Обонятельная луковица
| Обонятельная луковица | |
|---|---|
 Человеческий мозг, вид снизу. Vesalius ' Fabrica , 1543. Обонятельные луковицы и обонятельные тракты обведены красным. | |
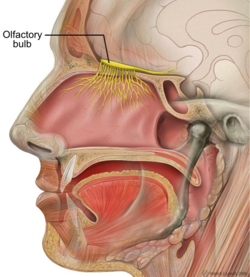 Сагиттальный разрез головы человека. | |
| Подробности | |
| Система | обонятельный |
| Идентификаторы | |
| латинский | обонятельная луковица |
| МеШ | D009830 |
| Нейроимена | 279 |
| НейроЛекс ID | бирнлекс_1137 |
| ТА98 | А14.1.09.429 |
| ТА2 | 5538 |
| ФМА | 77624 |
| Анатомические термины нейроанатомии | |

Синий – клубочковый слой;
Красный плексиформных и митральных клеток ; – внешний слой
Зеленый – внутренний слой плексиформных и гранулярных клеток .
Верхняя часть изображения — вид сверху , правая часть изображения — вид сбоку . Чешуя от вентральной до спинной части составляет примерно 2 мм.
( Обонятельная луковица лат . Bulbus olfactorius ) представляет собой структуру позвоночных, участвующую переднего мозга в обонянии , обонянии нервную . Он отправляет обонятельную информацию для дальнейшей обработки в миндалевидное тело , орбитофронтальную кору (ОФК) и гиппокамп , где он играет роль в эмоциях, памяти и обучении. Луковица разделена на две отдельные структуры: главную обонятельную луковицу и добавочную обонятельную луковицу. Основная обонятельная луковица соединяется с миндалевидным телом через грушевидную кору первичной обонятельной коры и напрямую выступает из главной обонятельной луковицы в определенные области миндалевидного тела. Дополнительная обонятельная луковица расположена в дорсально-задней области основной обонятельной луковицы и образует параллельный путь. Разрушение обонятельной луковицы приводит к ипсилатеральной аносмии , а ирритативные поражения ункуса могут привести к обонятельным и вкусовым галлюцинациям.

Структура
[ редактировать ]У большинства позвоночных обонятельная луковица является самой ростральной (передней) частью мозга, как это видно у крыс. Однако у людей обонятельная луковица находится на нижней (нижней) стороне мозга. Обонятельная луковица поддерживается и защищена решетчатой пластинкой решетчатой кости , которая у млекопитающих отделяет ее от обонятельного эпителия и перфорирована аксонами обонятельного нерва . Луковица разделена на две отдельные структуры: главную обонятельную луковицу и добавочную обонятельную луковицу.
Слои
[ редактировать ]Основная обонятельная луковица имеет многослойную клеточную архитектуру . В порядке от поверхности к центру слои располагаются:
- Клубочковый слой
- Внешний плексиформный слой
- митральных клеток Слой
- Внутренний плексиформный слой
- Слой гранулярных клеток
Обонятельная луковица передает информацию о запахах от носа к мозгу и, таким образом, необходима для правильного обоняния. Являясь нейронной цепью , слой клубочка получает прямую информацию от афферентных нервов , состоящих из аксонов примерно десяти миллионов нейронов обонятельных рецепторов обонятельной слизистой оболочки , области полости носа . Концы аксонов группируются в сферические структуры, известные как клубочки, так что каждый клубочек получает входные данные в первую очередь от нейронов обонятельных рецепторов, которые экспрессируют один и тот же обонятельный рецептор . Слой клубочков обонятельной луковицы представляет собой первый уровень синаптической обработки . [ 1 ] Слой клубочков представляет собой пространственную карту запахов, организованную по химической структуре пахучих веществ, такой как функциональные группы и длина углеродной цепи. Эта пространственная карта разделена на зоны и кластеры, которые представляют собой схожие клубочки и, следовательно, схожие запахи. Одна группа, в частности, связана с неприятными, испорченными запахами, которые представлены определенными химическими характеристиками. Эта классификация может быть эволюционной, чтобы помочь определить пищу, которую больше нельзя есть.
