Троодон
| Троодон Временной диапазон: поздний мел ( кампан ), | |
|---|---|
 | |
| Голотип зуба в нескольких проекциях | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Троодонтиды |
| Подсемейство: | † Троодонтина |
| Род: | † Троодон Лейди , 1856 г. |
| Типовой вид | |
| † Троодон красавец Лейди, 1856p. | |
Троодон ( / ˈ t r oʊ . ə d ɒ n / TROH -ə-don ; Троодон в более старых источниках) — бывший таксон мусорных корзин и потенциально сомнительный род относительно небольших птицеподобных окончательно -теропод, динозавров известных с кампанской эпохи Поздний меловой период (около 77 млн лет назад ). Он включает по крайней мере один вид, Troodon formosus , известный из Монтаны . Обнаруженный в октябре 1855 года, T. formosus был одним из первых динозавров, обнаруженных в Северной Америке , хотя до 1877 года его считали ящерицей. Когда -то считалось, что несколько известных экземпляров троодонтид из формации Парк динозавров в Альберте были представителями этого вида. род. Однако недавний анализ, проведенный в 2017 году, показал, что этот род недиагностичен, и некоторые из этих образцов были отнесены к роду Stenonychosaurus (долгое время считалось синонимом Troodon ), некоторые — к роду Latenivenatrix , а некоторые — к роду Pectinodon . Название рода в переводе с древнегреческого означает «ранящий зуб», имея в виду зубы, которые отличались от зубов большинства других теропод, известных на момент их открытия. Зубы имеют заметные апикально ориентированные зубцы. Однако эти «ранящие» зазубрины морфометрически более похожи на растительноядных рептилий и предполагают, возможно, всеядную диету. [1]
История открытия
[ редактировать ]Ранние исследования
[ редактировать ]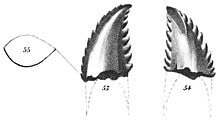
Первоначально название было написано в 1856 году как Troödon (с диарезисом ) Джозефом Лейди , а в 1876 году Sauvage официально изменил его до нынешнего статуса. [2] Типовой экземпляр троодона формации Джудит - вызвал проблемы с классификацией, поскольку весь род основан только на одном единственном зубе из Ривер . Троодон исторически представлял собой крайне нестабильную классификацию и был предметом многочисленных противоречивых синонимов с аналогичными экземплярами теропод. [3]
Зуб троодона переназначил его как «лацертилиан» ( ящерица первоначально был классифицирован Лейди как динозавра-мегалозавра ( ), но в 1901 году Франц Нопса фон Фельсо-Сильвас Megalosauridae исторически были таксоном мусорной корзины для большинства плотоядных динозавров). В 1924 году Гилмор предположил, что зуб принадлежал травоядному пахицефалозавру Stegoceras и что Stegoceras на самом деле был младшим синонимом троодона . Сходство зубов троодонтид с зубами травоядных динозавров продолжает заставлять многих палеонтологов полагать, что эти животные были всеядными. Классификация троодона как пахицефалозавра продолжалась в течение многих лет, в течение которых семейство Pachycephalosauridae было известно как Troodontidae . В 1945 году Чарльз Мортрам Штернберг отверг возможность того, что Троодон был пахицефалозавром из-за его большего сходства с зубами других плотоядных динозавров. Поскольку троодон теперь классифицируется как теропод, семейство Troodontidae больше нельзя было использовать для обозначения куполообразных динозавров, поэтому Штернберг назвал для них новое семейство. Пахицефалозавриды . [4]
Именование родственных видов
[ редактировать ]
Первые экземпляры, отнесенные к троодону и не являющиеся зубами, были найдены Штернбергом в начале 1930-х годов в формации Парка динозавров в Альберте . Первый был назван Stenonychosaurus inequalis Штернбергом в 1932 году на основании ступни, фрагментов руки и некоторых хвостовых позвонков. Примечательной особенностью этих останков был увеличенный коготь на втором пальце ноги, который теперь признан характерным для ранних паравианцев . Штернберг первоначально классифицировал Stenonychosaurus как члена семейства Coeluridae . Вторая, частичная кость нижней челюсти, была описана Гилмором (1932) как новый вид ящерицы, которую он назвал Polyodontosaurus grandis . В 1951 году Штернберг позже признал P. grandis возможным синонимом Троодона и предположил, что, поскольку у Стенонихозавра была «очень своеобразная стопа », а у Троодона «столь же необычные зубы», они могут быть тесно связаны. К сожалению, в то время не было сопоставимых образцов, позволяющих проверить эту идею. В недавней редакции материала, подготовленной ван дер Реестом и Карри, Полиодонтозавр был определен как nomen dubium , не пригодный для синонимии с другими таксонами. [5]
