Стенонихозавр
| Стенонихозавр Временной диапазон: кампан , Возможная позднего Маастрихта запись
| |
|---|---|
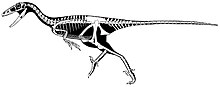
| |
| Реконструкция скелета с недостающими частями на основе родственных родов. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Троодонтиды |
| Подсемейство: | † Троодонтина |
| Род: | † Стенонихозавр Штернберг , 1932 год. |
| Типовой вид | |
| † Стенонихозавр неравный Штернберг, 1932 г.
| |
| Синонимы | |
| |
Stenonychosaurus (что означает «ящерица с узкими когтями») — род троодонтидных динозавров из позднемеловой формации Парка динозавров в Альберте , Канада, а также, возможно, из формации Два Медицина . Тип и единственный вид, inequalis , был назван Чарльзом Мортрамом Штернбергом в 1932 году на основе ступни, фрагментов руки и некоторых хвостовых позвонков из позднего мела Альберты S. . S. inequalis был переведен в 1987 году Филом Карри в род Troodon , который был отменен в результате признания Stenonychosaurus отдельным родом от, возможно, сомнительного Troodon в 2017 году Эвансом и др. а также позже в том же году Ван дер Реестом и Карри.
История открытия
[ редактировать ]Первые экземпляры, отнесенные в настоящее время к троодону и не являющиеся зубами, были найдены Штернбергом в 1928 году в формации Парка динозавров в Альберте . Первый был назван Stenonychosaurus inequalis Штернбергом в 1932 году на основании ступни, фрагментов руки и некоторых хвостовых позвонков. Примечательной особенностью этих останков был увеличенный коготь на втором пальце ноги, который теперь признан характерным для ранних паравианцев . Штернберг первоначально классифицировал Stenonychosaurus как члена семейства Coeluridae . Вторая, частичная кость нижней челюсти, была описана Гилмором (1932) как новый вид ящерицы, которую он назвал Polyodontosaurus grandis . Позже, в 1951 году, Штернберг признал P. grandis возможным синонимом Троодона и предположил, что, поскольку у Стенонихозавра была «очень своеобразная стопа », а у Троодона «столь же необычные зубы», они могут быть тесно связаны. В то время не было сопоставимых образцов, которые могли бы проверить эту идею.
Голотип Stenonychosaurus inequalis , CMN 8539, представляет собой частичный скелет, состоящий из шести хвостовых позвонков, костей кисти, дистального конца левой большеберцовой кости и таранной кости , а также полной левой стопы. К роду также было отнесено несколько черепных экземпляров: УАЛВП 52611 (почти полная крыша черепа) и ТМП 1986.036.0457 (частичная черепная коробка).

Более полный скелет Стенонихозавра был описан Дейлом Расселом в 1969 году из формации Парка Динозавров, которая в конечном итоге легла в основу научной основы для знаменитой скульптуры Стенонихозавра в натуральную величину , сопровождаемой его вымышленным гуманоидным потомком, «динозавроидом». [ 1 ]
Стенонихозавр стал широко известным тероподом в 1980-х годах, когда были более подробно описаны ступни и черепная коробка. Наряду с Saurornithoides образовал семейство Saurornithoididae . На основании различий в строении зубов и чрезвычайно фрагментарного характера оригинальных экземпляров Troodon formosus заурорнитоидиды считались близкими родственниками, тогда как троодон считался сомнительным возможным родственником этого семейства. Фил Карри , просматривая соответствующие образцы в 1987 году, показал, что предполагаемые различия в строении зубов и челюстей среди троодонтид и заурорнитоидид были основаны на возрасте и положении зуба в челюсти, а не на различиях в видах. Он реклассифицировал Stenonychosaurus inequalis, а также Polyodontosaurus grandis и Pectinodon bakkeri как младшие синонимы Troodon formosus . Карри также сделал Saurornithoididae младшим синонимом Troodontidae. [ 2 ] В 1988 году Грегори С. Пол пошел еще дальше и включил Saurornithoides mongoliensis в род Troodon как T. mongoliensis . [ 3 ] но эта реклассификация, как и многие другие односторонние синонимизации известных родов, не была принята другими исследователями. Классификация Карри всего североамериканского материала троодонтид, относящегося к одному виду Troodon formosus, получила широкое распространение среди других палеонтологов , и все экземпляры, когда-то называвшиеся Stenonychosaurus, назывались троодонами в начале 21 века в научной литературе .
