Ссыпанный
| Ссыпанный Временный диапазон: поздний меховой ,
~ | |
|---|---|

| |
| Два реконструированных скелета S. vaduum на основе образца UALVP 2, Музей Королевского Тиррелла | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | † Ornithischia |
| Клада : | † neornithischia |
| Клада : | † Pachycephalosauria |
| Семья: | †Pachycephalosauridae |
| Род: | † Шаги Ламбе , 1902 |
| Тип видов | |
| † stegoceras верно Ламбе, 1902
| |
| Разновидность | |
| |
| Синонимы | |
|
Список | |
Stegoceras является родом динозавра Pachycephalosaurid (головы купола) , который жил в том, что сейчас является Северной Америкой в поздний меховой период , от 77,5 до 74 миллионов лет назад (MYA). The first specimens from Alberta , Canada, were described in 1902, and the type species Stegoceras validum was based on these remains. The generic name means "horn roof", and the specific name means "strong". Несколько других видов были помещены в род на протяжении многих лет, но с тех пор они были перемещены в другие роды или считаются младшими синонимами . В настоящее время только S. vadum и S. novomexicanum , названные в 2011 году от окаменелостей, найденных в Нью -Мексико , остаются. Достоверность последнего вида также обсуждалась.
Stegoceras was a small, bipedal dinosaur about 2 to 2.5 metres (6.6 to 8.2 ft) long, and weighed around 10 to 40 kilograms (22 to 88 lb). The skull was roughly triangular with a short snout, and had a thick, broad, and relatively smooth dome on the top. The back of the skull had a thick "shelf" over the occiput , and it had a thick ridge over the eyes. Большая часть черепа была украшена бугорками (или круглыми «исходом») и узлами (или «ручками»), многими в рядах, и самые большие сформированные маленькие рога на полке. The teeth were small and serrated. The skull is thought to have been flat in juvenile animals and to have grown into a dome with age. It had a rigid vertebral column , and a stiffened tail. The pelvic region was broad, perhaps due to an extended gut.
Первоначально известный только по куполам черепа, Stegoceras был одним из первых известных пахицефалозавров, и неполнота этих первоначальных останков привела ко многим теориям о сродстве этой группы. A complete Stegoceras skull with associated parts of the skeleton was described in 1924, which shed more light on these animals. Pachycephalosaurs are today grouped with the horned ceratopsians in the group Marginocephalia . Stegoceras itself has been considered basal (or "primitive") compared to other pachycephalosaurs. Стегоцера, скорее всего, был травоядным, и, вероятно, было хорошее обоняние. Функция купола была обсуждена, а конкурирующие теории включают использование во внутриспецифических боевых действиях (голова или боковая нока), сексуальное отображение или распознавание видов . S. vaduum известен из формирования парка динозавров и формирования Oldman , тогда как S. novomexicanum взят из формирования Fruitland и Kirtland .
История открытия
[ редактировать ]
Первые известные останки стегокеров были собраны канадским палеонтологом Лоуренсом Ламбе из группы реки живота , в районе Красной Дир -Ривер в Альберте, Канада . Эти останки состояли из двух частичных куполов черепа (образцы CMN 515 и CMN 1423 в Канадском музее природы ) из двух животных разных размеров, собранных в 1898 году, и третьего частичного купола (CMN 1594), собранного в 1901 году. На основании этих образцов, Lambe described and named the new monotypic genus and species Stegoceras validus in 1902 . [ 1 ] [ 2 ] The generic name Stegoceras comes from the Greek stegè/στέγη, meaning "roof" and keras/κέρας meaning "horn". The specific name validus means "strong" in Latin, possibly in reference to the thick skull-roof.[ 3 ] Поскольку этот вид был основан на нескольких образцах ( серия Syntype ), CMN 515 был обозначен как образец лектотипа Джоном Беллом Хэтчер в 1907 году. [ 4 ] [ 2 ]
Поскольку в этом районе не было обнаружено никаких подобных останков, Ламбе не был уверен в том, каким они были динозавры, и представляли ли они один или несколько видов; Он предположил, что купола были «преназалами», расположенными перед носовыми костями на средней линии головы, и отметил их сходство с носовым рогом образца трицератопса . [ 1 ] В 1903 году венгерский палеонтолог Франц Нопцса фон Фелс-Шильвас предположил, что фрагментарные купола стегокеров были на самом деле лобными и носовыми костями, и что у животного было бы один непарный рог. Ламбе был сочувствую этой идее нового типа «динозавра единорога» в обзоре бумаги NOPSCSA 1903 года. В это время все еще была неопределенность в отношении того, стегокеры к какой группе динозавров принадлежала динозавров, с как цератопсианцами (рогатыми динозаврами), так и с стегозаврами (высеченными динозаврами) в качестве претендентов. [ 5 ] [ 6 ] Хэтчер сомневался, принадлежали ли образцы Stegoceras к одному и тому же виду, и были ли они вообще динозавры, и предположили, что купола состояли из лобных, затылочных и теменных костей черепа. [ 2 ] В 1918 году Ламбе направил еще один купол (CMN 138) S. vadusus и назвал новый вид, S. brevis , на основе образца CMN 1423 (который он первоначально включил в S. vadyus ). К этому времени он рассматривал этих животных как членов Стегозаврии (затем состоит из оба семейства бронированных динозавров, Stegosauridae и Ankylosauridae ), в новой семье, которое он назвал Psalisauridae (названным в честь сводчатой или купольной крыши черепа). [ 7 ]


В 1924 году американский палеонтолог Чарльз У. Гилмор описал полный череп S. vadus с связанными посткраниальными останками, к тому времени наиболее полными останками динозавра с куполом. Он был обнаружен в группе реки живота американским палеонтологом Джорджем Ф. Стернбергом в 1926 году и каталогизировал как образец UALVP 2 в лаборатории Палеонтологии позвоночных. Эта находка подтвердила интерпретацию Хэтчетера куполов как, состоящую из лобнопариетальной области черепа. UALVP 2 был обнаружен с небольшими, рассеянными косниционными элементами, которые, как считается, были гастралия (ребра брюшной полости), которые не известны у других орнитишских динозавров (одна из двух основных групп динозавров). Гилмор указал, что зубы S. vadus были очень похожи на зубы вида Troodon Formosus (названные в 1856 году и тогдашним только известным из изолированных зубов) и описал купол черепа, обнаруженный близко к местности, где был обнаружен Трудон . Поэтому Гилмор рассмотрел Stegoceras является недействительным младшим синонимом Troodon , и предположил , , тем самым переименуя S. vadus в T. vaduus что даже два вида могут быть одинаковыми. Кроме того, он обнаружил, что С. Бревиса идентичен С. Валирусу и, следовательно, младший синоним последнего. Он также поместил эти виды в новую семью Troodontidae (поскольку Lambe не выбрал род типа для своего Psalisauridae), который он считал наиболее близким к динозаврам Ornithopod . [ 8 ] [ 9 ] Поскольку череп казался таким специализированным по сравнению с довольно «примитивным» скелетом, NOPCSA сомневался, что эти части действительно принадлежали вместе, и предположил, что череп принадлежал узлокавру , скелет к орнитоподу и предполагаемой гастралии (ребра живота) рыба. Это утверждение было опровергнуто Гилмором и Лорисом С. Расселом в 1930 -х годах. [ 10 ]
Классификация Гилмора была поддержана американскими палеонтологами Барнумом Брауном и Эрихом Марен Шлаикджером в их обзоре динозавров в 1943 году, когда он был известен из 46 черепов. Из этих образцов Браун и Шлаикджер назвали новую виду Т. Стернберги и Т. Эдмонтоненсис (оба из Альберты), а также перемещение крупных видов Т. Вайомингенсис (который был назван в 1931 году) в новый род Пахицефалозавр , а также два Другие виды. Они обнаружили, что T. vaduus отличается от T. formosus , но считал S. brevis женской формой T. vaduus и, следовательно, младшим синонимом. К этому времени динозавры с куполом считались либо родственниками орнитоподов, либо анкилозаврами. [ 10 ] В 1945 году, после изучения акций T. formosus и S. vadus Teeth, американский палеонтолог Чарльз М. Стернберг продемонстрировал различия между ними и вместо этого предположил, что Трудон был динозавром теропода , и что динозавры с куполом должны быть помещены в их собственная семья. Хотя Stegoceras был первым членом этого семейства, который был назван, Sternberg назвал группу Pachycephalosauridae после второго рода, поскольку он нашел это имя (что означает «толстая ящерица») более описательным. Он также рассмотрел Т. Стернберги и Т. Эдмонтоненсиса членов в Stegoceras , нашел С. Бревиса действительным, и назвал новый вид, С. Ламбей , основанный на образце, ранее упоминавшемся S. vadiusus . [ 3 ] [ 11 ] раскол от Troodon Рассел в 1948 году поддержал , который описал тероподную зубную зубную зуба с зубами, почти идентичными для зубов T. formosus . [ 12 ]



