Гетеродонтозавриды
| Гетеродонтозавриды Временной диапазон: ранняя юра – ранний мел , Возможная запись позднего триаса
| |
|---|---|

| |
| Слепок экземпляра SAM-PK-K1332 Heterodontosaurus tucki , Палеонтологический музей Калифорнийского университета. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Сафорнитишия |
| Семья: | † Гетеродонтозавриды Ромер , 1966 (Кун, 1966 ) |
| Подгруппы [1] | |
| |
Heterodontosauridae — семейство птицетазовых , динозавров которые, вероятно, были одними из самых базальных (примитивных) членов группы. Их филогенетическое положение неясно, но чаще всего они оказываются примитивными, за пределами группы Genasauria . [2] Хотя их окаменелости относительно редки, а их группа небольшая по численности, они были обнаружены на всех континентах, кроме Австралии и Антарктиды , а их диапазон простирался от ранней юры до раннего мела .
Гетеродонтозавриды представляли собой динозавров размером с лисицу длиной менее 2 метров (6,6 футов), включая длинный хвост. Они известны главным образом своими характерными зубами, включая увеличенные клыкоподобные бивни и щечные зубы, приспособленные для жевания, аналогичные таковым у меловых гадрозаврид . Их диета была травоядной или, возможно, всеядной .
Описание
[ редактировать ]

Среди гетеродонтозаврид только сам Heterodontosaurus известен по полному скелету. Фрагментарные скелетные останки абриктозавра известны, но не были полностью описаны, в то время как большинство других гетеродонтозаврид известны только по фрагментам челюстей и зубов. Следовательно, большинство синапоморфий гетеродонтозаврид (определяющих особенностей) были описаны по зубам и костям челюсти. [3] [4] Гетеродонтозавр имел длину чуть более 1 метра (3,3 фута). [5] тогда как фрагментарные останки Lycorhinus могут указывать на более крупную особь. [6]
Тяньюлонг из Китая, по-видимому, сохранил нитевидный покров, который был интерпретирован как вариант протоперьев, обнаруженных у некоторых теропод. Эти нити имеют гребень вдоль хвоста. Наличие этого нитевидного покрова было использовано для предположения, что и птицетазовые, и ящеротазовые были эндотермными . [7]
Череп и зубы
[ редактировать ]И у абриктозавра , и у гетеродонтозавра были очень большие глаза . Под глазами скуловая кость выступала в сторону, что характерно и для цератопсов . Как и в челюстях большинства птицетазовых, передний край предчелюстной кости ( кость на кончике верхней челюсти) был беззубым и, вероятно, поддерживал ороговевший клюв ( рамфотека ), хотя у гетеродонтозаврид действительно были зубы в заднем отделе предчелюстной кости. Большой зазор, называемый диастемой , отделял эти предчелюстные зубы от зубов верхней челюсти (основной кости верхней челюсти) у многих птицетазовых животных, но у гетеродонтозаврид эта диастема была характерно изогнутой. ( Нижняя челюсть нижняя челюсть) имела на кончике предзубную кость, уникальную для птицетазовых. Эта кость также поддерживала клюв, похожий на тот, что был найден на предчелюстной кости . Все зубы нижней челюсти были обнаружены на зубной кости. [3]

Гетеродонтозавриды названы в честь их сильно гетеродонтного зубного ряда . Предчелюстных зубов было три. У раннеюрского Abrictosaurus , Heterodontosaurus и Lycorhinus первые два предчелюстных зуба были маленькими и коническими, тогда как гораздо более крупный третий зуб напоминал клыки хищных млекопитающих и часто назывался собачьим или «бивнем». Нижний клыковидный, более крупный, чем верхний, занимал первое положение в зубной кости и размещался в дугообразной диастеме верхней челюсти при закрытии рта. [3] Эти клыкообразные имели зазубрины как на переднем, так и на заднем краях у Heterodontosaurus и Lycorhinus , тогда как у Abrictosaurus зубцы были только на переднем крае. [8] [9] У раннемелового Echinodon , возможно, существовало два верхних клыкообразных, которые располагались на верхней челюсти, а не на предчелюстной кости. [10] и Fruitadens из поздней юры , возможно, имели по две нижних клыкообразных на каждой зубной кости. [11] [12]