Однако концепция о том, что все позднемеловые североамериканские троодонтиды принадлежат к одному виду, вскоре после публикации статьи Карри 1987 года стала подвергаться сомнению, в том числе самим Карри. Карри и его коллеги (1990) отметили, что, хотя они считали, что все троодонтиды реки Джудит были T. formosus , окаменелости троодонтид из других формаций, таких как формация Хелл-Крик и формация Лэнс , могли принадлежать к разным видам. В 1991 году Джордж Ольшевский отнес окаменелости формации Ланс, которые сначала были названы Pectinodon bakkeri , но позже стали синонимами Troodon formosus, к виду Troodon bakkeri , а несколько других исследователей (включая Карри) вернулись к сохранению окаменелостей формации Парк динозавров отдельно как Troodon inequalis. . [ 4 ]
В 2011 году Занно и его коллеги рассмотрели запутанную историю классификации троодонтид в позднемеловом периоде Северной Америки. Они последовали за Лонгричем (2008), рассматривая Pectinodon bakkeri как действительный род, и отметили, что вполне вероятно, что многочисленные позднемеловые экземпляры, в настоящее время отнесенные к Troodon formosus, почти наверняка представляют собой многочисленные новые виды, но требуется более тщательный анализ образцов. Поскольку голотип T. formosus представляет собой один зуб, это может сделать троодона nomen dubium . [ 5 ]
В 2017 году Эванс и его коллеги, основываясь на работе Занно и других, подтвердили недиагностируемую в настоящее время природу голотипа Troodon formosus и предложили использовать Stenonychosaurus для получения скелетного материала троодонтид из формации Парк динозавров. [ 6 ] Позже в 2017 году Ван дер Реест и Карри обнаружили, что Stenonychosaurus является действительным родом, но переназначили большую часть известного материала новому роду Latenivenatrix . [ 7 ]
Многие из Парка динозавров, экземпляры троодонтид когда-то отнесенные к троодонам, недавно были отнесены к Stenonychosaurus и Latenivenatrix . Экземпляры, которые сейчас относят к Stenonychosaurus, включают несколько лобных костей (UALVP 5282, TMP 1986.078.0040, TMP 1988.050.0088, TMP 1991.036.0690), частичную зубную кость (TMP 1982.019.0151) и дистальную часть левой плюсневой кости III (TMP). 1998.068.0090).
В 2021 году более полный повторный анализ морфологии и стратиграфического положения известного скелетного материала, отнесенного к Stenonychosaurus и Latenivenatrix, показал, что несколько признаков, описанных как диагностические для Latenivenatrix, на самом деле индивидуально изменчивы, что оба таксона стратиграфически перекрываются и что mcmasterae Latenivenatrix младший синоним Stenonychosaurus inequalis . [ 8 ] В результате S. inequalis остается единственным действительным таксоном троодонтид, идентифицированным в настоящее время из формации Парка динозавров.