Более полный скелет Стенонихозавра был описан Дейлом Расселом в 1969 году из формации Парка Динозавров, которая в конечном итоге легла в основу научной основы для знаменитой скульптуры Стенонихозавра в натуральную величину , сопровождаемой его вымышленным гуманоидным потомком, «динозавроидом» . [6] Стенонихозавр стал широко известным тероподом в 1980-х годах, когда были более подробно описаны ступни и черепная коробка. Наряду с Saurornithoides образовал семейство Saurornithoididae . На основании различий в строении зубов и чрезвычайно фрагментарного характера оригинальных экземпляров Troodon formosus заурорнитоидиды считались близкими родственниками, тогда как троодон считался сомнительным возможным родственником этого семейства. Фил Карри , просматривая соответствующие образцы в 1987 году, показал, что предполагаемые различия в строении зубов и челюстей среди троодонтид и заурорнитоидид были основаны на возрасте и положении зуба в челюсти, а не на различиях в видах. Он реклассифицировал Stenonychosaurus inequalis , Polyodontosaurus grandis и Pectinodon bakkeri как младшие синонимы Troodon formosus . Карри также сделал Saurornithoididae младшим синонимом Troodontidae. [7] В 1988 году Грегори С. Пол пошел еще дальше и включил Saurornithoides mongoliensis в род Troodon как T. mongoliensis . [8] но эта реклассификация, как и многие другие односторонние синонимизации известных родов, не была принята другими исследователями. Классификация Карри всего североамериканского материала троодонтид, относящегося к одному виду Troodon formosus, получила широкое распространение среди других палеонтологов , и все экземпляры, когда-то называвшиеся Stenonychosaurus, назывались троодонами в начале 21 века в научной литературе .
Распад одновидовой модели
[ редактировать ]
Однако концепция о том, что все позднемеловые североамериканские троодонтиды принадлежат к одному виду, вскоре после публикации статьи Карри в 1987 году стала подвергаться сомнению, в том числе самим Карри. Карри и его коллеги (1990) отметили, что, хотя они считали, что все троодонтиды реки Джудит были T. formosus , окаменелости троодонтид из других формаций, таких как формация Хелл-Крик и формация Лэнс , могли принадлежать к разным видам. В 1991 году Джордж Ольшевский отнес окаменелости формации Ланс, которые сначала были названы Pectinodon bakkeri , но позже стали синонимами Troodon formosus , к виду Troodon bakkeri , а несколько других исследователей (включая Карри) вернулись к разделению окаменелостей формации Парк динозавров. как Troodon inequalis (ныне Stenonychosaurus inequalis ). [9]
В 2011 году Занно и его коллеги рассмотрели запутанную историю классификации троодонтид в позднемеловом периоде Северной Америки. Они последовали примеру Лонгрича (2008) в рассмотрении Pectinodon bakkeri как действующего рода и отметили, что, вероятно, это многочисленные экземпляры позднего мела, которые в настоящее время отнесены к Troodon formosus , но требуется более тщательный анализ образцов. Поскольку голотип T. formosus представляет собой один зуб, это делает троодона nomen dubium . [3]