Как и характерные бивни, щечные зубы производных гетеродонтозаврид также были уникальными среди ранних птицетазовых. Небольшие гребни, или зубцы, располагались по краям щечных зубов птицетазовых, чтобы срезать растительность. Эти зубцы простираются только на треть длины коронки зуба от кончика у всех гетеродонтозаврид; у других птицетазовых зубцы простираются дальше вниз к корню. Базальные формы, такие как Abrictosaurus, имели щечные зубы как на верхней, так и на зубной кости, которые в целом были похожи на другие птицетазовые: широко расставленные, каждая из которых имела низкую коронку и сильно развитый гребень (цингулюм), отделяющий коронку от корня. У более производных форм, таких как Lycorhinus и Heterodontosaurus , зубы имели долотообразную форму, с гораздо более высокими коронками и без поясной извилины, так что не было разницы в ширине между коронками и корнями. [3]
Эти производные щечные зубы перекрывали друг друга, так что их коронки образовывали сплошную поверхность, на которой можно было пережевывать пищу. Зубные ряды были слегка вставлены со стороны рта, оставляя за пределами зубов пространство, которое, возможно, было ограничено мускулистой щекой , необходимой для жевания. Гадрозавры и цератопсы мелового периода, а также многие травоядные млекопитающие конвергентно развили несколько аналогичных зубных батарей . В отличие от гадрозавров, у которых постоянно заменялись сотни зубов, замена зубов у гетеродонтозаврид происходила гораздо медленнее, и было обнаружено несколько экземпляров без единого сменного зуба. Характерно, что у гетеродонтозаврид отсутствовали небольшие отверстия (отверстия) на внутренней стороне костей челюсти, которые, как полагают, способствовали развитию зубов у большинства других птицетазовых животных. Гетеродонтозавриды также могли похвастаться уникальным сфероидальным суставом между зубными и предзубными челюстями, позволяющим нижним челюстям вращаться наружу при закрытии рта, скрежетая щечные зубы друг о друга. Из-за медленной скорости замены такое шлифование приводило к сильному износу зубов, который обычно стирал большую часть зубчиков в старых зубах, хотя увеличенная высота коронок обеспечивала каждому зубу долгую жизнь. [13]
Скелет
[ редактировать ]
Посткраниальная анатомия Heterodontosaurus tucki хорошо описана, хотя H. tucki обычно считается наиболее производным из раннеюрских гетеродонтозаврид, поэтому невозможно узнать, сколько его особенностей было общим с другими видами. [3] Передние конечности были для динозавра длинными, более 70% длины задних конечностей. Хорошо развитый дельтопекторальный гребень (гребень для прикрепления грудных и плечевых мышц) плечевой кости и выступающий локтевой отросток (где мышцы, разгибающие предплечье прикреплялись ) локтевой кости указывают на то, что передняя конечность также была мощной. («руке») было пять цифр На манусе . Первый был большим, с остро изогнутым когтем на конце и при сгибании поворачивался внутрь; Роберт Баккер назвал это «поворотом большого пальца». [14] Вторая цифра была самой длинной, чуть длиннее третьей. На обоих этих пальцах были когти, тогда как четвертый и пятый пальцы без когтей были по сравнению с ними очень маленькими и простыми. В задних конечностях большеберцовая кость была на 30% длиннее бедренной , что обычно считается адаптацией к скорости. Большеберцовая и малоберцовая кость голени были слиты с таранной и пяточной костями лодыжки, образуя « тибиофибиотарзус », конвергентно с современными птицами . Также, как и у птиц, нижние кости предплюсны (лодыжки) и плюсны были слиты, образуя « предплюсне-плюсневую кость ». (задней стопе) четыре пальца На стопе , причем только второй, третий и четвертый пальцы касаются земли. Хвост, в отличие от многих других птицетазовых, не имел окостеневших сухожилий, позволяющих сохранять жесткую позу, и, вероятно, был гибким. [5] Фрагментарный скелет, известный у Abrictosaurus, никогда не был полностью описан, хотя передняя конечность и кисть были меньше, чем у Heterodontosaurus . Кроме того, на четвертом и пятом пальцах передней конечности на одну фаланговую кость меньше . [15]
Классификация
[ редактировать ]
Южноафриканский палеонтолог Роберт Брум в 1911 году придумал название «Геранозавр» для костей челюсти динозавра, у которых отсутствуют все зубы и некоторые частично связанные с ними кости конечностей. [16] В 1924 году Lycorhinus назвал и классифицировал его как цинодонта Сидни Хотон . [17] Гетеродонтозавр был назван в 1962 году, и он, Lycorhinus и Geranosaurus были признаны близкородственными птицетазовыми динозаврами. [18] Альфред Ромер назвал Heterodontosauridae в 1966 году семейством птицетазовых динозавров, включая Heterodontosaurus и Lycorhinus . [19] Кун независимо предложил Heterodontosauridae в том же году и иногда упоминается как его основной автор. [20] Он был определен как клада в 1998 году Полом Серено. [21] и переопределен им в 2005 году как стволовая клада, состоящая из Heterodontosaurus tucki и всех видов, более близких к Heterodontosaurus, чем к Parasaurolophus walkeri , Pachycephalosaurus wyomingensis , Triceratops horridus или Ankylosaurus magniventris . [22] Heterodontosaurinae — это стеблевой таксон, впервые филогенетически определенный Полом Серено в 2012 году как «наиболее инклюзивная клада, содержащая Heterodontosaurus tucki , но не Tianyulong confuciusi , Fruitadens haagarorum , Echinodon becklesii ». [1]