В 1953 году Биргер Болин назвал Трудон Бекселли на основе теменной кости из Китая. [ 13 ] В 1964 году Оскар Кун считал это однозначным видом стегокеров ; С. Бекселли . [ 14 ] В 1974 году польские палеонологи Тереза Марьянска и Халска Осмольска пришли к выводу, что «гастралия» стегокеров были окостененные сухожилия после идентификации таких структур в хвосте пахицефалозавра Хомалоцефале . [ 9 ] В 1979 году Уильям Патрик Уолл и Питер Галтон назвали новые виды Stegoceras Browni , основанные на сплюснутом куполе, ранее называемом женским S. validus в 1971 году. Специальное название чтит Барнум Браун, который нашел образец голотипа (образец Amnh. 5450 в Американском музее естественной истории ) в Альберте. [ 15 ] В 1983 году Гальтон и Ханс-Дитер переехали С. Брауни к своему собственному роду, Ornatotholus ( Ornatus является латинским для «украшенного» и Tholus для «Dome») и считает его первым известным американским членом группы «плоской» возглавляемые «Пахицефалозавры, ранее известные в Азии. [ 16 ] Pachycephalosaurs 1987 года Sues и Galton подняли конкретное имя Validus Validum В обзоре , которое впоследствии использовалось в научной литературе. Эти авторы синонимизировали С. Бревис , С. Стернберги и С. Ламбей с S. vaduum , обнаружили, что S. bexelli отличался от Stegoceras в нескольких особенностях и считал его неопределенным пахицефалозавром. [ 3 ] [ 4 ] В 1998 году Гудвин и его коллеги считали Ornatotholus юношеской S. vaduum , следовательно, младшим синонимом. [ 17 ]
Застройки 21 -го века
[ редактировать ]
In 2000, Robert M. Sullivan referred S. edmontonensis and S. brevis to the genus Prenocephale (until then only known from the Mongolian species P. prenes ), and found it more likely that S. bexelli belonged to Prenocephale than to Stegoceras , but Считается, что это номен дубий (сомнительное название, без различия символов) из -за его неполноты, и отметил, что его образец голотипа, по -видимому, был потерян. [ 18 ] В 2003 году Томас Э. Уильямсон и Томас Карр считали Ornatotholus именем сомнения или, возможно, ювенильными стегокерами . [ 19 ] В пересмотре Stegoceras 2003 года Салливан согласился с тем, что Ornatotholus был младшим синонимом Stegoceras , перенес S. lambei к новому роду Colepiocephale и С. Стернберги в Ханссюсея . Он заявил, что род Stegoceras стал таксоном мусора для мусора для малых и средних североамериканских пахицефалозавров до этого момента. К этому времени десятки образцов были переданы S. vaduum , включая многие купола, слишком неполные, чтобы быть определенными как стегокеры с уверенностью. UALVP 2 по -прежнему является наиболее полным образцом стегокеров , на котором основано наиболее научное понимание рода. [ 4 ] С. Бревис был перенесен в новый род Foraminacephale в 2016 году Райаном К. Шоттом Шоттом и Дэвидом С. Эвансом, [ 20 ] и С. Бекселли в Синоцефалу в 2021 году Эвансом и коллегами. [ 21 ] В 2023 году Аарон Д. Дайер и его коллеги проанализировали швы и отдельные элементы в черепах пахицефалозавров Gravitholus и Hanssuesia и не обнаружили существенного различия между ними и Stegoceras valum . Они считали оба младшими синонимами, с Gravitholus, представляющим конечную стадию в росте стегокеров . [ 22 ]
В 2002 году Уильямсон и Карр описали купол (образец NMMNH P-33983 в музее естественной истории и науки в Нью-Мексико ) из бассейна Сан-Хуан , Нью-Мексико , который они считали юношеским пахицефалозавром неопределенных видов (хотя, возможно, Sphaerotholus goodwini ) Полем В 2006 году Салливан и Спенсер Г. Лукас считали это юношеским S. vaduum , который значительно расширит диапазон видов. [ 23 ] [ 24 ] В 2011 году Стивен Э. Джасински и Салливан считали образца взрослого и сделали его голотипом новых видов stegoceras novomexicanum с двумя другими образцами (SMP VP-2555 и SMP VP-2790) в качестве паратипов. [ 25 ] 2011 года Филогенетический анализ Ватабе и его коллег не помещал два вида Stegoceras близко друг к другу. [ 26 ]
В 2016 году Уильямсон и Стивен Л. Брусатте воздержали голотип S. novomexicanum и обнаружили, что паратипы не принадлежали к тому же таксону, что и голотип, и что все задействованные образцы были несовершеннолетними. Кроме того, они не смогли определить, представлял ли образец голотипа различные виды S. novomexicanum , или это был юношеский из S. vaduum или sphaerotholus goodwini , или другой ранее известный пахицефалозавр. [ 27 ] В 2016 году Джасински и Салливан защитили достоверность S. novomexicanum ; Они согласились с тем, что некоторые особенности, используемые для диагностики видов, указывают на стадию суб-адульта, но представляли дополнительные диагностические особенности в голотипе, которые различают виды. Они также указали на некоторые черты взрослых, которые могут указывать на гетерохронность (разница в сроках онтогенетических изменений между родственными таксонами) у видов. Они признали, что паратипы и другие назначенные образцы отличались от голотипа, имея более высокие куполовые черепа, вместо этого назвав их как CF. S. novomexicanum (трудно идентифицировать), но обнаружил, что, вероятно, они все принадлежат к одному таксону (при назначенных образцах взрослые) из -за ограниченного стратиграфического интервала и географического диапазона. [ 28 ] Дайер и коллеги обнаружили, что голотип S. novomexicanum может быть незрелым Sphaerotholus , и, следовательно, младшим синонимом в 2023 году. [ 22 ]
Описание
[ редактировать ]
Stegoceras является одним из наиболее известных североамериканских пахицефалозавров и одним из немногих известных посткраниальных останков; S. Validum образец UALVP 2 является наиболее полным индивидуумом Stegoceras, известным на сегодняшний день. По оценкам, его длина составляла от 2 до 2,5 метров (от 6,6 до 8,2 фута), сравнимого с размером козы . [ 29 ] [ 30 ] [ 31 ] По оценкам, вес составляет от 10 до 40 килограммов (от 22 до 88 фунтов). [ 32 ] Stegoceras были от размера до среднего по сравнению с другими пахицефалозаврами. [ 3 ] S. novomexicanum , по -видимому, был меньше, чем S. vaduum , но оспаривается, являются ли известными образцами (неполными черепами) взрослыми или несовершеннолетними. [ 25 ] [ 27 ]
Череп и зубной зуб
[ редактировать ]
Череп стегокеров был примерно треугольной по форме при просмотре со стороны, с относительно короткой мордой. Фронтальные были очень толстыми и образовали и теменные кости приподнятый купол. Швар между этими двумя элементами был уничтожен (только слабо видим в некоторых образцах), и они в совокупности называют «фронтопариеталем». Фронтопариетальный купол был широким и имел относительно гладкую поверхность, причем только боковые стороны были морщинистыми (морщинистыми). Он был сужен выше и между орбитой (глазные розетки). Фронтопариеталь сузился сзади, был втиснут между сквамозальными костями и заканчивался депрессией над затылкой в задней части черепа. Париетальные и сквомозальные кости сформировали толстый шельф над затылкой, названной паритосковой шельфом, чья степень варьировалась между образцами. Сквамозал был большой, а не частью купола, а задняя часть была опухшей. Он был украшен нерегулярно разнесенными бугорками (или округлыми исходами), и ряд узлов (ручек) простирался вдоль его верхних краев, заканчивающиеся заостренным бугорком (или маленьким рогом) с каждой стороны на задней части черепа. Внутренний ряд небольших бугорков проходил параллельно с большим. За исключением верхней поверхности купола, большая часть черепа была украшена узлами, многие из которых расположены в рядах. [ 3 ]
Большая орбита была сформирована как несовершенный эллипс (с самой длинной осью спереди к спине), и была направлена в сторону и слегка вперед. Инфратвральный фенестра (открытие) за глаза была узкой и наклоненной назад, а супратропоральный фенестра на верхней части задней части черепа был очень уменьшен по размеру из -за утолщения фронтопариеталя. Основнойран мозговой (этаж части ) был сокращен и дистанцирован от областей под орбитами и вокруг неба . Затылка наклонилась назад и вниз, а затылочный мыщел был отклонен в том же направлении. Лесковая кость образовала нижний передний край орбиты, а ее поверхность имела ряды узлоподобных украшений. Префронтальные пальпебральные и кости были слиты и образовали толстый гребень над орбитой. Относительно большая югальная кость сформировала нижний край орбиты, простирался далеко вперед и вниз к челюстному суставу. Это было украшено хребтами и узлами в излучающем расположении. [ 3 ]