Пространственная карта слоя клубочков может быть использована для восприятия запаха в обонятельной коре. [ 2 ] Следующий уровень синаптической обработки в обонятельной луковице происходит во внешнем плексиформном слое, между слоем клубочков и слоем митральных клеток. Внешний плексиформный слой содержит астроциты , интернейроны и некоторые митральные клетки. Он содержит не много клеточных тел , а в основном дендриты митральных клеток и ГАМКергические гранулярные клетки. [ 3 ] также пронизаны дендритами нейронов, называемыми митральными клетками , которые, в свою очередь, выводятся в обонятельную кору . В обонятельной луковице существуют многочисленные типы интернейронов, включая перигломерулярные клетки , которые образуют синапсы внутри клубочков и между ними, и гранулярные клетки, которые образуют синапсы с митральными клетками. Слой гранулярных клеток — самый глубокий слой обонятельной луковицы. Он состоит из дендродендритных гранулярных клеток, которые образуют синапсы со слоем митральных клеток. [ 4 ]
Функция
[ редактировать ]Эта часть мозга воспринимает запахи. Как нервная цепь, обонятельная луковица имеет один источник сенсорной входной информации (аксоны обонятельных рецепторных нейронов обонятельного эпителия) и один выходной сигнал (аксоны митральных клеток). В результате обычно предполагается, что он функционирует как фильтр , в отличие от ассоциативной схемы, имеющей множество входов и множество выходов. Однако обонятельная луковица также получает информацию «сверху вниз» от таких областей мозга, как обонятельная кора , миндалевидное тело , неокортекс , гиппокамп , голубое пятно и черная субстанция . [ 5 ] Его потенциальные функции можно разделить на четыре неисключительные категории: [ нужна ссылка ]
- различать запахи
- повышение чувствительности обнаружения запаха
- фильтрация многих фоновых запахов для улучшения передачи некоторых избранных запахов
- позволяя высшим областям мозга, участвующим в возбуждении и внимании, изменять обнаружение или различение запахов.
Хотя все эти функции теоретически могут возникнуть из-за схемы схемы обонятельной луковицы, неясно, какие из этих функций выполняются исключительно обонятельной луковицей. По аналогии с аналогичными частями мозга, такими как сетчатка , многие исследователи сосредоточились на том, как обонятельная луковица фильтрует поступающую информацию от рецепторных нейронов в пространстве или как она фильтрует поступающую информацию во времени. В основе этих предложенных фильтров лежат два класса интернейронов; перигломерулярные клетки и гранулярные клетки. Обработка происходит на каждом уровне основной обонятельной луковицы, начиная с пространственных карт, которые классифицируют запахи в слое клубочков. [ 2 ]
Интернейроны внешнего плексиформного слоя реагируют на пресинаптические потенциалы действия и проявляют как возбуждающие постсинаптические потенциалы , так и тормозные постсинаптические потенциалы . Нейронная активация варьируется во времени: существуют периоды быстрой, спонтанной активации и медленной модуляции активации. Эти закономерности могут быть связаны с обнюхиванием или изменением интенсивности и концентрации запаха. [ 3 ] Временные закономерности могут влиять на последующую обработку пространственного восприятия запаха. [ нужна ссылка ] Например, синхронизированные серии спайков митральных клеток, по-видимому, помогают различать похожие запахи лучше, чем когда эти серии спайков не синхронизированы. [ 6 ] Известная модель [ 7 ] заключается в том, что бульбарная нервная цепь преобразует информацию о запахе в рецепторах в популяционный паттерн нейронной колебательной активности. [ 8 ] в популяции митральных клеток, [ 7 ] и этот паттерн затем распознается ассоциативными воспоминаниями обонятельных объектов в обонятельной коре. [ 9 ] [ 10 ] Обратная связь сверху вниз от обонятельной коры к обонятельной луковице модулирует бульбарные реакции, так что, например, луковица может адаптироваться к уже существующему обонятельному фону, чтобы выделить запах переднего плана из смеси запахов для распознавания. [ 10 ] [ 11 ] или может повысить чувствительность к целевому запаху во время поиска запаха. [ 12 ] [ 10 ] Разрушение обонятельной луковицы приводит к ипсилатеральной аносмии тогда как ирритативное поражение ункуса , может привести к обонятельным и вкусовым галлюцинациям. [ нужна ссылка ]
Латеральное торможение
[ редактировать ]- Внешний плексиформный слой
Интернейроны во внешнем плексиформном слое осуществляют торможение обратной связи на митральных клетках, чтобы контролировать обратное распространение . Они также участвуют в латеральном торможении митральных клеток. Это торможение является важной частью обоняния, поскольку оно помогает распознавать запахи, уменьшая возбуждение в ответ на фоновые запахи и дифференцируя реакции обонятельных нервов в слое митральных клеток. [ 1 ] Подавление слоя митральных клеток другими слоями способствует распознаванию запахов и обработке более высокого уровня путем модуляции выходного сигнала обонятельной луковицы. Эти гиперполяризации во время стимуляции запахом формируют реакцию митральных клеток, делая их более специфичными к запаху. [ 4 ]
Отсутствует информация о функции внутреннего плексиформного слоя, который лежит между слоем митральных клеток и слоем гранулярных клеток. [ нужна ссылка ]
