Стегоцерас
| Стегоцерас Временной диапазон: поздний мел ,
~ | |
|---|---|

| |
| Два реконструированных скелета S. validum на основе экземпляра UALVP 2, Королевский музей Тиррелла. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Пахицефалозаврия |
| Семья: | † Пахицефалозавриды |
| Род: | † Стегоцерас Ламбе , 1902 год. |
| Типовой вид | |
| † Стегоцерас сильный Ламбе, 1902 год.
| |
| Разновидность | |
| |
| Синонимы | |
|
Список | |
Stegoceras — это род пахицефалозавридных , (купологоловых) динозавров которые жили на территории современной Северной Америки в позднемеловой период , около 77,5–74 миллионов лет назад (млн лет назад). Первые экземпляры из Альберты , Канада, были описаны в 1902 году, и типовой вид Stegoceras validum на основе этих останков был основан . Родовое название означает «роговая крыша», а видовое – «крепкий». На протяжении многих лет к этому роду было отнесено несколько других видов, но с тех пор они были перенесены в другие роды или признаны младшими синонимами . В настоящее время сохранились только S. validum и S. nomexicanum , названные в 2011 году по окаменелостям, найденным в Нью-Мексико . Достоверность последнего вида также обсуждалась.
Стегоцерас был небольшим двуногим динозавром длиной от 2 до 2,5 метров (от 6,6 до 8,2 футов) и весом от 10 до 40 кг (от 22 до 88 фунтов). Череп имел примерно треугольную форму с короткой мордой и толстым, широким и относительно гладким куполом наверху. Задняя часть черепа имела толстый «полок» над затылком и толстый гребень над глазами. Большая часть черепа была украшена бугорками (или круглыми «выростами») и узлами (или «шишечками»), многие из которых располагались рядами, а самые крупные образовывали небольшие рога на полке. Зубы были маленькими и зазубренными. Считается, что у молодых животных череп был плоским, а с возрастом превратился в купол. У него был жёсткий позвоночник и жёсткий хвост. Тазовая область была широкой, возможно, из-за расширенного кишечника.
Первоначально известный только по куполам черепов, Stegoceras был одним из первых известных пахицефалозавров, и неполнота этих первоначальных останков привела к появлению множества теорий о родстве этой группы. Полный череп Stegoceras с соответствующими частями скелета был описан в 1924 году, что пролило больше света на этих животных. Пахицефалозавры сегодня объединены с рогатыми цератопсами в группу Marginocephalia . Сам Stegoceras считался базальным (или «примитивным») по сравнению с другими пахицефалозаврами. Стегоцерас , скорее всего, был травоядным и, вероятно, обладал хорошим обонянием. Функция купола обсуждается, и конкурирующие теории включают использование во внутривидовом бою (бодание головой или флангом), сексуальную демонстрацию или распознавание вида . S. validum известен из формации Парк динозавров и формации Олдман , тогда как S. nomexicanum происходит из формации Фрутленд и Киртланд .
История открытия
[ редактировать ]
Первые известные останки Stegoceras были собраны канадским палеонтологом Лоуренсом Ламбе из группы реки Белли , в районе реки Ред-Дир в Альберте, Канада . Эти останки состояли из двух частичных куполов черепа (образцы CMN 515 и CMN 1423 в Канадском музее природы ) двух животных разного размера, собранных в 1898 году, и третьего частичного купола (CMN 1594), собранного в 1901 году. На основе этих образцов Ламбе описал и назвал новый монотипический род и вид Stegoceras validus в 1902 году . [ 1 ] [ 2 ] Родовое название Stegoceras происходит от греческого stegè /στέγη, что означает «крыша», и keras /κέρας, что означает «рог». Видовое название validus на латыни означает «сильный», возможно, в связи с толстой крышей черепа. [ 3 ] Поскольку вид был основан на нескольких экземплярах ( серия синтипов в 1907 году определил CMN 515 как образец лектотипа ), Джон Белл Хэтчер . [ 4 ] [ 2 ]
Поскольку ранее в этом районе не было обнаружено подобных останков, Ламбе не был уверен, что это были за динозавры и представляли ли они один вид или несколько; он предположил, что купола представляют собой «преназальные кости», расположенные перед носовыми костями на средней линии головы, и отметил их сходство с сердцевиной носового рога экземпляра трицератопса . [ 1 ] В 1903 году венгерский палеонтолог Франц Нопча фон Фельсо-Силвас предположил, что фрагментарные купола Stegoceras на самом деле представляли собой лобные и носовые кости и, следовательно, у животного был один непарный рог. Ламбе с пониманием отнесся к этой идее нового типа «динозавра-единорога» в обзоре статьи Нопски в 1903 году. В то время все еще существовала неопределенность относительно того, Stegoceras к какой группе динозавров принадлежал как цератопсы (рогатые динозавры), так и стегозавры (пластинчатые динозавры). , причем претендентами были [ 5 ] [ 6 ] Хэтчер усомнился в принадлежности экземпляров Stegoceras к одному и тому же виду и были ли они вообще динозаврами, и предположил, что купола состоят из лобной, затылочной и теменной костей черепа. [ 2 ] В 1918 году Ламбе отнес еще один купол (CMN 138) к S. validus и назвал новый вид S. brevis на основе экземпляра CMN 1423 (который он первоначально включил в S. validus ). К этому времени он считал этих животных представителями Stegosauria (тогда состоявшего из обоих семейств панцирных динозавров, Stegosauridae и Ankylosauridae ), в новом семействе, которое он назвал Psalisauridae (названном в честь сводчатой или куполообразной крыши черепа). [ 7 ]


В 1924 году американский палеонтолог Чарльз В. Гилмор описал полный череп S. validus с соответствующими посткраниальными остатками, которые на тот момент были наиболее полными останками куполообразного динозавра. Он был обнаружен в группе реки Белли американским палеонтологом Джорджем Ф. Штернбергом в 1926 году и внесен в каталог как образец UALVP 2 в Университета Альберты Лаборатории палеонтологии позвоночных . Эта находка подтвердила интерпретацию Хэтчером куполов как состоящих из лобно-теменной области черепа. UALVP 2 был обнаружен с небольшими разрозненными костными элементами, которые тогда считались гастралиями (брюшными ребрами), которые не известны у других птицетазовых динозавров (одна из двух основных групп динозавров). Гилмор отметил, что зубы S. validus очень похожи на зубы вида Troodon formosus (названного в 1856 году и к тому времени известного только по отдельным зубам), и описал купол черепа, обнаруженный недалеко от места, где был найден троодон . Поэтому Гилмор считал Stegoceras недействительный младший синоним Троодона — , тем самым переименовав S. validus в T. validus , и предположил, что даже эти два вида могут быть одинаковыми. Более того, он обнаружил, что S. brevis идентичен S. validus и, следовательно, является младшим синонимом последнего. Он также поместил эти виды в новое семейство Troodontidae (поскольку Ламбе не выбрал типовой род для своих Psalisauridae), которое он считал наиболее близким к динозаврам -орнитоподам . [ 8 ] [ 9 ] Поскольку череп казался настолько специализированным по сравнению с довольно «примитивным» на вид скелетом, Нопча усомнился в том, что эти части на самом деле принадлежали друг другу, и предположил, что череп принадлежал нодозавру , скелет — орнитоподу, а предполагаемые гастралии (брюшные ребра) — нодозавру. рыба. Это утверждение было опровергнуто Гилмором и Лорисом С. Расселом в 1930-х годах. [ 10 ]
Классификация Гилмора была поддержана американскими палеонтологами Барнумом Брауном и Эрихом Мареном Шлайкьером в их обзоре купологоловых динозавров, известных к тому времени по 46 черепам, в 1943 году. Из этих экземпляров Браун и Шлайкьер назвали новые виды T. sternbergi и T. edmontonensis (оба из Альберты), а также переместили крупный вид T. wyomingensis (который был назван в 1931 году) в новый род Pachycephalosaurus вместе с двумя другие виды. Они обнаружили, что T. validus отличается от T. formosus , но считали S. brevis женской формой T. validus и, следовательно, младшим синонимом. К этому времени куполообразные динозавры считались родственниками либо орнитопод, либо анкилозавров. [ 10 ] В 1945 году, исследовав слепки зубов T. formosus и S. validus , американский палеонтолог Чарльз М. Штернберг продемонстрировал различия между ними и вместо этого предположил, что троодон был динозавром -тероподом , а купологоловых динозавров следует поместить в их собственная семья. Хотя Stegoceras был первым представителем этого семейства, получившим название, Штернберг назвал группу Pachycephalosauridae в честь второго рода, поскольку он нашел это название (означающее «толстоголовая ящерица») более описательным. Он также считал T. sternbergi и T. edmontonensis членами Stegoceras , нашел S. brevis действительным и назвал новый вид S. lampei на основе экземпляра, ранее отнесенного к S. validus . [ 3 ] [ 11 ] Отделение от Троодона было поддержано Расселом в 1948 году, который описал зубную кость теропода с зубами, почти идентичными зубам T. formosus . [ 12 ]