В 2017 году Эванс и его коллеги продолжили обсуждение недиагностической природы голотипа Troodon formosus и предложили использовать Stenonychosaurus для получения скелетного материала троодонтид из формации Парк динозавров. [10] Позже в том же году Аарон Дж. ван дер Реест и Карри пришли к такому же выводу, что и Эванс и его коллеги, а также разделили большую часть материала, отнесенного к Stenonychosaurus, на новый род: Latenivenatrix . [5] В 2018 году Варриккио и его коллеги не согласились с Эвансом и его коллегами, сославшись на то, что Stenonychosaurus не использовался в течение тридцати лет с тех пор, как Карри и его коллеги сделали его синонимом Троодона , и указали, что « Troodon formosus остается собственным названием для этого таксона». [11] С этим выводом Варриккио согласились Селлес и его коллеги в их описании Тамарро в 2021 году . [12] Комментарии Варриккио позже были рассмотрены Калленом и его коллегами в их обзоре биоразнообразия формации Парка динозавров в 2021 году, где они отметили, что, хотя Stenonychosaurus действительно не использовался в течение 30 лет, первоначальная гипотеза Карри о субъективной синонимии (основанной на морфологии зубов и челюстей) была никогда не проверялась напрямую, и, учитывая, что более поздние исследования показали, что зубы у троодонтид ниже семейного уровня не являются диагностическими, первоначальная гипотеза Карри, следовательно, не подтверждается имеющимися данными, независимо от того, сколько времени прошло с момента ее первоначального предложения. [13] Они предположили, что описание более полного скелетного материала (т.е. содержащего зубные, лобные и посткраниальные элементы), который можно связать с голотипом, могло бы позволить провести прямую проверку гипотезы синонимии, но вновь подтвердили, что на данный момент, учитывая отсутствие Из подтверждающих доказательств синонимия троодона и стенонихозавра не может быть сохранена, и что простое отсутствие испытаний в течение 30 лет не является достаточным оправданием, чтобы принять предлагаемое объединение таксонов, не имеющих перекрывающихся диагностических материалов. [13] Однако Варриккио и другие по-прежнему настаивают на своем методе наименования.
Классификация
[ редактировать ]
Троодон считается одним из наиболее производных представителей своего семейства. Наряду с Занабазаром , Саурорнитоидесом и Талосом он образует кладу специализированных троодонтид. [3]
Ниже представлена кладограмма Troodontidae, составленная Занно и др. в 2011 году. [3]
| Троодонтиды |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]
Одно исследование было основано на нескольких зубах троодона , собранных в позднемеловых отложениях на севере Аляски. Эти зубы намного крупнее, чем те, что были собраны в более южных местах, что свидетельствует о том, что популяции троодонов на северной Аляске выросли до более крупных средних размеров тела, что намекает на правило Бергмана . В этом исследовании также представлен анализ пропорций и характера износа большой выборки зубов троодона . Предполагается, что характер износа зубов всех троодонов предполагает диету, состоящую из мягкой пищи, несовместимой с пережевыванием костей, экзоскелетами беспозвоночных или жесткими растительными продуктами. Это исследование предполагает, что диета в основном состоит из мяса. [14] Судя по грануле, возможно принадлежащей Троодону, он охотился на ранних млекопитающих, таких как Альфадон . [15]
В 2011 году из Внутренней Монголии был описан еще один производный троодонтид, Линхевенатор . Авторы отметили, что у него относительно короткие и крепкие передние конечности, а также увеличенная вторая ногтевая пластина, похожая на таковую у дромеозавридов по сравнению с более базальными троодонтидами. Было высказано предположение, что у производных троодонтид конвергентно развились большие вторые когти педали в стиле дромеозаврид, вероятно, как адаптация, связанная с хищничеством. Авторы отметили, что вполне вероятно, что это может быть применимо к другим производным троодонтидам, включая троодона , хотя в настоящее время это неясно из-за нехватки достаточных останков последнего рода. [16]
Коммунальное гнездование
[ редактировать ]В исследовании 2023 года с использованием предполагаемой яичной скорлупы троодонов из формации Олдман использовалась термометрия слипшихся изотопов для определения их формирования и развития. Исследование показало, что в отличие от ускоренной минерализации яиц у современных птиц, троодон и, вероятно, другие нептичьи манирапторы замедляли кальцификацию яиц, как и другие рептилии. Это указывает на то, что, в отличие от птиц, троодон и другие манирапторы имели два функциональных яичника , что ограничивало количество производимых яиц. Таким образом, исследование пришло к выводу, что большие кладки окаменелых яиц, присутствующие в формации, несмотря на ограниченное производство яиц у каждой особи, указывают на то, что троодон имел коллективное гнездовое поведение, при котором яйца откладывались в одном гнезде несколькими самками, образуя большие клатчи. Эту стратегию также используют некоторые современные птицы, например страусы . [17] [18]