- Слой гранулярных клеток
Базальные дендриты митральных клеток соединены с интернейронами, известными как гранулярные клетки , которые, согласно некоторым теориям, вызывают латеральное торможение между митральными клетками. Синапс между митральными и гранулярными клетками относится к редкому классу синапсов, которые являются «дендродендритными», что означает, что обе стороны синапса представляют собой дендриты, высвобождающие нейромедиатор. В этом конкретном случае митральные клетки выделяют возбуждающий нейромедиатор глутамат , а гранулярные клетки выделяют тормозной нейромедиатор гамма-аминомасляную кислоту (ГАМК). В результате своей двунаправленности дендро-дендритный синапс может заставлять митральные клетки тормозить себя (аутоингибирование), а также соседних митральных клеток (латеральное торможение). Более конкретно, слой гранулярных клеток получает возбуждающие сигналы глутамата от базальных дендритов митральных и тафтинговых клеток. Гранулярная клетка, в свою очередь, высвобождает ГАМК, оказывая ингибирующее действие на митральную клетку. Больше нейротрансмиттера высвобождается из активированной митральной клетки в связанный с ней дендрит гранулярной клетки, что делает ингибирующий эффект гранулярной клетки на активированную митральную клетку сильнее, чем на окружающие митральные клетки. [ 4 ] Неясно, какова будет функциональная роль латерального торможения, хотя оно может быть задействовано в повышении соотношения сигнал/шум сигналов запаха за счет подавления базовой частоты импульсов окружающих неактивированных нейронов. Это, в свою очередь, помогает различать запахи. [ 1 ] Другие исследования показывают, что латеральное торможение способствует дифференцированной реакции на запахи, что помогает в обработке и восприятии различных запахов. [ 4 ] Имеются также данные о холинергических эффектах на гранулярные клетки, которые усиливают деполяризацию гранулярных клеток, делая их более возбудимыми, что, в свою очередь, усиливает торможение митральных клеток. Это может способствовать более специфическому выходному сигналу обонятельной луковицы, который будет больше напоминать карту запахов клубочков. [ 13 ] [ 14 ] Обоняние отличается от других сенсорных систем , где периферические сенсорные рецепторы имеют реле в промежуточном мозге . играет обонятельная луковица Следовательно, такую роль для обонятельной системы .
Дополнительная обонятельная лампочка
[ редактировать ]У позвоночных добавочная обонятельная луковица (АОБ), расположенная в дорсально-задней области основной обонятельной луковицы, образует параллельный путь, независимый от основной обонятельной луковицы. Вомероназальный орган посылает проекции в добавочную обонятельную луковицу. [ 15 ] [ 16 ] что делает его вторым этапом обработки дополнительной обонятельной системы . Как и в основной обонятельной луковице, вход аксонов в добавочную обонятельную луковицу образует синапсы с митральными клетками внутри клубочков. Дополнительная обонятельная луковица получает аксональную информацию от вомероназального органа , отдельного сенсорного эпителия от основного обонятельного эпителия , который обнаруживает химические стимулы, важные для социального и репродуктивного поведения, а также, вероятно, общие запахи. [ 17 ] Было высказано предположение, что для включения вомерназальной помпы основной обонятельный эпителий должен сначала обнаружить соответствующий запах. [ 18 ] Однако возможность того, что вомероназальная система работает параллельно или независимо от общих обонятельных сигналов, еще не исключена.
Вомероназальные сенсорные нейроны обеспечивают прямые возбуждающие сигналы к основным нейронам АОБ, называемым митральными клетками. [ 19 ] которые передаются в миндалевидное тело и гипоталамус и, следовательно, непосредственно участвуют в активности половых гормонов и могут влиять на агрессивность и брачное поведение. [ 20 ] Аксоны вомероназальных сенсорных нейронов экспрессируют определенный тип рецепторов, которые, в отличие от того, что происходит в главной обонятельной луковице, расходятся между 6 и 30 клубочками AOB. Дендритные окончания митральных клеток проходят через драматический период нацеливания и кластеризации сразу после пресинаптического объединения аксонов сенсорных нейронов. Связь нейронов вомерназальной сенсорной клубочки с митральными клетками точная, при этом дендриты митральных клеток нацелены на клубочки . [ 19 ] Есть данные против наличия функциональной дополнительной обонятельной луковицы у человека и других высших приматов. [ 21 ]
AOB разделен на две основные субрегионы, переднюю и заднюю, которые получают отдельные синаптические входы от двух основных категорий вомероназальных сенсорных нейронов, V1R и V2R соответственно. Это выглядит как четкая функциональная специализация, учитывая различную роль двух популяций сенсорных нейронов в обнаружении химических стимулов различного типа и молекулярной массы. Хотя, по-видимому, он не поддерживается централизованно, где сходятся проекции митральных клеток с обеих сторон АОБ. Явным отличием схемы АОБ от остальной части луковицы является ее гетерогенная связь между митральными клетками и вомероназальными сенсорными афферентами внутри нейропильных клубочков. Митральные клетки AOB действительно контактируют через апикальные дендритные отростки клубочков, образованные афферентами различных рецепторных нейронов, тем самым нарушая правило «один рецептор-один нейрон», которое обычно справедливо для основной обонятельной системы. Это означает, что стимулы, воспринимаемые через VNO и вырабатываемые в AOB, подвергаются другому и, вероятно, более сложному уровню обработки. Соответственно, митральные клетки AOB демонстрируют явно разные паттерны возбуждения по сравнению с другими нейронами бульбарных проекций. [ 22 ] Кроме того, входной сигнал, поступающий сверху вниз в обонятельную луковицу, по-разному влияет на обонятельные выходы. [ 23 ]
