Пахицефалозаврия
| Пахицефалозавры Временной диапазон:
| |
|---|---|

| |
| Монтаж четырех пахицефалозавров. По часовой стрелке сверху слева: Stegoceras , Prenocephale , Pachycephalosaurus и Homalocephale. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Пиродонтия |
| Клэйд : | † Церапода |
| Клэйд : | † Маргиноцефалия |
| Клэйд : | † Пахицефалозаврия Штернберг , 1945 год. ( сохранившееся название ) |
| Типовой вид | |
| † Пахицефалозавр вайомингенсис Гилмор , 1931 г. (сохранилось название)
| |
| Роды | |
| |
Пахицефалозаврия ( / ˌ p æ k ɪ s ɛ f əl ə ˈ s ɔː r i ə , - ˌ k ɛ f -/ ; [2] от греческого «толстоголовые ящерицы») — клада птицетазовых παχυκεφαλόσαυρος — динозавров . Вместе с Ceratopsia составляет кладу Marginocephalia . За исключением двух видов, большинство пахицефалозавров жили в позднемеловом периоде , примерно между 85,8 и 66 миллионами лет назад. [3] Они обитают исключительно в Северном полушарии , все они обитают в Северной Америке и Азии. Все они были двуногими , травоядными / всеядными животными с толстыми черепами. Черепа могут быть куполообразными, плоскими или клиновидными в зависимости от вида, и все они сильно окостеневшие. Купола часто были окружены узлами и/или шипами. Частичные скелеты были обнаружены у нескольких видов пахицефалозавров, но на сегодняшний день полных скелетов обнаружено не было. Часто отдельные фрагменты черепа являются единственными найденными костями. [4]
Кандидатами на звание самого раннего из известных пахицефалозавров являются Ferganocephale adenticulatum из среднеюрского периода слоев Киргизии и Stenopelix valdensis из раннего мела слоев в Германии , хотя Р.М. Салливан сомневался, что какой-либо из этих видов является пахицефалозавром. [5] Альбалофозавр из раннемеловых отложений Японии также может представлять собой базального пахицефалозавра. [1] Самый старый известный окончательный пахицефалозавр — bexelli из позднего мела Китая Sinocephale . [6] В 2017 году филогенетический анализ, проведенный Ханом и его коллегами, идентифицировал Стенопеликса как представителя Ceratopsia . [7]
Описание
[ редактировать ]

Пахицефалозавры были двуногими птицетазами, характеризующимися утолщенными черепами. У них было массивное туловище с расширенной брюшной полостью и широкими бедрами, короткие передние конечности, длинные ноги, короткая толстая шея и тяжелый хвост. Большие глазницы и большой зрительный нерв указывают на то, что у пахицефалозавров было хорошее зрение, а нехарактерно большие обонятельные доли указывают на то, что у них было хорошее обоняние по сравнению с другими динозаврами. [4] Это были довольно маленькие динозавры, длина большинства из которых составляла 2–3 метра (6,6–9,8 футов), а самый крупный из них, Pachycephalosaurus wyomingensis , имел длину 4,5 метра (14,8 футов) и весил 450 кг (990 фунтов). . [3] [8] Характерный череп пахицефалозавров является результатом слияния и утолщения лобных теменных и костей , сопровождающегося закрытием надвисочного окна . У некоторых видов он имеет форму приподнятого купола; у других череп плоский или клиновидный. Хотя плоскоголовые пахицефалозавры традиционно считаются отдельными видами или даже семействами, они могут представлять собой молодых особей куполообразной особи. [5] У всех имеются сильно орнаментированные скуловые кости , чешуйчатые кости и заглазничные кости в виде тупых рогов и узлов. Многие виды известны только по фрагментам черепа, а полный скелет пахицефалозавра еще не найден. [4]
Представители Pachycephalosauria характерно имеют необычно куполообразную голову, напоминающую более раннюю Protopyknosia в примере конвергентной эволюции . [9]
Классификация
[ редактировать ]
Большинство остатков пахицефалозаврид не являются полными и обычно состоят из частей лобно-теменной кости, образующей характерный купол. Это может затруднить таксономическую идентификацию, поскольку классификация родов и видов Pachycephalosauria почти полностью опирается на черепные характеристики. Следовательно, исторически к кладе относили неправильные виды. Например, Majungatholus , когда-то считавшийся пахицефалозавром, теперь признан экземпляром абелизавридного теропода Majungasaurus , а Yaverlandia , еще один динозавр, первоначально описанный как пахицефалозаврид, также с тех пор был реклассифицирован как целурозавр (Naish in Sullivan (2006) ). Еще больше усложняют ситуацию различные интерпретации онтогенетических и половых особенностей пахицефалозавров.
