Обонятельный бугорок
| Обонятельный бугорок | |
|---|---|
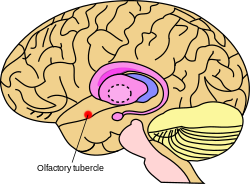 Примерное расположение обонятельного бугорка в головном мозге | |
| Подробности | |
| Часть | Мезолимбический путь Вентральное полосатое тело ; Обонятельная кора |
| Части | Медиальный бугорок Латеральный бугорок |
| Идентификаторы | |
| латинский | обонятельный бугорок |
| Акроним(ы) | OT |
| МеШ | D066208 |
| ТА98 | A14.1.09.433 |
| ТА2 | 5544 |
| Анатомическая терминология | |
Обонятельный бугорок ( ОТ ), также известный как обонятельный бугорок , представляет собой мультисенсорный центр обработки, который содержится в обонятельной коре и вентральном полосатом теле и играет роль в познании вознаграждения . Также было показано, что ОТ играет роль в локомоторном поведении и поведении внимания , особенно в отношении социальной и сенсорной реакции . [1] и это может быть необходимо для поведенческой гибкости. [2] ОТ взаимосвязан с многочисленными областями мозга, особенно с сенсорными центрами, центрами возбуждения и вознаграждения, что делает его потенциально важным интерфейсом между обработкой сенсорной информации и последующими поведенческими реакциями. [3]
ОТ представляет собой сложную структуру, которая получает прямую информацию от обонятельной луковицы и содержит морфологические и гистохимические характеристики вентрального паллидума и полосатого тела переднего мозга. [4] Дофаминергические нейроны мезолимбического пути проецируются на ГАМКергической шиповидные нейроны и среды прилежащего ядра обонятельного бугорка. [5] ( рецептор D3 в изобилии присутствует в этих двух областях [6] ). Кроме того, ОТ содержит плотно упакованные скопления клеток, известные как островки Каллеха , которые состоят из гранулярных клеток . Несмотря на то, что он является частью обонятельной коры и получает прямую информацию от обонятельной луковицы , не было доказано, что он играет роль в обработке запахов.
Структура
[ редактировать ]Обонятельный бугорок различается по расположению и относительному размеру у людей, других приматов, грызунов, птиц и других животных. В большинстве случаев обонятельный бугорок определяется как округлая выпуклость вдоль основания переднего мозга впереди зрительного перекреста и сзади от обонятельной ножки . [7] У человека и других приматов визуальная идентификация обонятельного бугорка затруднена, поскольку у этих животных базальная выпуклость переднего мозга невелика. [8] Что касается функциональной анатомии , обонятельный бугорок можно рассматривать как часть трех более крупных сетей. Во-первых, его считают частью базального переднего мозга, прилежащего ядра и миндалевидных ядер из-за его расположения вдоль рострально-вентральной области мозга, то есть передне-нижней части. Во-вторых, он считается частью обонятельной коры, поскольку получает прямую информацию от обонятельной луковицы . он также считается частью вентрального полосатого тела В-третьих, на основании анатомических, нейрохимических и эмбриологических данных .
Одной из наиболее ярких особенностей обонятельного бугорка являются плотно упакованные скопления клеток серповидной формы, которые расположены преимущественно в слое III, а иногда и в слое II. Эти скопления клеток, называемые островками Каллея , иннервируются дофаминергическими проекциями прилежащего ядра и черной субстанции , что указывает на роль, которую обонятельный бугорок играет в системе вознаграждения .