В 1953 году Биргер Болин назвал Troodon bexelli на основе теменной кости из Китая. [ 13 ] В 1964 году Оскар Кун считал это однозначным видом Stegoceras ; С. Бекселли . [ 14 ] В 1974 году польские палеонтологи Тереза Марьянска и Хальшка Осмолска пришли к выводу, что «гастралии» Stegoceras представляют собой окостеневшие сухожилия, после идентификации таких структур в хвосте пахицефалозавра Homalocephale . [ 9 ] В 1979 году Уильям Патрик Уолл и Питер Гальтон назвали новый вид Stegoceras Browni на основе уплощенного купола, ранее описанного как самка S. validus Гальтоном в 1971 году. Видовое название дано в честь Барнума Брауна, который нашел образец голотипа (образец AMNH). 5450 в Американском музее естественной истории ) в Альберте. [ 15 ] В 1983 году Гальтон и Ханс-Дитер Сьюс выделили S. Browni в отдельный род Ornatotholus ( ornatus в переводе с латыни означает «украшенный», а tholus — «купол») и считали его первым известным американским представителем группы «плоских» растений. «головоголовые» пахицефалозавры, ранее известные из Азии. [ 16 ] В обзоре пахицефалозавров 1987 года Сьюс и Гальтон изменили видовое название validus на validum , которое впоследствии использовалось в научной литературе. Эти авторы синонимизировали S. brevis , S. sternbergi и S. lampei с S. validum , обнаружили, что S. bexelli отличается от Stegoceras по нескольким признакам, и считали его неопределенным пахицефалозавром. [ 3 ] [ 4 ] В 1998 году Гудвин и его коллеги считали Ornatotholus ювенильным S. validum , следовательно, младшим синонимом. [ 17 ]
События 21 века
[ редактировать ]
В 2000 году Роберт М. Салливан отнес S. edmontonensis и S. brevis к роду Prenocephale (до этого известного только из монгольского вида P. prenes ) и обнаружил, что более вероятно, что S. bexelli принадлежал к Prenocephale, чем к Stegoceras , но счел его nomen dubium (сомнительное имя без отличительных знаков) из-за его неполноты и отметил, что его голотипный образец, по-видимому, потеряться. [ 18 ] В 2003 году Томас Э. Уильямсон и Томас Карр посчитали Орнатотолус или сомнительным названием , возможно, молодым Stegoceras . [ 19 ] В пересмотре Stegoceras в 2003 году Салливан согласился, что Ornatotholus был младшим синонимом Stegoceras , переместил S. lampei в новый род Colepiocephale , а S. sternbergi в Hanssuesia . Он заявил, что до этого момента род Stegoceras стал таксоном мусорной корзины для мелких и средних североамериканских пахицефалозавров. были отнесены десятки экземпляров К этому времени к S. validum , в том числе многие купола, слишком неполные, чтобы их можно было с уверенностью идентифицировать как Stegoceras . UALVP 2 по-прежнему является наиболее полным экземпляром Stegoceras , на котором основано большинство научных представлений об этом роде. [ 4 ] S. brevis был перенесен в новый род Foraminacephale в 2016 году Райаном К. Шоттом Шоттом и Дэвидом К. Эвансом. [ 20 ] и S. bexelli в Sinocephale в 2021 году Эвансом и его коллегами. [ 21 ] В 2023 году Аарон Д. Дайер и его коллеги проанализировали швы и отдельные элементы черепов пахицефалозавров Gravitholus и Hanssuesia и не обнаружили существенных различий между ними и Stegoceras validum . Они считали оба младшими синонимами, причем Gravitholus представлял конечную стадию роста Stegoceras . [ 22 ]
В 2002 году Уильямсон и Карр описали купол (образец NMMNH P-33983 в Музее естественной истории и науки Нью-Мексико ) из бассейна Сан-Хуан , штат Нью-Мексико , которого они считали молодым пахицефалозавром неопределенного вида (хотя, возможно, Sphaerotholus Goodwini ). . В 2006 году Салливан и Спенсер Г. Лукас считали его молодым S. validum , что значительно расширит ареал этого вида. [ 23 ] [ 24 ] В 2011 году Стивен Э. Ясински и Салливан сочли этот экземпляр взрослым и сделали его голотипом нового вида Stegoceras nomomexicanum , а два других экземпляра (SMP VP-2555 и SMP VP-2790) — паратипами. [ 25 ] 2011 года Филогенетический анализ , проведенный Ватабе и его коллегами, не выявил близости двух видов Stegoceras друг к другу. [ 26 ]
В 2016 году Уильямсон и Стивен Л. Брусатте повторно изучили голотип S. noomexicanum и обнаружили, что паратипы не принадлежали к тому же таксону, что и голотип, и что все использованные экземпляры были молодыми. Кроме того, они не смогли определить, представляет ли образец голотипа отдельный вид S. noomexicanum , или это была молодь либо S. validum , либо Sphaerotholus Goodwini , либо другого ранее известного пахицефалозавра. [ 27 ] В 2016 году Ясински и Салливан защитили обоснованность S. noomexicanum ; они согласились, что некоторые признаки, используемые для диагностики этого вида, указывают на полувзрослую стадию, но представили дополнительные диагностические признаки голотипа, которые отличают этот вид. Они также указали на некоторые особенности взрослых особей, которые могут указывать на гетерохронию (разницу во времени онтогенетических изменений между родственными таксонами) у этого вида. Они признали, что паратипы и другие присвоенные экземпляры отличались от голотипа более куполообразными черепами, вместо этого называя их ср . S. noomexicanum (трудно идентифицировать), но обнаружил, что все они принадлежали к одному и тому же таксону (при этом назначенные экземпляры были взрослыми особями) из-за ограниченного стратиграфического интервала и географического ареала. [ 28 ] Дайер и его коллеги обнаружили, что голотип S. noomexicanum может быть незрелым Sphaerotholus и, следовательно, младшим синонимом в 2023 году. [ 22 ]
Описание
[ редактировать ]
Stegoceras — один из наиболее известных североамериканских пахицефалозавров и один из немногих, известных по посткраниальным останкам; Образец S. validum UALVP 2 — наиболее полный экземпляр Stegoceras, известный на сегодняшний день. По оценкам, его длина составляла от 2 до 2,5 метров (от 6,6 до 8,2 футов), что сравнимо с размером козы . [ 29 ] [ 30 ] [ 31 ] По оценкам, вес составляет от 10 до 40 килограммов (от 22 до 88 фунтов). [ 32 ] Стегоцерас был от маленького до среднего размера по сравнению с другими пахицефалозаврами. [ 3 ] S. noomexicanum , по-видимому, был меньше S. validum , но спорен вопрос о том, являются ли известные экземпляры (неполные черепа) взрослыми или молодыми особями. [ 25 ] [ 27 ]
Череп и зубной ряд
[ редактировать ]
Череп Stegoceras имел примерно треугольную форму, если смотреть сбоку, с относительно короткой мордой. Лобная и теменная кости были очень толстыми и образовывали приподнятый купол. Шов между этими двумя элементами был стерт (у некоторых экземпляров он был лишь слабо заметен), и их вместе называют «лобно-теменными». Лобно-теменной купол был широким, имел относительно гладкую поверхность, только бока были морщинистыми (морщинистыми). Она сужена сверху и между орбитами (глазницами). Лобно-теменная кость сужена сзади, вклинивалась между чешуйчатыми костями и заканчивалась впадиной над затылком в задней части черепа. Теменная и чешуйчатая кости образовывали толстый выступ над затылком, называемый теменно-чешуйчатым выступом, протяженность которого варьировалась у разных экземпляров. Чешуйчатая кость была большой, а не частью купола, а задняя часть была опухшей. Он был орнаментирован неравномерно расположенными бугорками (или круглыми выростами), а по его верхним краям простирался ряд узлов (шишечек), заканчивающихся заостренным бугорком (или небольшим рогом) с каждой стороны в задней части черепа. Внутренний ряд более мелких бугорков шел параллельно большему. За исключением верхней поверхности купола, большая часть черепа была украшена узлами, многие из которых располагались рядами. [ 3 ]
Большая орбита имела форму несовершенного эллипса (с самой длинной осью спереди назад), обращенной в сторону и немного вперед. Подвисочное окно (отверстие) позади глаза было узким и наклонено назад, а надвисочное окно в верхней части задней части черепа было сильно уменьшено в размерах из-за утолщения лобно-теменной области. Базикраний черепной (дно коробки ) был укорочен и отодвинут от областей ниже глазниц и вокруг неба . Затылок наклонен назад и вниз, а затылочный мыщелок отклонен в том же направлении. Слезная кость образовывала нижний передний край глазницы, поверхность ее имела ряды узловатого орнамента. Префронтальная глазная и кости срослись и образовали толстый гребень над глазницей. Относительно большая скуловая кость образовывала нижний край глазницы, простираясь далеко вперед и вниз по направлению к челюстному суставу. Он был украшен гребнями и узлами, расположенными радиально. [ 3 ]
Носовые отверстия были большими и обращены вперед. была Носовая кость толстой, рельефной и имела выпуклый профиль. Вместе с лобной костью он образовывал выступ (щит) на средней вершине черепа. Нижняя передняя часть предчелюстной кости (передняя кость верхней челюсти) была морщинистой и утолщенной. небольшое отверстие В шве между предчелюстными костями имелось (отверстие), ведущее в полость носа и, возможно, соединенное с органом Якобсона ( органом обоняния ). Верхняя челюсть была короткой и глубокой и, вероятно, содержала пазуху . На верхней челюсти был ряд отверстий, которые соответствовали положению каждого зуба и служили проходами для прорезывания сменных зубов. Нижняя челюсть сочленяется с черепом ниже задней части орбиты. Зубная часть нижней челюсти была длинной, а задняя часть довольно короткой. Хотя она и не сохранилась, на наличие предзубной кости указывают фасетки в передней части нижней челюсти. [ 3 ] Как и у других пахицефалозавров, у него был небольшой клюв. [ 33 ]