Носовые отверстия были большими и сталкивались спереди. была Носовая кость толстой, сильно скульптурной и имела выпуклый профиль. Он сформировал босс (щит) на средней вершине черепа вместе с лобной костью. Нижний передний фронт предчелюстной кости (передняя кость верхней челюсти) был ворваком и утолщен. Небольшое отверстие (отверстие) присутствовало в шове между преднамерками, ведущими в полость носа и, возможно, связан с органом Якобсона ( орган обонятельного чувства). Верхняя челюсть была короткими и глубокими, и, вероятно, содержала синус . У верхней челюсти была серия отверстий, которые соответствовали каждому положению зуба, и они функционировали как отрывки для извержения замены зубов. Мябильщика сформулирована с черепом под задней частью орбиты. Зубчатая часть нижней челюсти была длинной, а часть была довольно короткой. Хотя это и не сохранилось, присутствие предпринятой кости обозначено границами в передней части нижней челюсти. [ 3 ] Как и другие пахицефалозавры, у него был бы небольшой клюв. [ 33 ]

У Stegoceras были зубы, которые были гетеродонтами (дифференцированными) и Thecodont (помещены в розетки). У него были маргинальные ряды относительно маленьких зубов, и ряды не образовали прямой режущей кромки. Зубы были наклонно установлены по длине челюстей и слегка перекрывали друг друга спереди назад. С каждой стороны наиболее полный образец (UALVP 2) имел три зуба в премейсилле, шестнадцать в верхней челюсти (обе часть верхней челюсти) и семнадцать в зубном состоянии нижней челюсти. The teeth in the premaxilla were separated from those behind in the maxilla by a short diastema (space), and the two rows in the premaxilla were separated by a toothless gap at the front. Зубы в передней части верхней челюсти (премейсилла) и передняя нижняя челюсть были одинаковыми; У них были более высокие, более заостренные и рекрустные короны, а также «каблук» сзади. Передние зубы в нижней челюсти были больше, чем у верхней челюсти. Передние края коронок имели восемь зубчатых зубцов (зубцов), а задний край - девять до одиннадцати. Зубы в задней части верхней (верхняя челюсть) и нижняя челюсть были треугольными по визу и сжаты спереди. У них были длинные корни, которые были овальными в секции, а короны были отмечены пояс на их базах. Данкуля здесь были сжаты и направлены на вершину коронок. Как внешняя, так и внутренняя сторона зубной коронок имела эмаль , и обе стороны были разделены вертикально на гребне. У каждого края было около семи или восьми зубных зубьев, а передний край обычно больше всего. [ 3 ]
Череп стегокеров можно отличить от числа других пахицефалозавров по таким особенностям, как его выраженная париосоквамозальная шельфа (хотя это стало меньше с возрастом), «зарождающийся» доминг его фронтопариента (хотя доминг увеличился с возрастом), его надутые носовые кости. , его орнамент бугорков по бокам и задней части сквомозальных костей, ряды до шести бугорков на верхней стороне каждого сквомозали и до двух узлов на обратной проекции теменной. Он также отличается от отсутствия носового орнамента и имеет уменьшенную диастему. [ 4 ] [ 34 ] Череп S. novomexicanum можно отличить от такого у S. vadum в таких особенностях, как обратное расширение теменной кости, более уменьшенной и треугольной, имея более высокий супратропоральный фенестра (хотя это может быть связано с возможным ювенальным статусом образца ), и имея примерно параллельные контакты шва между сквомозальными и теменными. Похоже, у него был меньший фронтальный босс, чем S. vadum , [ 25 ] [ 27 ] и, кажется, в целом было более грейлевым. [ 28 ]
Посткраниальный скелет
[ редактировать ]




стегокеров Позвоночный столб не полностью известен. Артикуляция между зигагофиз (суставные процессы) последовательных дорсальных (заднего) позвонков, по -видимому, предотвратило движение бокового столбца позвоночной колонны, что сделало его очень жестким, и оно было дополнительно укреплено с помощью окостененных сухожилий . [ 3 ] Несмотря на то, что позвонки шеи не известны, проникновение в затылочный мыщел (который сочтен с первой вертерой шеи) указывает на то, что шея удерживалась в изогнутой позе, такой как «S»- или «U»- пояс большинства шеей динозавров. [ 35 ] Основываясь на их положении в Homalocephale , сортифицированные сухожилия, найденные в UALVP 2, сформировали бы сложную « хвостовую корзину » в хвосте, состоящие из параллельных рядов, причем конечности каждого сухожилия связываются с следующей последовательно. Такие структуры называются Myorhabdoi и в остальном известны только в телеострной рыбе ; Эта особенность уникальна для пахицефалозавров среди тетраподных (четырехногленных) животных, и, возможно, функционировал в жесткости хвоста. [ 9 ]
Лопатка плечевой (лопатка) была длиннее кости (кость верхней части руки); Его лезвие было стройным и узким, и слегка скрученным после контура ребер. Лопатка не расширилась на верхней части, но была очень расширена у основания. Коракоид был в основном тонким и похожим на тарелку. У плечевой кости был стройный вал, был слегка скручен по его длине и был слегка поклоняться. Дельтопекторный гребень (где прикреплены дельтовидные и грудные мышцы) был слабо развит. Концы локтевой кости были расширены, а хребты простирались вдоль вала. Радиус был более надежным, чем локтевая, что необычна. Когда они видели сверху, тазовый пояс был очень широким для двуночного архозавра и стал шире к задней части. Широта области таза, возможно, приспособила к заднему расширению кишечника. был Подвздошной подвздошной кости удлинен, а иший был длинным и стройным. Хотя лобки не известен, он, вероятно, был уменьшен в размерах, как у Homalocephale . Бедра а (бедра) была стройной и изогнутой, голени была стройной и извращенной, Невозь была стройной и широкой на верхней части. ноги Похоже, что плюсневка была узкой, а единственная известная конусная (костяная кость) ног было стройным и слегка изогнутым. [ 3 ] Хотя конечности стегокеров не совсем известны, они, скорее всего, были похожи на другие пахицефалозавры, имеющие руки с пятью пальцами и четыре пальца. [ 33 ]
Классификация
[ редактировать ]
В течение 1970 -х годов было описано больше родов Pachycephalosaur от азиатских окаменелостей, которые предоставили больше информации о группе. В 1974 году Марьянска и Осмольска пришли к выводу, что пахицефалозавры достаточно различны, чтобы оправдать свой собственный подряд в Ornithischia, Pachycephalosauria. В 1978 году китайский палеонтолог Донг Жиминг раскололся пахицефалозаурию на две семьи; Голова купола Pachycephalosauridae (включая Stegoceras ) и плоские головы Homalocephalidae (первоначально пишется Homalocephaleridae). [ 36 ] Уолл и Галтон не нашли статус подряд для пахицефалозавров, оправданных в 1979 году. [ 15 ] К 1980 -м годам сродство пахицефалозавров в Ornithischia было нерешенным. Основными конкурирующими взглядами было то, что группа была наиболее близкой к орнитоподам или цератопсам, последний взгляд из -за сходства между скелетом стегокеров и «примитивными» цератопскими протокератопами . В 1986 году американский палеонтолог Пол Серено поддержал взаимосвязь между пахицефалозаврами и цератопсами, а также объединила их в групповой марниноцефалии , основываясь на аналогичных черепных чертах, таких как «шельфа»-структуру над затылкой. Он признал, что доказательства этой группировки не были ошеломляющими, но достоверность группы была поддержана Суэс и Гальтоном в 1987 году. [ 3 ]
By the early 21st century, few pachycephalosaur genera were known from postcranial remains, and many taxa were only known from domes, which made classification within the group difficult. Pachycephalosaurs are thus mainly defined by cranial features, such as the flat to domed frontoparietal, the broad and flattened bar along the postorbital and squamosal bones, and the squamosal bones being deep plates on the occiput.[ 31 ] В 1986 году Серено разделил пахицефалозавров на различные группы, основываясь на масштабах доминга их черепов (сгруппированных в неверные таксоны, такие как «Tholocephalidae» и «Domocephalinae»), а в 2000 году он рассматривал «частично Стегокер » Переход между предположительно «примитивным» плоским головом и продвинутыми «полностью» куполообразными родами (такими как Pachycephalosaurus ). [ 37 ] В последующие годы отделение пахицефалозавров с головами/плоским головом было заброшено, поскольку плоские головы считались педоморфными (юноподобными) или полученными признаками в большинстве ревизий, но не сексуально- диморфными чертами. В 2006 году Салливан выступил против идеи, что степень доминга была полезна при определении таксономических сродств между пахицефалозаврами. [ 31 ] В 2003 году Салливан обнаружил, что сам Stegoceras более базальными (или «примитивом»), чем «полностью домовые» члены подсемейства Pachycephalosaurinae, разработав выводы, сделанные Серено в 1986 году. [ 4 ]
2013 года Филогенетический анализ , проведенный Эвансом и его коллегами, показал, что некоторые плоские роды пахицефалозавра были более тесно связаны с «полностью» куполообразными таксонами, чем с «не полностью» куполообразными стегокерами , которые предполагают, что они представляют несовершеннолетни Укажите таксономическую сродство. [ 38 ] Кладограмма в Pachycephalosauridae , ниже показывает размещение Stegoceras согласно Schott and Colleagues, 2016: [ 20 ]

| Pachycephalosauria |
| ||||||||||||||||||||||||