Обонятельный бугорок является мультисенсорным обрабатывающим центром из-за множества иннерваций, идущих к другим областям мозга и от них, таких как миндалевидное тело , таламус , гипоталамус , гиппокамп , ствол мозга, слуховые и зрительные сенсорные волокна, а также ряд структур в систему вознаграждения-возбуждения, а также обонятельную кору. Благодаря многочисленным иннервациям из других областей мозга обонятельный бугорок участвует в объединении информации от органов чувств, таких как обонятельная/слуховая и обонятельная/зрительная интеграция, возможно, поведенчески значимым образом. Таким образом, повреждение обонятельного бугорка, скорее всего, повлияет на функциональность всех этих областей мозга. Примеры таких нарушений включают изменения в нормальном поведении, ориентированном на запахи, а также нарушения в модуляции состояния и мотивационном поведении. [3] которые часто встречаются при психических расстройствах, таких как шизофрения , [9] деменция [10] и депрессия . [11]
Было показано, что обонятельный бугорок играет большую роль в поведении. Было показано, что односторонние поражения обонятельного бугорка изменяют внимание, социальную и сенсорную реакцию и даже двигательное поведение. [2] Было показано, что двусторонние поражения снижают копулятивное поведение у крыс-самцов. Также было показано, что обонятельный бугорок особенно участвует в поведении, связанном с вознаграждением и привыканием. Было показано, что крысы вводят кокаин в обонятельный бугорок чаще, чем в прилежащее ядро и вентральное паллидум , другие центры вознаграждения в мозге. [12] Фактически, они будут вводить кокаин в обонятельный бугорок примерно 200 раз в час и даже до самой смерти.
Функциональный вклад обонятельного бугорка в обоняние в настоящее время неясен; однако есть свидетельства того, что он может играть перцептивную роль. Работа Зелано и др. предполагают, что обонятельный бугорок может иметь решающее значение в выяснении источников обонятельной информации . [13] Это говорит о том, что он также может играть роль в поведении, управляемом запахом. Таким образом, он может связывать восприятие запаха с действием посредством связей с системами внимания, вознаграждения и мотивации базального отдела переднего мозга . [3] Данные функциональной визуализации из этой же группы также показывают, что обонятельный бугорок сильно активируется во время задач, требующих внимания, тем самым играя большую роль в системах, связанных с возбуждением.
Поскольку обонятельный бугорок является компонентом вентрального полосатого тела , он тесно связан с несколькими центрами мозга, связанными с аффективностью, вознаграждением и мотивацией. Он также находится на стыке обонятельных сенсорных сигналов и зависимых от состояния поведенческих модуляторных цепей , то есть области, которая модулирует поведение во время определенных физиологических и психических состояний. Таким образом, обонятельный бугорок может также играть важную роль в опосредовании подхода к запаху и поведения избегания запаха, вероятно, в зависимости от состояния. [14]
Анатомия
[ редактировать ]Обычно обонятельный бугорок расположен в базальном отделе переднего мозга животного в медиальной височной доле. В частности, части бугорка включены в обонятельную кору и располагаются между зрительным перекрестом и обонятельным трактом и вентральнее прилежащего ядра . Обонятельный бугорок состоит из трех слоев: молекулярного слоя (слой I), слоя плотных клеток (слой II) и многообразного слоя (слой III). [8] За исключением островов Каллеха , характерных для бугорка, [15] он также известен тем, что иннервируется дофаминергическими нейронами из вентральной покрышки. Обонятельный бугорок также состоит из разнородных элементов, например медиального пучка переднего мозга, и имеет вентральное продолжение полосатого комплекса. В 1970-х годах было обнаружено, что бугорок содержит стриарный компонент , состоящий из ГАМКергических средних шиповатых нейронов . [5] ГАМКергические нейроны проецируются на вентральное паллидум и получают глутаматергические входы из корковых областей и дофаминергические входы из вентральной области покрышки. [16] [17]
Морфологические и нейрохимические особенности
[ редактировать ]Вентральная часть обонятельного бугорка состоит из трех слоев, тогда как дорсальная часть содержит плотные скопления клеток и примыкает к вентральному паллидуму (внутри базальных ганглиев). Строение наиболее вентральной и передней частей бугорка можно определить как анатомически очерченные холмики (состоящие из извилин и борозд) и скопления клеток.