У Stegoceras были зубы гетеродонтные (дифференцированные) и текодонтные (помещенные в лунки). Он имел краевые ряды относительно мелких зубов, причем ряды не образовывали прямую режущую кромку. Зубы были расположены косо по длине челюстей и слегка перекрывали друг друга спереди назад. С каждой стороны наиболее полный экземпляр (УАЛВП 2) имел по три зуба в предчелюстной кости, шестнадцать в верхней челюсти (оба являются частью верхней челюсти) и семнадцать в зубной кости нижней челюсти. Зубы предчелюстной кости были отделены от задних зубов верхней челюсти короткой диастемой (промежутком), а два ряда предчелюстной кости были разделены беззубым промежутком спереди. Зубы в передней части верхней челюсти (предчелюстной кости) и передней нижней челюсти были сходны; у них были более высокие, более заостренные и изогнутые короны, а также «пятка» сзади. Передние зубы нижней челюсти были крупнее, чем верхние. На передних краях коронок было восемь зубчиков (зубцов), на заднем — от девяти до одиннадцати. Зубы в задней части верхней (верхней) и нижней челюсти имели треугольную форму при виде сбоку и сжатые при виде спереди. У них были длинные корни овального сечения, а короны имели выраженную выпуклость. цингулумы у их оснований. Зубцы здесь сжаты и направлены к вершине коронок. И внешняя, и внутренняя стороны зубных коронок были покрыты эмалью , и обе стороны были разделены вертикально гребнем. На каждом крае было около семи или восьми зубчиков, причем на переднем крае обычно их было больше всего. [ 3 ]
Череп Stegoceras можно отличить от черепа других пахицефалозавров по таким особенностям, как выраженный теменно-чешуйчатый выступ (хотя с возрастом он стал меньше), «зарождающийся» купол лобно-теменной части (хотя с возрастом купол увеличивался), раздутые носовые кости. , орнаментация бугорков по бокам и задней части чешуйчатых костей, ряды до шести бугорков на верхней стороне каждой чешуйчатой кости и до два узла в задней проекции теменной кости. Он также отличается отсутствием украшений на носу и уменьшенной диастемой. [ 4 ] [ 34 ] Череп S. noomexicanum можно отличить от черепа S. validum по таким особенностям, как вытянутая назад теменная кость, более редуцированная и треугольная форма, наличие более крупных надвисочных окон (хотя это может быть связано с возможным ювенильным статусом экземпляров). ), и имеющие примерно параллельные шовные контакты между чешуйчатой и теменной костью. Также, по-видимому, у него был меньший лобный выступ, чем у S. validum . [ 25 ] [ 27 ] и, кажется, в целом был более грациозным. [ 28 ]
Посткраниальный скелет
[ редактировать ]




Позвоночный столб Stegoceras . известен не полностью Сочленение между зигагофизами (суставными отростками) последовательных спинных (задних) позвонков, по-видимому, предотвращало боковое движение позвоночного столба, что делало его очень жестким, и оно дополнительно укреплялось окостеневшими сухожилиями . [ 3 ] Хотя шейные позвонки неизвестны, наклоненный вниз затылочный мыщелок (который сочленяется с первым шейным позвонком) указывает на то, что шея находилась в изогнутом положении, подобном S- или U-образной форме шеей большинства динозавров. [ 35 ] Судя по их положению у Homalocephale , окостеневшие сухожилия, обнаруженные при UALVP 2, образовывали сложную « хвостовую корзину » в хвосте, состоящую из параллельных рядов, при этом концы каждого сухожилия последовательно контактировали со следующим. Такие структуры называются myorhabdoi и известны только у костистых рыб ; Эта особенность уникальна для пахицефалозавров среди четвероногих (четырехногих) животных и, возможно, способствовала укреплению хвоста. [ 9 ]
Лопатка плечевой (лопатка) была длиннее кости (кость плеча); его лезвие было тонким и узким и слегка изогнутым, повторяя контур ребер. Лопатка не расширена на верхнем конце, но сильно расширена у основания. Коракоид . преимущественно тонкий и пластинчатый Плечевая кость имела тонкий стержень, слегка искривлена по длине и слегка изогнута. Дельтопекторальный гребень (где прикрепляются дельтовидная и грудная мышцы) развит слабо. Концы локтевой кости были расширены, а вдоль туловища простирались гребни. Лучевая кость была более прочной, чем локтевая, что необычно. При взгляде сверху тазовый пояс был очень широким для двуногого архозавра и становился шире к задней части. Широкая область таза могла вместить заднее расширение кишечника. была Подвздошная кость удлиненной, а седалищная кость — длинной и тонкой. Хотя лобковая кость неизвестна, вероятно, она была уменьшена в размерах, как и у Homalocephale . Бедренная кость (бедренная кость) была тонкой и изогнутой внутрь, большеберцовая кость была тонкой и изогнутой, а малоберцовая кость была тонкой и широкой на верхнем конце. стопы Плюсневая кость , по-видимому, была узкой, а единственная известная когтевая кость (когтевая кость) пальца была тонкой и слегка изогнутой. [ 3 ] Хотя конечности Stegoceras до конца не известны, скорее всего, они, как и другие пахицефалозавры, имели пять пальцев на руках и четыре пальца на ногах. [ 33 ]
Классификация
[ редактировать ]
В 1970-е годы по азиатским окаменелостям было описано больше родов пахицефалозавров, что дало больше информации об этой группе. В 1974 году Марьянска и Осмольска пришли к выводу, что пахицефалозавры достаточно различны, чтобы гарантировать отдельный подотряд внутри Ornithischia, Pachycephalosauria. В 1978 году китайский палеонтолог Дун Чжимин разделил Pachycephalosauria на два семейства; куполообразные Pachycephalosauridae (включая Stegoceras ) и плоскоголовые Homalocephalidae (первоначально называвшиеся Homalocephaleridae). [ 36 ] Уолл и Гальтон не нашли оправданным статус подотряда пахицефалозавров в 1979 году. [ 15 ] К 1980-м годам родство пахицефалозавров с Ornithischia оставалось невыясненным. Основные конкурирующие мнения заключались в том, что эта группа была ближе всего либо к орнитоподам, либо к цератопсам, последняя точка зрения объяснялась сходством между скелетом Stegoceras и «примитивным» цератопсовым протоцератопсом . В 1986 году американский палеонтолог Пол Серено поддержал родство между пахицефалозаврами и цератопсами и объединил их в группу Marginocephalia , основываясь на схожих черепных особенностях, таких как «полочная» структура над затылком. Он признал, что доказательства существования этой группы не являются подавляющими, но достоверность группы была подтверждена Сьюсом и Гальтоном в 1987 году. [ 3 ]
К началу 21 века по посткраниальным останкам было известно лишь несколько родов пахицефалозавров, а многие таксоны были известны только по куполам, что затрудняло классификацию внутри группы. Таким образом, пахицефалозавры в основном определяются чертами черепа, такими как плоская или куполообразная лобно-теменная кость, широкая и уплощенная полоса вдоль заглазничных и чешуйчатых костей, а также чешуйчатые кости, представляющие собой глубокие пластины на затылке. [ 31 ] В 1986 году Серено разделил пахицефалозавров на разные группы в зависимости от степени выпуклости их черепов (сгруппированных в ныне недействительные таксоны, такие как «Tholocephalidae» и «Domocephalinae»), а в 2000 году он считал «частично» Stegoceras куполообразных переход между предположительно «примитивными» плоскоголовыми и развитыми «полностью» куполообразными родами (такими как Pachycephalosaurus ). [ 37 ] В последующие годы от разделения пахицефалозавров на куполообразные и плоскоголовые отказались, поскольку в большинстве ревизий плоские головы считались педоморфными (ювенильными) или производными признаками, но не признаком полового диморфизма . В 2006 году Салливан выступил против идеи о том, что степень выпуклости полезна для определения таксономического сходства между пахицефалозаврами. [ 31 ] В 2003 году Салливан обнаружил, что сам Stegoceras является более базальным (или «примитивным»), чем «полностью куполообразные» представители подсемейства Pachycephalosaurinae, развивая выводы, к которым пришел Серено в 1986 году. [ 4 ]
проведенный Эвансом и его коллегами в 2013 году, Филогенетический анализ, показал, что некоторые роды плоскоголовых пахицефалозавров были более тесно связаны с «полностью» куполообразными таксонами, чем с «неполностью» куполообразными Stegoceras , что позволяет предположить, что они представляют собой молодь куполообразных таксонов, и что плоские головы не указывают на таксономическое родство. [ 38 ] ниже На кладограмме показано расположение Stegoceras среди Pachycephalosauridae по данным Шотта и его коллег, 2016: [ 20 ]

| Пахицефалозаврия |