Дальнейшая обработка
[ редактировать ]Обонятельная луковица отправляет обонятельную информацию для дальнейшей обработки в миндалину , орбитофронтальную кору (ОФК) и гиппокамп , где она играет роль в эмоциях, памяти и обучении. Основная обонятельная луковица соединяется с миндалевидным телом через грушевидную кору первичной обонятельной коры и напрямую выступает из главной обонятельной луковицы в определенные области миндалевидного тела. [ 24 ] Миндалевидное тело передает обонятельную информацию в гиппокамп . Орбитофронтальная кора, миндалевидное тело, гиппокамп, таламус и обонятельная луковица имеют множество взаимосвязей, прямо и опосредованно через коры первичной обонятельной коры. Эти связи указывают на связь между обонятельной луковицей и высшими областями обработки информации, особенно теми, которые связаны с эмоциями и памятью. [ 24 ]
Миндалевидное тело
[ редактировать ]Ассоциативное обучение между запахами и поведенческими реакциями происходит в миндалевидном теле . Запахи служат подкреплением или наказанием в процессе ассоциативного обучения; запахи, возникающие при положительном состоянии, усиливают поведение, которое привело к положительному состоянию, тогда как запахи, возникающие при отрицательном состоянии, действуют наоборот. Сигналы запаха кодируются нейронами миндалевидного тела с поведенческим эффектом или эмоциями, которые они вызывают. Таким образом, запахи отражают определенные эмоции или физиологические состояния. [ 25 ] Запахи начинают ассоциироваться с приятными и неприятными реакциями, и в конечном итоге запах становится сигналом и может вызвать эмоциональную реакцию. Эти ассоциации запахов способствуют возникновению эмоциональных состояний, таких как страх. Снимки мозга показывают, что активация миндалевидного тела коррелирует с приятными и неприятными запахами, что отражает связь между запахами и эмоциями. [ 25 ]
Гиппокамп
[ редактировать ]Гиппокамп . также способствует обонятельной памяти и обучению В гиппокампе происходит несколько процессов обонятельной памяти. Подобно процессу в миндалевидном теле, запах связан с определенным вознаграждением, т. е. запахом пищи с получением средств к существованию. [ 26 ] Запахи в гиппокампе также способствуют формированию эпизодической памяти ; воспоминания о событиях в определенном месте или времени. Время, в которое активируются определенные нейроны в гиппокампе, связывается нейронами со стимулом, например с запахом. Представление запаха в разное время может вызвать воспроизведение воспоминаний, поэтому запах помогает вспомнить эпизодические воспоминания. [ 26 ]
Обонятельное кодирование у Хабенулы
[ редактировать ]У низших позвоночных (миноги и костистые рыбы) аксоны митральных клеток (главных обонятельных нейронов) асимметрично отходят исключительно в правое полушарие Habenula. Сообщается, что дорсальные Habenula (Hb) функционально асимметричны с преобладанием запаховых реакций в правом полушарии. Было также показано, что нейроны Hb спонтанно активны даже в отсутствие обонятельной стимуляции. Эти спонтанно активные нейроны Hb организованы в функциональные кластеры, которые, как предполагается, управляют обонятельными реакциями. (Джетти, СК и др., 2014, Current Biology)
- Модели депрессии
Дальнейшие доказательства связи между обонятельной луковицей, эмоциями и памятью продемонстрированы на моделях депрессии у животных . Удаление обонятельной луковицы у крыс эффективно вызывает структурные изменения в миндалевидном теле и гиппокампе, а также поведенческие изменения, аналогичные таковым у человека с депрессией. Исследователи используют крыс с обонятельной бульбэктомией для исследования антидепрессантов. [ 27 ] Исследования показали, что удаление обонятельной луковицы у крыс приводит к реорганизации дендритов , нарушению роста клеток в гиппокампе и снижению нейропластичности в гиппокампе. Эти изменения гиппокампа из-за удаления обонятельной луковицы связаны с поведенческими изменениями, характерными для депрессии, что демонстрирует корреляцию между обонятельной луковицей и эмоциями. [ 28 ] Гиппокамп и миндалевидное тело влияют на восприятие запахов. Во время определенных физиологических состояний, таких как голод, запах еды может казаться более приятным и полезным из-за ассоциаций в миндалевидном теле и гиппокампе стимула запаха еды с вознаграждением за еду. [ 25 ]
Орбитофронтальная кора