Наиболее распространенными типами клеток обонятельного бугорка являются плотные шипиковые клетки среднего размера, встречающиеся преимущественно во II слое (плотный клеточный слой). Дендриты этих клеток покрыты аксонами иммунореактивного вещества p (SPI) до слоя III (мультиформного слоя). [8] Эти клетки также проецируются в прилежащее ядро и хвостатую скорлупу, связывая, таким образом, обонятельный бугорок с паллидумом. [18] Другие клетки среднего размера находятся также во II и III слоях обонятельного бугорка. К ним относятся нейроны с бедными шипами и веретенообразные клетки, и они отличаются от плотных клеток шипиков среднего размера тем, что имеют редкие дендритные деревья. Самые крупные клетки и наиболее яркая особенность обонятельного бугорка представляют собой плотно упакованные скопления клеток серповидной формы, островки Каллея, которые расположены в основном в дорсальной части обонятельного бугорка, слое III, а также могут быть обнаружены в слое II. Обонятельный бугорок также содержит три класса мелких клеток, встречающихся преимущественно в I и II слоях. Первыми являются пиальные клетки (названные так из-за расположения вблизи пиальной поверхности), которые выглядят как миниатюрные плотные шиповидные клетки среднего размера. Вторые представляют собой лучистые клетки и легко идентифицируются по многочисленным разнонаправленным бесшипистым дендритам. Третьи, маленькие шиповидные клетки, похожи на пиальные клетки тем, что они также выглядят как шиповидные клетки среднего размера, за исключением того, что они не расположены вблизи пиальной поверхности. [19]
Разработка
[ редактировать ]Мигрирующие клетки из нескольких мест развития собираются вместе, образуя обонятельный бугорок. Сюда входят вентральные ганглиозные возвышения (находятся в вентральной части конечного мозга , где они образуют выпуклости в желудочках , которые позже становятся базальными ганглиями, присутствующими только на эмбриональных стадиях ) и ростромедиальная стенка конечного мозга (переднего мозга). [20] Нейроны обонятельных бугорков возникают уже на 13-й день эмбриона (E13), и развитие клеток происходит послойно-специфическим образом. Возникновение трех основных слоев обонятельного бугорка начинается почти одновременно. Большие нейроны слоя III возникают от E13 до E16, а мелкие и средние — между E15 и E20. Подобно мелким и средним клеткам слоя III, клетки слоя II и полосатых мостиков также возникают между E15 и E20 и развиваются в латеральном к медиальному градиенту. [21] Зернистые клетки островков Calleja возникают между E19 и E22 и продолжают мигрировать в островки еще долгое время после рождения. [22] [23]
Волокна латерального обонятельного тракта начинают разветвляться в обонятельный бугорок в районе Е17. Латеральная часть обонятельного бугорка (которая примыкает к обонятельному тракту) получает самые плотные входные волокна, а медиальная часть - отростки легких волокон. [24] Это ветвление продолжается до завершения примерно к концу первой недели после рождения.
Функция
[ редактировать ]Мультисенсорные процессы
[ редактировать ]Обонятельный бугорок играет функциональную роль в мультисенсорной интеграции обонятельной информации с экстрамодальными органами чувств. Слуховая сенсорная информация может поступать в обонятельный бугорок через сети, включающие гиппокамп и вентральный паллидум , или непосредственно из обонятельной коры, что указывает на возможную роль обонятельного бугорка в обонятельно-слуховой сенсорной интеграции . [25] Было показано, что эта конвергенция вызывает восприятие звука, вызванное взаимодействием запаха и звука. Эта возможность была подтверждена работой [7] где обонятельный бугорок демонстрировал обонятельно-слуховую конвергенцию.
Ретинальные проекции также были обнаружены во втором слое обонятельного бугорка, что позволяет предположить, что он представляет собой область обонятельной и зрительной конвергенции. [26] Эти зрительные сенсорные волокна приходят от ганглиозных клеток сетчатки. Таким образом, обонятельный бугорок может играть роль в восприятии запахов при идентификации зрительного источника.