| ||||||||||||||||||||||||
Биогеография и ранняя эволюционная история пахицефалозавров плохо изучены и могут быть прояснены только благодаря новым открытиям. Пахицефалозавры внезапно появляются в летописи окаменелостей и присутствуют как в Северной Америке, так и в Азии, поэтому неизвестно, когда они впервые возникли и в каком направлении рассеялись. Самые старые известные представители группы (такие как Acrotholus ) имеют «полностью куполообразную форму» и известны с сантонского этапа позднего мелового периода (около 84 миллионов лет назад). Это произошло до предположительно более примитивных Stegoceras из среднего кампана (77 миллионов лет назад) и Homalocephale из раннего маастрихта (70 миллионов лет назад), поэтому куполообразный череп может быть гомопластической чертой (формой конвергентной эволюции ). Позднее появление пахицефалозавров по сравнению с родственными цератопсами указывает на длинную призрачную линию (предполагаемую, но отсутствующую в летописи окаменелостей), охватывающую 66 миллионов лет, от поздней юры до мела. Поскольку пахицефалозавры были в основном небольшими, это может быть связано с тафономическая предвзятость ; более мелкие животные с меньшей вероятностью сохранятся в результате окаменелости. Более тонкие кости также с меньшей вероятностью сохранятся, поэтому пахицефалозавры известны в основном по их крепким черепам. [ 4 ] [ 38 ]
Палеобиология
[ редактировать ]Механика кормления
[ редактировать ]
Неизвестно, чем питались пахицефалозавры; Имея очень маленькие ребристые зубы, они не могли жевать жесткие волокнистые растения так же эффективно, как другие динозавры того же периода. Предполагается, что их острые, зазубренные зубы идеально подходили для смешанной диеты, состоящей из листьев, семян, фруктов и насекомых. [ 39 ] Stegoceras , возможно, питались полностью травоядными животными, поскольку их зубные коронки были похожи на таковые у игуанидных ящериц. На предчелюстных зубах имеются фасетки износа из-за контакта с предзубной костью, а на верхнечелюстных зубах имеются двойные фасетки износа, подобные тем, которые наблюдаются у других птицетазовых динозавров. Каждый третий верхнечелюстной зуб у UALVP 2 является прорезывающимся замещающим зубом , а замена зубов происходила в обратном порядке в последовательных трех зубах. Затылочная область Stegoceras была хорошо разграничена для прикрепления мышц, и считается, что движение челюсти Stegoceras и других пахицефалозавров в основном ограничивалось движениями вверх и вниз с лишь небольшой способностью к вращению челюсти. Это основано на строении челюсти, микроизносе зубов и фасетках износа зубов, что указывает на то, что сила укуса использовалась больше для сдвига, чем для дробления. [ 3 ] [ 40 ]
В 2021 году канадский палеонтолог Майкл Н. Хаджинс и его коллеги исследовали зубы Stegoceras и Thescelosaurus и обнаружили, что, хотя у обоих были гетеродонтные зубы, их можно статистически отличить друг от друга. Из-за своего широкого рострума и более однородных зубов Stegoceras питался неразборчивыми массами и собирал большое количество растительности, в то время как зубы и узкий рострум Thescelosaurus указывают на то, что он питался избирательно. Пахицефалозавры и тесцелозавриды встречаются в одних и тех же формациях Северной Америки, и похоже, что их сосуществование стало возможным благодаря тому, что они занимали разные экоморфопространства (хотя сами стегоцеры и тесцелозавры не были современниками). [ 41 ]
Носовые ходы
[ редактировать ]В 1989 году Эмили Б. Гриффин обнаружила, что Stegoceras и другие пахицефалозавры обладали хорошим обонянием (обонянием), основываясь на изучении краниальных эндокастов , которые показали большие обонятельные луковицы в мозгу. [ 42 ] В 2014 году Джейсон М. Бурк и его коллеги обнаружили, что Stegoceras потребовались бы хрящевые носовые раковины в передней части носовых ходов, чтобы поток воздуха достигал обонятельной области. Доказательством присутствия этой структуры является костный гребень, к которому она могла прикрепиться. Размер обонятельной области также указывает на то, что у Stegoceras было острое обоняние. Исследователи обнаружили, что у динозавра могли быть носовые раковины в форме свитка (как у индейки ) или разветвленные (как у страуса ), поскольку оба они могли направлять воздух в обонятельную область. Система кровеносных сосудов в проходах также позволяет предположить, что носовые раковины служили для охлаждения теплой артериальной крови из тела, направлявшейся в мозг. Череп S. validum экз. УАЛВП 2 пригоден для такого рода исследований благодаря исключительной сохранности; в полости носа имеются оссифицированные мягкие ткани, которые в противном случае были бы хрящевыми и, следовательно, не сохранились бы за счет минерализации. [ 43 ]
Онтогенетические изменения
[ редактировать ]
Исторически было предложено несколько объяснений вариаций, наблюдаемых в черепах Stegoceras и других пахицефалозавров. Браун и Шлайкьер предположили, что существует половой диморфизм в степени выпуклости, и выдвинули гипотезу, что экземпляры с плоской головой, такие как AMNH 5450 ( Ornatotholus ), представляют собой женскую морфу Stegoceras . Эта идея была поддержана морфометрическим исследованием 1981 года, проведенным Чампаном и его коллегами, которое показало, что у самцов купола больше и толще. [ 34 ] [ 44 ] После того, как были обнаружены другие плоскоголовые пахицефалозавры, степень выпуклости была предложена как признак, имеющий таксономическую важность, и поэтому AMNH 5450 считался отдельным таксоном с 1979 года. Вместо этого в 1998 году Гудвин и его коллеги предположили, что вздутие купола является онтогенетической особенностью, которая меняется с возрастом, основываясь на гистологическом исследовании черепа S. validum , которое показало, что купол состоит из сосудистой, быстрорастущей кости, что соответствует увеличение доминирования с возрастом. Эти авторы обнаружили, что предположительно отличительные черты Ornatotholus легко могли быть результатом онтогенеза. [ 34 ] [ 17 ]
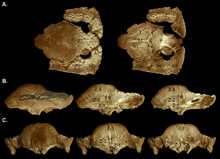
В 2003 году Уильямсон и Карр опубликовали гипотетические серии исследований роста S. validum , показывающие Ornatotholus как ювенильную стадию. Они предположили, что молодые особи характеризуются плоской, утолщенной лобно-теменной крышей, с более крупными надвисочными окнами и усеяны близко расположенными бугорками и узлами. Теменно-чешуйчатый выступ не уменьшен в размерах, лобно-теменной шов открыт. У полувзрослых особей купола были курганообразными, задняя часть теменной и черепной крыши плоская. Надвисочные окна имели асимметрию размеров, а закрытие лобно-теменного шва было вариабельным. Узлы были растянуты или почти стерты по мере расширения купола во время роста, при этом оставалась мозаичная поверхность. Узор часто стирался в самой высокой точке (вершине) купола, области, где происходило максимальное расширение. Бугорки на черепе были вытянуты в разные стороны, а на краю теменно-чешуйчатого выступа, возможно, представляли собой гипертрофированные (увеличенные) бугорки. Задняя и боковые стороны черепов молодых и взрослых особей были украшены менее видоизмененными бугорками. Прежде чем быть включенными в расширяющийся купол, кости черепа расширились, в результате чего между этими костями образовались соединения. Купол взрослой особи был широким и выпуклым и включал в себя большую часть выступа, который уменьшился в размерах и нависал над затылком в виде толстой «губы». Надвисочные окна были закрыты, но шов между лобно-теменной и соединяющимися костями черепа не всегда был закрытым у взрослых и подростков. [ 19 ]
В 2011 году Шотт и его коллеги провели более всесторонний анализ онтогенеза краниального купола у S. validum . Исследование показало, что теменно-чешуйчатый выступ сохранял расположение орнаментов на протяжении всего роста, а васкуляризация лобно-теменных куполов уменьшалась с увеличением размера. Также было обнаружено, что форма и размер купола сильно коррелировали с ростом, и что рост был аллометрическим (в отличие от изометрического ) от плоского до куполообразного, что подтверждает статус Ornatotholus как молодого Stegoceras . Они также предположили, что эта модель роста купола с резкими изменениями от юного возраста к взрослому была общей траекторией развития пахицефалозавров. Эти исследователи отметили, что, хотя наблюдение Уильямсона и Карра о том, что надвисочные отверстия закрываются с возрастом, в целом верно, все же существует высокая степень индивидуальных различий в размерах этих окон, независимо от размера лобно-теменных окон, и поэтому эта особенность может иметь значение. были независимыми от онтогенеза. [ 34 ]
Исследование Шотта и Эванса, проведенное в 2012 году, показало, что количество и форма отдельных узлов на чешуйчатой полке исследованных черепов S. validum значительно различаются, и что эта изменчивость, по-видимому, не коррелирует с онтогенными изменениями, а обусловлена индивидуальными вариациями. . Эти исследователи не обнаружили корреляции между шириной надвисочных окон и размером чешуйчатой кости. [ 45 ]