В статье 2009 года было высказано предположение, что Dracorex и Stygimoloch были всего лишь ранними стадиями роста пахицефалозавра , а не отдельными родами. [10]
Переработка Cerapoda 2020 года от Dieudonné et al. обнаружили животных, традиционно считавшихся « гетеродонтозавридами » как базальную группу внутри Pachycephalosauria, парафилетическую по отношению к традиционным куполообразным пахицефалозаврам. [11] К такому же выводу ранее пришел Джордж Ольшевский в 1991 году, который классифицировал гетеродонтозаврид как базальных пахицефалозавров на основании предполагаемого краниального кинеза , наличия клыковидных предчелюстных зубов и выраженной диастемы , присутствующей у многих родов. [12]
Таксономия
[ редактировать ]Pachycephalosauria была впервые названа как подотряд отряда Ornithischia Марьянской и Осмольской (1974) . Они включили в него только одно семейство — Pachycephalosauridae. [13] Более поздние исследователи, такие как Майкл Бентон, причислили его к инфраотряду подотряда Cerapoda , объединяющего цератопсов и орнитопод . [14] В 2006 году Роберт Салливан опубликовал повторную оценку таксономии пахицефалозавров. Салливан считал попытки Марьянской и Осмольской ограничить определение Pachycephalosauria излишними с их Pachycephalosauridae, поскольку они были диагностированы по одним и тем же анатомическим признакам. Салливан также отверг попытки Серено (1986) в его филогенетических исследованиях: [15] переопределить Pachycephalosauridae, включив в него только виды с «куполообразным черепом» (включая Stegoceras и Pachycephalosaurus ), оставив при этом больше «базальных» видов за пределами этого семейства в Pachycephalosauria. Таким образом, использование Салливаном Pachycephalosauridae эквивалентно использованию Серено и Бентоном Pachycephalosauria.
Салливан диагностировал Pachycephalosauridae только на основании признаков черепа, причем определяющим признаком была куполообразная лобно-теменная кость черепа. По мнению Салливана, отсутствие этой особенности у некоторых видов, считавшихся примитивными, привело к разделению классификации на куполообразных и некупольных пахицефалозавров; однако открытие более продвинутых и, возможно, молодых пахицефалозавров с плоскими черепами (таких как Dracorex hogwartsia ) показывает, что это различие неверно. Салливан также отметил, что первоначальный диагноз Pachycephalosauridae основывался на черепах от «плоских до куполообразных», поэтому формы с плоской головой следует включить в это семейство. [16] В статье, опубликованной в 2003 году, Томас Э. Уильямсон и Томас Д. Карр обнаружили кладу Pachycephalosauridae, которая была сестринским таксоном рода Stegoceras , состоящим из «всех других куполообразных пахицефалозавров; она называлась Pachycephalosaurinae». [17]
Филогения