Что касается обоняния, данные некоторых исследований in vitro показывают, что единицы обонятельных бугорков обладают функциональной способностью нейронов других обонятельных центров обрабатывать запахи. Было высказано предположение, что обонятельный бугорок может иметь решающее значение в определении источника обонятельной информации и реагирует на вдыхание запаха, к которому обращаются внимание. [13]
Роль в поведении
[ редактировать ]Показано, что обонятельный бугорок отвечает главным образом за прием сенсорных импульсов от обонятельных рецепторов. [27] Из-за своих связей с такими областями, как миндалевидное тело и гиппокамп, обонятельный бугорок может играть роль в поведении. Крысы в значительной степени полагаются на обонятельную сенсорную информацию от обонятельных рецепторов при формировании поведенческого отношения. [28] Исследования показывают, что двустороннее поражение обонятельного бугорка значительно снижает стереотипное поведение. [29] [30] такие как копулятивное поведение у самцов крыс и снижение нюхания и жевания. [2] Эти стереотипные торможения могли быть вызваны удалением центральных нейрональных отростков, отличных от дофаминергических клеток в обонятельном бугорке. Было показано, что односторонние поражения изменяют внимание, социальную и сенсорную реакцию и даже двигательное поведение у крыс. [2]
Возбуждение и вознаграждение
[ редактировать ]Дофаминергические нейроны вентральной покрышки, иннервирующие обонятельный бугорок, позволяют бугорку играть роль в вознаграждении и возбуждении и, по-видимому, частично опосредуют подкрепление кокаина. [12] Было показано, что переднемедиальные части бугорка опосредуют некоторые полезные эффекты таких наркотиков, как кокаин и амфетамин. Это было показано в исследованиях, в которых крысы учатся самостоятельно вводить кокаин со значительно высокой скоростью в бугорок. Инъекции кокаина в бугорок вызывают у крыс устойчивое передвижение и вставание на ноги. [31]
Клиническое значение
[ редактировать ]Мультисенсорная природа обонятельного бугорка и множество иннерваций, которые он получает из других областей мозга, особенно прямой входной сигнал от обонятельной луковицы и иннервации из вентральной покрышки , делают возможным участие его в ряде психических расстройств , при которых обоняние и поражаются дофаминовые рецепторы. Многие исследования обнаружили снижение обонятельной чувствительности у пациентов с большими депрессивными расстройствами (БДР) , деменцией и шизофренией . Было показано, что у пациентов с БДР наблюдается уменьшение обонятельной луковицы и обонятельной коры головного мозга. [11] по сравнению с нормальными людьми. При деменции, особенно типа болезни Альцгеймера , поражаются обонятельная луковица, переднее обонятельное ядро и орбитофронтальная кора , все области мозга, отвечающие за обоняние. Дефициты, наблюдаемые при деменции, включают снижение порога обонятельной чувствительности, [32] [33] идентификация запаха [34] и память запахов. [10] У пациентов с шизофренией наблюдаются нарушения обонятельной дискриминации, которые не наблюдаются у пациентов с другими психическими расстройствами, не упомянутыми здесь. Рупп и др. [9] обнаружили, что у больных шизофренией снижены обонятельная чувствительность и способность к различению, а также способности к идентификации более высокого порядка. Как упоминалось ранее, обонятельный бугорок может участвовать в восприятии запахов благодаря сигналам, поступающим от луковицы, и, таким образом, в более широком смысле может играть роль в этих психических расстройствах.