Палеоэкология
[ редактировать ]
Типовой экземпляр Troodon formosus был найден в формации Джудит-Ривер в Монтане . Породы формации Джудит-Ривер по возрасту эквивалентны формации Олдман в Альберте . [19] который датируется периодом от 77,5 до 76,5 миллионов лет назад. [20]
относили к тому же роду, что и троодон В прошлом останки из множества других геологических формаций реки Джудит. Сейчас признано маловероятным, что все эти окаменелости, происходящие из мест, находящихся на расстоянии сотен или тысяч миль друг от друга и разделенных миллионами лет, представляют собой один вид или даже один род троодонтид. Чтобы определить, сколько видов троодонов существовало, необходимы дальнейшие исследования и больше окаменелостей. Сомнительно, что после дальнейшего изучения к Troodon можно будет отнести какой-либо дополнительный вид , и в этом случае род будет считаться nomen dubium . [3]
Останки, относящиеся к Троодону, известны из формации Принс-Крик , слоя горных пород на Аляске, который датируется от позднего кампана до маастрихта периодом позднего мела . [21] Судя по присутствию в породах гипса и пирита , можно предположить, что формация была ограничена крупным водоемом. Похоже, что, судя по наличию окаменелостей пыльцы, доминирующими растениями были деревья , кустарники , травы и цветковые растения . Температура колебалась от 2 до 12°C, что примерно соответствует 36-54°F, и, судя по положению Аляски в позднем меловом периоде, в этом районе было около 120 дней зимней темноты. [22] Этот манирапторан жил рядом со многими другими рептилиями, такими как центрозаврин Pachyrhinosaurus perotorum , разновидность зауролофинового гадрозаврида Edmontosaurus , пахицефалозаврин Alaskacephale Gangloffi , безымянный аждархидный птерозавр и тираннозавр Nanuqsaurus hoglundi . Он также жил рядом с метатерийным млекопитающим Unnuakomys hutchisoni . [23] Судя по количеству обнаруженных зубов, этот троодонтид был наиболее распространенным тероподом формации , составляя 2/3 всех экземпляров, что резко контрастирует с более южными отложениями в Монтане , где троодонтиды составляют лишь 6% всех останков теропод. . [24] Это, наряду с доказательством того, что троодон был более многочисленным в более прохладные периоды, такие как ранний маастрихт , может указывать на то, что троодон предпочитал более прохладный климат. [25]
Дополнительные экземпляры, которые в настоящее время относят к Троодону, происходят из верхней части формации Ту Медисин в Монтане . Зубы, похожие на троодона, были обнаружены в нижней части формации Джавелина в Техасе и в формации Наашойбито формации Охо Аламо в Нью-Мексико . [26] [27]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хольц, Томас Р., Бринкман, Дэниел Л., Чендлер, Кристин Л. (1998) Морфометрия зубчиков и, возможно, всеядная привычка питания теропод-динозавра Троодона. Гайя № 15. Декабрь 1998 г., стр. 159-166.
- ^ Соваж, Х.-Э. (1876). «Заметки об ископаемых рептилиях» . Бюллетень Геологического общества Франции (на французском языке) (3-я серия 4). Геологическое общество Франции: 434–444.
- ^ Перейти обратно: а б с д и Линдси Э. Занно, Дэвид Дж. Варриккио, Патрик М. О'Коннор, Алан Л. Титус и Майкл Дж. Нелл (2011). Лалуэса-Фокс, Карлес (ред.). «Новый теропод-троодонтид, Talos sampsoni gen. et sp. nov., из верхнемелового западного внутреннего бассейна Северной Америки» . ПЛОС ОДИН . 6 (9): e24487. Бибкод : 2011PLoSO...624487Z . дои : 10.1371/journal.pone.0024487 . ПМК 3176273 . ПМИД 21949721 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Штернберг, К. (1945). «Описаны Pachycephalosauridae, предложенные для куполообразных динозавров, Stegoceras lampei n. sp.». Журнал палеонтологии . 19 : 534–538.