Биогеография и ранняя эволюционная история пахицефалозавров плохо изучены и могут быть прояснены только новыми открытиями. Пахицефалозавры внезапно появляются в ископаемом отчете и присутствуют как в Северной Америке, так и в Азии, поэтому они неизвестны, когда они впервые возникли, и с какого направления они рассеялись. Самые старые известные члены группы (такие как Acrotholus ) являются «полностью куполообразными» и известны на Сантонианской стадии позднего мелового периода (около 84 миллионов лет назад). Это до того, как якобы более примитивные стегокеры из среднего кампании (77 миллионов лет назад) и Хомалоцефала от раннего маастрихта (70 миллионов лет назад), поэтому добыча черепа может быть гомопластичной чертой (форма сходящейся эволюции ). Позднее появление пахицефалозавров по сравнению со связанными цератопсинами указывает на длительную линию призрака (выводится, но отсутствует в результате ископаемого), охватывающей 66 миллионов лет, от поздней юры до мелового. Поскольку пахицефалозавры были в основном небольшие, это может быть связано с Тафономическая предвзятость ; Меньшие животные с меньшей вероятностью будут сохранены с помощью окаменелости. Более деликатные кости также менее склонны к сохранению, поэтому пахицефалозавры в основном известны из их надежных черепов. [ 4 ] [ 38 ]
Палеобиология
[ редактировать ]Механика кормления
[ редактировать ]
Неясно, что съели пахицефалозавры; Имея очень маленькие, жесткие зубы, они не могли жевать жесткие, волокнистые растения так эффективно, как другие динозавры того же периода. Предполагается, что их острые зубчатые зубы идеально подходили для смешанной диеты листьев, семян, фруктов и насекомых. [ 39 ] У Stegoceras , возможно, была полностью травоядная диета, так как зубные короны были похожи на короны игуанидных ящериц. Легкие зубов показывают аспекты износа от контакта с предпринятой костью, а верхнечелюстные зубы имеют двойные аспекты, похожие на те, которые наблюдаются у других орнитишских динозавров. Каждый третий верхнечелюстный зуб UALVP 2 извергает замену зубов , а замена зубов произошла в обратном прогрессии в последовательных тройках. Затылочная область Stegoceras была хорошо сработана для мышечной привязки, и считается, что движение челюсти стегокеров и других пахицефалозавров в основном ограничивалось движениями вверх и вниз с небольшими возможностями вращения челюсти. Это основано на структуре челюсти и стоматологического микровождества, а также износ зубов указывают на то, что для сдвига использовался больший укус, чем для раздавливания. [ 3 ] [ 40 ]
В 2021 году канадский палеонтолог Майкл Н. Хаджинс и его коллеги исследовали зубы стегокеров и Thescelosaurus и обнаружили, что, хотя у обоих были зубы гетеродонта, они могут быть статистически отличаются друг от друга. Из-за широкого ростюма и более однородных зубов стегоцерас был неизбирательным корм для объема, который обрезал большое количество растительности, в то время как зубы и узкая рострума Thescelosaurus указывают, что это был селективный питатель. Пахицефалозавры и теселозавриды встречаются в одних и тех же североамериканских формациях, и, похоже, их сосуществование стало возможным благодаря им, занимающим различные экоморфопросцы (хотя стегокеры и сами Thescelosaurus не были современниками). [ 41 ]
Носовые отрывки
[ редактировать ]В 1989 году Эмили Б. Гриффин обнаружила, что у стегокеров и других пахицефалозавров было хорошее обоняние (олихание), основанное на изучении краниальных эндокастов , которые показали большие обонятельные луковицы в мозге. [ 42 ] В 2014 году Джейсон М. Бурк и его коллеги обнаружили, что Stegoceras потребовались бы хрящевые носовые турбинаты в передней части носовых проходов для воздушного потока, чтобы достичь обонятельной области. Свидетельством наличия этой структуры является костный гребень, к которому он мог прикрепить. Размер обонятельной области также указывает на то, что у стегокеров было острое обоняние. The researchers found that the dinosaur could have had either a scroll-shaped turbinate (like in a turkey ) or a branched one (as in an ostrich ) as both could have directed air to the olfactory region. The blood vessel system in the passages also suggest that the turbinates served to cool down warm arterial blood from the body that was heading to the brain. Череп S. vaduum образец UALVP 2 был подходит для исследования такого рода из -за его исключительного сохранения; Он сортировал мягкие ткани в носовой полости, которая в противном случае была бы хрящной и, следовательно, не сохранилась с помощью минерализации. [ 43 ]
Онтогенетические изменения
[ редактировать ]
Исторически было предложено несколько объяснений для вариаций, наблюдаемых в черепах стегокеров и других пахицефалозавров. был сексуального диморфизма в степени предположили, что половой диморфиз Браун и Шлайкджер Эта идея была поддержана морфометрическим исследованием 1981 года Шампоном и его коллегами, которое обнаружило, что у мужчин были более крупные и более толстые купола. [ 34 ] [ 44 ] After other flat-headed pachycephalosaurs were discovered, the degree of doming was proposed to be a feature with taxonomic importance, and AMNH 5450 was therefore considered a distinct taxon from 1979 onwards. In 1998, Goodwin and colleagues instead proposed that the inflation of the dome was an ontogenetic feature that changed with age, based on a histological study of an S. validum skull that showed the dome consisted of vascular, fast-growing bone, consistent with an Увеличение домунга до возраста. Эти авторы обнаружили, что предположительно различные особенности Ornatotholus могут легко быть результатами онтогенерии. [ 34 ] [ 17 ]
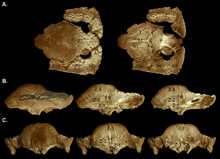
В 2003 году Уильямсон и Карр опубликовали гипотетическую серию роста S. vaduum , показывающая Ornatotholus как юношескую стадию. Они предположили, что несовершеннолетние характеризовались плоской, утолщенной лобовой крышей, с большей супратропоральной фенестрами, и усыпаны с близко расположенными бугорками и узлами. Покрытие Parietosquamosal не уменьшилось в размере, а фронтопариетальный шов был открыт. У суб-взрослев были куполы, похожие на насыпью, с задней частью париетальной и черепа-крышки плоской. Супратропорный фенестра показал асимметрию по размеру, и закрытие лобнопариетального шва было переменным. Узлы растягивались или почти уничтожены, когда купол расширился во время роста, при этом оставалась тесраированная поверхность. Схема часто стерла в самой высокой точке (вершина) купола, площадь, где произошло максимальное расширение. Бугорья на черепе были растянуты в разных направлениях, и те, кто находился на краю на шельфе Parietosquamosal, могли быть гипертрофированными (увеличенными) бугорками. Задняя и боковые стороны суб-адольты и взрослых черепа были украшены менее модифицированными бугорками. Прежде чем быть включенным в увеличивающий купол, кости черепа расширились, что привело к соединениям между этими костями. Взрослый купол был широким и выпуклым, и включал большую часть полки, которая была уменьшена в размерах и затягивала затылку как толстую «губу». Супратипуральные фенестра были закрыты, но шов между лобнопариетальными и подключенными костями черепа не всегда был закрыт у взрослых и субаду. [ 19 ]
В 2011 году Шотт и его коллеги провели более полный анализ онтогенерию черепного купола в S. vadum . Исследование показало, что на шельфе Parietosquamosal сохранилась расположение орнамента на протяжении всего роста, и что сосудистость лобнопариетальных куполов уменьшалась с размерами. Он также обнаружил, что форма и размер купола сильно коррелировали с ростом, и что рост был аллометрическим (в отличие от изометрического ) от плоского до купола, поддерживающего Ornatotholus в качестве ювенильных стегокеров . Они также предположили, что эта модель роста купола, с драматическими изменениями от ювенильного до взрослого, была общей траекторией развития пахицефалозавров. Эти исследователи отметили, что, хотя наблюдение Уильямсона и Карра о том, что супратропоральные фенестра были закрыты с возрастом, в целом было правильным, все еще была высокая степень индивидуальных различий в размере этих фенестров, независимо от размера фронтопариетали. не зависел от онтогенез. [ 34 ]
A 2012 study by Schott and Evans found that the number and shape of the individual nodes on the squamosal shelf of the examined S. validum skulls varied considerably, and that this variability does not seem to correlate with ontogenic changes, but was due to individual variation Полем Эти исследователи не обнаружили корреляции между шириной супратропоральных фенестров и размером сквомозали. [ 45 ]
Купольная функция
[ редактировать ]Функция Pachycephalosaur Doomes обсуждается, и Stegoceras использовалась в качестве модели для экспериментов в различных исследованиях. Купол был в основном интерпретируется как оружие, используемое во внутриспецифическом бою , сексуальном дисплее или средстве для распознавания видов . [ 46 ] [ 47 ]