Функция купола
[ редактировать ]Функция куполов пахицефалозавра обсуждалась, а Stegoceras использовался в качестве модели для экспериментов в различных исследованиях. Купол в основном интерпретировался как оружие, используемое во внутривидовых боях , структура сексуального проявления или средство распознавания видов . [ 46 ] [ 47 ]
Бой
[ редактировать ]
Гипотеза о том, что куполообразные черепа Stegoceras и других пахицефалозавров использовались для бодания голов, была впервые выдвинута американским палеонтологом Эдвином Колбертом в 1955 году. В 1970 и 1971 годах Гальтон развил эту идею и утверждал, что, если бы купол был просто декоративным, он бы были менее плотными, и что структура была идеальной для сопротивления силе. Гальтон предположил, что, когда Стегоцерас держит череп вертикально, перпендикулярно шее, сила будет передаваться от черепа с небольшой вероятностью его смещения, и поэтому купол можно будет использовать в качестве тарана. Он полагал, что он вряд ли использовался в основном для защиты от хищников, поскольку на самом куполе отсутствовали шипы, а шипы теменно-чешуйчатого выступа находились в «неэффективном» положении, но счел его совместимым с внутривидовой конкуренцией. Гальтон представлял, что купола сколоты, а позвоночник удерживается в горизонтальном положении. Это можно было делать либо лицом друг к другу при нанесении ударов, либо атакуя друг друга с опущенными головами (аналог современных овец и коз). Он также отметил, что жесткость спины была бы полезна при использовании для этой цели головы. В 1978 году Сьюс согласился с Гальтоном в том, что анатомия пахицефалозавров соответствует передаче ударного напряжения от купола к куполу, основываясь на тестах с модели из оргстекла . Удар будет поглощаться шеей и телом, а связки и мышцы шеи предотвратят травмы от скользящих ударов (как у современных снежных баранов ). Сьюс также предположила, что животные могли бодать друг друга боками. [ 35 ] [ 48 ] [ 49 ]
В 1997 году американский палеонтолог Кеннет Карпентер отметил, что спинные позвонки со спины пахицефалозавра Homalocephale показывают, что спина изогнута вниз непосредственно перед шеей (которая не сохранилась), и если шея не изогнута вверх, голова будет указывать на земля. Поэтому он пришел к выводу, что шеи Stegoceras и других пахицефалозавров находились в изогнутой позе (что является нормой для динозавров), и поэтому они не могли выровнять голову, шею и тело горизонтально прямо, что было бы необходим для передачи стресса. Их шеи пришлось бы держать ниже уровня спины, что могло бы привести к повреждению спинного мозга при ударе. Современные снежные бараны и бизоны решают эту проблему за счет наличия прочных связок от шеи до высоких нервных отростков над плечами (которые поглощают силу удара), но у пахицефалозавров такие особенности неизвестны. Эти животные также поглощают силу удара через пазухи у основания рогов, а их лоб и рога образуют широкую контактную поверхность, в отличие от узкой поверхности куполов пахицефалозавров. Потому что купол Стегоцерас был закругленным, это давало бы очень небольшую площадь для потенциального удара, и купола отскакивали бы друг от друга (если только удар не был точно центрирован). Сражающимся пахицефалозаврам было бы трудно видеть друг друга, когда их головы были опущены, из-за костных гребней над глазами. [ 35 ]


Из-за проблем, которые он обнаружил в гипотезе удара головой, Карпентер вместо этого предположил, что купола были адаптацией для удара с фланга (как это наблюдается у некоторых крупных африканских млекопитающих); он представлял себе, что два животных будут стоять параллельно, лицом друг к другу или в одном направлении, и наносить удары в сторону противника. Следовательно, относительно большая ширина тела пахицефалозавров могла служить для защиты жизненно важных органов от повреждений во время бодания с фланга. Не исключено, что Stegoceras и подобные ему пахицефалозавры наносили удары движением шеи сбоку и поворотом головы. Верхние стороны купола имеют наибольшую площадь поверхности и, возможно, были местом удара. Толщина купола увеличила бы силу удара в стороны, и это гарантировало бы, что противник ощутит силу удара, не получив при этом серьезных травм. Костный ободок над глазницей мог защищать глаз агрессора при нанесении удара. Карпентер предположил, что пахицефалозавры сначала занимались демонстрация угрозы , покачивая и подставляя головы, чтобы показать размер своих куполов (запугивание), а затем нанося друг другу удары, пока один из противников не подал сигнал о подчинении. [ 35 ]
В 2008 году Эрик Снивели и Адам Кокс проверили характеристики черепов пахицефалозавров в 2D и 3D с помощью анализа методом конечных элементов и обнаружили, что они могут выдерживать значительные удары; больший свод куполов позволял увеличить силу удара. Они также считали вероятным, что купола пахицефалозавров были покрыты ) , и поэтому кератином, прочным материалом, который может выдерживать большую энергию без необратимых повреждений (как остеодермы крокодилов включили кератин в свою тестовую формулу. [ 50 ] В 2011 году Снивели и Джессика М. Теодор провели анализ методом конечных элементов, моделируя удары головой с помощью компьютерной томографии черепов S. validum (UALVP 2), Prenocephale prenes , бьющихся головой и нескольких существующих парнокопытных . Они обнаружили, что корреляция между ударением головой и морфологией черепа, обнаруженная у живых животных, также существовала у изученных пахицефалозавров. Stegoceras и Prenocephale имели форму черепа, похожую на снежного барана, с губчатой костью, защищающей мозг. Они также имели сходство в распределении компактных и губчатых областей с снежным бараном, белобрюхим дукером и жирафом . Белобрюхий дукер оказался ближайшим морфологическим аналогом Stegoceras ; у этого вида, бьющегося головой, купол меньшего размера, но такой же округлый. Stegoceras был лучше способен рассеивать силу, чем парнокопытные, которые бодались головами с большой силой, но менее васкуляризированные купола старых пахицефалозавров и, возможно, пониженная способность к заживлению после травм, выступали против такого боя у пожилых людей. В ходе исследования также проверили влияние кератинового покрытия купола и обнаружили, что оно улучшает производительность. Хотя У Stegoceras отсутствовали пневматические пазухи, которые находятся ниже точки удара в черепах парнокопытных, ударяющих головой, вместо этого у них были сосудистые стойки, которые могли аналогичным образом действовать как скобы, а также каналы для питания развития кератинового покрытия. [ 51 ]


В 2012 году Калеб М. Браун и Энтони П. Рассел предположили, что затвердевшие хвосты, вероятно, не использовались в качестве защиты от ударов с флангов, но, возможно, позволяли животным принимать треножную стойку во время внутривидового боя, используя хвост в качестве опоры. . Браун и Рассел обнаружили, что таким образом хвост может помочь противостоять сжимающим, растягивающим и скручивающим нагрузкам, когда животное наносит или получает удары куполом. [ 9 ] Исследование Джозефа Э. Петерсона и его коллег в 2013 году выявило повреждения черепов Stegoceras и других пахицефалозавров, которые были интерпретированы как инфекции, вызванные травмой. Поражения были обнаружены на 22% отобранных черепов пахицефалозавров (частота соответствует всем родам), но отсутствовали у плоскоголовых особей (которые были интерпретированы как молодые особи или самки), что согласуется с использованием во внутривидовых боях (за территорию). или товарищи). Распределение поражений у этих животных имело тенденцию концентрироваться в верхней части купола, что подтверждает поведение, связанное с боданием головой. Удар с фланга, вероятно, приведет к меньшему количеству травм, которые вместо этого будут сосредоточены по бокам купола. Эти наблюдения были сделаны при сравнении повреждений с повреждениями на черепах и боках скелетов современных овец. Исследователи отметили, что современные животные, бьющиеся головой, используют свое оружие как для боя, так и для демонстрации, и поэтому пахицефалозавры могли также использовать свои купола для того и другого. Демонстрации оружия и готовности его использовать может быть достаточно для разрешения споров у некоторых животных. [ 46 ]
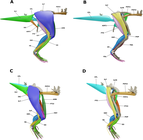
