014). «Заметки о посткраниальной анатомии гетеродонтозавридного динозавра Heterodontosaurus tucki , базального птицетазового животного из нижней юры Южной Африки» (PDF) . Revue de Paléobiologie, Женева . 1. 33 : 97–141. ISSN 1661-5468 .
- ^ Муди, RTJ; Нэйш, Д. (2010). «Алан Джек Чариг (1927–1997): обзор его научных достижений и роли в мире исследований ископаемых рептилий». Геологическое общество, Лондон, специальные публикации . 343 (1): 89–109. Бибкод : 2010ГСЛСП.343...89М . дои : 10.1144/SP343.6 . S2CID 129586311 .
- ^ Перейти обратно: а б с д и ж Батлер, Р.Дж.; Порро, Л.Б.; Норман, Д.Б. (2008). «Молодой череп примитивного птицетазового динозавра Heterodontosaurus tucki из Штормберга на юге Африки». Журнал палеонтологии позвоночных . 28 (3): 702–711. doi : 10.1671/0272-4634(2008)28[702:AJSOTP]2.0.CO;2 . S2CID 86739244 .
- ^ «Окаменелость динозавра, найденная в ЮАР, наконец-то раскрывает свои тайны» . 27 июля 2016 г. Проверено 27 июля 2016 г.
- ^ «ESRF сканирует самый полный из когда-либо найденных скелетов гетеродонтозавра » . www.esrf.eu. 2016 . Проверено 27 июля 2016 г.
- ^ Перейти обратно: а б с Радермахер, В.Дж.; Фернандес, В.; Шахнер, ER; Батлер, Р.Дж.; Борди, EM; Нэйлор Хаджинс, М.; де Клерк, WJ; Шапель, KEJ; Шуаньер, Ж. Н. (2021). «Новый экземпляр гетеродонтозавра проливает свет на уникальную дыхательную макроэволюцию птицетазовых динозавров» . электронная жизнь . 10 : e66036. doi : 10.7554/eLife.66036 . ПМК 8260226 . ПМИД 34225841 .
- ^ Перейти обратно: а б Талборн, РА (1970). «Систематическое положение триасового птицетазового динозавра Lycorhinus angustidens ». Зоологический журнал Линнеевского общества . 49 (3): 235–245. дои : 10.1111/j.1096-3642.1970.tb00739.x .
- ^ Перейти обратно: а б с д Талборн, РА (1974). «Новый динозавр-гетеродонтозаврид (Reptilia: Ornithischia) из красных слоев верхнего триаса Лесото». Зоологический журнал Линнеевского общества . 55 (2): 151–175. дои : 10.1111/j.1096-3642.1974.tb01591.x .
- ^ Чариг, Эй Джей; Кромптон, AW (1974). «Предполагаемая синонимия Lycorhinus и Heterodontosaurus ». Анналы Южноафриканского музея . 64 : 167–189.
- ^ Хопсон, Дж. А. (1975). «О родовом выделении птицетазовых динозавров Lycorhinus и Heterodontosaurus из серии Штормберг (верхний триас) Южной Африки». Южноафриканский научный журнал . 71 : 302–305.
- ^ Сибахер, Ф. (2001). «Новый метод расчета аллометрических отношений длины и массы динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. CiteSeerX 10.1.1.462.255 . doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 . ISSN 0272-4634 . S2CID 53446536 .
- ^ Перейти обратно: а б с д Порро, Л.Б.; Батлер, Р.Дж.; Барретт, премьер-министр; Мур-Фэй, С.; Абель, Р.Л. (2011). «Новые экземпляры гетеродонтозаврид из нижней юры южной Африки и ранняя радиация птицетазовых динозавров» (PDF) . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 101 (Специальный выпуск 3–4): 351–366. дои : 10.1017/S175569101102010X . ISSN 1755-6929 . S2CID 128636991 . Архивировано из оригинала (PDF) 11 августа 2017 г. Проверено 30 декабря 2015 г.
- ^ Ламберт, Д. (1993). Самая лучшая книга о динозаврах . Нью-Йорк: Дорлинг Киндерсли. стр. 134–135 . ISBN 978-1-56458-304-8 .