Heterodontosauridae включает роды Abrictosaurus , Lycorhinus и Heterodontosaurus , все из Южной Африки. Хотя Ричард Талборн однажды передал всех троих Ликоринусу , [15] все остальные авторы считают эти три рода отдельными. [9] Внутри семейства Heterodontosaurus и Lycorhinus считаются сестринскими таксонами , а Abrictosaurus является базальным членом. [4] Геранозавр также является гетеродонтозавридом, но обычно считается nomen dubium, поскольку у типового экземпляра отсутствуют все зубы, что делает его неотличимым от любого другого рода семейства. [3] Совсем недавно в нескольких исследованиях род Echinodon считался гетеродонтозавридом. [10] [11] Ланазавр был назван в честь верхней челюсти в 1975 году. [23] но более поздние открытия показали, что вместо этого он принадлежит Lycorhinus , что делает Lanasaurus младшим синонимом этого рода. [6] Дианчунгозавр когда-то считался гетеродонтозавридом из Азии . [24] тех пор было показано, что останки представляли собой химеру прозауроподов но с и останков мезоэвкрокодилов . [25] Хосе Бонапарт одно время также классифицировал южноамериканского пизанозавра как гетеродонтозаврид. [26] но теперь известно, что это животное является более базальным птицетазовым животным. [27]

Принадлежность к Heterodontosauridae хорошо известна по сравнению с его неопределенным филогенетическим положением. Несколько ранних исследований показали, что гетеродонтозавриды были очень примитивными птицетазами. [5] [18] Из-за предполагаемого сходства в морфологии передних конечностей Роберт Баккер предположил родство между гетеродонтозавридами и ранними зауроподоморфами , такими как анхизавр , соединяющими отряды Saurischia и Ornithischia . [14] Доминирующая гипотеза последних нескольких десятилетий относит гетеродонтозаврид к базальным орнитоподам. [3] [4] [8] [28] Однако другие предположили, что гетеродонтозавриды имеют общего предка с маргиноцефалиями ( цератопсы и пахицефалозавры ). [29] [30] гипотеза, которая нашла поддержку в некоторых исследованиях начала 21 века. [31] [32] Клада, содержащая гетеродонтозаврид и маргиноцефалов, была названа Heterodontosauriformes . [33] Гетеродонтозавриды также считались базальными как для орнитопод, так и для маргиноцефалов. [34] [35] В 2007 году кладистический анализ показал, что гетеродонтозавриды являются базальными для всех известных птицетазовых, за исключением Pisanosaurus , и этот результат перекликается с некоторыми из самых ранних работ по этому семейству. [36] [37] Однако исследование Бонапарта показало, что Pisanosauridae являются синонимами Heterodontosauridae, а не отдельным семейством само по себе, тем самым включая Pisanosaurus как гетеродонтозавра. [38] Батлер и др. (2010) обнаружили, что Heterodontosauridae являются наиболее базовой из известных значимых птицетазовых радиаций. [39]
Кладограмма ниже показывает взаимосвязи внутри Heterodontosauridae и соответствует анализу Серено, 2012: [40]
| Гетеродонтозавриды | |
Переработка Cerapoda, проведенная Дьедонне и его коллегами в 2020 году, обнаружила животных, традиционно считавшихся «гетеродонтозавридами», как базальную группу внутри Pachycephalosauria, парафилетическую по отношению к традиционным куполообразным пахицефалозаврам. Этот результат был основан на многочисленных характеристиках черепа, включая зубной ряд , а также на том факте, что окаменелости пахицефалозавров совершенно неизвестны из юрского периода. Современное понимание филогении птицетазовых предполагает, что юрские пахицефалозавры должны существовать, поскольку множество юрских цератопсов было обнаружено , но ни один такой пахицефалозавр не был уверенно идентифицирован. Этот анализ был проведен для уточнения выводов Барона и его коллег (2017), согласно которым чилезавр был базальным птицетазовым животным . [41] Филогенетический анализ был проведен с участием чилезавра, закодированного как птицетазовый, что также имело значение для филогении орнитопод .