История
[ редактировать ]Обонятельный бугорок был впервые описан Альбертом фон Кёлликером в 1896 году, изучавшим его на крысах. С тех пор было проведено несколько гистологических и гистохимических исследований; [2] [8] было сделано в этой области, чтобы идентифицировать его у других грызунов, кошек, людей, приматов и других видов. Подобные исследования были проведены несколькими авторами, чтобы определить клеточный состав и иннервацию других областей ОТ. За прошедшие годы использовалось несколько других методов, чтобы выяснить возможные функции и роль ОТ в мозге. Они начались с изучения поражений [2] [14] [29] и ранние электрофизиологические записи. [35] [36] Усовершенствования в технологии позволили теперь размещать несколько электродов в обонятельном бугорке и вести запись с анестезированных и даже бодрствующих животных, участвующих в поведенческих задачах. [7] [12] [31] [34]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хитт, Дж. К.; Брайон, DM; Модианос, Д.Т. (1973). «Влияние рострально-медиального пучка переднего мозга и поражений обонятельных бугорков на сексуальное поведение самцов крыс». Журнал сравнительной и физиологической психологии . 82 (1): 30–36. дои : 10.1037/h0033797 . PMID 4567890 .
- ^ а б с д и ж Кооб, Г.Ф.; Райли, С.Дж.; Смит, Южная Каролина; Роббинс, ТВ (1978). «Влияние 6-гидроксидофаминовых поражений прилежащего ядра перегородки и обонятельного бугорка на питание, двигательную активность и амфетаминовую анорексию у крыс». Журнал сравнительной и физиологической психологии . 92 (5): 917–927. дои : 10.1037/h0077542 . ПМИД 282297 .
- ^ а б с Вессон, Д.В.; Уилсон, Д.А. (2011). «Выявляем вклад обонятельного бугорка в обоняние: гедоника, сенсорная интеграция и многое другое?» . Неврологические и биоповеденческие обзоры . 35 (3): 655–668. doi : 10.1016/j.neubiorev.2010.08.004 . ПМК 3005978 . ПМИД 20800615 .
- ^ Хеймер, Л.; Уилсон, Р.Д. (1975). «Подкорковые проекции аллокортекса: сходство нейронных связей гиппокампа, грушевидной коры и неокортекса». В Сантини, Маурицио (ред.). Материалы симпозиума столетия Гольджи . Нью-Йорк: Рэйвен. стр. 177–193. ISBN 978-0911216806 .
- ^ а б Икемото С (2010). «Схема вознаграждения мозга за пределами мезолимбической дофаминовой системы: нейробиологическая теория» . Неврологические и биоповеденческие обзоры . 35 (2): 129–50. doi : 10.1016/j.neubiorev.2010.02.001 . ПМЦ 2894302 . ПМИД 20149820 .
Недавние исследования по внутричерепному самостоятельному введению нейрохимических веществ (лекарств) показали, что крысы учатся самостоятельно вводить различные лекарства в мезолимбические дофаминовые структуры — заднюю вентральную область покрышки, медиальное прилежащее ядро оболочки и медиальный обонятельный бугорок. ... В 1970-х годах было признано, что обонятельный бугорок содержит полосатый компонент, который заполнен ГАМКергическими средними шиповидными нейронами, получающими глутаматергические входы из кортикальных областей и дофаминергические входы из ВТА и проецирующимися на вентральное паллидум, как и прилежащее ядро.
Рисунок 3. Вентральное полосатое тело и самостоятельный прием амфетамина. - ^ Брантон, Лоуренс Л.; Хилал-Дандан, Ранда; Ноллманн, Бьорн К. (2018). Гудман и Гилман - Фармакологические основы терапии . Образование МакГроу Хилл. п. 329. ИСБН 978-1-25-958473-2 .
- ^ а б с Вессон, Д.В.; Уилсон, Д.А. (2010). «Обонятельные звуки: обонятельно-слуховое сенсорное схождение в обонятельном бугорке» . Журнал неврологии . 30 (8): 3013–3021. doi : 10.1523/JNEUROSCI.6003-09.2010 . ПМЦ 2846283 . ПМИД 20181598 .
- ^ а б с д Миллхаус, Огайо; Хеймер, Л. (1984). «Конфигурация клеток обонятельного бугорка крысы». Журнал сравнительной неврологии . 228 (4): 571–597. дои : 10.1002/cne.902280409 . ПМИД 6490970 . S2CID 9261393 .