[ редактировать ]Обонятельная информация отправляется в первичную обонятельную кору, откуда проекции отправляются в орбитофронтальную кору . OFC вносит свой вклад в связь запаха и вознаграждения, а также оценивает ценность вознаграждения, то есть пищевую ценность пищи. OFC получает проекции из грушевидной коры , миндалевидного тела и парагиппокампальной коры. [ 25 ] Нейроны OFC, которые кодируют информацию о пищевом вознаграждении, при стимуляции активируют систему вознаграждения, связывая процесс еды с вознаграждением. OFC далее проецируется на переднюю поясную извилину , где играет роль в формировании аппетита. [ 29 ] OFC также связывает запахи с другими стимулами, такими как вкус. [ 25 ] Восприятие и распознавание запахов также связаны с OFC. Пространственная карта запахов в слое клубочков обонятельной луковицы может способствовать этим функциям. Карта запахов начинает обработку обонятельной информации путем пространственной организации клубочков. Эта организация помогает обонятельной коре выполнять функции восприятия и различения запахов. [ 2 ]
Взрослый нейрогенез
[ редактировать ]Обонятельная луковица, наряду с субвентрикулярной зоной и субгранулярной зоной зубчатой извилины гиппокампа, является одной из трех структур головного мозга, которые, как наблюдалось, подвергаются продолжающемуся нейрогенезу у взрослых млекопитающих. [ нужна ссылка ] У большинства млекопитающих новые нейроны рождаются из нейральных стволовых клеток в субвентрикулярной зоне и мигрируют рострально по направлению к основному стволу. [ 30 ] и аксессуар [ 31 ] обонятельные луковицы. Внутри обонятельной луковицы эти незрелые нейробласты развиваются в полнофункциональные интернейроны гранулярных клеток и интернейроны перигломерулярных клеток, которые располагаются в слое гранулярных клеток и слоях клубочков соответственно. Аксоны обонятельных сенсорных нейронов, которые образуют синапсы в клубочках обонятельных луковиц, также способны к регенерации после повторного роста обонятельного сенсорного нейрона, расположенного в обонятельном эпителии. Несмотря на динамический оборот сенсорных аксонов и интернейронов, проекционные нейроны (митральные и тафтинговые нейроны), образующие синапсы с этими аксонами, структурно не пластичны. [ нужна ссылка ]
Функция взрослого нейрогенеза в этом регионе остается предметом изучения. Выживание незрелых нейронов при их входе в цепь очень чувствительно к обонятельной активности и, в частности, к задачам ассоциативного обучения. Это привело к гипотезе, что новые нейроны участвуют в процессах обучения. [ 32 ] Никакого определенного поведенческого эффекта не наблюдалось в экспериментах с потерей функции, что позволяет предположить, что функция этого процесса, если вообще связана с обонятельной обработкой, может быть незначительной. [ нужна ссылка ]
Клиническое значение
[ редактировать ]Обонятельная доля — это структура переднего мозга позвоночных, участвующая в обонянии или обонянии. Разрушение обонятельной луковицы приводит к ипсилатеральной аносмии .
Другие животные
[ редактировать ]Эволюция
[ редактировать ]
Сравнение структуры обонятельной луковицы у видов позвоночных, таких как леопардовая лягушка и лабораторная мышь , показывает, что все они имеют одну и ту же фундаментальную структуру (пять слоев, содержащих ядра трех основных типов клеток; подробности см. в разделе «Анатомия»). несмотря на то, что они непохожи по форме и размеру. Аналогичное строение имеет аналогичный обонятельный центр плодовой мухи Drosophila melanogaster — усиковая доля . Одна из возможностей заключается в том, что структура обонятельной луковицы позвоночных и усиков насекомых может быть схожей, поскольку они содержат оптимальное решение вычислительной проблемы, с которой сталкиваются все обонятельные системы, и, таким образом, могли развиваться независимо в разных типах - явление, обычно известное как конвергентная эволюция . [ 33 ] [ 34 ]
«Увеличение размера мозга по сравнению с размером тела — энцефализация — тесно связано с эволюцией человека. Однако две генетически разные эволюционные линии, неандертальцы и современные люди , произвели на свет человеческие виды с одинаковым большим мозгом. Таким образом, понимание эволюции человеческого мозга должно включать в себя исследование специфической реорганизации головного мозга, возможно, отражающееся в изменениях формы мозга. Здесь мы используем интеграцию развития между мозгом и лежащей в его основе скелетной основой, чтобы проверить гипотезы об эволюции мозга у Homo . Трехмерный геометрический морфометрический анализ эндобазикраниальной формы раскрывает ранее недокументированные детали эволюции. Изменения у Homo sapiens . Более крупные обонятельные луковицы, относительно широкая орбитофронтальная кора, относительно увеличенные и выступающие вперед полюса височных долей кажутся уникальными для современных людей. Такая реорганизация мозга, помимо физических последствий для общей формы черепа, могла способствовать эволюции H. sapiens. « обучение и социальные способности, в которых высшие обонятельные функции и их когнитивные и неврологические поведенческие последствия могли быть до сих пор недооцененными факторами». [ 35 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Гамильтон, Калифорния; Хейнбокель, Т.; Эннис, М.; Сабо, Г.; Эрдели, Ф.; Хаяр, А. (2005). «Свойства интернейронов внешнего плексиформного слоя в срезах обонятельной луковицы мыши» . Нейронаука . 133 (3): 819–829. doi : 10.1016/j.neuroscience.2005.03.008 . ISSN 0306-4522 . ПМК 2383877 . ПМИД 15896912 .