«Динозавроид»
[ редактировать ]
В 1982 году Дейл А. Рассел , тогдашний куратор окаменелостей позвоночных в Национальном музее Канады в Оттаве, предположил возможный путь эволюции Стенонихозавра , если бы он не погиб в результате мел-палеогенового вымирания , предполагая, что он мог эволюционировать в разумные существа, сходные по строению тела с человеком. наблюдался устойчивый рост коэффициента энцефализации Рассел отметил, что с течением геологического времени среди динозавров или EQ (относительной массы мозга по сравнению с другими видами с такой же массой тела). Рассел обнаружил первый череп троодонтида и отметил, что, хотя его EQ был низким по сравнению с человеческим, он был в шесть раз выше, чем у других динозавров. Рассел предположил, что, если бы эта тенденция в эволюции стенонихозавра продолжалась до настоящего времени, размер его мозговой оболочки к настоящему времени мог бы составлять 1100 см. 3 (67 куб. дюймов), что сопоставимо с ростом человека (в среднем 1260 см). 3 (77 у.е. дюймов) для мужчин и 1130 см. 3 (69 у.е. дюймов) для женщин). [ 1 ]
У троодонтидов были полуманипулятивные пальцы, способные в определенной степени захватывать и удерживать предметы, а также бинокулярное зрение. [ 1 ] Рассел предположил, что его «Динозавроид», как и представители семейства троодонтид, имел бы большие глаза и по три пальца на каждой руке, один из которых был бы частично противоположен . Рассел также предположил, что у «Динозавроида» был беззубый клюв. Как и у большинства современных рептилий (и птиц), он считал их гениталии внутренними. Рассел предположил, что для этого потребовался бы пупок, поскольку плацента способствует развитию большого мозга. Однако у него не было молочных желез, и он кормил бы своих детенышей, как это делают некоторые птицы, срыгивающей пищей. Он предположил, что его язык чем-то напоминал пение птиц . [ 1 ] [ 9 ]
Однако мысленный эксперимент Рассела был встречен критикой со стороны других палеонтологов с 1980-х годов, многие из которых отмечают, что его динозавроид чрезмерно антропоморфен. Грегори С. Пол (1988) и Томас Р. Хольц-младший считают его «подозрительно человеческим», а Даррен Нэйш утверждал, что высокоинтеллектуальный троодонтид с большим мозгом сохранит более стандартное строение тела теропода, с горизонтальной позой и длинный хвост и, вероятно, манипулировал объектами мордой и ногами, как птица, а не человеческими «руками». [ 9 ]
Описание
[ редактировать ]
Стенонихозавр был небольшим динозавром, длиной до 2,5 метров (8,2 фута) и массой тела 35 килограммов (77 фунтов). [ 10 ] Наиболее крупные экземпляры по размерам сравнимы с дейнонихом и уненлагией . [ 11 ] У них были очень длинные и тонкие задние конечности, что позволяло предположить, что эти животные умели быстро бегать. У них были большие втягивающиеся серповидные когти на вторых пальцах ног, которые при беге поднимались над землей.
Их глаза были очень большими (возможно, свидетельствующими о частичном ночном образе жизни) и слегка обращенными вперед, что давало Стенонихозавру некоторую степень восприятия глубины . [ 12 ]
Мозг и внутреннее ухо
[ редактировать ]
У Стенонихозавра был один из самых крупных известных мозгов среди всех динозавров по сравнению с массой его тела (сравнимой с современными птицами). [ 13 ] Это было рассчитано как соотношение объема головного мозга к объему 31,5–63% от пропорции нептичьих рептилий до истинно птичьих. [ 14 ] Кроме того, у него были костные кристы, поддерживающие барабанные перепонки , которые были окостеневшими, по крайней мере, в верхней и нижней частях. Остальные кристы были либо хрящевыми, либо слишком хрупкими, чтобы их можно было сохранить. Метотическая стойка стенонихозавра была увеличена из стороны в сторону, подобно дромеозавру и примитивным птицам, таким как археоптерикс и гесперорнис . [ 14 ]
Палеобиология
[ редактировать ]
Стенонихозавр , как полагают, был хищником , и это мнение подтверждается его серповидным когтем на ноге и, по-видимому, хорошим бинокулярным зрением .
Однако зубы стенонихозавра отличаются от зубов большинства других теропод . Одно сравнительное исследование пищевого аппарата позволяет предположить, что Стенонихозавр мог быть всеядным . [ 15 ] Челюсти сходились в широком U-образном симфизе, похожем на таковой у игуаны , вида ящериц, приспособленного к растительноядному образу жизни. Кроме того, зубы Стенонихозавра имели большие зазубрины, каждая из которых называется зубчиком . На пересечении зубчиков есть ямки, а кончики зубчиков направлены к кончику или вершине каждого зуба. На боках зубов видны следы износа. Хольц (1998) также отметил, что характеристики, используемые для поддержки хищнической привычки Стенонихозавра - хватательные руки, большой мозг и стереоскопическое зрение - все это характеристики, общие с травоядными или всеядными приматами и всеядным Проционом (енотом).