- ^ Серено, ПК (2012). стр. 100-1 161–162.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Вейшампель, Д.Б.; Уитмер, LM (1990). «Гетеродонтозавриды». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия . Издательство Калифорнийского университета. стр. 486–497. ISBN 978-0-520-06726-4 .
- ^ Батлер, Р.Дж.; Гальтон, премьер-министр; Порро, Л.Б.; Кьяппе, LM; Хендерсон, DM; Эриксон, генеральный директор (2010). «Нижние пределы размера тела птицетазовых динозавров, выведенные на основе нового верхнеюрского гетеродонтозаврид из Северной Америки» . Труды Королевского общества B: Биологические науки . 277 (1680): 375–381. дои : 10.1098/rspb.2009.1494 . ISSN 0080-4649 . ПМЦ 2842649 . ПМИД 19846460 .
- ^ Издательство Пенсофт (2012). «Новый клыкастый карликовый динозавр из южной Африки питался растениями» . ЭврикаАлерт! . Проверено 28 января 2016 г.
- ^ Перейти обратно: а б Серен, ПК (2012). п. 219.
- ^ Набавизаде, А. (2018). «Новая реконструкция черепной мускулатуры птицетазовых динозавров: значение для механизмов питания и анатомии щеки» . Анатомическая запись . 303 (2): 347–362. дои : 10.1002/ar.23988 . ПМИД 30332723 . S2CID 52986589 .
- ^ Калверт, Сесилия; Хант, Тайлер; Уэлен, Найл; Шуаньер, Иона; Норелл, Марк; Эриксон, Грегори (2023). « Микроструктура эмали Heterodontosaurus tucki и гистология зубов» . Acta Palaeontologica Polonica . 68 . дои : 10.4202/app.01060.2023 .
- ^ Бентон, MJ (2012). Доисторическая жизнь . Эдинбург, Шотландия: Дорлинг Киндерсли. п. 271. ИСБН 978-0-7566-9910-9 .
- ^ Додсон, П.; Бритт, Б.; Карпентер, К.; Форстер, Калифорния; Джилетт, Д.Д.; Норелл, Массачусетс; Ольшевский Г.; Пэрриш, Дж. М.; Вейшампель, Д.Б., ред. (1 января 1993 г.). « Гетеродонтозавр ». Эпоха динозавров . Линкольнвуд: Publications International, LTD. п. 37. ИСБН 978-0-7853-0443-2 .
- ^ Серено, ПК (2012). стр. 100-1 29–30.
- ^ Ромер, А.С. (1966). Палеонтология позвоночных (3-е изд.). Чикаго: Издательство Чикагского университета. 468 стр . ISBN 978-0-7167-1822-2 .
- ^ Кун, О. (1966) Рептилии. Верлаг Обен, Крайлинг под Мюнхеном, 154 стр.
- ^ Серено, ПК (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Новогодний альбом по геологии и палеонтологии, Трактаты . 210 (1): 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Сюй, Х.; Форстер, Калифорния; Кларк, Дж. М.; Мо, Дж. (2006). «Базальный цератопс с переходными чертами из поздней юры северо-западного Китая» . Труды Королевского общества B: Биологические науки . 273 (1598): 2135–2140. дои : 10.1098/rspb.2006.3566 . ПМЦ 1635516 . ПМИД 16901832 .
- ^ Перейти обратно: а б Серено, ПК (2012). стр. 100-1 193–206.
- ^ Барон, МГ; Норман, Д.Б.; Барретт, премьер-министр (2017). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров» (PDF) . Природа . 543 (7646): 501–506. Бибкод : 2017Natur.543..501B . дои : 10.1038/nature21700 . ПМИД 28332513 . S2CID 205254710 .
- ^ Дьедонне, П.-Э.; Крузадо-Кабальеро, П.; Годфруа, П.; Тортоса, Т. (2020). «Новая филогения цераподских динозавров» . Историческая биология . 33 (10): 2335–2355. Бибкод : 2021HBio...33.2335D . дои : 10.1080/08912963.2020.1793979 .
- ^ Перейти обратно: а б с Серен, ПК (2012). стр. 100-1 162–193.