Боевой
[ редактировать ]
The hypothesis that the domed skulls of Stegoceras and other pachycephalosaurs were used for butting heads was first suggested by American palaeontologist Edwin Colbert in 1955. In 1970 and 1971, Galton elaborated on this idea, and argued that if the dome was simply ornamental, it would были менее плотными, и что структура была идеальной для сопротивления силы. Galton suggested that when Stegoceras held its skull vertically, perpendicular to the neck, force would be transmitted from the skull, with little chance of it being dislocated, and the dome could therefore be used as a battering-ram. He believed it was unlikely to have been used mainly as defence against predators, because the dome itself lacked spikes, and those of the parietosquamosal shelf were in an "ineffective" position, but found it compatible with intra-specific competition. Галтон представлял, что купола были избиты вместе, в то время как позвоночная колонна удерживалась в горизонтальном положении. This could either be done while facing each other while dealing blows, or while charging each other with lowered heads (analogous to modern sheep and goats). Он также отметил, что жесткость спины была бы полезна при использовании головы для этой цели. In 1978, Sues agreed with Galton that the anatomy of pachycephalosaurs was consistent with transmitting dome-to-dome impact stress, based on tests with модели с плекси-стеклом . Влияние будет поглощено через шею и тело, а связки и мышцы шеи предотвратят травмы, продлив удары (как у современных снежных баров ). Сувсы также предположили, что животные могли бы заострить фланги друг друга. [ 35 ] [ 48 ] [ 49 ]
In 1997, the American palaeontologist Kenneth Carpenter pointed out that the dorsal vertebrae from the back of the pachycephalosaur Homalocephale show that the back curved downwards just before the neck (which was not preserved), and unless the neck curved upwards, the head would point to земля. He therefore inferred that the necks of Stegoceras and other pachycephalosaurs were held in a curved posture (as is the norm in dinosaurs), and that they would therefore not have been able to align their head, neck, and body horizontally straight, which would be необходимо передавать стресс. Их шеи должны быть удерживаны ниже уровня спины, что рискует нанести ущерб спинному мозгу при ударе. Modern bighorn sheep and bison overcome this problem by having strong ligaments from the neck to the tall neural spines over the shoulders (which absorb the force of impact), but such features are not known in pachycephalosaurs. Эти животные также поглощают силу удара через пазухие камеры у основания их рогов, а лоб и рога образуют широкую контактную поверхность, в отличие от узкой поверхности куполов пахицефалозавра. Потому что купол Stegoceras был округлен, он дал бы очень небольшую площадь для потенциального воздействия, и купола оторвались бы друг от друга (если бы воздействие не было сосредоточено). У борьбы с пахицефалозаврами было бы трудно видеть друг друга, пока их головы были понижены из -за костных хребтов над глазами. [ 35 ]


Из-за проблем, которые он обнаружил с гипотезой с головой, Карпентер вместо этого предположил, что купола были адаптацией для бокала-бока (как видно у некоторых крупных африканских млекопитающих); Он представлял, что два животных будут стоять параллельно, обращенные друг на друга или одно и то же направление, и прямое удары в сторону противника. Следовательно, относительно большая ширина тела пахицефалозавров может служить для защиты жизненно важных органов от вреда во время рукавов. Вполне возможно, что стегокеры и подобные пахицефалозары могли бы нанести удары с движением шеи с боковой стороны и вращением головы. Верхние стороны купола имеют наибольшую площадь поверхности и, возможно, были точкой удара. Толщина купола увеличила бы силу удара по бокам, и это гарантирует, что противник почувствовал силу удара, не будучи серьезно ранеными. Костный обод над орбитой мог защитить глаз агрессора при нанесении удара. Карпентер предположил, что пахицефалозавры впервые занялись Показ угрозы , блуждая и представляя свои головы, чтобы показать размер их куполов (запугивание), а затем нанести удары друг другу, пока один противник не сигнализирует о подчинении. [ 35 ]
В 2008 году Эрик Снайти и Адам Кокс проверили производительность 2D и 3D -пахицефалозавра с помощью анализа конечных элементов и обнаружили, что они могут противостоять значительному воздействию; Большая сводка куполов позволил привести к более высоким силам воздействия. Они также считали вероятным, что купола пахицефалозавра были покрыты в ) и , следовательно кератине, сильный материал, который может выдерживать большую энергию, не будучи постоянно поврежденным (например, остеодермы крокодилов , включить кератин в их тестовую формулу. [ 50 ] In 2011, Snively and Jessica M. Theodor conducted a finite element analysis by simulating head-impacts with CT scanned skulls of S. validum (UALVP 2), Prenocephale prenes and several extant head-butting artiodactyls . They found that the correlations between head-striking and skull morphologies found in the living animals also existed in the studied pachycephalosaurs. Stegoceras and Prenocephale both had skull shapes similar to the bighorn sheep with cancellous bone protecting the brain. They also shared similarities in the distribution of compact and cancellous regions with the bighorn sheep, white-bellied duiker and the giraffe . The white-bellied duiker was found to be the closest morphological analogue to Stegoceras ; У этого вида с головой есть купол, который меньше, но аналогично округл. Stegoceras was better capable of dissipating force than artiodactyls that butt heads at high forces, but the less vascularized domes of older pachycephalosaurs, and possibly diminished ability to heal from injuries, argued against such combat in older individuals. Исследование также проверило влияние кератинового покрытия купола и обнаружило, что он помогает в производительности. Хотя Stegoceras lacked the pneumatic sinuses that are found below the point of impact in the skulls of head-striking artiodactyls, it instead had vascular struts which could have similarly acted as braces, as well as conduits to feed the development of a keratin covering.[ 51 ]


В 2012 году Калеб М. Браун и Энтони П. Рассел предположили, что жесткие хвосты, вероятно, не использовались в качестве защиты от боя, но, возможно, позволили животным занять тройную позицию во время внутриспецифического боя, а хвост в качестве поддержки Полем Браун и Рассел обнаружили, что хвост может помочь в сопротивлении сжимающему, растяжению и кручке, когда животное наносило или получило удары с куполом. [ 9 ] Исследование, проведенное Джозефом Э. Петерсоном и коллегами в 2013 году, выявило поражения в черепах стегокеров и других пахицефалозавров, которые интерпретировались как инфекции, вызванные травмой. Поражения были обнаружены на 22% отбранных черепа пахицефалозавра (частота, согласованная по родам), но отсутствовали в образцах с плоской головой (которые были интерпретированы как несовершеннолетние или женщины), что согласуется с использованием во внутриспецифической боевых действиях (для территории или товарищи). Распределение поражений у этих животных имело тенденцию концентрироваться на вершине купола, что поддерживает поведение с головой. Благодаря флангу, вероятно, приведет к меньшему количеству травм, что вместо этого будет сосредоточено на сторонах купола. Эти наблюдения были сделаны при сравнении поражений с тем, что на черепах и флангах современных скелетов овец. Исследователи отметили, что современные носительские животные используют свое оружие как для боя, так и для демонстрации, и что пахицефалозавры могли, следовательно, также могли бы использовать свои купола для обоих. Показ оружия и готовность использовать его может быть достаточно, чтобы урегулировать споры у некоторых животных. [ 46 ]
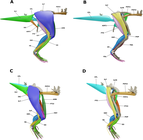
Брайан Р.С. Мур и его коллеги исследовали и реконструировали мускулатуру конечностей стегокеров в 3D в 2022 году, используя очень полный образец UALVP 2 в качестве основы. Они обнаружили, что мускулатура передних конечностей была консервативной, особенно по сравнению с ранними двуногими сауришскими динозаврами, но мускулатура таза и задних конечностей вместо этого была более полученной (или «продвинутой») из -за особенностей скелета. В этих областях были большие мышцы, и в сочетании с широким тазом и крепкими задние конечности (и, возможно, увеличенные связки), это привело к сильной стабильной структуре таза, которая помогла бы во время головы между людьми. Поскольку купола-черепа пахицефалозавров росли с положительной аллометрией и, возможно, использовались в бою, эти исследователи предположили, что это могло быть и для мышц задних конечностей, если они использовались для продвижения вперед организма во время выпадения головы. Они предупредили, что, хотя UALVP 2 очень завершен для пахицефалозавра, их исследование было ограничено тем, что ему не хватало больших частей его позвоночных колонн и элементов внешней конечности. [ 52 ]