Исследования по определению возраста , проведенные на троодонте Two Medicine с использованием подсчета годичных колец, показывают, что этот динозавр достиг своего взрослого размера, вероятно, через 3–5 лет. [ 16 ]
Обнаружен частичный скелет с сохранившимися следами проколов, возможно, нанесенных хищником. [ 17 ]
Воспроизведение
[ редактировать ]
Яйца и гнезда динозавров были обнаружены Джоном Р. Хорнером в 1983 году в формации Ту Медисин в Монтане. Варрикчо и др. (2002) описали восемь из таких гнезд, обнаруженных на сегодняшний день. Все они находятся в коллекции Музея Скалистых гор и имеют инвентарные номера MOR 246, 299, 393, 675, 676, 750, 963, 1139. Хорнер (1984) обнаружил отдельные кости и частичные скелеты гипсилофодонта Orodromeus. очень близко гнезда на том же горизонте и описал яйца как яйца Ородромея . [ 18 ] Хорнер и Вейшампель (1996) повторно исследовали эмбрионы, сохранившиеся в яйцах, и определили, что это эмбрионы троодона , а не ородромеуса . [ 19 ] Варриккио и др. (1997) сделали это определение с еще большей уверенностью, когда описали частичный скелет взрослого троодона (MOR 748), находившегося в контакте с кладкой, состоящей как минимум из пяти яиц (MOR 750), вероятно, в насиживающем положении. [ 20 ] Ван дер Реест и Карри считали возможным, что троодонт Two Medicine принадлежал к тому же виду, что и Stenonychosaurus . [ 7 ]
Варриккио и др. (1997) описали точное строение гнезд. Они были построены из отложений, имели форму тарелки, внутренний диаметр около 100 см (39 дюймов) и с ярко выраженным приподнятым краем, окружающим яйца. В более полных гнездах было от 16 (минимальное количество для MOR 246) до 24 (MOR 963) яиц. Яйца имеют форму удлиненных капель, с более заостренными концами, направленными вниз и примерно наполовину погруженными в осадок. Яйца откладывают под углом так, что в среднем верхняя половина оказывается ближе к центру гнезда. Нет никаких свидетельств присутствия в гнезде растительных остатков.

Варриккио и др. (1997) смогли извлечь из гнезд достаточно данных, чтобы сделать вывод о некоторых характеристиках репродуктивной биологии троодонтов. В результате выяснилось, что они, по-видимому, имели тип размножения, промежуточный между крокодилами и птицами, как и предсказывает филогения. Яйца статистически сгруппированы парами, что позволяет предположить, что у животного было два функциональных яйцевода, как у крокодилов, а не один, как у птиц. Крокодилы откладывают много яиц, небольших размеров, пропорциональных размеру тела взрослой особи. Птицы откладывают меньше, но более крупные яйца. Троодонт Two Medicine был промежуточным, откладывая яйцо весом около 0,5 кг (1,1 фунта) на взрослую особь весом 50 кг (110 фунтов). Это в 10 раз больше, чем у рептилий той же массы, но два яйца троодонтов примерно эквивалентны яйцу весом 1,1 кг (2,4 фунта), предсказанному для птицы весом 50 кг (110 фунтов).