[ редактировать ]Филогенетический анализ, проведенный многими авторами, показал, что Pachycephalosauria представляет собой группу, в которой Stegoceras является одним из самых ранних полностью куполообразных представителей, с плоскоголовыми и потенциально ювенильными таксонами, такими как Homalocephale и Goyocephale, либо сразу за пределами, либо внутри ее клады, а также более производными пахицефалозаврами. . Эти исследования начались с филогенетической работы Пола Серено , которая во многих итерациях была модифицирована для включения новых таксонов и дополнительных признаков. [18] [19] [20] Ниже представлена версия анализа, опубликованная Вудраффом и его коллегами в 2023 году. [21]
| Пахицефалозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ниже представлена кладограмма, опубликованная Дьедонне и его коллегами (2020 г.). [22] которые противоречиво обнаружили, что гетеродонтозавриды являются парафилетическими по отношению к пахицефалозаврам. Этот анализ был предложен в качестве гипотезы полного отсутствия окаменелостей пахицефалозавров юрского и раннего мела, хотя они должны были существовать, если современное понимание филогении птицетазовых верно. Однако эта гипотеза не получила широкого признания среди других палеонтологов. [1]
| Пахицефалозаврия |
| |||||||
Палеобиология
[ редактировать ]Кормление
[ редактировать ]Небольшой размер большинства видов пахицефалозавров и отсутствие скелетной адаптации указывают на то, что они не были альпинистами и в основном питались близко к земле. Мэллон и др. (2013) исследовали сосуществование травоядных животных на островном континенте Ларамидия в позднем меловом периоде и пришли к выводу, что пахицефалозавриды обычно ограничивались питанием растительностью на высоте 1 метр или ниже. [23] Они демонстрируют гетеродонтию , имея разную морфологию зубов между предчелюстными и верхнечелюстными зубами. Передние зубы маленькие, колышевидные, с овальным поперечным сечением и, скорее всего, использовались для хватания пищи. У некоторых видов последний предчелюстной зуб был увеличен и напоминал клык. Задние зубы маленькие, треугольной формы с зубчиками на передней и задней части коронки, служащими для обработки рта. У видов, у которых зубная кость обнаружена , зубы нижней челюсти по размеру и форме аналогичны зубам верхней челюсти. Характер износа зубов варьируется в зависимости от вида, что указывает на ряд пищевых предпочтений, которые могут включать семена, стебли, листья, фрукты и, возможно, насекомых. Очень широкая грудная клетка и большая кишечная полость, простирающаяся до основания хвоста, позволяют предположить, что для переваривания пищи используется ферментация. [4]
Бодание головой
[ редактировать ]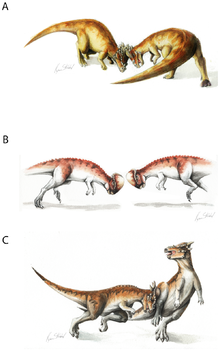
Адаптивное значение купола черепа активно обсуждается. Популярная среди широкой публики гипотеза о том, что череп использовался для бодания головой, как своего рода таран динозавров, была впервые предложена Кольбером (1955) , с. [ нужна страница ] . Эта точка зрения была популяризирована в научно-фантастическом рассказе « Пистолет для динозавров » Л. Спрэга де Кампа 1956 года . Многие палеонтологи с тех пор отстаивали гипотезу бодания головой, в том числе Гальтон (1970) и Сьюс (1978) . Согласно этой гипотезе, пахицефалозавры таранили друг друга в лоб, как это делают современные снежные бараны и овцебыки .