- ^ Перейти обратно: а б ван дер Реест, AJ; Карри, Пи Джей (2017). «Троодонтиды (Theropoda) из формации Парк динозавров, Альберта, с описанием уникального нового таксона: значение для разнообразия дейнонихозавров в Северной Америке» (PDF) . Канадский журнал наук о Земле . 54 (9): 919–935. Бибкод : 2017CaJES..54..919V . doi : 10.1139/cjes-2017-0031 . hdl : 1807/78296 .
- ^ Рассел, Д.А.; Сеген, Р. (1982). «Реконструкция небольшого мелового теропода Stenonychosaurus inequalis и гипотетического динозавроида» . Силлогей . 37 : 1 –43.
- ^ Карри, П. (1987). «Тероподы формации реки Джудит». Периодическая статья Палеонтологического музея Тиррелла . 3 : 52–60.
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 398–399 . ISBN 978-0-671-61946-6 .
- ^ Карри, П. (2005). «Тероподы, включая птиц». в Карри и Коппельхус (ред.). Провинциальный парк динозавров, впечатляющая экосистема. Часть вторая. Флора и фауна парка. Издательство Университета Индианы, Блумингтон. Стр. 367–397.
- ^ Эванс, округ Колумбия; Каллен, ТМ; Ларсон, Д.В.; Рего, А. (2017). «Новый вид троодонтидного теропода (Dinosauria: Maniraptora) из формации Каньон Подкова (Маастрихт) в Альберте, Канада» . Канадский журнал наук о Земле . 54 (8): 813–826. Бибкод : 2017CaJES..54..813E . doi : 10.1139/cjes-2017-0034 .
- ^ Варриккио, диджей; Кундрат, М.; Хоган, Дж. (2018). «Промежуточный инкубационный период и примитивное вынашивание динозавра-теропода» . Научные отчеты . 8 (1): 12454. Бибкод : 2018НатСР...812454В . дои : 10.1038/s41598-018-30085-6 . ПМК 6102251 . ПМИД 30127534 .
- ^ Селлес, АГ; Вилла, Б.; Брусатте, СЛ; Карри, Пи Джей; Галобарт, А. (2021). «Быстрорастущий базальный троодонтид (Dinosauria: Theropoda) из позднего мела Европы» . Научные отчеты . 11 (1): 4855. Бибкод : 2021NatSR..11.4855S . дои : 10.1038/s41598-021-83745-5 . ПМЦ 7921422 . ПМИД 33649418 .
- ^ Перейти обратно: а б Каллен, Томас М.; Занно, Линдси; Ларсон, Дерек В.; Тодд, Эринн; Карри, Филип Дж.; Эванс, Дэвид К. (30 июня 2021 г.). «Анатомический, морфометрический и стратиграфический анализ биоразнообразия теропод в формации парка динозавров верхнего мела (кампана)» . Канадский журнал наук о Земле . 58 (9): 870–884. doi : 10.1139/cjes-2020-0145 .
- ^ Фиорилло, Энтони Р. (2008) «О появлении исключительно больших зубов троодона (Dinosauria: Saurischia) из позднего мела Северной Аляски», том 23 Палайоса, стр. 322-328
- ^ Фреймут, Уильям (2021). «Желудочные гранулы, содержащие млекопитающих, потенциально относящиеся к Troodon formosus в местности мелового периода Эгг-Маунтин, формация Ту-Медисин, Монтана, США» . Палеонтология . 64 (5): 699–725. Бибкод : 2021Palgy..64..699F . дои : 10.1111/пала.12546 . S2CID 237659529 .