- ^ а б Рупп, CI; Флейшхакер, WW; Кеммлер, Г.; Кремсер, К.; Билдер, Р.М.; Меччеряков С.; Сешко, PR (2005). «Обонятельные функции и объемные показатели орбитофронтальной и лимбической областей при шизофрении». Исследования шизофрении . 74 (2–3): 149–161. doi : 10.1016/j.schres.2004.07.010 . ПМИД 15721995 . S2CID 11026266 .
- ^ а б Мерфи, К.; Нордин, С.; Джинич, С. (1999). «Очень раннее снижение способности узнавать запахи при болезни Альцгеймера». Старение, нейропсихология и познание . 6 (3): 229–240. дои : 10.1076/anec.6.3.229.777 .
- ^ а б Негояс, С.; Крой, И.; Гербер, Дж.; Пушманн, С.; Петровский, К.; Йорашки, П.; Хаммел, Т. (2010). «Снижение объема обонятельной луковицы и обонятельной чувствительности у пациентов с острой большой депрессией». Нейронаука . 169 (1): 415–421. doi : 10.1016/j.neuroscience.2010.05.012 . ПМИД 20472036 . S2CID 207248484 .
- ^ а б с Икемото, С. (2003). «Участие обонятельного бугорка в вознаграждении за кокаин: исследования внутричерепного самостоятельного приема» . Журнал неврологии . 23 (28): 9305–9311. doi : 10.1523/JNEUROSCI.23-28-09305.2003 . ПМК 6740580 . ПМИД 14561857 .
- ^ а б Зелано, К.; Монтэг, Дж.; Джонсон, Б.; Хан, Р.; Собель, Н. (2007). «Диссоциированные представления раздражения и валентности в первичной обонятельной коре человека». Журнал нейрофизиологии . 97 (3): 1969–1976. дои : 10.1152/Jn.01122.2006 . ПМИД 17215504 .
- ^ а б Жерве, Г. (1979). «Односторонние поражения обонятельного бугорка, изменяющие общие эффекты возбуждения обонятельной луковицы крысы». Электроэнцефалография и клиническая нейрофизиология . 46 (6): 665–674. дои : 10.1016/0013-4694(79)90104-4 . ПМИД 87311 .
- ^ Каллеха, К. (1893). Обонятельная область мозга . Мадрид: Николас Мойя.
- ^ Мейер, Г.; Гонсалес-Эрнандес, Т.; Каррильо-Падилья, Ф.; Феррес-Торрес, Р. (1989). «Агрегации гранулярных клеток в базальном отделе переднего мозга (острова Каллеха): Гольджи и цитоархитектоническое исследование у различных млекопитающих, включая человека». Журнал сравнительной неврологии . 284 (3): 405–428. дои : 10.1002/cne.902840308 . ПМИД 2474005 . S2CID 29824764 .
- ^ Миллхаус, Огайо (1987). «Зернитчатые клетки обонятельного бугорка и вопрос об островах каллеи». Журнал сравнительной неврологии . 265 (1): 1–24. дои : 10.1002/cne.902650102 . ПМИД 3693600 . S2CID 21826194 .
- ^ Фэллон, Дж. Х. (июнь 1983 г.). «Комплекс островов Каллея базального переднего мозга II крысы: соединения клеток среднего и крупного размера». Бюллетень исследований мозга . 10 (6): 775–93. дои : 10.1016/0361-9230(83)90210-1 . ПМИД 6616269 . S2CID 4723010 .
- ^ Рыбак, CE.; Фэллон, Дж. Х. (март 1982 г.). «Комплекс базального переднего мозга крысы на острове Каллея. I. Световые и электронно-микроскопические наблюдения». Журнал сравнительной неврологии . 205 (3): 207–18. дои : 10.1002/cne.902050302 . ПМИД 7076893 . S2CID 44954144 .