Анатомические свидетельства боевого поведения включают сочленения позвонков, обеспечивающие жесткость позвоночника, а также форму спины, указывающую на сильную мускулатуру шеи. [24] Было высказано предположение, что пахицефалозавр мог выпрямлять голову, шею и тело горизонтально, чтобы передавать напряжение во время тарана. Однако ни у одного известного динозавра голова, шея и тело не могли быть ориентированы в таком положении. Вместо этого шейные и передние спинные позвонки пахицефалозавров показывают, что шея имела S- или U-образную кривую. [25]
Кроме того, закругленная форма черепа уменьшила площадь соприкасающейся поверхности во время удара головой, что привело к скользящим ударам. Другие возможности включают удар с фланга, защиту от хищников или и то, и другое. Относительно широкое телосложение пахицефалозавров (которое защищало жизненно важные внутренние органы от повреждений во время удара с бока) и чешуйчатые рога стигимолоха ( которые могли бы с большим успехом использоваться во время удара с бока) добавляют убедительности гипотезе о боде с бока. [ нужна ссылка ]

Гистологическое исследование, проведенное Гудвином и Хорнером (2004), выступило против гипотезы тарана. Они утверждали, что купол был «эфемерной онтогенетической стадией», губчатая костная структура не могла выдержать удары боя, а радиальный рисунок был просто эффектом быстрого роста. [26] Однако более поздние биомеханические анализы, проведенные Снивели и Коксом (2008) и Снивели и Теодором (2011) , пришли к выводу, что купола могут выдерживать боевые нагрузки. [24] Леман (2010) утверждал, что модели роста, обсуждаемые Гудвином и Хорнером, не противоречат поведению «бодания головой». [27]
Вместо этого Гудвин и Хорнер (2004) утверждали, что купол служит для распознавания видов. Есть свидетельства того, что купол имел какое-то внешнее покрытие, и разумно предположить, что купол мог быть ярко покрыт или мог менять цвет в зависимости от сезона. [26] Однако из-за характера летописи окаменелостей невозможно проследить, играл ли цвет роль в функции купола.
Лонгрич, Санки и Танке (2010) утверждали, что распознавание видов является маловероятной эволюционной причиной возникновения купола, поскольку формы купола существенно не различаются между видами. Из-за этого общего сходства несколько родов Pachycephalosauridae иногда ошибочно объединяли. Это не похоже на случай с цератопсами и гадрозавридами , у которых черепной орнамент был гораздо более отчетливым. Лонгрич и др. утверждал, что вместо этого купол выполнял механическую функцию, например боевую, которая была достаточно важной, чтобы оправдать инвестиции в ресурсы. [19]
Купольная палеопатология
[ редактировать ]

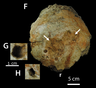
Петерсон, Дишлер и Лонгрич (2013) изучали краниальные патологии среди Pachycephalosauridae и обнаружили, что 22% всех исследованных куполов имели поражения, соответствующие остеомиелиту , инфекции кости, возникающей в результате проникающей травмы или травмы ткани, покрывающей череп, ведущей к к инфицированию костной ткани. Этот высокий уровень патологии еще больше подтверждает гипотезу о том, что купола пахицефалозаврид использовались во внутривидовых боях. [28] Частота травм была сопоставима у разных родов этого семейства , несмотря на то, что эти роды различаются размером и архитектурой своих куполов, а также тем фактом, что они существовали в разные геологические периоды. [28] Эти результаты резко контрастировали с результатами анализа относительно плоскоголовых пахицефалозаврид, у которых не было патологии. Это поддержало бы гипотезу о том, что эти особи представляют собой либо самок, либо несовершеннолетних. [19] где не предполагается внутривидовое боевое поведение.