- ^ Сюй X, Тан Q, Салливан С, Хан Ф, Сяо Д (2011) Короткорукий троодонтидный динозавр из верхнего мела Внутренней Монголии и его последствия для эволюции троодонтид. PLoS ONE 6(9): e22916. doi:10.1371/journal.pone.0022916
- ^ Тальявенто, Маттиа; Дэвис, Амелия Дж.; Бернекер, Мигель; Фибиг, Йенс (3 апреля 2023 г.). «Доказательства гетеротермической эндотермии и минерализации яичной скорлупы, подобной рептилии, у троодона, нептичьего теропода-манираптора» . ПНАС . 120 (15): e2213987120. Бибкод : 2023PNAS..12013987T . дои : 10.1073/pnas.2213987120 . ПМЦ 10104568 . ПМИД 37011196 .
- ^ «Троодоны откладывают яйца в общие гнезда, как современные страусы» . Популярная наука . 03.04.2023 . Проверено 5 апреля 2023 г.
- ^ Эберт, Дэвид А. (1997). «Джудит Ривер Ведж». В Карри, Филип Дж .; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 199–204 . ISBN 978-0-12-226810-6 .
- ^ Арбур, ВМ ; Бернс, Мэн; Сиссонс, Р.Л. (2009). «Переописание анкилозавридного динозавра Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) и пересмотр рода». Журнал палеонтологии позвоночных . 29 (4): 1117–1135. Бибкод : 2009JVPal..29.1117A . дои : 10.1671/039.029.0405 . S2CID 85665879 .
- ^ Фиорилло, Арканзас ; Тыкоски, RS ; Карри, Пи Джей ; Маккарти, Пи Джей ; Флайг, П. (2009). «Описание двух частичных черепных коробок троодонов из формации Принс-Крик (верхний мел), Норт-Слоуп, Аляска». Журнал палеонтологии позвоночных . 29 (1): 178–187. дои : 10.1080/02724634.2009.10010370 . S2CID 197535475 .
- ^ Дракенмиллер, Патрик С.; Эриксон, Грегори М.; Бринкман, Дональд; Браун, Калеб М.; Эберле, Джаэлин Дж. (23 августа 2021 г.). «Гнездование нептичьих динозавров в крайних полярных широтах» . Современная биология . 31 (16): 3469–3478.e5. Бибкод : 2021CBio...31E3469D . дои : 10.1016/j.cub.2021.05.041 . ПМИД 34171301 . S2CID 235631483 .
- ^ Эберле, Джаэлин Дж.; Клеменс, Уильям А.; Маккарти, Пол Дж.; Фиорилло, Энтони Р.; Эриксон, Грегори М.; Друкенмиллер, Патрик С. (26 апреля 2019 г.). «Самая северная находка Metatheria: новый позднемеловой педиомид с Северного склона Аляски» . Тейлор и Фрэнсис. doi : 10.6084/m9.figshare.8047169.v2 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ «Гигантские динозавры-троодонтиды Аляски» . Eartharchives.org .
- ^ Фиорилло, Энтони Р.; Ганглофф, Роланд А. (2000). «Зубы теропода из формации Принс-Крик (мел) на Северной Аляске, с размышлениями о палеоэкологии арктических динозавров». Журнал палеонтологии позвоночных . 20 (4): 675–682. doi : 10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2 . S2CID 130766946 .
- ^ Лэнгстон, Стендхардт и Стивенс (1989). «Коллекционирование ископаемых позвоночных в Биг-Бенд - история и ретроспектива». в области палеонтологии позвоночных, биостратиграфии и условий осадконакопления, последний мел и третичный период, район Биг-Бенд, Техас . Путеводитель полевой поездки, номера 1 a, B и 49-е ежегодное собрание Общества палеонтологии позвоночных, Остин, Техас, 29 октября - 1 ноября 1989 г. 11-21.
- ^ Вейл и Уильямсон (2000). «Разнообразная маастрихтская фауна наземных позвоночных члена Наашойбито, формация Киртланд (бассейн Сан-Хуан, Нью-Мексико) подтверждает «ланкийскую» фаунистическую гетерогенность в западной части Северной Америки». Рефераты с программами Геологического общества Америки , 32 : A-498.
- Рассел, Д.А. (1987). «Модели и картины североамериканских динозавров». В: Черкас, С. Дж. и Олсон, ЕС (ред.) Динозавры в прошлом и настоящем, Том I. Музей естественной истории округа Лос-Анджелес / Университет Вашингтона (Сиэтл и Вашингтон), стр. 114–131.