- ^ Jump up to: а б с Мори К., Такахаши Ю.К., Игараси К.М., Ямагути М. (апрель 2006 г.). «Карты молекулярных особенностей запахов в обонятельной луковице млекопитающих». Физиол. Преподобный . 86 (2): 409–33. doi : 10.1152/physrev.00021.2005 . ПМИД 16601265 .
- ^ Jump up to: а б Спорс, Х.; Албеану, DF; Мурти, В.Н.; Ринберг, Д.; Учида, Н.; Вачовяк, М.; Фридрих, RW (2012). «Освещение обонятельной обработки позвоночных» . Журнал неврологии . 32 (41): 14102–14108а. doi : 10.1523/JNEUROSCI.3328-12.2012 . ПМЦ 3752119 . ПМИД 23055479 .
- ^ Jump up to: а б с д Скотт Дж.В., Уэллис Д.П., Ригготт М.Дж., Буонвизо Н. (февраль 1993 г.). «Функциональная организация главной обонятельной луковицы». Микроск. Рез. Тех . 24 (2): 142–56. дои : 10.1002/jemt.1070240206 . ПМИД 8457726 . S2CID 6135866 .
- ^ Проф. Леон Зуравицкий (2 сентября 2010 г.). Нейромаркетинг: исследование мозга потребителя . Springer Science & Business Media. п. 22. ISBN 978-3-540-77828-8 . Проверено 4 июля 2015 г.
- ^ Линстер, Кристиана; Клеланд, Томас (17 июня 2013 г.). «Пространственно-временное кодирование в обонятельной системе». 20 лет вычислительной нейронауки . Том. 9. С. 229–242. дои : 10.1007/978-1-4614-1424-7_11 . ISBN 978-1-4614-1423-0 .
- ^ Jump up to: а б Ли, Чжаопин; Хопфилд, Джей-Джей (1 сентября 1989 г.). «Моделирование обонятельной луковицы и ее нейронных колебательных процессов» . Биологическая кибернетика . 61 (5): 379–392. дои : 10.1007/BF00200803 . ISSN 1432-0770 . ПМИД 2551392 . S2CID 7932310 .
- ^ Фриман, Уолтер Дж (1 мая 1978 г.). «Пространственные свойства ЭЭГ-событий в обонятельной луковице и коре головного мозга» . Электроэнцефалография и клиническая нейрофизиология . 44 (5): 586–605. дои : 10.1016/0013-4694(78)90126-8 . ISSN 0013-4694 . ПМИД 77765 .
- ^ Хаберли, Льюис Б.; Бауэр, Джеймс М. (1 января 1989 г.). «Обонятельная кора: модельная схема для изучения ассоциативной памяти?» . Тенденции в нейронауках . 12 (7): 258–264. дои : 10.1016/0166-2236(89)90025-8 . ISSN 0166-2236 . ПМИД 2475938 . S2CID 3947494 .
- ^ Jump up to: а б с Чжаопин, Ли (01 октября 2016 г.). «Распознавание, сегментация, адаптация, поиск цели и распознавание обонятельных объектов с помощью сети обонятельной луковицы и коры головного мозга: вычислительная модель и экспериментальные данные» . Современное мнение в области поведенческих наук . Компьютерное моделирование. 11 :30–39. дои : 10.1016/j.cobeha.2016.03.009 . ISSN 2352-1546 . S2CID 27989941 .
- ^ Фрэнк, Мэрион Э.; Флетчер, Дэйн Б.; Хеттингер, Томас П. (1 сентября 2017 г.). «Распознавание запахов компонентов в смесях» . Химические чувства . 42 (7): 537–546. doi : 10.1093/chemse/bjx031 . ISSN 1464-3553 . ПМЦ 5863551 . ПМИД 28641388 .