Гистологическое исследование показывает, что купола пахицефалозаврид состоят из уникальной формы фиброламеллярной кости. [29] который содержит фибробласты , которые играют решающую роль в заживлении ран и способны быстро откладывать костную ткань во время ремоделирования. [10] Петерсон и др. (2013) пришли к выводу, что в совокупности частота распределения поражений и костная структура лобно-теменных куполов убедительно подтверждают гипотезу о том, что пахицефалозавриды использовали свои уникальные черепные структуры для агонистического поведения . [28]
Палеоэкология
[ редактировать ]
Азиатские и североамериканские виды пахицефалозавров жили в совершенно разных условиях. Азиатские экземпляры обычно более неповреждены, что указывает на то, что их не перевозили далеко от места смерти до окаменения. Вероятно, они жили в большом пустынном регионе Центральной Азии с жарким и засушливым климатом . Североамериканские экземпляры обычно встречаются в породах, образовавшихся в результате эрозии Скалистых гор . Образцы гораздо менее целы; обычно обнаруживаются только черепные коробки, а у найденных регулярно наблюдается отслоение поверхности и другие признаки того, что они были перенесены водой на большие расстояния до окаменения. Предполагается, что они жили в горах в умеренном климате и были перенесены эрозией после смерти к месту своего последнего упокоения. [30]
Распределение
[ редактировать ]Пахицефалозавры жили исключительно в Лавразии , их находили в западной части Северной Америки и Центральной Азии. Пахицефалозавры возникли в Азии и пережили два крупных события расселения, в результате которых в Азии наблюдались две отдельные волны эволюции пахицефалозавров. Первый, произошедший до позднего сантона или раннего кампана , включал миграцию из Азии в Северную Америку, скорее всего, через Берингов мост . благодаря общему предку Stygimoloch , Stegoceras , Tylocephale , Prenocephale и Эта миграция произошла Pachycephalosaurus . Второе событие произошло до среднего кампана и включало миграцию обратно в Азию из Северной Америки общего предка Prenocephale и Tylocephale . Два вида, первоначально заявленные как пахицефалозавры, обнаруженные за пределами этого ареала: Yaverlandia bitholus из Англии и Majungatholus atopus из Мадагаскара , недавно было показано, что на самом деле они являются тероподами . [4] [31]
См. также
[ редактировать ]- Хронология исследований пахицефалозавров
- Triopticus Конвергентная эволюция
Примечания
[ редактировать ]- ^ Jump up to: а б с д Фонсека, АО; Рид, Ай-Джей; Веннер, А.; Дункан, Р.Дж.; Гарсия, MS; Мюллер, RT (2024). «Всесторонний филогенетический анализ ранней птицетазовой эволюции». Журнал систематической палеонтологии . 22 (1). 2346577. дои : 10.1080/14772019.2024.2346577 .
- ^ «Пахицефалозаврия» . Оксфордские словари Британско-английского словаря . Издательство Оксфордского университета . [ мертвая ссылка ]
- ^ Jump up to: а б Хольц, Томас Р. младший (2007 г.) [последнее обновление 28 января 2011 г.]. «Список родов динозавров Хольца (2007)» (PDF) .
- ^ Jump up to: а б с д и Марьянская, Т.; Чепмен, RE; Вейшампель, Д.Б. (2004). «Пахицефалозаврия». Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 464–477. ISBN 978-0520242098 . OCLC 493366196 .
- ^ Jump up to: а б Салливан 2006 г. [ нужна проверка ]
- ^ Эванс, Дэвид; Браун, Калеб М.; Ты, Хайлу; Кампионе, Николас Э. (2021). «Описание и пересмотренный диагноз первого зарегистрированного пахицефалозавриды в Азии, Sinocephale bexelli gen. nov., из верхнего мела Внутренней Монголии, Китай» . Канадский журнал наук о Земле . е – Первый (10): 981–992. doi : 10.1139/cjes-2020-0190 . S2CID 244227050 .
- ^ Хан, Флорида; Форстер, Калифорния; Кларк, Дж. М.; Сюй, X. (2016). «Краниальная анатомия Иньлун Даунси (Ornithischia: Ceratopsia) из верхнеюрской формации Шишугоу в Синьцзяне, Китай». Журнал палеонтологии позвоночных . 36 (1): e1029579. дои : 10.1080/02724634.2015.1029579 . S2CID 130791053 .
- ^ Пол, Грегори С. (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. ISBN 9780691137209 . OCLC 930852339 . [ нужна страница ]
- ^ Стокер, MR; Несбитт, С.Дж.; Крисвелл, Кентукки; Паркер, В.Г.; Уитмер, Л.М.; Роу, ТБ; Риджли, Р.; Браун, Массачусетс (2016). «Куполаголовый стеблевой архозавр является примером конвергенции динозавров и их дальних родственников» . Современная биология . 26 (19): 2674–2680. дои : 10.1016/j.cub.2016.07.066 . ПМИД 27666971 .