Варриккио и др. также нашли доказательства повторяющейся откладки, когда взрослая особь могла откладывать пару яиц каждые один или два дня, а затем обеспечивать одновременное вылупление, откладывая высиживание до тех пор, пока все яйца не будут отложены. MOR 363 был обнаружен с 22 пустыми (вылупившимися) яйцами, а эмбрионы, обнаруженные в яйцах MOR 246, находились в очень схожих стадиях развития, что означает, что все детеныши вылупились примерно одновременно. Эмбрионы имели продвинутую степень развития скелета, а пустые яйца были относительно нераздавлены, что означает, что вылупившиеся птенцы были преждевременными . По оценкам авторов, общее время пребывания взрослых особей в гнезде для кладки, высиживания и вылупления составляет от 45 до 65 дней. [ 21 ]
Варриккио и др. (2008) исследовали гистологию кости образца троодонта Two Medicine MOR 748 и обнаружили, что у него отсутствуют закономерности резорбции кости, которые указывали бы на то, что это была самка, откладывающая яйца. Они также измерили соотношение общего объема яиц в кладках к массе тела взрослой особи. Они изобразили корреляцию между этим соотношением и типом родительской стратегии, используемой современными птицами и крокодилами, и обнаружили, что это соотношение у троодонтов соответствует такому же соотношению у птиц, у которых яйца высиживает только взрослый самец. Из этого они пришли к выводу, что самки троодонтов, скорее всего, не высиживают яйца, в отличие от самцов, и это может быть общим признаком между динозаврами-манирапторами и базальными птицами. [ 22 ] Однако более поздний анализ массы птичьей кладки показал, что тип родительской заботы не может быть определен с использованием традиционных аллометрических методов, таких как тот, который использовался Варриккио и др. [ 23 ]
Палеоэкология
[ редактировать ]
Stenonychosaurus inequalis известен из формации Парка динозавров на юге Альберты , Канада, которая в то время представляла собой теплую прибрежную пойму, покрытую лесами умеренного пояса. Высшими хищниками были тираннозавриды , такие как дасплетозавр и горгозавр . Травоядные животные включали гадрозавридов , таких как ламбеозавр , коритозавр и прозауролоф ; цератопсиды , такие как стиракозавр , центрозавр и хасмозавр ; анкилозавры , такие как Scolosaurus , Euoplocephalus и Edmontonia ; и пахицефалозавры, такие как Stegoceras и Foraminacephale . [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Рассел, Д.А.; Сеген, Р. (1982). «Реконструкция небольшого мелового теропода Stenonychosaurus inequalis и гипотетического динозавроида» . Силлогей . 37 : 1–43.
- ^ Карри, П. (1987). «Тероподы формации реки Джудит». Периодическая статья Палеонтологического музея Тиррелла . 3 : 52–60.
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 398–399 . ISBN 978-0-671-61946-6 .
- ^ Карри, П. (2005). «Тероподы, включая птиц». в Карри и Коппельхус (ред.). Провинциальный парк динозавров, впечатляющая экосистема. Часть вторая. Флора и фауна парка. Издательство Университета Индианы, Блумингтон. Стр. 367–397.
- ^ Занно, Линдси Э.; Варриккио, Дэвид Дж.; О'Коннор, Патрик М.; Титус, Алан Л.; Нелл, Майкл Дж. (2011). Лалуэса-Фокс, Карлес (ред.). «Новый теропод-троодонтид, Talos sampsoni gen. et sp. nov., из верхнемелового западного внутреннего бассейна Северной Америки» . ПЛОС ОДИН . 6 (9): e24487. Бибкод : 2011PLoSO...624487Z . дои : 10.1371/journal.pone.0024487 . ПМК 3176273 . ПМИД 21949721 .
- ^ Эванс, округ Колумбия; Каллен, ТМ; Ларсон, Д.В.; Рего, А. (2017). «Новый вид троодонтидного теропода (Dinosauria: Maniraptora) из формации Каньон Подкова (Маастрихт) в Альберте, Канада» . Канадский журнал наук о Земле . 54 (8): 813–826. Бибкод : 2017CaJES..54..813E . doi : 10.1139/cjes-2017-0034 .
- ^ Jump up to: а б ван дер Реест, AJ; Карри, Пи Джей (2017). «Троодонтиды (Theropoda) из формации Парк динозавров, Альберта, с описанием уникального нового таксона: значение для разнообразия дейнонихозавров в Северной Америке». Канадский журнал наук о Земле . 54 (9): 919–935. Бибкод : 2017CaJES..54..919V . doi : 10.1139/cjes-2017-0031 . hdl : 1807/78296 .