- ^ Стивенсон, Ричард Дж.; Кейс, Тревор И. (1 апреля 2005 г.). «Обонятельные образы: обзор» . Психономический бюллетень и обзор . 12 (2): 244–264. дои : 10.3758/BF03196369 . ISSN 1531-5320 . ПМИД 16082803 . S2CID 43129982 .
- ^ Пресслер, RT; Иноуэ, Т.; Строубридж, BW (2007). «Активация мускариновых рецепторов модулирует возбудимость гранулярных клеток и усиливает ингибирование митральных клеток в обонятельной луковице крысы» . Журнал неврологии . 27 (41): 10969–10981. doi : 10.1523/JNEUROSCI.2961-07.2007 . ПМК 6672850 . ПМИД 17928438 .
- ^ Смит, Р.С.; Ху, Р; ДеСуза, А; Эберли, CL; Крахе, К; Чан, В; Аранеда, RC (29 июля 2015 г.). «Дифференциальная мускариновая модуляция в обонятельной луковице» . Журнал неврологии . 35 (30): 10773–85. doi : 10.1523/JNEUROSCI.0099-15.2015 . ПМЦ 4518052 . ПМИД 26224860 .
- ^ Танигучи, К.; Сайто, С.; Танигучи, К. (февраль 2011 г.). «Филогенетический очерк обонятельной системы позвоночных» . J Vet Med Sci . 73 (2): 139–47. дои : 10.1292/jvms.10-0316 . ПМИД 20877153 .
- ^ Карлсон, Нил Р. (2013). Физиология поведения (11-е изд.). Бостон: Пирсон. п. 335. ИСБН 978-0205239399 .
- ^ Трин, К.; Шторм ДР. (2003). «Вомероназальный орган обнаруживает запахи при отсутствии передачи сигналов через основной обонятельный эпителий». Нат Нейроски . 6 (5): 519–25. дои : 10.1038/nn1039 . ПМИД 12665798 . S2CID 8289755 .
- ^ Слотник, Б.; Рестрепо, Д.; Шеллинк, Х.; Арчболд, Г.; Прайс, С.; Лин, В. (март 2010 г.). «Функция добавочной обонятельной луковицы модулируется сигналами основного обонятельного эпителия» . Eur J Neurosci . 31 (6): 1108–16. дои : 10.1111/j.1460-9568.2010.07141.x . ПМЦ 3745274 . ПМИД 20377623 .
- ^ Jump up to: а б Ховис, КР.; Рамнатх, Р.; Дален, Дж.Э.; Романова, Ал.; ЛаРокка, Г.; Бир, Мэн; Урбан, НН. (июнь 2012 г.). «Активность регулирует функциональные связи от вомероназального органа с добавочной обонятельной луковицей» . Дж. Нейроски . 32 (23): 7907–16. doi : 10.1523/JNEUROSCI.2399-11.2012 . ПМЦ 3483887 . ПМИД 22674266 .
- ^ Тротье, Д. (сентябрь 2011 г.). «Вомероназальный орган и феромоны человека» . Европейские анналы оториноларингологии, болезней головы и шеи . 128 (4): 184–90. дои : 10.1016/j.anorl.2010.11.008 . ПМИД 21377439 .
- ^ Бреннан П.А., Зуфал Ф. (ноябрь 2006 г.). «Феромональная коммуникация у позвоночных». Природа . 444 (7117): 308–15. Бибкод : 2006Natur.444..308B . дои : 10.1038/nature05404 . ПМИД 17108955 . S2CID 4431624 .
- ^ Шпак, Г.; Зильберталь, А.; Яром, Ю.; Вагнер, С. (2012). «Активируемые кальцием устойчивые огневые реакции отличают вспомогательные клетки от митральных клеток основной обонятельной луковицы» . Журнал неврологии . 32 (18): 6251–62. doi : 10.1523/JNEUROSCI.4397-11.2012 . ПМК 6622135 . ПМИД 22553031 .
- ^ Смит, Р.С.; Ху, Р; ДеСуза, А; Эберли, CL; Крахе, К; Чан, В; Аранеда, RC (29 июля 2015 г.). «Дифференциальная мускариновая модуляция в обонятельной луковице» . Журнал неврологии . 35 (30): 10773–85. doi : 10.1523/JNEUROSCI.0099-15.2015 . ПМЦ 4518052 . ПМИД 26224860 .
- ^ Jump up to: а б Ройе Дж. П., Плайи Дж. (октябрь 2004 г.). «Латерализация обонятельных процессов» . хим. Чувства . 29 (8): 731–45. дои : 10.1093/chemse/bjh067 . ПМИД 15466819 .
- ^ Jump up to: а б с д и Кадохиса М (2013). «Влияние запаха на эмоции с последствиями» . Передняя система нейронов . 7 : 66. дои : 10.3389/fnsys.2013.00066 . ПМЦ 3794443 . ПМИД 24124415 .