Кладограмма ниже представляет собой сокращенную версию выводов Дьедонна и его коллег: [42]
| Орнитишия |
| |||||||||||||||||||||
Распределение
[ редактировать ]
Хотя первоначально останки гетеродонтозаврид были известны только из ранней юры на юге Африки, сейчас они известны на четырех континентах . В начале истории гетеродонтозаврид суперконтинент Пангея практически по всему миру все еще оставался в значительной степени нетронутым, что позволило этому семейству распространиться . [10] Самые старые из известных остатков гетеродонтозаврид — это фрагмент челюсти и отдельные зубы из формации Лагуна-Колорада в Аргентине , которая датируется позднем триасом . Эти останки имеют производную морфологию, аналогичную Heterodontosaurus , включая клыкообразную форму с зубцами как на переднем, так и на заднем краях, а также зубы верхней челюсти с высокой коронкой без поясной части. [43] Ирмис и др. (2007) предварительно согласились, что этот ископаемый материал представляет собой гетеродонтозаврид, но заявили, что для подтверждения этого отнесения необходим дополнительный материал, поскольку образец плохо сохранился. [44] в то время как Серено (2012) лишь заявил, что этот материал может представлять собой птицетазового животного или даже, в частности, гетеродонтозаврид. [1] Олсен , Кент и Уайтсайд (2010) отметили, что возраст самой формации Лагуна-Колорада плохо определен, и поэтому не было окончательно определено, относится ли предполагаемый гетеродонтозаврид из этой формации к триасовому или юрскому возрасту. [45] Самая разнообразная фауна гетеродонтозаврид происходит из ранней юры южной Африки, где окаменелости Heterodontosaurus , Abrictosaurus , Lycorhinus и сомнительного Geranosaurus . обнаружены [3]
Неописанные раннеюрские гетеродонтозавриды также известны из США. [46] и Мексика , [47] соответственно. Кроме того, начиная с 1970-х годов, большое количество ископаемого материала было обнаружено в позднеюрской формации Моррисон недалеко от Фруты , штат Колорадо , США. [11] Описанный в печати в 2009 году, этот материал был отнесен к роду Fruitadens . [12] Зубы гетеродонтозаврид без поясной части также были описаны в позднеюрских и раннемеловых образованиях Испании и Португалии . [48] Останки Echinodon были переописаны в 2002 году, что показало, что они могут представлять собой поздно выживших гетеродонтозаврид из берриасского этапа раннего мела на юге Англии. [10] Дианчунгозавр из ранней юры Китая больше не считается гетеродонтозавридом; хотя известна одна средне-позднеюрская азиатская форма ( Тяньюлун ). [7] Неопределенные щечные зубы, возможно, представляющие собой гетеродонтозаврид, также известны из барремского возраста формации Уэссекс на юге Англии, и в случае подтверждения это будет самая молодая находка в группе. [49]
Палеобиология
[ редактировать ]
Большинство окаменелостей гетеродонтозаврид обнаружено в геологических формациях , которые представляют собой засушливые и полузасушливые среды, включая формацию Аппер-Эллиот в Южной Африке и пласты Пурбек на юге Англии. [4] Было высказано предположение, что гетеродонтозавриды впадали в сезонную спячку или спячку в самые засушливые времена года. Из-за отсутствия сменных зубов у большинства гетеродонтозаврид было предложено заменять весь набор зубов в этот период покоя, поскольку казалось, что постоянная и спорадическая замена зубов нарушит функцию зубного ряда как единой жевательной поверхности. . [15] Однако это было основано на неправильном понимании механики челюстей гетеродонтозаврид. [50] Считалось, что гетеродонтозавриды на самом деле постоянно меняли свои зубы, хотя и медленнее, чем у других рептилий , но компьютерная томография черепов молодых и взрослых гетеродонтозавров не выявила замены зубов. [51] В настоящее время нет доказательств, подтверждающих гипотезу эстивации у гетеродонтозаврид. [3] но его нельзя отвергнуть, судя по сканированию черепа. [51]
Хотя щечные зубы гетеродонтозаврид явно приспособлены для измельчения жесткого растительного материала, их диета могла быть всеядной. Заостренные предчелюстные зубы и острые изогнутые когти на передних конечностях предполагают некоторую степень хищнического поведения. Было высказано предположение, что длинные и мощные передние конечности Heterodontosaurus могли быть полезны для разрывания гнезд насекомых, как и у современных муравьедов . Эти передние конечности, возможно, также служили орудиями для рытья корней и клубней . [3]