- ^ Jump up to: а б Хорнер, младший ; Гудвин, МБ (2009). «Чрезвычайная краниальная онтогенез верхнемелового динозавра пахицефалозавра » . ПЛОС ОДИН . 4 (10): е7626. Бибкод : 2009PLoSO...4.7626H . дои : 10.1371/journal.pone.0007626 . ПМК 2762616 . ПМИД 19859556 .
- ^ Дьёдонне П., Крузадо-Кабальеро П., Годфруа П., Тортоса Т. (2020). «Новая филогения цераподских динозавров». Историческая биология . 33 (10): 2335–2355. дои : 10.1080/08912963.2020.1793979 .
- ^ Ольшевский, Георгий (1991). «Пересмотр параинфракласса Archosauria Cope, 1869 г., за исключением продвинутых крокодилов» (PDF) . Библиотека наук о Земле : 104.
- ^ Марианска Т , Осмольска Х (1974). «Пахицефалозаврия, новый подотряд птицетазовых динозавров». Польская палеонтология . 30 : 45–102.
- ^ Бентон, Майкл Дж. (2004). Палеонтология позвоночных (3-е изд.). Издательство Блэквелл. п. 472. ИСБН 978-0-632-05637-8 .
- ^ Серено, ПК (1986). «Филогения птицетазовых динозавров (отряд Ornithischia)». Национальное географическое исследование . 2 : 234–256.
- ^ Салливан Р.М. (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 347–365. Архивировано из оригинала (PDF) 27 сентября 2007 г.
- ^ Уильямсон, Томас Э.; Карр, Томас Д. (14 января 2003 г.). «Новый род производных пахицефалозавров из западной части Северной Америки» . Журнал палеонтологии позвоночных . 22 (4): 779–801. doi : 10.1671/0272-4634(2002)022[0779:ANGODP]2.0.CO;2 . ISSN 0272-4634 . S2CID 86112901 .
- ^ Уильямсон Т.Э., Карр Т.Д. (2002). «Новый род производных пахицефалозавров из западной части Северной Америки». Журнал палеонтологии позвоночных . 22 (4): 779–801. doi : 10.1671/0272-4634(2002)022[0779:ANGODP]2.0.CO;2 . ISSN 0272-4634 . S2CID 86112901 .
- ^ Jump up to: а б с Лонгрич, Северная Каролина; Санки, Дж.; Танке, Д. (2010). «Texacephale langstoni, новый род пахицефалозаврид (Dinosauria: Ornithischia) из верхней кампанской формации Агуджа, южный Техас, США». Меловые исследования . 31 (2): 274–284. дои : 10.1016/j.cretres.2009.12.002 .
- ^ Эванс, округ Колумбия, Шотт Р.К., Ларсон Д.В., Браун СМ, Райан М.Дж. (2013). «Старейший североамериканский пахицефалозавр и скрытое разнообразие мелких птицетазовых динозавров» . Природные коммуникации . 4 . 1828. Бибкод : 2013NatCo...4.1828E . дои : 10.1038/ncomms2749 . ПМИД 23652016 .
- ^ Вудрафф, Д. Кэри; Шотт, Райан К.; Эванс, Дэвид К. (15 ноября 2023 г.). «Два новых вида пахицефалозаврин с мелким телом (Dinosauria, Marginocephalia) из верхнего мела Северной Америки предполагают скрытое разнообразие в хорошо отобранных образованиях». Статьи по палеонтологии . 9 (6). е1535. дои : 10.1002/spp2.1535 . ISSN 2056-2799 .
- ^ Дьедонне, П.-Э.; Крузадо-Кабальеро, П.; Годфруа, П.; Тортоса, Т. (2021). «Новая филогения цераподских динозавров» . Историческая биология . 33 (10): 2335–2355. дои : 10.1080/08912963.2020.1793979 . S2CID 221854017 .