Брайан Р.С. Мур и его коллеги исследовали и реконструировали мускулатуру конечностей Stegoceras в 3D в 2022 году, используя в качестве основы очень полный образец UALVP 2. Они обнаружили, что мускулатура передних конечностей была консервативной, особенно по сравнению с таковой у ранних двуногих ящеротазовых динозавров, но мускулатура таза и задних конечностей была более развитой (или «продвинутой») из-за особенностей скелета. Эти области имели большие мышцы, а в сочетании с широким тазом и толстыми задними конечностями (и, возможно, увеличенными связками) это приводило к сильной и стабильной структуре таза, которая помогла бы во время столкновений головами между людьми. Поскольку купола черепа пахицефалозавров росли с положительной аллометрией и, возможно, использовались в бою, эти исследователи предположили, что то же самое могло произойти и с мышцами задних конечностей, если они использовались для продвижения тела вперед во время удара головой. Они предупредили, что, хотя UALVP 2 является очень полным для пахицефалозавра, их исследование было ограничено отсутствием больших частей его позвоночника и элементов внешних конечностей. [ 52 ]
Другие предлагаемые функции
[ редактировать ]
В 1987 году Дж. Кейт Ригби и его коллеги предположили, что купола пахицефалозавров были теплообменными органами, используемыми для терморегуляции , на основе их внутренних «излучающих структур» ( трабекул ). Эту идею поддержали и некоторые другие писатели в середине 1990-х годов. [ 47 ] В 1998 году Гудвин и его коллеги сочли отсутствие пазух в черепе Stegoceras и « сотовую » сеть сосудистой кости купола плохо подходящей для ударов головой и отметили, что кости, прилегающие к куполу, рискуют сломаться. во время такого контакта. Основываясь на идее о том, что окостеневшие сухожилия, которые укрепляли хвосты Stegoceras и других пахицефалозавров, позволяли им принимать треножную позицию (впервые предложенную Марьянской и Осмольской в 1974 году), Гудвин и др. предположил, что эти структуры могли защитить хвост от ударов с фланга или что сам хвост мог использоваться в качестве оружия. [ 17 ] В 2004 году Гудвин и его коллеги изучили краниальную гистологию пахицефалозавров и обнаружили, что васкуляризация (включая трабекулы) куполов уменьшается с возрастом, что, по их мнению, не соответствует функции ни ударения головой, ни теплообмена. Они также предположили, что плотный слой шарпиевых волокон вблизи поверхности купола указывает на то, что при жизни он имел внешнее покрытие, из-за чего узнать форму купола у живого животного невозможно. Вместо этого эти исследователи пришли к выводу, что купола предназначались в основном для распознавания видов и общения (как у некоторых африканских коров ), и что их использование в сексуальных проявлениях было лишь вторичным. Они также предположили, что внешнее покрытие куполов было ярко окрашено при жизни или могло менять цвет в зависимости от сезона. [ 47 ]
В 2011 году американские палеонтологи Кевин Падиан и Джон Р. Хорнер предположили, что «причудливые структуры» у динозавров в целом (включая купола, оборки, рога и гребни) в первую очередь использовались для распознавания видов, и отвергли другие объяснения как неподтвержденные доказательствами. Среди других исследований эти авторы процитировали статью Гудвина и др. 2004 года о куполах пахицефалозавров как подтверждение этой идеи и отметили, что такие структуры, по-видимому, не являются сексуально диморфными. [ 53 ] В ответ Падиану и Хорнеру в том же году Роб Дж. Нелл и Скотт Д. Сэмпсон утверждали, что распознавание видов вполне возможно как вторичная функция «причудливых структур» у динозавров, но что половой отбор (используемый в демонстрации или бою для конкурировать за партнеров) было более вероятным объяснением из-за высокой стоимости их развития, а также из-за того, что такие структуры, по-видимому, сильно варьируются внутри вида. [ 54 ] В 2013 году британские палеонтологи Дэвид Э. Хоун и Даррен Нэйш раскритиковали «гипотезу распознавания видов» и заявили, что ни одно из ныне живущих животных не использует такие структуры в первую очередь для распознавания видов, и что Падиан и Хорнер проигнорировали возможность взаимного полового отбора (где оба пола украшены орнаментами). [ 55 ]
В 2012 году Шотт и Эванс предположили, что регулярность чешуйчатого орнамента на протяжении всего онтогенеза Stegoceras соответствует распознаванию видов, но переход от плоских к куполообразным лобно-теменным отделам в позднем возрасте предполагает, что функция этого признака менялась в онтогенезе и, возможно, была обусловлена половым путем. выбран, возможно, для внутривидового боя. [ 45 ] В 2023 году Дайер и его коллеги обнаружили, что экземпляры Stegoceras различаются по толщине лобно-носового выступа и что черепа с наибольшим количеством костных патологий были черепами с самыми высокими выступами, что, по их мнению, указывает на то, что изменение толщины выступа представляет собой межполовые вариации. [ 22 ] В 2023 году Хорнер и его коллеги заявили, что, поскольку купол и связанный с ним орнамент Стегоцераса и орнамент Пахицефалозавра развились в раннем возрасте, это указывает на то, что они использовались для визуальной коммуникации, чтобы молодые особи могли узнавать других молодых особей, а взрослые - других взрослых. Они не исключали, что эти особенности могли быть использованы и для других целей, в том числе для ударов головой, но не считали травмы, наблюдаемые у экземпляров, доказательством этого. Они также предположили, что особенности черепов некоторых пахицефалозаврид указывают на то, что купол мог поддерживать более крупную ороговевшую структуру, чем просто шляпу. [ 56 ]
Палеосреда
[ редактировать ]
S. validum известен из позднемеловой группы реки Белли (канадский эквивалент группы реки Джудит в США), а образцы были обнаружены в формации Парка динозавров (поздний кампан, 76,5–75 млн лет назад) в Провинциальном парке динозавров. (включая образец лектотипа) и формацию Олдман (средний кампан, 77,5–76,5 млн лет назад) в Альберте, Канада. пахицефалозавры Hanssuesia (если не синоним Stegoceras ) и Foraminacephale . Из обеих формаций также известны [ 4 ] [ 22 ] S. noomexicanum известен из Фрутленда (поздний кампан, около 75 млн лет назад) и нижней части формации Киртланд (поздний кампан , около 74 млн лет назад) в Нью-Мексико, и если бы этот вид правильно принадлежал Stegoceras , этот род имел бы широкое географическое распространение. . [ 25 ] Присутствие подобных пахицефалозавров как на западе, так и на севере Северной Америки в позднем меловом периоде показывает, что они были важной частью тамошней фауны динозавров. [ 27 ]
Традиционно предполагалось, что пахицегалозавры населяли горные районы; износ их черепов предположительно был результатом того, что их скатило водой с возвышенностей, и сравнение с снежными баранами подтвердило эту теорию. В 2014 году Джордан К. Мэллон и Эванс оспорили эту идею, поскольку износ и первоначальное расположение черепов не соответствуют транспортировке таким способом, и вместо этого предположили, что североамериканские пахицефалозавры населяли аллювиальные (связанные с водой) и прибрежные равнинные условия. [ 57 ]
Формация «Парк динозавров» интерпретируется как низкий рельеф рек и пойм , которые со временем стали более заболоченными и находились под влиянием морских условий по мере того, как Западный внутренний морской путь продвигался на запад. [ 58 ] Климат был теплее , чем в современной Альберте, без морозов , но с более влажными и сухими сезонами. Хвойные деревья, по-видимому, были доминирующими растениями кроны , с подлеском из папоротников , древовидных папоротников и покрытосеменных растений . [ 59 ] Парк динозавров известен своим разнообразным сообществом травоядных животных. Помимо Stegoceras , в формации также обнаружены окаменелости цератопсов Centrosaurus , Styracosaurus и Chasmosaurus , гадрозавридов Prosaurolophus , Lambeosaurus , Gryposaurus , Corythosaurus и Parasaurolophus , а также анкилозавров Edmontonia и Euoplocephalus . Присутствующие тероподы включают тираннозаврид горгозавра и дасплетозавра . [ 60 ] Другие динозавры, известные из формации Олдмана, включают гадрозавров Brachylophosaurus , цератопсов Coronosaurus и Albertaceratops , орнитомимид , теризинозавров и, возможно, анкилозавров. К тероподам относились троодонтиды , овирапторозавры , дромеозавриды Saurornitholestes и, возможно, тираннозавр-альбертозавр. [ 61 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Ламбе, LM (1902). «Новые роды и виды из серии Белли-Ривер (середина мела)» . Геологическая служба Канады, Вклад в канадскую палеонтологию . 3:68 .
- ^ Jump up to: а б с Хэтчер, Дж. Б.; Лулл, РС; Марш, ОК; Осборн, HF (1907). «Цератопсия» . Монографии Геологической службы США . XLIX . дои : 10.5962/bhl.title.60500 .
- ^ Jump up to: а б с д и ж г час я дж к л Сьюс, HD и Гальтон, PM (1987). «Анатомия и классификация североамериканских пахицефалозавров (Dinosauria: Ornithischia)» . Палеонтографика Абтейлунг А. 198 : 1–40.
- ^ Jump up to: а б с д и ж г Салливан, РМ (2003). «Ревизия динозавра Stegoceras Lambe (Ornithischia, Pachycephalosauridae)». Журнал палеонтологии позвоночных . 23 (1): 181–207. doi : 10.1671/0272-4634(2003)23[181:rotdsl]2.0.co;2 . S2CID 85894105 .
- ^ Нопца, Ф. (1903). «О стегоцерасах и стереоцефалах ». Centralblatt по минералогии, геологии и палеонтологии . 1903 : 266–267.
- ^ Ламбе, LM (1903). «Новейшая зоопалеонтология» . Наука . 18 (445): 60. Бибкод : 1903Sci....18...60L . дои : 10.1126/science.18.445.60 . JSTOR 1631645 . ПМИД 17746863 .