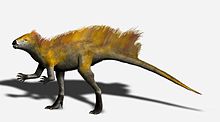
Длина передней конечности по сравнению с задней конечностью предполагает, что гетеродонтозавр мог быть частично четвероногим , а выступающий локтевой отросток и сверхразгибаемые пальцы передней конечности встречаются у многих четвероногих. Однако кисть явно предназначена для захвата, а не для поддержки веса. Многие особенности задних конечностей, включая длинную голень и стопу, а также слияние тибиофибиотарсуса и предплюснепредплюсны, указывают на то, что гетеродонтозавриды были приспособлены к быстрому бегу на задних конечностях, поэтому маловероятно, что гетеродонтозавр передвигался на всех четырех конечностях, за исключением, возможно, случаев, когда кормление. [5]
Короткие бивни, обнаруженные у всех известных гетеродонтозаврид, сильно напоминают бивни современных кабарги , пекари и свиней . с более длинными бивнями У многих из этих животных (а также у моржей и азиатских слонов ) это половой диморфный признак: бивни встречаются только у самцов. Типовой экземпляр абриктозавра лишен бивней и первоначально был описан как самка. [15] Хотя это остается возможным, несросшиеся крестцовые позвонки и короткая морда указывают на то, что этот экземпляр представляет собой молодое животное. Второй, более крупный экземпляр, первоначально предположительно принадлежавший абриктозавру , явно обладает бивнями, что использовалось для подтверждения идеи о том, что бивни встречаются только у взрослых, а не являются вторичным половым признаком самцов. Эти бивни могли использоваться для боя или демонстрации представителей того же вида или других видов. [3] Отсутствие бивней у молодых абриктозавров также может быть еще одной характеристикой, отличающей его от других гетеродонтозаврид, поскольку бивни известны и у молодых гетеродонтозавров . Другие предполагаемые функции бивней включают защиту и использование в периодической всеядной диете. [51] Однако в 2012 году Серено в качестве альтернативы отнес этот экземпляр к Lycorhinus , который, как уже известно, обладал бивнями, и поэтому их отсутствие у Abrictosaurus могло быть не результатом возраста. [1]
был обнаружен небольшой полный окаменелый скелет гетеродонтозавра возрастом более 200 миллионов лет В 2005 году в Южной Африке . В июле 2016 года его отсканировала группа южноафриканских исследователей с использованием Европейской установки синхротронного излучения ; сканирование зубного ряда выявило кости неба толщиной менее миллиметра. [52]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Серено, Пол К. (2012). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридных динозавров» . ZooKeys (226): 1–225. дои : 10.3897/zookeys.226.2840 . ПМК 3491919 . ПМИД 23166462 .
- ^ Мадзия, Дэниел; Арбур, Виктория М.; Бойд, Клинт А.; Фарке, Эндрю А.; Крусадо-Кабальеро, Пенелопа; Эванс, Дэвид К. (9 декабря 2021 г.). «Филогенетическая номенклатура птицетазовых динозавров» . ПерДж . 9 : е12362. дои : 10.7717/peerj.12362 . ISSN 2167-8359 . ПМЦ 8667728 . ПМИД 34966571 .
- ^ Jump up to: а б с д и ж г час я дж к Вейшампель, Дэвид Б .; Уитмер, Лоуренс М. (1990). «Гетеродонтозавриды». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия . Беркли: Издательство Калифорнийского университета. стр. 486–497. ISBN 978-0-520-06727-1 .
- ^ Jump up to: а б с д Норман, Дэвид Б .; Сьюс, Ханс-Дитер; Уитмер, Лоуренс М.; Кориа, Родольфо А. (2004). «Базальные орнитоподы». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 393–412. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б с д Санта-Лука, Альберт П. (1980). «Посткраниальный скелет Heterodontosaurus tucki (Reptilia, Ornithischia) из Штормберга в Южной Африке». Анналы Южноафриканского музея . 79 (7): 159–211.
- ^ Jump up to: а б Гоу, Кристофер Э. (1990). «Верхняя челюсть с зубами, относящаяся к Lycorhinus angustidens Haughton, 1924 (Dinosauria, Ornithischia)». Анналы Южноафриканского музея . 99 (10): 367–380.
- ^ Jump up to: а б Чжэн, Сяо-Тин; Ты, Хай-Лу; Сюй, Син; Донг, Чжи-Мин (19 марта 2009 г.). «Раннемеловой гетеродонтозавридный динозавр с нитевидными покровными структурами». Природа . 458 (7236): 333–336. Бибкод : 2009Natur.458..333Z . дои : 10.1038/nature07856 . ПМИД 19295609 . S2CID 4423110 .
- ^ Jump up to: а б Талборн, Ричард А. (1970). «Систематическое положение триасового птицетазового динозавра Lycorhinus angustidens ». Зоологический журнал Линнеевского общества . 49 (3): 235–245. дои : 10.1111/j.1096-3642.1970.tb00739.x .