Другие предлагаемые функции
[ редактировать ]
В 1987 году Дж. Кейт Ригби и его коллеги предположили, что пахицефалозавры купола были органами теплообмена , используемыми для терморегуляции , на основе их внутренних «излучающих структур» ( TrabeCulae ). Эта идея была поддержана несколькими другими авторами в середине 1990-х годов. [ 47 ] В 1998 году Гудвин и его коллеги рассмотрели отсутствие пазух в черепе стегокеров и сети сосудистой кости, похожей на « сото » в куполе, не подходящей для разбивания головы, и указал, что кости, прилегающие к риску купола. Во время такого контакта. Опираясь на идею о том, что окостененные сухожилия, которые укрепляли хвосты стегокеров и других пахицефалозавров, позволили им занять тройку (впервые предложено Марианской и Осмольской в 1974 году), Goodwin et al. предположил, что эти структуры могли бы защитить хвост от боковых ног, или что сам хвост мог быть использован в качестве оружия. [ 17 ] В 2004 году Гудвин и его коллеги изучили черепную гистологию пахицефалозавров и обнаружили, что сосудистость (включая трабекулы) куполов снизилась с возрастом, что они обнаружили, что они не соответствуют функции в рамках головы или тепло. Они также предположили, что плотный слой волокон Шарпи вблизи поверхности купола указывал на то, что в жизни было внешнее покрытие, что делает невозможным узнать форму купола у живого животного. Эти исследователи вместо этого пришли к выводу, что купола были в основном для распознавания видов и общения (как в некоторых африканских боидах ), и что использование в сексуальном дисплее было только вторичным. Они также предположили, что внешнее покрытие куполов было ярко окрашено в жизни или, возможно, изменило цвет сезонно. [ 47 ]
В 2011 году американские палеонтологи Кевин Падиан и Джон Р. Хорнер предположили, что «странные структуры» в динозаврах в целом (включая купола, излишеств, рога и гребни) в основном использовались для распознавания видов и отклоняли другие объяснения, как не поддерживаемые доказательствами. Среди других исследований эти авторы назвали статью Goodwin et al. 2004 года о куполах Pachycephalosaur как поддержку этой идеи, и они указали, что такие структуры не представляются сексуально диморфным. [ 53 ] В ответ на Падиан и Хорнер в том же году Роб Дж. Кнелл и Скотт Д. Сэмпсон утверждали, что распознавание видов не маловероятно в качестве вторичной функции для «странных структур» у динозавров, но и этот половой отбор (используется в дисплее или борьбе с конкурировать за друзей) было более вероятным объяснением из -за высокой стоимости их развития и потому, что такие структуры, по -видимому, сильно варьируются в видах. [ 54 ] В 2013 году британские палеонтологи Дэвид Э. Хон и Даррен Найш раскритиковали «гипотезу распознавания видов» и утверждали, что ни один существующий животный не использует такие структуры, прежде всего, для распознавания видов, и что Падиан и Хорнер проигнорировали возможность взаимного сексуального отбора (где Оба пола украшены). [ 55 ]
В 2012 году Шотт и Эванс предположили, что регулярность в сквамозной орнаменте по всему онтогенерию стегокеров соответствовала распознаванию видов, но переход от плоской к куполообразным фронталам в позднем возрасте предполагает, что функция этой особенности изменилась через онтоген. Выбран, возможно для внутриспецифического боя. [ 45 ] Дайер и коллеги, обнаруженные в 2023 году, что образцы Stegoceras различались по толщине фронназального босса, и что черепа с большинством патологий кости были таковыми с самыми высокими боссами, которые, по их мнению, показали, что вариация толщины босса представляет межсексуальную вариацию. [ 22 ] В 2023 году Хорнер и его коллеги заявили, что с тех пор, как купол и связанное с этим орнамент стегокеров и орнамент Pachycephalosaurus развивались в начале жизни, это указывает на то, что они использовались для визуального общения, так что несовершеннолетние могли распознавать другие несовершеннолетние и взрослые. Они не исключали, что эти функции могли быть использованы для других целей, включая опадение головы, но не рассматривали травму, рассматриваемую в образцах в качестве доказательства для этого. Они также предположили, что особенности в некоторых черепах Pachycephalosaurid указывают на то, что купол поддерживал бы большую кератиновую структуру, чем просто кепку. [ 56 ]
Палеоэрона
[ редактировать ]
S. Validum известен из поздней меловой группы Belly River Group (канадской эквивалент группы Джудит Ривер в США), и образцы были обнаружены в формировании парка динозавров (покойный кампанан, от 76,5 до 75 млн лет) в Парке провинции Динозавр (включая образец лектотипа) и формация Oldman (средний кампаний, от 77,5 до 76,5 млн лет) Альберты, Канада. Пахицефалозавры Ханссюсея (если не синоним стегокеров ) и форамефейл также известны из обоих формирования. [ 4 ] [ 22 ] S. novomexicanum известен из Fruitland (покойный кампания, около 75 млн лет) и нижней киртлендской формы (покойный кампанайский , около 74 млн лет) Нью -Мексико, и если бы этот вид правильно принадлежит к стегокерам , у этого рода было бы широкое географическое распределение Полем [ 25 ] Присутствие подобных пахицефалозавров на западе и севере от Северной Америки во время последних меловых показывает, что они были важной частью фаун динозавров. [ 27 ]
Традиционно предполагалось, что пахицегалозавры населяли горную среду; Износ их черепов, предположительно, был результатом того, что они были проткнуты водой из нагорья, а сравнения с снежными короками усиливали теорию. В 2014 году Джордан С. Мэллон и Эванс оспаривали эту идею, поскольку износ и оригинальные места черепа не соответствуют тому, чтобы быть транспортируемыми таким образом, и вместо этого они предположили, что североамериканские пахицефалозавры населяли аллювиальные (связанные с водой) и и и Прибрежная равнина . [ 57 ]
Формирование парка динозавров интерпретируется как низкоприемная обстановка рек и поймы , которые стали более болотистыми и под влиянием морских условий с течением времени, когда западный внутренний морской путь нарушался на запад. [ 58 ] Климат был теплее , чем современная Альберта, без мороза , но с более влажными и более сухими сезонами. Хвойные деревья, были доминирующими растениями навеса , с подлеской папоротников по -видимому , , папоротников и покрытосеменных . [ 59 ] Парк Динозавров известен своим разнообразным сообществом травоядных. Помимо стегокеров , формация также принесла окаменелости цератопса , стиракозавра и хасмозавра , хадрозавридов просоуролофуса , ламбеозавра , грипозавра , коритозавра и парасавралофа а анкилозавра , также центрозавра . Присутствующие тероподы включают тиранозавриды Gorgosaurus и Daspletosaurus . [ 60 ] Другие динозавры, известные из формирования Олдмана, включают хадрозавр брахилофозавра , цератопсанские коронозавра и альбертацератопс , орнитомимиды , теризинозавры и, возможно, анкилозавры. Тероподы включали Troodontids , Oviraptorosaurs , Dromaeosaurid Sauronitholestes и, возможно, тираннозавр альбертозавра. [ 61 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Lambe, LM (1902). «Новые роды и виды из серии реки живота (средний цвет)» . Геологическая служба Канады, вклад в канадскую палеонтологию . 3 : 68.
- ^ Jump up to: а беременный в Хэтчер, JB; Lull, Rs; Болото, OC; Осборн, HF (1907). «Цератопсия» . Монографии Геологической службы США . Xlix . doi : 10.5962/bhl.title.60500 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Sues, HD & Galton, PM (1987). «Анатомия и классификация североамериканской пахицефалозаврии (Dinosauria: Ornithischia)» . Palaeontographica abteilung a . 198 : 1–40.
- ^ Jump up to: а беременный в дюймовый и фон глин Салливан, 2003). Пересмотр . А 23 (1): 181–2 doi : 2003) [181: TDSL] 10.1671/0272-4634 ( S2CID 85894105 .
- ^ Nopcsa, F. (1903). «О стегокерах и стереоцефалии ». Центральный лист для минералогии, геологии и палеонтологии . 1903 : 266–267.
- ^ Lambe, LM (1903). «Недавняя зоопалеонтология» . Наука . 18 (445): 60. Bibcode : 1903sci .... 18 ... 60L . doi : 10.1126/science.18.445.60 . JSTOR 1631645 . PMID 17746863 .
- ^ Lambe, LM (1918). «Блицкий род Stegoceras, типичный новую семью, направленную временно в стегозаврию» . Сделки Королевского общества Канады . 12 (4): 23–36.
- ^ Гилмор, CW (1924). «На Troodon Validus , ортоподном динозавре из мелового чулота реки живота Альберта, Канада». Кафедра геологии, Университет Альберты Бюллетень . 1 : 1–43.