- ^ Ламбе, LM (1918). «Меловой род Stegoceras, олицетворяющий новое семейство, условно отнесенный к Stegosauria» . Труды Королевского общества Канады . 12 (4): 23–36.
- ^ Гилмор, CW (1924). «О Troodon validus , ортоподном динозавре из мелового периода реки Белли в Альберте, Канада». Вестник факультета геологии Университета Альберты . 1 :1–43.
- ^ Jump up to: а б с д Браун, СМ; Рассел, AP; Фарке, А.А. (2012). «Гомология и архитектура хвостовой корзины пахицефалозавров (Dinosauria: Ornithischia): первое появление Myorhabdoi у четвероногих» . ПЛОС ОДИН . 7 (1): e30212. Бибкод : 2012PLoSO...730212B . дои : 10.1371/journal.pone.0030212 . ПМК 3260247 . ПМИД 22272307 .
- ^ Jump up to: а б Браун, Б.; Э.М., Шлайкьер (1943). «Исследование динозавров-троодонтов с описанием нового рода и четырех новых видов». Бюллетень АМНХ . 82 . HDL : 2246/387 .
- ^ Штернберг, CM (1945). «Pachycephalosauridae, предложенные для куполообразного динозавра, Stegoceras lampei , n. sp., описаны». Журнал палеонтологии . 19 (5): 534–538. JSTOR 1299007 .
- ^ Рассел, Л.С. (1948). «Дентария троодона , рода тероподовых динозавров». Журнал палеонтологии . 22 (5): 625–629. JSTOR 1299599 .
- ^ Болин, Б., 1953. Ископаемые рептилии из Монголии и Ганьсу. Отчеты научной экспедиции в северо-западные провинции Китая под руководством доктора Свена Хедина. VI. Палеонтология позвоночных 6. Публикация китайско-шведской экспедиции 37: 1–113.
- ^ Кун, О., 1964, Каталог окаменелостей I: Animalia, часть 105. Ornithischia (Приложение I) , IJsel Pers, Deventer, 80 стр.
- ^ Jump up to: а б Уолл, WP; Гальтон, премьер-министр (1979). «Заметки о динозаврах-пахицефалозавридах (Reptilia: Ornithischia) из Северной Америки с комментариями об их статусе орнитопод». Канадский журнал наук о Земле . 16 (6): 1176–1186. Бибкод : 1979CaJES..16.1176W . дои : 10.1139/e79-104 .
- ^ Гальтон, премьер-министр; Сьюс, Х.-Д. (1983). «Новые данные о динозаврах-пахицефалозавридах (Reptilia: Ornithischia) из Северной Америки». Канадский журнал наук о Земле . 20 (3): 462–472. Бибкод : 1983CaJES..20..462G . дои : 10.1139/e83-043 .
- ^ Jump up to: а б с Гудвин, МБ; Бухгольц, Э.А.; Джонсон, Р.Э. (1998). «Краниальная анатомия и диагностика Stygimoloch spinifer (Ornithischia: Pachycephalosauria) с комментариями к структурам черепного дисплея в агонистическом поведении». Журнал палеонтологии позвоночных . 18 (2): 363–375. Бибкод : 1998JVPal..18..363G . дои : 10.1080/02724634.1998.10011064 .
- ^ Салливан, Роберт М. (2000). « Новый гребешок Prenocephale edmontonensis (Brown и Schlaikjer) и новый гребешок P. brevis (Lambe). (Dinosauria: Ornithischia: Pachycephalosauria) из верхнего мела Северной Америки» . Бюллетень Музея естественной истории и науки Нью-Мексико . 17 : 177–90.
- ^ Jump up to: а б Уильямсон, TE; Карр, Т.Д. (2003). «Новый род производных пахицефалозавров из западной части Северной Америки». Журнал палеонтологии позвоночных . 22 (4): 779–801. doi : 10.1671/0272-4634(2002)022[0779:ANGODP]2.0.CO;2 . S2CID 86112901 .
- ^ Jump up to: а б Шотт, РК; Эванс, округ Колумбия (2016). «Краниальные вариации и систематика Foraminacephale brevis gen. nov. и разнообразие пахицефалозавридных динозавров (Ornithischia: Cerapoda) в группе реки Белли в Альберте, Канада». Зоологический журнал Линнеевского общества . дои : 10.1111/zoj.12465 .
- ^ Эванс, Дэвид С.; Браун, Калеб М.; Ты, Хайлу; Кампионе, Николас Э. (октябрь 2021 г.). «Описание и пересмотренный диагноз первого зарегистрированного в Азии пахицефалозавра, Sinocephale bexelli gen. nov., из верхнего мела Внутренней Монголии, Китай». Канадский журнал наук о Земле . 58 (10): 981–992. Бибкод : 2021CaJES..58..981E . doi : 10.1139/cjes-2020-0190 . S2CID 244227050 .
- ^ Jump up to: а б с д Дайер, Аарон; Пауэрс, Марк; Карри, Филип (2023). «Проблемные предполагаемые пахицефалозавриды: синхротронная микрокомпьютерная томография проливает новый свет на анатомию и таксономическую достоверность Gravitholus albertae из группы реки Белли (кампан) в Альберте, Канада» . Анатомия позвоночных Морфология Палеонтология . 10 (1). дои : 10.18435/vamp29388 .
- ^ Салливан, РМ; Лукас, СГ (2006). «Пахицефалозавридный динозавр Stegoceras validum из верхнемеловой формации Fruitland, бассейн Сан-Хуан, Нью-Мексико» . Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 329–330.
- ^ Уильямсон, TE; Карр, Т.Д. (2002). «Молодой пахицефалозавр (Dinosauria: Pachycephalosauridae) из формации Fruitland». Нью-Мексико: Геология Нью-Мексико . 24 : 67–68.
- ^ Jump up to: а б с д Ясински, SE; Салливан, РМ (2011). «Повторная оценка пахицефалозаврид из перехода Фрутленд-Киртленд (Киртланд, поздний кампан), бассейн Сан-Хуан, Нью-Мексико, с описанием нового вида Stegoceras и повторной оценкой Texascephale langstoni » (PDF) . Ископаемая запись 3. Музей естественной истории и науки Нью-Мексико, Бюллетень . 53 : 202–215.
- ^ Ватабе, М.; Цогтбаатар, К.; Салливан, РМ (2011). «Новый пахицефалозаврид из формации Байншир (сеноман-поздний сантон), пустыня Гоби, Монголия» (PDF) . Ископаемая запись 3. Музей естественной истории и науки Нью-Мексико, Бюллетень . 53 : 489–497.
- ^ Jump up to: а б с д Уильямсон, TE; Брусатте, СЛ (2016). «Пахицефалозавры (Dinosauria: Ornithischia) из верхнего мела (верхний кампан) Нью-Мексико: переоценка Stegoceras noomexicanum » . Меловые исследования . 62 : 29–43. Бибкод : 2016CrRes..62...29W . дои : 10.1016/j.cretres.2016.01.012 .
- ^ Jump up to: а б Ясински, SE; Салливан, РМ (2016). «Достоверность позднемелового пахицефалозавра Stegoceras nomomexicanum (Dinosauria: Pachycephalosauridae)» . В Салливане, Роберт М.; Лукас, Спенсер Г. (ред.). Летопись окаменелостей 5: Бюллетень 74 . Музей естественной истории и науки Нью-Мексико. стр. 107–116.
- ^ Перенасыщение, Д.Ф. (1997). Динозавры: Энциклопедия . Джефферсон, Северная Каролина: McFarland & Co., стр. 834–838. ISBN 978-0-89950-917-4 .
- ^ Ламберт, Д. (1993). Самая лучшая книга о динозаврах . Нью-Йорк: Дорлинг Киндерсли. п. 155 . ISBN 978-1-56458-304-8 .
- ^ Jump up to: а б с Салливан, РМ (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» . Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 347–365. S2CID 4243316 .
- ^ Печкис, Дж. (1995). «Последствия оценок массы тела для динозавров». Журнал палеонтологии позвоночных . 14 (4): 520–533. Бибкод : 1995JVPal..14..520P . дои : 10.1080/02724634.1995.10011575 . JSTOR 4523591 .
- ^ Jump up to: а б Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. стр. 241–242 . ISBN 978-0-691-13720-9 .
- ^ Jump up to: а б с д Шотт, Райан К.; Эванс, Дэвид С.; Гудвин, Марк Б.; Хорнер, Джон Р.; Браун, Калеб Маршалл; Лонгрич, Николас Р. (29 июня 2011 г.). «Краниальный онтогенез Stegoceras validum (Dinosauria: Pachycephalosauria): количественная модель роста и изменения купола пахицефалозавра» . ПЛОС ОДИН . 6 (6): e21092. Бибкод : 2011PLoSO...621092S . дои : 10.1371/journal.pone.0021092 . ПМК 3126802 . ПМИД 21738608 .
- ^ Jump up to: а б с д Карпентер, Кеннет (1 декабря 1997 г.). «Агонистическое поведение пахицефалозавров (орнитисхия, динозаврозаврия); новый взгляд на поведение, связанное с боданием головой» . Геология Скалистых гор . 32 (1): 19–25.