- ^ Jump up to: а б Хопсон, Джеймс А. (1975). «О родовом выделении птицетазовых динозавров Lycorhinus и Heterodontosaurus из серии Штормберг (верхний триас) Южной Африки». Южноафриканский научный журнал . 71 : 302–305.
- ^ Jump up to: а б с д Норман, Дэвид Б .; Барретт, Пол М. (2002). «Порнитишские динозавры нижнего мела (берриаса) Англии». В Милнере, Эндрю; Баттен, Дэвид Дж. (ред.). Жизнь и окружающая среда во времена Пурбека . Специальные статьи по палеонтологии 68 . Лондон: Палеонтологическая ассоциация. стр. 161–189. ISBN 978-0-901702-73-9 .
- ^ Jump up to: а б с Гальтон, Питер М. (2007). «Зубы птицетазовых динозавров (в основном Ornithopoda) из формации Моррисон (верхняя юра) на западе США». В Карпентер, Кеннет (ред.). Рога и клювы: цератопсы и динозавры-орнитоподы . Блумингтон: Издательство Университета Индианы. стр. 17–47. ISBN 978-0-253-34817-3 .
- ^ Jump up to: а б Батлер, Ричард Дж.; Гальтон, Питер М.; Порро, Лаура Б.; Кьяппе, Луис М.; Хендерсон, DM; Эриксон, Грегори М. (2010). «Нижние пределы размеров тела птицетазовых динозавров, выведенные на основе нового верхнеюрского гетеродонтозаврид из Северной Америки» . Труды Королевского общества Б. 277 (1680): 375–381. дои : 10.1098/rspb.2009.1494 . ПМЦ 2842649 . ПМИД 19846460 .
- ^ Вейшампель, Дэвид Б. (1984). "Введение". Эволюция челюстных механизмов динозавров-орнитопод . Достижения в анатомии, эмбриологии и клеточной биологии. Том. 87. Берлин; Нью-Йорк: Springer-Verlag. стр. 1–109. дои : 10.1007/978-3-642-69533-9_1 . ISBN 978-0-387-13114-6 . ПМИД 6464809 .
- ^ Jump up to: а б Баккер, Роберт Т. (1986). Ереси динозавров: новые теории, раскрывающие тайну динозавров и их исчезновения . Нью-Йорк: Уильям Морроу. п. 453 стр. ISBN 978-0-14-010055-6 .
- ^ Jump up to: а б с д Талборн, Ричард А. (1974). «Новый гетеродонтозавридный динозавр (Reptilia: Ornithischia) из красных пластов верхнего триаса Лесото». Зоологический журнал Линнеевского общества . 55 (2): 151–175. дои : 10.1111/j.1096-3642.1974.tb01591.x .
- ^ Брум, Роберт. (1911). «О динозаврах Стормберга, Южная Африка». Анналы Южноафриканского музея . 7 : 291–308.
- ^ Хотон, Сидни Х. (1924). «Фауна и стратиграфия серии Штормберг». Анналы Южноафриканского музея . 12 : 323–497.
- ^ Jump up to: а б Кромптон, штат Аризона; Чариг, Алан (1962). «Новый птицетаз из верхнего триаса Южной Африки». Природа . 196 (4859): 1074–1077. Бибкод : 1962Natur.196.1074C . дои : 10.1038/1961074a0 . S2CID 4198113 .
- ^ Ромер, Альфред С. (1966). Палеонтология позвоночных (Третье изд.). Чикаго: Издательство Чикагского университета. п. 468 стр . ISBN 978-0-7167-1822-2 .
- ^ Серено, Пол (10 марта 2012 г.). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридных динозавров» . ZooKeys (226): 1–225. дои : 10.3897/zookeys.226.2840 . ISSN 1313-2970 . ПМК 3491919 . ПМИД 23166462 .
- ^ Серено, Пол К. (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Новогодний альбом по геологии и палеонтологии, Трактаты . 210 (1): 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Серено, Пол К. (07 ноября 2005 г.). «Стеблевая архозаврия — TaxonSearch» . Архивировано из оригинала 19 февраля 2007 г. Проверено 24 февраля 2007 г.
- ^ Гоу, Кристофер Э. (1975). «Новый гетеродонтозаврид из Redbeds Южной Африки, демонстрирующий явные доказательства замены зубов». Зоологический журнал Линнеевского общества . 57 (4): 335–339. дои : 10.1111/j.1096-3642.1975.tb01895.x .