- ^ Каллен, Томас М.; Занно, Линдси; Ларсон, Дерек В.; Тодд, Эринн; Карри, Филип Дж.; Эванс, Дэвид К. (30 июня 2021 г.). «Анатомический, морфометрический и стратиграфический анализ биоразнообразия теропод в формации парка динозавров верхнего мела (кампана)» . Канадский журнал наук о Земле . 58 (9): 870–884. doi : 10.1139/cjes-2020-0145 .
- ^ Jump up to: а б Нэйш, Д. (2006). Возвращение к динозавроидам Даррена Нэйша: Зоология четвероногих, 23 апреля 2011 г.
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам, 2-е издание . Издательство Принстонского университета. п. 160. ИСБН 978-0-691-16766-4 .
- ^ Тернер, Алан Х.; Марк А. Норелл; Диего Пол; Джулия А. Кларк; Грегори М. Эриксон (2007). «Базальный дромеозаврид и эволюция размеров перед полетом птиц» . Научный журнал . 317 (5843): 1378–81. Бибкод : 2007Sci...317.1378T . дои : 10.1126/science.1144066 . ПМИД 17823350 .
- ^ Карри, Пи Джей (1987). «Птичьи характеристики челюстей и зубов троодонтидных теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных . 7 : 72–81. дои : 10.1080/02724634.1987.10011638 .
- ^ Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон, Великобритания: Marshall Editions. стр. 112–113. ISBN 978-1-84028-152-1 .
- ^ Jump up to: а б Ларссон, HCE (2001). «Эндокраниальная анатомия Carcharodontosaurus saharicus (Theropoda: Allosauroidea) и ее значение для эволюции мозга теропод». В Танке, DH; Карпентер, К.; Скрепник, М.В. (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. стр. 19–33.
- ^ Хольц, Томас Р., Бринкман, Дэниел Л., Чендлер, Чистин Л. (1998) Морфометрия зубчиков и, возможно, всеядная привычка питания теропод-динозавра Троодона. Гайя, номер 15. Декабрь 1998 г., стр. 159–166.
- ^ Варриккио, диджей (1993). Микроструктура костей верхнемелового тероподного динозавра Troodon formosus. Дж. Вертебр. Палеонтология. 13 , 99–1 JSTOR 4523488
- ^ Якобсен, А.Р. 2001. Маленькая кость теропода с отмеченными зубами: чрезвычайно редкий след. п. 58-63. В: Жизнь мезозойских позвоночных . Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Университета Индианы.
- ^ Хорнер, Джон Р. (1984) «Гнездовое поведение динозавров». «Научный американец», 250: 130–137.
- ^ Хорнер, Джон Р., Вейшампель, Дэвид Б. (1996) «Сравнительное эмбриологическое исследование двух птицетазовых динозавров - исправление». «Природа» 383: 256–257.
- ^ Варриккио, диджей; Джексон, Ф.; Борковски, Джей Джей; Хорнер, младший (1997). «Кладки гнезд и яиц динозавра Troodon formosus и эволюция репродуктивных признаков птиц». Природа . 385 (6613): 247–250. Бибкод : 1997Natur.385..247V . дои : 10.1038/385247a0 . S2CID 4313286 .
- ^ Варриккио, Дэвид Дж.; Хорнер, Джон Дж.; Джексон, Фрэнки Д. (2002). «Эмбрионы и яйца мелового теропода динозавра Troodon formosus ». Журнал палеонтологии позвоночных . 22 (3): 564–576. doi : 10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2 . S2CID 85728452 .
- ^ Варриккио, диджей; Мур, младший; Эриксон, генеральный директор; Норелл, Массачусетс; Джексон, Флорида; Борковски, Джей Джей (2008). «Птичья отцовская забота имела происхождение от динозавров» . Наука 322 (5909): 1826–8. Бибкод : 2008Науч... 322.1826В дои : 10.1126/science.1163245 . ПМИД 19095938 .
- ^ Бирчард Джеффри Ф.; Рута Марчелло; Диминг Д. Чарльз (23 августа 2013 г.). «Эволюция инкубационного поведения родителей у динозавров не может быть выведена из массы кладки у птиц» . Письма по биологии . 9 (4): 20130036. doi : 10.1098/rsbl.2013.0036 . ПМЦ 3730617 . ПМИД 23676654 .