- ^ Гарсиа-Морено, Ф.; Лопес-Маскарак, Л.; де Карлос, Дж.А. (июнь 2008 г.). «Ранняя телэнцефальная миграция, топографически сходящаяся в обонятельной коре» . Кора головного мозга . 18 (6): 1239–52. дои : 10.1093/cercor/bhm154 . hdl : 10261/12293 . ПМИД 17878174 .
- ^ Байер, SA. (1985). «Производство нейронов в гиппокампе и обонятельной луковице мозга взрослой крысы: добавление или замена?». Анналы Нью-Йоркской академии наук . 457 (1): 163–72. Бибкод : 1985NYASA.457..163B . дои : 10.1111/j.1749-6632.1985.tb20804.x . ПМИД 3868311 . S2CID 26832068 .
- ^ Бедард, А.; Левеск, М.; Бернье, П.Дж.; Родитель, А. (2002). «Ростральный миграционный поток у взрослых обезьян: вклад новых нейронов в обонятельный бугорок и участие антиапоптотического белка bcl-2». Европейский журнал неврологии . 16 (10): 1917–1924. дои : 10.1046/j.1460-9568.2002.02263.x . ПМИД 12453055 . S2CID 31096044 .
- ^ Де Марчис, С.; Фазоло, А.; Пуче, AC. (август 2004 г.). «Происходящие из субвентрикулярной зоны предшественники нейронов мигрируют в подкорковый передний мозг постнатальных мышей». Журнал сравнительной неврологии . 476 (3): 290–300. дои : 10.1002/cne.20217 . hdl : 2318/101068 . ПМИД 15269971 . S2CID 25911933 .
- ^ Швоб, Дж.Э.; Прайс, Дж.Л. (февраль 1984 г.). «Развитие аксональных связей в центральной обонятельной системе крыс». Журнал сравнительной неврологии . 223 (2): 177–202. дои : 10.1002/cne.902230204 . ПМИД 6200518 . S2CID 25870173 .
- ^ Дедвайлер, ЮАР; Фостер, TC; Хэмпсон, Р.Э. (1987). «Обработка сенсорной информации в гиппокампе». Критические обзоры CRC в клинической нейробиологии . 2 (4): 335–355. ПМИД 3297494 .
- ^ Мик, Г.; Купер, Х.; Маньен, М. (1993). «Проекция сетчатки на обонятельный бугорок и базальный конечный мозг у приматов». Журнал сравнительной неврологии . 327 (2): 205–219. дои : 10.1002/cne.903270204 . ПМИД 8425942 . S2CID 21784363 .
- ^ Эйди, WR (1959). «ГЛАВА XXI» . В Дж. Филде (ред.). Обоняние . Вашингтон, округ Колумбия: Американская физиологическая ассоциация. стр. 535–548 . Проверено 6 ноября 2013 г.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Барнетт, SA (1963). Крыса; исследование поведения . Чикаго: Паб Aldine. Компания OCLC 558946 .
- ^ а б Ашер, IM; Агаджанян, ГК (1974). «6-гидроксидофаминовые поражения обонятельных бугорков и хвостатых ядер: влияние на стереотипное поведение крыс, вызванное амфетамином». Исследования мозга . 82 (1): 1–12. дои : 10.1016/0006-8993(74)90888-9 . ПМИД 4373138 .
- ^ Маккензи, генеральный менеджер. (1972). «Роль обонятельного бугорка в стереотипном поведении, вызванном апоморфином у крыс». Психофармакология . 23 (3): 212–9. дои : 10.1007/bf00404127 . ПМИД 5026945 . S2CID 6928275 .
- ^ а б Икемото, С.; Мудрый, РА (2002). «Полезные эффекты холинергических агентов карбахола и неостигмина в задне-вентральной области покрышки» . Журнал неврологии . 22 (22): 9895–9904. doi : 10.1523/JNEUROSCI.22-22-09895.2002 . ПМЦ 6757811 . ПМИД 12427846 .