- ^ Jump up to: а беременный в дюймовый Браун, CM; Рассел, AP; Фарке, А.А. (2012). «Гомология и архитектура каудальной корзины Пахицефалозаурии (Dinosauria: Ornithischia): первое вхождение миоруса в тетрапеде» . Plos один . 7 (1): E30212. BIBCODE : 2012PLOSO ... 730212B . doi : 10.1371/journal.pone.0030212 . PMC 3260247 . PMID 22272307 .
- ^ Jump up to: а беременный Браун, Б.; EM, Schlaikjer (1943). «Изучение динозавров Троудонта, с описанием нового рода и четырех новых видов». Бюллетень AMNH . 82 HDL : 2246/387 .
- ^ Стернберг, CM (1945). «Pachycephalosauridae, предложенные для динозавров с куполом, Stegoceras Lambei , n. Sp., Описано». Журнал палеонтологии . 19 (5): 534–538. JSTOR 1299007 .
- ^ Рассел, Л.С. (1948). «ДАНТАРИЯ ТРОЕДОНА , род динозавров Theropod». Журнал палеонтологии . 22 (5): 625–629. JSTOR 1299599 .
- ^ Bohlin, B., 1953. Fossil Reptilies от Монголии и Кансу. Отчеты из научной экспедиции в северо-западные провинции Китая под руководством доктора Свена Хедина. VI Палеонтология позвоночных 6. Сино-шведская экспедиционная публикация 37: 1–113
- ^ Kuhn O., 1964, Fossilis Catalog 1: Animalia 105. Ornithischia (приложение 1) , ijsel pers, deventer, 80 pp
- ^ Jump up to: а беременный Стена, WP; Galton, PM (1979). «Заметки о динозаврах Pachycephalosaurid (Reptilia: Ornithischia) из Северной Америки, с комментариями о их статусе Ornithopods». Канадский журнал наук о Земле . 16 (6): 1176–1186. Bibcode : 1979cajes..16.1176w . doi : 10.1139/e79-104 .
- ^ Галтон, премьер -министр; Суды, H.-D. (1983). «Новые данные о динозаврах Pachycephalosaurid (Reptilia: Ornithischia) из Северной Америки». Канадский журнал наук о Земле . 20 (3): 462–472. Bibcode : 1983cjes..20..462g . doi : 10.1139/e83-043 .
- ^ Jump up to: а беременный в Гудвин, МБ; Буххольц, EA; Джонсон, Re (1998). «Анатомия черепа и диагностика стигимолоха (Ornithischia: Pachycephalosauria) с комментариями о структурах краниальных дисплеев в агонистическом поведении». Журнал палеонтологии позвоночных . 18 (2): 363–375. Bibcode : 1998jvpal..18..363g . doi : 10.1080/02724634.1998.10011064 .
- ^ Салливан, Роберт М. (2000). « Prenocephale Edmontonensis (Brown и Schlaikjer) New Comb. И P. Brevis (Lambe) New Comb. (Dinosauria: Ornithischia: Pachycephalosauria) из верхней меловой части Северной Америки» . Музей естественной истории и науки Нью -Мексико . 17 : 177–90.
- ^ Jump up to: а беременный Уильямсон, те; Карр, Т.Д. (2003). «Новый род полученного пахицефалозавриана из Западной Северной Америки». Журнал палеонтологии позвоночных . 22 (4): 779–801. doi : 10.1671/0272-4634 (2002) 022 [0779: angodp] 2.0.co; 2 . S2CID 86112901 .
- ^ Jump up to: а беременный Шотт, RK; Эванс, округ Колумбия (2016). «Черноальные вариации и систематика отверстия бревиса на форуме и разнообразие динозавров пахицефалозавридов (Ornithischia: Cerapoda) в группе реки Belly of Alberta, Канада». Зоологический журнал Линневого общества . doi : 10.1111/Zoj.12465 .
- ^ Эванс, Дэвид С.; Браун, Калеб М.; Ты, Хайлу; Кампионе, Николас Э. (октябрь 2021 г.). «Описание и пересмотренный диагноз первого зарегистрированного Азии Пахицефалозаврида, Синоцефала Бекселли Генерал. Ноябрь, из верхнего мела внутренней Монголии, Китай». Канадский журнал наук о Земле . 58 (10): 981–992. Bibcode : 2021cajes..58..981e . doi : 10.1139/cjes-2020-0190 . S2CID 244227050 .
- ^ Jump up to: а беременный в дюймовый Дайер, Аарон; Силы, Марк; Керри, Филипп (2023). «Проблемные предполагаемые пахицефалозавриды: синхротрона µCT -визуализация светит новый свет на анатомию и таксономическую достоверность Gravitholus albertae из группы реки живота (Кампаниан) Альберты, Канада» . Анатомия позвоночных морфологии палеонтология . 10 (1). doi : 10.18435/VAMP29388 .
- ^ Салливан, RM; Люк, С.Г. (2006). " Бассейн Сан -Хуан , Нью -Мексико, Нью -Мексико Булитин 35 : 329–3
- ^ Уильямсон, те; Карр, Т.Д. (2002). «Ювенильный пахицефалозавр (Dinosauria: Pachycephalosauridae) из формации фруктов». Нью -Мексико: Геология Нью -Мексико . 24 : 67–68.
- ^ Jump up to: а беременный в дюймовый Jasinski, SE; Салливан, RM (2011). «Переоценка пахицефалозавридов из перехода Фрукленд-Киртланда (Kirtlandian, покойный кампаний), бассейн Сан-Хуан, Нью-Мексико, с описанием нового вида стегокеров и переоценкой Texascephale Langstoni » (PDF) . Запись ископаемых 3. Музей естественной истории и науки Нью -Мексико, Бюллетень . 53 : 202–215.
- ^ Watabe, M.; TSTOTBAATAR, K.; Салливан, 2011). (Pdoliaia » (PDFF) . Бюллетень 53 : 489–4
- ^ Jump up to: а беременный в дюймовый Уильямсон, те; Brusatte, SL (2016). «Пахицефалозавры (Dinosauria: Ornithischia) из верхнего мела (верхнего кампаниана) Нью -Мексико: переоценка стегокеров Novomexicanum » . Мерашные исследования . 62 : 29–43. Bibcode : 2016crres..62 ... 29w . doi : 10.1016/j.cretres.2016.01.012 .
- ^ Jump up to: а беременный Jasinski, SE; Салливан, RM (2016). «Достоверность позднего мела пахицефалозаурида stegoceras novomexicanum (Dinosauria: Pachycephalosauridae)» . В Салливане, Роберт М.; Лукас, Спенсер Г. (ред.). Запись ископаемого 5: Бюллетень 74 . Музей естественной истории и науки Нью -Мексико. С. 107–116.
- ^ GLUT, DF (1997). Динозавры: энциклопедия . Джефферсон, Северная Каролина: McFarland & Co. с. 834–838. ISBN 978-0-89950-917-4 .
- ^ Ламберт Д. (1993). Конечная книга динозавров . Нью -Йорк: Дорлинг Киндерсли. п. 155 ISBN 978-1-56458-304-8 .
- ^ Jump up to: а беременный в Салливан, 2006). Орнишка " Булитин 35 : 347–3 S2CID 4243316 .
- ^ Peczkis, J. (1995). «Последствия оценки массы тела для динозавров». Журнал палеонтологии позвоночных . 14 (4): 520–533. Bibcode : 1995jvpal..14..520p . doi : 10.1080/02724634.1995.10011575 . JSTOR 4523591 .
- ^ Jump up to: а беременный Пол, GS (2010). Полевой гид Принстона по динозаврам . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 241–242 . ISBN 978-0-691-13720-9 .
- ^ Jump up to: а беременный в дюймовый Шотт, Райан К.; Эванс, Дэвид С.; Гудвин, Марк Б.; Хорнер, Джон Р.; Браун, Калеб Маршалл; Лонгрич, Николас Р. (29 июня 2011 г.). «Черновой онтоген в стегокере validum (Dinosauria: Pachycephalosauria): количественная модель роста и вариации купола пахицефалозавра» . Plos один . 6 (6): E21092. Bibcode : 2011ploso ... 621092S . doi : 10.1371/journal.pone.0021092 . PMC 3126802 . PMID 21738608 .
- ^ Jump up to: а беременный в дюймовый Карпентер, Кеннет (1 декабря 1997 г.). «Агонистическое поведение у пахицефалозавров (Ornithischia, Dinosauria); новый взгляд на поведение с головой» . Геология Скалистых гор . 32 (1): 19–25.
- ^ Перл, А.; Osmólska, H. (1982). « Goyocephale lattimorei Gen. Et sp. N., новый плосколовый пахицефалозавр (Ornlthlschia, Dinosauria) из верхнего мела Монголии-Acta palaeontologica Polonica» . Acta Palaeontologica Polonica . 27 : 115–127.