- ^ Перл, А.; Осмольска, Х. (1982). « Goyocephale lattimorei gen. et sp. n., новый плоскоголовый пахицефалозавр (Ornlthlschia, Dinosauria) из верхнего мела Монголии – Acta Palaeontologica Polonica» . Acta Palaeontologica Polonica . 27 : 115–127.
- ^ Серено, ПК, 2000. Летопись окаменелостей, систематика и эволюция пахицефалозавров и цератопсов из Азии. 480–516 в Бентоне, М.Дж., М.А. Шишкине, Д.М. Анвине и Е.Н. Курочкине (ред.), Эпоха динозавров в России и Монголии . Издательство Кембриджского университета, Кембридж.
- ^ Jump up to: а б Эванс, округ Колумбия; Шотт, РК; Ларсон, Д.В.; Браун, СМ; Райан, MJ (2013). «Старейший североамериканский пахицефалозавр и скрытое разнообразие мелких птицетазовых динозавров» . Природные коммуникации . 4 : 1828. Бибкод : 2013NatCo...4.1828E . дои : 10.1038/ncomms2749 . ПМИД 23652016 .
- ^ Марьянская, Т.; Чепмен, RE; Вейшампель, Д.Б. (2004). «Пахицефалозаврия». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 464–477 . ISBN 978-0-520-24209-8 .
- ^ Набавизаде, А. (2016). «Эволюционные тенденции в механике приводящих челюстей птицетазовых динозавров» . Анатомическая запись . 299 (3): 271–294. дои : 10.1002/ar.23306 . ПМИД 26692539 .
- ^ Хаджинс, Майкл Нейлор; Карри, Филип Дж.; Салливан, Корвин (16 октября 2021 г.). «Стоматологическая оценка Stegoceras validum (Ornithischia: Pachycephalosauridae) и Thescelosaurus ignores (Ornithischia: Thescelosauridae): палеоэкологические выводы». Меловые исследования . 130 : 105058. doi : 10.1016/j.cretres.2021.105058 . S2CID 239253658 .
- ^ Гиффин, Э.Б. (1989). «Пахицефалозавр Палеоневрология (Archosauria: Ornithischia)». Журнал палеонтологии позвоночных . 9 (1): 67–77. Бибкод : 1989JVPal...9...67G . дои : 10.1080/02724634.1989.10011739 . JSTOR 4523238 .
- ^ Бурк, Дж. М.; Портер, Вм. Р.; Риджли, Колорадо; Лисон, ТР; Шахнер, ER; Белл, PR; Витмер, LM (2014). «Вдохнуть жизнь в динозавров: решить проблемы восстановления мягких тканей и носового дыхания у вымерших видов» . Анатомическая запись . 297 (11): 2148–2186. дои : 10.1002/ar.23046 . ПМИД 25312371 . S2CID 4660680 .
- ^ Чепмен, RE; Гальтон, Пе. М.; Сепкоски, Джей Джей; Уолл, WP (1981). «Морфометрическое исследование черепа пахицефалозавридного динозавра Stegoceras ». Журнал палеонтологии . 55 (3): 608–618. JSTOR 1304275 .
- ^ Jump up to: а б Шотт, РК; Эванс, округ Колумбия (2012). «Чешуйчатый онтогенез и вариации пахицефалозаврового динозавра Stegoceras validum Lambe, 1902, из формации Парк динозавров, Альберта». Журнал палеонтологии позвоночных . 32 (4): 903–913. Бибкод : 2012JVPal..32..903S . дои : 10.1080/02724634.2012.679878 . JSTOR 23251281 . S2CID 84526252 .
- ^ Jump up to: а б Петерсон, Дж. Э.; Дишлер, К.; Лонгрич, Северная Каролина; Додсон, П. (2013). «Распределение черепных патологий доказывает, что у куполообразноголовых динозавров (Pachycephalosauridae) бьются головой» . ПЛОС ОДИН . 8 (7): е68620. Бибкод : 2013PLoSO...868620P . дои : 10.1371/journal.pone.0068620 . ПМЦ 3712952 . ПМИД 23874691 .
- ^ Jump up to: а б с Гудвин, МБ; Хорнер, младший (2004). «Краниальная гистология пахицефалозавров (Ornithischia: Marginocephalia) выявляет временные структуры, несовместимые с поведением бодания головой» (PDF) . Палеобиология . 30 (2): 253–267. Бибкод : 2004Pbio...30..253G . doi : 10.1666/0094-8373(2004)030<0253:chopom>2.0.co;2 . JSTOR 4096846 . S2CID 84961066 .
- ^ Гальтон, премьер-министр (1971). «Примитивный купологоловый динозавр (Ornithischia: Pachycephalosauridae) из нижнего мела Англии и функция купола пахицефалозаврид». Журнал палеонтологии . 45 (1): 40–47. JSTOR 1302750 .
- ^ Суэц, HD (1978). «Функциональная морфология купола динозавров-пахицефалозаврид». Новый ежегодник по геологии и палеонтологии - ежемесячные выпуски . 8 : 459–472.
- ^ Снайвли, Э.; Кокс, А. (2008). «Структурная механика черепа пахицефалозавра допускает бодание головой» . Электронная палеонтология . 11 : 1–17.
- ^ Снайвли, Э.; Теодор, Дж. М. (2011). «Общие функциональные корреляты поведения при ударе головой у пахицефалозавра Stegoceras validum (Ornithischia, Dinosauria) и боевых парнокопытных» . ПЛОС ОДИН . 6 (6): e21422. Бибкод : 2011PLoSO...621422S . дои : 10.1371/journal.pone.0021422 . ПМК 3125168 . ПМИД 21738658 .
- ^ Мур, Брайан Р.С.; Ролосон, Мэтью Дж.; Карри, Филип Дж.; Райан, Майкл Дж.; Паттерсон, Р. Тимоти; Мэллон, Джордан К. (2022). «Аппендикулярная миология Stegoceras validum (Ornithischia: Pachycephalosauridae) и значение гипотезы удара головой» . ПЛОС ОДИН . 17 (9): e0268144. Бибкод : 2022PLoSO..1768144M . дои : 10.1371/journal.pone.0268144 . ПМЦ 9436104 . ПМИД 36048811 .
- ^ Падиан, К.; Хорнер, младший (2011). «Эволюция «причудливых структур» у динозавров: биомеханика, половой отбор, социальный отбор или распознавание видов?». Журнал зоологии . 283 (1): 3–17. дои : 10.1111/j.1469-7998.2010.00719.x .
- ^ Нелл, Р.Дж.; Сэмпсон, С. (январь 2011 г.). «Причудливые структуры динозавров: распознавание видов или половой отбор? Ответ Падиану и Хорнеру: причудливые структуры динозавров» . Журнал зоологии . 283 (1): 18–22. дои : 10.1111/j.1469-7998.2010.00758.x .
- ^ Хон, DWE; Нэйш, Д. (2013). «Гипотеза распознавания видов» не объясняет наличие и эволюцию преувеличенных структур у нептичьих динозавров» . Журнал зоологии . 290 (3): 172–180. дои : 10.1111/jzo.12035 .
- ^ Хорнер, Джон Р.; Гудвин, Марк Б.; Эванс, Дэвид К. (2023). «Новый пахицефалозавр из формации Хелл-Крик, округ Гарфилд, Монтана, США». Журнал палеонтологии позвоночных . 42 (4). дои : 10.1080/02724634.2023.2190369 .
- ^ Мэллон, Джей Си; Эванс, округ Колумбия (2014). «Тафономия и предпочтения среды обитания североамериканских пахицефалозаврид (Dinosauria, Ornithischia)». Летайя . 47 (4): 567–578. Бибкод : 2014Лета..47..567М . дои : 10.1111/лет.12082 .
- ^ Эберт, Дэвид А. (2005). «Геология» . В Карри, Филип Дж.; Коппельхус, Ева Бундгаард (ред.). Провинциальный парк динозавров . Издательство Университета Индианы. стр. 54–82 . ISBN 978-0-253-34595-0 .
- ^ Браман, Деннис Р.; Коппельхус, Ева Б. (2005). «Кампанские палиноморфы » В Карри, Филип Дж.; Коппельхус, Ева Бундгаард (ред.). Провинциальный парк динозавров . Издательство Университета Индианы. стр. 100-1 101–30 . ISBN 978-0-253-34595-0 .
- ^ Вейшампель, Д.Б.; Барретт, премьер-министр; Кориа, РА; Ле Лефф, Ж.; Сюй Син; ЗХ; Сахни, А.; Член парламента Гомани, Элизабет; Ното, ЧР (2004). «Распространение динозавров» В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. стр. 100-1 517 –606 . ISBN 978-0-520-24209-8 .
- ^ Эберт, Д.А. (1997). «Джудит Ривер Ведж». В Карри, Филип Дж .; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 199–204 . ISBN 978-0-12-226810-6 .
Внешние ссылки
[ редактировать ] СМИ, связанные со Стегоцерасом, на Викискладе?
СМИ, связанные со Стегоцерасом, на Викискладе?  Данные, относящиеся к Stegoceras, в Wikispecies
Данные, относящиеся к Stegoceras, в Wikispecies