- ^ Перейти обратно: а б Барретт, премьер-министр (2000). «Прозауроподы динозавры и игуаны: размышления о рационе вымерших рептилий». В Сьюс, HD. (ред.). Эволюция травоядных у наземных позвоночных. Перспективы летописи окаменелостей . Издательство Кембриджского университета. стр. 42–78 . дои : 10.1017/CBO9780511549717.004 . ISBN 978-0-521-59449-3 .
- ^ Перенасыщение, Д.Ф. (1997). «Гетеродонтозавр». Динозавры, энциклопедия . Издательство McFarland & Company, Inc. стр. 467–469. ISBN 978-0-375-82419-7 .
- ^ Перейти обратно: а б с д Батлер, Ричард Дж; Порро, Лаура Б; Гальтон, Питер М; Кьяппе, Луис М (2012). «Анатомия и краниальная функциональная морфология мелкого динозавра Fruitadens haagarorum из верхней юры США» . ПЛОС ОДИН . 7 (4): e31556. Бибкод : 2012PLoSO...731556B . дои : 10.1371/journal.pone.0031556 . ПМЦ 3324477 . ПМИД 22509242 .
- ^ Набавизаде, А. (2016). «Эволюционные тенденции в механике приводящих челюстей птицетазовых динозавров» . Анатомическая запись . 299 (3): 271–294. дои : 10.1002/ar.23306 . ПМИД 26692539 .
- ^ Талборн, РА (1978). «Праздник среди динозавров-орнитопод африканского триаса». Летайя . 11 (3): 185–198. Бибкод : 1978Лета..11..185Т . дои : 10.1111/j.1502-3931.1978.tb01226.x .
- ^ Перейти обратно: а б Хопсон, Дж. А. (1980). «Функция и замена зубов у птицетазовых динозавров раннего мезозоя: последствия для летнего периода». Летайя . 13 (1): 93–105. Бибкод : 1980Лета..13...93Ч . дои : 10.1111/j.1502-3931.1980.tb01035.x .
- ^ Батлер, Р.Дж.; Барретт, премьер-министр (2012). «Орнитоподы». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Блумингтон и Индианаполис: Издательство Университета Индианы. п. 563 . ISBN 978-0-253-35701-4 .
- ^ Марьянская, Т.; Осмольска, Х. (1985). «О филогении птицетазовых» . Acta Palaeontologica Polonica . 30 (3–4): 137–150.
- ^ Пол, GS (1987). «Наука и искусство восстановления жизненного облика динозавров и их родственников: подробное практическое руководство» . В Черкасе, SJ; Олсон, ЕС (ред.). Динозавры, прошлое и настоящее . 2. Издательство Вашингтонского университета. стр. 4–49 . ISBN 978-0-295-96570-3 .
- ^ Норман, Д.Б .; Сьюс, Х.-Д.; Уитмер, Л.М.; Кориа, РА (2004). «Базальные орнитоподы». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 393–412 . ISBN 978-0-520-24209-8 .
- ^ Понцер, Х.; Аллен, В.; Хатчинсон, младший (2009). «Биомеханика бега указывает на эндотермию у прямоногих динозавров» . ПЛОС ОДИН . 4 (11): е7783. Бибкод : 2009PLoSO...4.7783P . дои : 10.1371/journal.pone.0007783 . ПМЦ 2772121 . ПМИД 19911059 .
- ^ Нолл, Ф. (2005). «Фауна четвероногих формаций Аппер-Эллиот и Кларенс в основном бассейне Кару (Южная Африка и Лесото)». Бюллетень геологического общества Франции . 176 : 81–91. дои : 10.2113/176.1.81 .
Цитируемые работы
[ редактировать ]- Серено, ПК (2012). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридных динозавров» . ZooKeys (226): 1 –225. дои : 10.3897/zookeys.226.2840 . ПМК 3491919 . ПМИД 23166462 .
Внешние ссылки
[ редактировать ]- Музей естественной истории: «Большие зубы для крошечного динозавра» — трехминутное видео о гетеродонтозавре, представленное Ричардом Батлером на YouTube
- Синхротрон ESRF: «Цифровой скелет гетеродонтозавра, произведенный в ESRF, показывающий полный образец и анатомию» - одноминутное видео о синхротронном сканировании на YouTube.