- ^ Jump up to: а б Rolls ET (декабрь 2010 г.). «Вычислительная теория формирования эпизодической памяти в гиппокампе». Поведение. Мозговой Рес . 215 (2): 180–96. дои : 10.1016/j.bbr.2010.03.027 . ПМИД 20307583 . S2CID 13605251 .
- ^ Песня, К.; Леонард, BE. (2005). «Обонятельная бульбэктомированная крыса как модель депрессии». Неврологические и биоповеденческие обзоры . 29 (4–5): 627–47. doi : 10.1016/j.neubiorev.2005.03.010 . ПМИД 15925697 . S2CID 42450349 .
- ^ Моралес-Медина, JC.; Хуарес, И.; Венансио-Гарсия, Э.; Кабрера, Сан-Франциско; Менар, К.; Ю, В.; Флорес, Г.; Мечавар, Н.; Кирион, Р. (апрель 2013 г.). «Нарушение структурной пластичности гиппокампа связано с эмоциональными нарушениями и нарушениями памяти у крыс, подвергшихся обонятельной бульбэктомии». Нейронаука . 236 : 233–43. doi : 10.1016/j.neuroscience.2013.01.037 . ПМИД 23357118 . S2CID 32020391 .
- ^ Роллс, ET (ноябрь 2012 г.). «Вкус, обоняние и текстура пищи вознаграждают мозг за обработку и контроль аппетита» . Труды Общества питания . 71 (4): 488–501. дои : 10.1017/S0029665112000821 . ПМИД 22989943 .
- ^ Лазарини, Ф.; Лледо, премьер-министр. (январь 2011 г.). «Необходим ли нейрогенез взрослых для обоняния?» (PDF) . Тенденции в нейронауках . 34 (1): 20–30. дои : 10.1016/j.tins.2010.09.006 . ПМИД 20980064 . S2CID 40745757 .
- ^ Оботи, Л; Савалли Дж; Джакино С; Де Марчис С; Панзика ГК; Фазоло А; Перетто П. (2009). «Интеграция и зависящее от сенсорного опыта выживание вновь созданных нейронов в добавочной обонятельной луковице самок мышей». Eur J Neurosci . 29 (4): 679–92. дои : 10.1111/j.1460-9568.2009.06614.x . ПМИД 19200078 . S2CID 36990333 .
- ^ Лепусез, Г.; Вэлли, Монтана; Лледо, премьер-министр. (2013). «Влияние нейрогенеза взрослых на цепи и вычисления обонятельных луковиц». Ежегодный обзор физиологии . 75 : 339–63. doi : 10.1146/annurev-psyol-030212-183731 . ПМИД 23190074 .
- ^ Аче, Б.В. (сентябрь 2010 г.). «Специфические для запаха способы передачи сигналов при обонянии млекопитающих» . Химические чувства . 35 (7): 533–9. doi : 10.1093/chemse/bjq045 . ПМЦ 2924424 . ПМИД 20519266 .
- ^ Ван, Дж.В. (январь 2012 г.). «Пресинаптическая модуляция ранней обонятельной обработки у дрозофилы» . Дев Нейробиол . 72 (1): 87–99. дои : 10.1002/днеу.20936 . ПМК 3246013 . ПМИД 21688402 .
- ^ Бастир, М.; Росас, А.; Гунц, П.; Пенья-Мелиан, А.; Манзи, Г.; Харвати, К.; Крушинский, Р.; Стрингер, К.; Хаблин, Дж.Дж. (2011). «Эволюция основания мозга у людей с высокой энцефализацией» (PDF) . Нат Коммун . 2 : 588. Бибкод : 2011NatCo...2..588B . дои : 10.1038/ncomms1593 . ПМИД 22158443 .
Дальнейшее чтение
[ редактировать ]- Шепард, Г. Синаптическая организация мозга , Oxford University Press, 5-е издание (ноябрь 2003 г.). ISBN 0-19-515956-X
- Халперн, М; Мартинес-Маркос, А (2003). «Структура и функции вомероназальной системы: обновленная информация» (PDF) . Прогресс нейробиологии . 70 (3): 245–318. дои : 10.1016/S0301-0082(03)00103-5 . ПМИД 12951145 . S2CID 31122845 . Архивировано из оригинала (PDF) 7 ноября 2017 г.
- Аче, BW; Янг, Дж. М. (2005). «Обоняние: разнообразие видов, сохранившиеся принципы» . Нейрон . 48 (3): 417–30. дои : 10.1016/j.neuron.2005.10.022 . ПМИД 16269360 . S2CID 12078554 .
Внешние ссылки
[ редактировать ]- «Анатомическая схема: 13048.000-1» . Roche Lexicon – иллюстрированный навигатор . Эльзевир. Архивировано из оригинала 07.11.2014.