- ^ Мэллон, Джордан К.; Эванс, Дэвид С.; Райан, Майкл Дж.; Андерсон, Джейсон С. (2013). «Стратификация по высоте питания травоядных динозавров из формации Парк динозавров (верхний кампан) Альберты, Канада» . БМК Экология . 13:14 . дои : 10.1186/1472-6785-13-14 . ПМЦ 3637170 . ПМИД 23557203 .
- ^ Jump up to: а б Снавли Э., Кокс А. (март 2008 г.). «Структурная механика черепов пахицефалозавра допускала бодание головой» . Электронная палеонтология . 11 (1). ISSN 1094-8074 .
- ^ Карпентер, К. (1997). «Агонистическое поведение пахицефалозавров (Ornithischia:Dinosauria): новый взгляд на поведение с боданием головой» (PDF) . Вклад в геологию . 32 (1): 19–25. Архивировано из оригинала (PDF) 3 марта 2016 г. Проверено 16 июля 2011 г.
- ^ Jump up to: а б Гудвин М.Б., Хорнер-младший (июнь 2004 г.). «Краниальная гистология пахицефалозавров (Ornithischia: Marginocephalia) выявляет временные структуры, несовместимые с поведением бодания головой» (PDF) . Палеобиология . 30 (2): 253–267. doi : 10.1666/0094-8373(2004)030<0253:CHOPOM>2.0.CO;2 . S2CID 84961066 .
- ^ Леман ТМ (2010). «Pachycephalosauridae из формаций Сан-Карлос и Агуджа (верхний мел) западного Техаса и наблюдения за лобно-теменным куполом». Журнал палеонтологии позвоночных . 30 (3): 786–798. дои : 10.1080/02724631003763532 . ISSN 0272-4634 . S2CID 129592428 .
- ^ Jump up to: а б с Петерсон Дж. Э., Дишлер С., Лонгрич Н. Р. (2013). «Распределение черепных патологий доказывает, что у куполообразноголовых динозавров (Pachycephalosauridae) бьются головой» . ПЛОС ОДИН . 8 (7): е68620. Бибкод : 2013PLoSO...868620P . дои : 10.1371/journal.pone.0068620 . ПМЦ 3712952 . ПМИД 23874691 .
- ^ Рид, REH (1997). «Гистология костей и зубов». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Сан-Диего, Калифорния: Academic Press. стр. 329–339.
- ^ Вейшампель, Дэвид Б. (2009). Динозавры: краткая естественная история . Издательство Кембриджского университета. ISBN 9780511479410 . OCLC 476234422 . [ нужна страница ]
- ^ Нэйш, Д .; Мартилл, DM (2008). «Динозавры Великобритании и роль Лондонского геологического общества в их открытии: Ornithischia». Журнал Геологического общества . 165 (3): 613–623. Бибкод : 2008JGSoc.165..613N . дои : 10.1144/0016-76492007-154 . S2CID 129624992 .
Ссылки
[ редактировать ]- Бентон, Майкл Дж (2004). Палеонтология позвоночных (3-е изд.). Издательство Блэквелл. ISBN 978-0-632-05637-8 .
- Карпентер, Кеннет (1997). «Агонистическое поведение пахицефалозавров (Ornithischia: Dinosauria): новый взгляд на поведение с боданием головой» (PDF) . Вклад в геологию . 32 (1): 19–25. Архивировано из оригинала (PDF) 3 марта 2016 г. Проверено 16 июля 2011 г.
- Кольбер, Эдвин (1955). Эволюция позвоночных . Нью-Йорк: Джон Уайли.
- Гальтон, П. (1970). «Пахицефалозавриды — динозавровые тараны». Открытие . 6 : 23–32.