- ^ Янг, CC (1982). «[Новый род динозавров из округа Луфэн, провинция Юньнань]». Ин Чжоу М. (ред.). [Собрание сочинений Ян Чжунцзяня] (на китайском языке). Пекин: Академика Синика. стр. 38–42.
- ^ Барретт, Пол М.; Сюй Син (2005). «Повторная оценка Dianchungosaurus lufengensis Yang, 1982a, загадочной рептилии из формации Нижний Луфэн (нижняя юра) провинции Юньнань, Китайская Народная Республика». Журнал палеонтологии . 79 (5): 981–986. doi : 10.1666/0022-3360(2005)079[0981:ARODLY]2.0.CO;2 . ISSN 0022-3360 . S2CID 89542580 .
- ^ Бонапарт, Хосе Ф. (1976). « Pisanosaurus mertii Casamiquela и происхождение птицетазовых». Журнал палеонтологии . 50 : 808–820.
- ^ Вейшампель, Дэвид Б .; Уитмер, Лоуренс М. (1990). « Лесотозавр , Пизанозавр и Технозавр ». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (Первое изд.). Беркли: Издательство Калифорнийского университета. стр. 416–425. ISBN 978-0-520-06727-1 .
- ^ Серено, Пол К. (1986). «Филогения птицетазовых динозавров». Национальное географическое исследование . 2 (2): 234–256.
- ^ Чжао Сицзинь (1983). «Филогения и этапы эволюции динозавров». Acta Palaeontologica Polonica . 28 (1–2): 295–306.
- ^ Купер, Майкл А. (1985). «Пересмотр птицетазового динозавра Kangnasaurus coetzeei Haughton с классификацией птицетазовых». Анналы Южноафриканского музея . 95 : 281–317.
- ^ Чжао Сицзинь ; Чэн Чжэнву; Сюй Син (1999). «Самый ранний цератопс из формации Тучэнцзы в Ляонине, Китай». Журнал палеонтологии позвоночных . 19 (4): 681–691. Бибкод : 1999JVPal..19..681X . дои : 10.1080/02724634.1999.10011181 .
- ^ Ю Хайлу; Сюй Син ; Ван Сяолинь (2003). «Новый род Psittacosauridae (Dinosauria: Ornithopoda) и происхождение и ранняя эволюция маргиноцефальных динозавров». Acta Geologica Sinica (английское издание) . 77 (1): 15–20. Бибкод : 2003AcGlS..77...15Y . дои : 10.1111/j.1755-6724.2003.tb00105.x . S2CID 89051352 .
- ^ Сюй Син, X ; Форстер, Кэтрин А.; Кларк, Джеймс М.; Мо Цзинью. (2006). «Базальный цератопс с переходными чертами из поздней юры северо-западного Китая» . Труды Королевского общества B: Биологические науки . 273 (1598): 2135–2140. дои : 10.1098/rspb.2006.3566 . ПМЦ 1635516 . ПМИД 16901832 .
- ^ Марианска, Тереза ; Осмольска, Гальшка. (1985). «О филогении птицетазовых». Acta Palaeontologica Polonica . 30 : 137–150.
- ^ Батлер, Ричард Дж. (2005). «Фаброзавриды» птицетазовые динозавры формации Верхний Эллиот (нижняя юра) Южной Африки и Лесото» . Зоологический журнал Линнеевского общества . 145 (2): 175–218. дои : 10.1111/j.1096-3642.2005.00182.x .
- ^ Батлер, Ричард Дж .; Смит, Роджер М.Х.; Норман, Дэвид Б. (2007). «Примитивный птицетазовый динозавр из позднего триаса Южной Африки, ранняя эволюция и разнообразие птицетазовых динозавров» . Труды Королевского общества B: Биологические науки . 274 (опубликовано в Интернете): 2041–6. дои : 10.1098/rspb.2007.0367 . ПМК 2275175 . ПМИД 17567562 .
- ^ Батлер, Ричард Дж.; Апчерч, Пол; Норман, Дэвид Б. (2008). «Филогения птицетазовых динозавров». Журнал систематической палеонтологии . 6 (1): 1–40. Бибкод : 2008JSPal...6....1B . дои : 10.1017/S1477201907002271 . S2CID 86728076 .
- ^ Бонапарт, JF (1976). «Pisanosaurus mertii Casamiquela и происхождение попугаев». Журнал палеонтологии. 50 (5): 808–820. JSTOR 1303575 .
- ^ Р. Дж. Батлер. 2010. Анатомия базального птицетазового динозавра Eocursor parvus из нижней части формации Эллиот (поздний триас) Южной Африки. Зоологический журнал Линнеевского общества 160:648-684.