- ^ Sereno, PC, 2000. Окаменечная запись, систематика и эволюция пахицефалозавров и цератопсийцев из Азии. 480–516 в Бентоне, MJ, MA Shishkin, DM Unwin & En Kurochkin (Eds.), Возраст динозавров в России и Монголии . Издательство Кембриджского университета, Кембридж.
- ^ Jump up to: а беременный Эванс, округ Колумбия; Шотт, RK; Ларсон, DW; Браун, CM; Райан, MJ (2013). «Самый старый североамериканский пахицефалозаврид и скрытое разнообразие малых тел-орнитишских динозавров» . Природная связь . 4 : 1828. Bibcode : 2013natco ... 4.1828e . doi : 10.1038/ncomms2749 . PMID 23652016 .
- ^ Marynska, T.; Чепмен, re; Weshampel, DB (2004). "Pachyceeplosauria" В Вешампелле, дБ; Додсон, П.; Osmóllska, H. (Eds.). Данурия (2 -е изд.). Беркли: Университет Калифорнийской прессы. стр. 464–477 . ISBN 978-0-520-24209-8 .
- ^ Набавизаде А. (2016). «Эволюционные тенденции в механике аддуктора челюсти орнитишских динозавров» . Анатомическая запись . 299 (3): 271–294. doi : 10.1002/ar.23306 . PMID 26692539 .
- ^ Хаджинс, Майкл Нейлор; Керри, Филипп Дж.; Салливан, Корвин (16 октября 2021 г.). «Стоматологическая оценка Stegoceras vaduum (Ornithischia: Pachycephalosaurdae) и Thescelosaurus Parnerectus (Ornithischia: Thescelosaurdae): выводы палеоэкологии» Мерашные исследования 130 : 105058. DOI : 10.1016/ j.cretres.2021.105058 S2CID 239253658
- ^ Гиффин, EB (1989). «Пахицефалозавр палеонеуролагии (Архосаурия: Орнитиша)». Журнал палеонтологии позвоночных . 9 (1): 67–77. Bibcode : 1989jvpal ... 9 ... 67G . doi : 10.1080/02724634.1989.10011739 . JSTOR 4523238 .
- ^ Бурк, JM; Портер, Wm. R.; Ridgely, RC; Лайсон, Тр; Шахнер, эр; Белл, PR; Witmer, LM (2014). «Вдыхая жизнь в динозавров: решание проблем восстановления мягких тканей и носового воздушного потока у вымерших видов» . Анатомическая запись . 297 (11): 2148–2186. doi : 10.1002/ar.23046 . PMID 25312371 . S2CID 4660680 .
- ^ Чепмен, re; Galton, PE. М.; Sepkoski, JJ; Уолл, WP (1981). «Морфометрическое исследование черепа пахицефалозавридного динозавра стегокеров ». Журнал палеонтологии . 55 (3): 608–618. JSTOR 1304275 .
- ^ Jump up to: а беременный Шотт, RK; Эванс, округ Колумбия (2012). «Сквамосовая онтогенция и вариация в пахицефалозаврском динозавре Стегокерас Валум Ламбе, 1902, из формации парка Динозавров, Альберта». Журнал палеонтологии позвоночных . 32 (4): 903–913. Bibcode : 2012jvpal..32..903s . doi : 10.1080/02724634.2012.679878 . JSTOR 23251281 . S2CID 84526252 .
- ^ Jump up to: а беременный Петерсон, JE; Dischler, C.; Longrich, NR; Додсон, П. (2013). «Распределения патологий черепа предоставляют доказательства для выпадения головы у динозавров с куполом (Pachycephalosauridae)» . Plos один . 8 (7): E68620. Bibcode : 2013ploso ... 868620p . doi : 10.1371/journal.pone.0068620 . PMC 3712952 . PMID 23874691 .
- ^ Jump up to: а беременный в Гудвин, МБ; Хорнер, младший (2004). «Граниальная гистология пахицефалозавров (Ornithischia: Marginocephalia) выявляет временные структуры, несовместимые с поведением с головой» (PDF) . Палеобиология . 30 (2): 253–267. Bibcode : 2004pbio ... 30..253g . doi : 10.1666/0094-8373 (2004) 030 <0253: Chopom> 2.0.co; 2 . JSTOR 4096846 . S2CID 84961066 .
- ^ Galton, PM (1971). «Примитивный динозавр с куполом (Ornithischia: Pachycephalosauridae) из нижнего мела Англии и функции купола пахицефалозавридов». Журнал палеонтологии . 45 (1): 40–47. JSTOR 1302750 .
- ^ Sues, HD (1978). «Функциональная морфология купола в динозавре пахицефалозавра». Новый ежегодный книга по геологии и палеонтологии - ежемесячные виды . 8 : 459–472.
- ^ Snivy, E.; Кокс А. (2008). «Структурная механика Pachycephalosaur Crania разрешила поведение с головой» . Palaeontologia Electronica . 11 : 1–17.
- ^ Snivy, E.; Theodor, JM (2011). «Обычные функциональные корреляты поведения на головном ударе в пахицефалозаврах стегокерас (Ornithischia, Dinosauria) и боевые артиодактилы» . Plos один . 6 (6): e21422. Bibcode : 2011ploso ... 621422S . doi : 10.1371/journal.pone.0021422 . PMC 3125168 . PMID 21738658 .
- ^ Мур, Брайан Р.С.; Ролосон, Мэтью Дж.; Керри, Филипп Дж.; Райан, Майкл Дж.; Паттерсон, Р. Тимоти; Маллон, Джордан С. (2022). «Аппендикулярная миология стегокера Validum (Ornithischia: Pachycephalosauridae) и последствия для гипотезы с головой» . Plos один . 17 (9): E0268144. Bibcode : 2022ploso..1768144M . doi : 10.1371/journal.pone.0268144 . PMC 9436104 . PMID 36048811 .
- ^ Падиан, К.; Хорнер, младший (2011). «Эволюция« странных структур »в динозаврах: биомеханика, половой отбор, социальное отбор или распознавание видов?». Журнал зоологии . 283 (1): 3–17. doi : 10.1111/j.1469-7998.2010.00719.x .
- ^ Кнелл, RJ; Сэмпсон С. (январь 2011 г.). «Странные структуры в динозаврах: распознавание видов или половой отбор? Ответ на Падиана и Хорнер: странные структуры у динозавров» . Журнал зоологии . 283 (1): 18–22. doi : 10.1111/j.1469-7998.2010.00758.x .
- ^ Hone, DWE; Найш Д. (2013). «Гипотеза распознавания видов» не объясняет наличие и эволюцию преувеличенных структур у неавиановых динозавров » . Журнал зоологии . 290 (3): 172–180. doi : 10.1111/jzo.12035 .
- ^ Хорнер, Джон Р.; Гудвин, Марк Б.; Эванс, Дэвид С. (2023). «Новый пахицефалозаврид из Адского Крик Формирования, графство Гарфилд, Монтана, США». Журнал палеонтологии позвоночных . 42 (4). doi : 10.1080/02724634.2023.2190369 .
- ^ Mallon, JC; Эванс, округ Колумбия (2014). «Тафономия и предпочтения среды обитания североамериканских пахицефалозавридов (Dinosauria, Ornithischia)». Летая . 47 (4): 567–578. Bibcode : 2014Letha..47..567m . doi : 10.1111/let.12082 .
- ^ Эберт, Дэвид А. (2005). «Геология» . В Керри, Филипп Дж.; Koppelhus, Eva Bundgaard (ред.). Парк провинции Динозавр . Издательство Университета Индианы. С. 54–82 . ISBN 978-0-253-34595-0 .
- ^ Btas, Dennis R.; Koppelhus, Eva B. (2005). "Кампанский палиноморфы " В Керри, Филипп Дж.; Koppelhus, Eva Bundgaard (ред.). провинции Динозавр Парк Издательство Университета Индианы. Стр. 101–3 ISBN 978-0-253-34595-0 .
- ^ Weishampel, DB; Барретт, премьер -министр; Кория, Ра; Le Loeuff, J.; Сюй Син; Zx; Sahni, A.; Депутат Гомани, Элизабет; Нет, CR (2004). "Распределение динозавров" В Вайшампеле, дБ; Додсон, П.; Osmólska, H. (Eds.). The Dinosauria (2nd ed.). Калифорнийский университет. Стр. 517 –606. ISBN 978-0-520-24209-8 .
- ^ Эберт, Д.А. (1997). "Джудит Ривер Клин". In Currie, Philip J. ; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан -Диего: академическая пресса. С. 199–204 . ISBN 978-0-12-226810-6 .
Внешние ссылки
[ редактировать ] СМИ, связанные со стегокерами в Wikimedia Commons
СМИ, связанные со стегокерами в Wikimedia Commons  Данные, связанные со стегокерами в Wikispecies
Данные, связанные со стегокерами в Wikispecies