- Гудвин, МБ; Хорнер, младший (июнь 2004 г.). «Краниальная гистология пахицефалозавров (Ornithischia: Marginocephalia) выявляет временные структуры, несовместимые с поведением бодания головой» (PDF) . Палеобиология . 30 (2): 253–267. doi : 10.1666/0094-8373(2004)030<0253:CHOPOM>2.0.CO;2 . S2CID 84961066 .
- Хорнер, младший; Гудвин, МБ (2009). Серено, Пол (ред.). «Чрезвычайная краниальная онтогенез верхнемелового динозавра пахицефалозавра » . ПЛОС ОДИН . 4 (10): е7626. Бибкод : 2009PLoSO...4.7626H . дои : 10.1371/journal.pone.0007626 . ПМК 2762616 . ПМИД 19859556 .
- Леман, ТМ (2010). «Pachycephalosauridae из формаций Сан-Карлос и Агуджа (верхний мел) западного Техаса и наблюдения за лобно-теменным куполом». Журнал палеонтологии позвоночных . 30 (3): 786–798. дои : 10.1080/02724631003763532 . ISSN 0272-4634 . S2CID 129592428 .
- Лонгрич, Северная Каролина; Санки, Дж; Танке, Д. (2010). « Texacephale langstoni , новый род пахицефалозаврид (Dinosauria: Ornithischia) из верхней кампанской формации Агуджа, южный Техас, США». Меловые исследования . 31 (2): 274–284. дои : 10.1016/j.cretres.2009.12.002 .
- Марьянская, Т ; Осмольска, Х (1974). «Пахицефалозаврия, новый подотряд птицетазовых динозавров». Палеонтологика Полоника (30): 45–102.
- Серено, ПК (1986). «Филогения птицетазовых динозавров (отряд Ornithischia)». Национальное географическое исследование . 2 : 234–256.
- Снайвли, Э; Кокс, А. (март 2008 г.). «Структурная механика черепов пахицефалозавра допускала бодание головой» . Электронная палеонтология . 11 (1). ISSN 1094-8074 .
- Снайвли, Э; Теодор, Дж. М. (2011). Карпентер, Кеннет (ред.). «Общие функциональные корреляты поведения при ударе головой у пахицефалозавра Stegoceras validum (Ornithischia, Dinosauria) и боевых парнокопытных» . ПЛОС ОДИН . 6 (6): e21422. Бибкод : 2011PLoSO...621422S . дои : 10.1371/journal.pone.0021422 . ПМК 3125168 . ПМИД 21738658 .
- Стокстад, Эрик (ноябрь 2007 г.). «Собрание Общества палеонтологии позвоночных: вызвали ли возбужденные молодые динозавры иллюзию существования отдельных видов?» . Наука . 318 (5854): 1236. doi : 10.1126/science.318.5854.1236 . ПМИД 18033861 . S2CID 36443204 .
- Суэц, HD (1978). «Функциональная морфология купола динозавров-пахицефалозаврид». Новогодний альбом по геологии и палеонтологии : 459–472.
- Салливан, РМ (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико (35): 347–365. Архивировано из оригинала (PDF) 27 сентября 2007 г.
- Ватабе, Махито; Цогтбаатар, Хишигджау; Салливан, Роберт М. (2011). «Новый пахицефалозавр из формации Байншир (сеноман-поздний сантон), пустыня Гоби, Монголия» (PDF) . Ископаемая запись 3. Музей естественной истории и науки Нью-Мексико, Бюллетень . 53 : 489–497. Архивировано из оригинала (PDF) 29 сентября 2011 г.
- Уильямсон, TE; Карр, Т.Д. (2002). «Новый род производных пахицефалозавров из западной части Северной Америки». Журнал палеонтологии позвоночных . 22 (4): 779–801. doi : 10.1671/0272-4634(2002)022[0779:ANGODP]2.0.CO;2 . ISSN 0272-4634 . S2CID 86112901 .
Внешние ссылки
[ редактировать ]- TEDx YouTube на на Выступление Джека Хорнера об меняющих форму черепах динозавров и неправильной классификации динозавров