- ^ Серено, ПК (2012). стр. 100-1 193-2
- ^ Барон, Мэтью Г.; Барретт, Пол М. (01 марта 2018 г.). «Поддержка размещения чилезавра в пределах Ornithischia: ответ Мюллеру и др.» . Письма по биологии . 14 (3): 20180002. doi : 10.1098/rsbl.2018.0002 . ISSN 1744-9561 . ПМЦ 5897612 . ПМИД 29593075 .
- ^ Дьедонне, ЧП; Крузадо-Кабальеро, П.; Годфруа, П.; Тортоса, Т. (2020). «Новая филогения цераподских динозавров» . Историческая биология . 33 (10): 2335–2355. дои : 10.1080/08912963.2020.1793979 .
- ^ Баэс, Ана Мария; Марсикано, Клаудия А. (2001). «Гетеродонтозаврид-птицетазовый динозавр из верхнего триаса Патагонии». Амегиниана . 38 (3): 271–279.
- ^ Рэндалл Б. Ирмис; Уильям Дж. Паркер; Стерлинг Дж. Несбитт; Цзюнь Лю (2007). «Ранние птицетазовые динозавры: триасовая летопись». Историческая биология . 19 (1): 3–22. Бибкод : 2007HBio...19....3I . CiteSeerX 10.1.1.539.8311 . дои : 10.1080/08912960600719988 . S2CID 11006994 .
- ^ Пол Э. Олсен; Деннис В. Кент; Джессика Х. Уайтсайд (2010). «Последствия астрохронологии и временной шкалы геомагнитной полярности, основанной на супергруппе Ньюарка (Newark-APTS), для темпа и режима ранней диверсификации динозавров». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 101 (3–4): 201–229. Бибкод : 2010EETR.101..201O . дои : 10.1017/S1755691011020032 . S2CID 16123732 .
- ^ Сьюс, Ханс-Дитер; Кларк, Джеймс М.; Дженкинс, Фариш А. (1994). «Обзор раннеюрских четвероногих из группы Глен-Каньон на юго-западе Америки». У Фрейзера, Николаса К.; Сьюс, Ханс-Дитер (ред.). В тени динозавров: четвероногие раннего мезозоя . Кембридж: Издательство Кембриджского университета. стр. 285–294. ISBN 978-0-521-45899-3 .
- ^ Кларк, Джеймс; Монтеллано, Марисоль; Хопсон, Джеймс А.; Эрнандес, Рене; Фастовский, Дэвид А. (1994). «Набор четвероногих ранней или средней юры из формации Ла Бока, северо-восток Мексики». Во Фрейзере, Северная Каролина; Сьюс, Х.-Д. (ред.). В тени динозавров: четвероногие раннего мезозоя . Кембридж: Издательство Кембриджского университета. стр. 295–302. ISBN 978-0-521-45899-3 .
- ^ Санчес-Эрнандес, Барбара; Бентон, Майкл Дж .; Нэйш, Даррен (2007). «Динозавры и другие ископаемые позвоночные животные из поздней юры и раннего мела в районе Гальве, северо-восток Испании» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 249 (1–2): 180–215. Бибкод : 2007PPP...249..180S . дои : 10.1016/j.palaeo.2007.01.009 .
- ^ Свитман, Стивен С. (март 2016 г.). «Сравнение барремско-раннеаптских сообществ позвоночных из группы Джехол, северо-восточный Китай, и группы Уэлден, южная Великобритания: ценность исследований микропозвоночных в неблагоприятных условиях сохранения» . Палеобиоразнообразие и палеосреда . 96 (1): 149–167. Бибкод : 2016ПдПе...96..149С . дои : 10.1007/s12549-015-0217-9 . ISSN 1867-1594 . S2CID 129956539 .
- ^ Хопсон, Джеймс А. (1980). «Функция и замена зубов у птицетазовых динозавров раннего мезозоя: значение для летнего периода». Летайя . 13 (1): 93–105. Бибкод : 1980Лета..13...93Ч . дои : 10.1111/j.1502-3931.1980.tb01035.x .
- ^ Jump up to: а б с Батлер, Ричард Дж.; Порро, Лаура Б.; Норман, Дэвид Б. (2008). «Молодой череп примитивного птицетазового динозавра Heterodontosaurus tucki из Штормберга на юге Африки». Журнал палеонтологии позвоночных . 28 (3): 702–711. doi : 10.1671/0272-4634(2008)28[702:AJSOTP]2.0.CO;2 . ISSN 0272-4634 . S2CID 86739244 .
- ^ «Окаменелость динозавра, найденная в ЮАР, наконец-то раскрывает свои тайны» . 27 июля 2016 г. Архивировано из оригинала 28 июля 2016 г. Проверено 27 июля 2016 г.