- ^ Бэкон, AW; Бонди, МВт; Лосось, ДП; Мерфи, К. (1998). «Очень ранние изменения в обонятельном функционировании из-за болезни Альцгеймера и роли аполипопротеина Е в обонянии». Анналы Нью-Йоркской академии наук . 855 (1): 723–731. Бибкод : 1998NYASA.855..723B . дои : 10.1111/j.1749-6632.1998.tb10651.x . ПМИД 9929677 . S2CID 29775199 .
- ^ Нордин, С.; Мерфи, К. (1996). «Нарушение сенсорной и когнитивной обонятельной функции при сомнительной болезни Альцгеймера». Нейропсихология . 10 (1): 112–119. дои : 10.1037/0894-4105.10.1.113 .
- ^ а б Доти, РЛ; Перл, ДП; Стил, Джей Си; Чен, К.М.; Пирс, Джей Ди младший; Рейес, П.; Курляндия, Литва (1991). «Дефицит идентификации запаха комплекса паркинсонизма-деменции на Гуаме: эквивалент болезни Альцгеймера и идиопатической болезни Паркинсона». Неврология . 41 (5 Приложение 2): 77–80, обсуждение 80–81. дои : 10.1212/WNL.41.5_Suppl_2.77 . ПМИД 2041598 . S2CID 36051446 .
- ^ Детари, Л.; Юхас, Г.; Кукорелли, Т. (1984). «Энергетические свойства базальных нейронов переднего мозга кошки во время цикла сон-бодрствование». Электроэнцефалография и клиническая нейрофизиология . 58 (4): 362–368. дои : 10.1016/0013-4694(84)90062-2 . ISSN 0013-4694 . ПМИД 6207005 .
- ^ Суо-Шаньи, МФ; Понец, Дж.; Гонон, Ф. (1991). «Пресинаптическое аутоингибирование электрически вызванного высвобождения дофамина, изученное в обонятельном бугорке крысы с помощью электрохимии in vivo». Нейронаука . 45 (3): 641–652. дои : 10.1016/0306-4522(91)90277-У . ISSN 0306-4522 . ПМИД 1775239 . S2CID 46471029 .
Дальнейшее чтение
[ редактировать ]- Хеймер, Л. (2003). «Новая анатомическая основа нервно-психических расстройств и злоупотребления наркотиками». Американский журнал психиатрии . 160 (10): 1726–1739. дои : 10.1176/appi.ajp.160.10.1726 . ПМИД 14514480 .
- Икемото, С. (2007). «Схема вознаграждения дофамина: две проекционные системы от вентральной части среднего мозга к комплексу прилежащего ядра и обонятельного бугорка» . Обзоры исследований мозга . 56 (1): 27–78. дои : 10.1016/j.brainresrev.2007.05.004 . ПМК 2134972 . ПМИД 17574681 .
- Икемото, С. (2010). «Схема вознаграждения мозга за пределами мезолимбической дофаминовой системы: нейробиологическая теория» . Неврологические и биоповеденческие обзоры . 35 (2): 129–150. doi : 10.1016/j.neubiorev.2010.02.001 . ПМЦ 2894302 . ПМИД 20149820 .
- Паксино, Г.; Франклин, К. (2000). Мозг мыши в стереотаксических координатах (Второе изд.). Сан-Диего: Академическая пресса. ISBN 978-0125476379 .
Внешние ссылки
[ редактировать ]- http://brainmaps.org/ajax-viewer.php?datid=1&sname=1201&hname=olfactory%20tubercle&hlabel=OLT
- Загадочный обонятельный бугорок. Обзор текущих исследований обонятельного бугорка в NIDCD.
- Области мозга: обонятельный бугорок Основная информация о бугорке на сайте НейроЛекс.
- Разделы, содержащие обонятельный бугорок. Иллюстрированные фрагменты всех типов, показывающие точное расположение бугорка в мозге на BrainMaps.