Тиранозавр
| Тиранозавр Временной диапазон: поздний мел , | |
|---|---|
 | |
| Реконструкция тираннозавра экземпляра типа в Музее естественной истории Карнеги. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Тираннозавриды |
| Подсемейство: | † Тираннозавры |
| Клэйд : | † Тираннозаврин |
| Род: | † Тираннозавр Осборн, 1905 год. |
| Типовой вид | |
| † Тираннозавр рекс Осборн, 1905 год. | |
| Другие виды | |
| |
| Синонимы | |
Синонимия рода Синонимия видов | |
Тираннозавр ( / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ -/ ) [а] — род крупных -теропод динозавров . Типовой вид Tyrannosaurus rex ( rex означает «король» на латыни ), часто сокращаемый до T. rex или в просторечии T-Rex , является одним из наиболее представленных теропод. Они обитали на территории нынешней западной части Северной Америки , на тогдашнем островном континенте, известном как Ларамидия . Тираннозавр имел гораздо более широкий ареал, чем другие тираннозавриды . Окаменелости обнаружены в различных скальных образованиях, относящихся к позднему кампанско - маастрихтскому периоду позднего мелового периода , от 72,7 до 66 миллионов лет назад . Это был последний известный представитель тираннозаврид и один из последних нептичьих динозавров , существовавших до мел-палеогенового вымирания .
Как и другие тираннозавриды, тираннозавр был двуногим хищником с массивным черепом, уравновешенным длинным тяжелым хвостом. По сравнению с большими и мощными задними конечностями, передние конечности тираннозавра были короткими, но необычайно мощными для своего размера и имели два когтистых пальца. Самый полный экземпляр имеет длину 12,3–12,4 м (40–41 фут), но, согласно большинству современных оценок, тираннозавра размеры могли превышать 13 м (43 фута) в длину и 3,7–4 м (12–13 футов). по высоте бедра и 8,8 тонны (8,7 длинных тонн; 9,7 коротких тонн) по массе. Хотя некоторые другие тероподы могли соперничать или превосходить тираннозавра по размеру , он по-прежнему остается одним из крупнейших известных наземных хищников, а его расчетная сила укуса является самой большой среди всех наземных животных. Безусловно, самый крупный хищник в своей среде обитания, Тираннозавр Рекс, скорее всего, был высшим хищником , охотившимся на гадрозавров , молодых бронированных травоядных, таких как цератопсы и анкилозавры , и, возможно, зауроподов . Некоторые эксперты предполагают, что динозавр был в первую очередь мусорщик . Вопрос о том, был ли тираннозавр высшим хищником или чистым падальщиком, был одним из самых долгих споров в палеонтологии . Сегодня большинство палеонтологов признают, что тираннозавр был одновременно активным хищником и падальщиком.
Среди экземпляров тираннозавра рекса есть почти полные скелеты. мягкие ткани и белки По крайней мере, в одном из этих образцов были обнаружены . Обилие ископаемого материала позволило провести значительные исследования многих аспектов его биологии, включая историю его жизни и биомеханику . Пищевые привычки, физиология и потенциальная скорость тираннозавра рекса являются предметом споров. Его таксономия также противоречива, поскольку некоторые ученые считают Tarbosaurus bataar из Азии третьим видом тираннозавров , в то время как другие считают, что Tarbosaurus является отдельным родом. других родов североамериканских тираннозаврид также были синонимами тираннозавра Несколько . два вида тираннозавров В настоящее время действительными считаются ; типовой вид T. rex и более ранний и недавно обнаруженный T. mcraeensis .
Как архетипический теропод, тираннозавр был одним из самых известных динозавров с начала 20-го века и появлялся в фильмах, рекламе, на почтовых марках и во многих других средствах массовой информации.
История исследований
Самые ранние находки

Зуб того, что сейчас задокументировано как тираннозавр рекс, был найден в июле 1874 года на Южной Столовой горе (Колорадо) студентом Джарвис-Холла (Колорадо) Питером Т. Дотсоном под эгидой профессора Артура Лейкса недалеко от Голдена, штат Колорадо . [1] В начале 1890-х годов Джон Белл Хэтчер собирал посткраниальные элементы в восточном Вайоминге . Считалось, что окаменелости принадлежат крупному виду Ornithomimus grandis (ныне Deinodon ), но теперь считаются останками T. rex . [2]
В 1892 году Эдвард Дринкер Коуп нашел два фрагмента позвонков большого динозавра. Коуп полагал, что фрагменты принадлежали динозавру «агатаумид» ( цератопсид ), и назвал их Manospondylus gigas , что означает «гигантский пористый позвонок», в связи с многочисленными отверстиями для кровеносных сосудов, которые он обнаружил в кости. [2] Останки M. gigas в 1907 году были идентифицированы Хэтчером как останки теропода, а не цератопсида. [3]
Генри Фэйрфилд Осборн признал сходство между Manospondylus gigas и T. rex еще в 1917 году, когда второй позвонок был потерян. Из-за фрагментарной природы Manospondylus vertebrae Осборн не стал синонимизировать эти два рода, вместо этого посчитав более старый род неопределенным. [4] В июне 2000 года Институт Блэк-Хиллз обнаружил около 10% скелета тираннозавра ( BHI 6248) на месте, которое могло быть первоначальным местонахождением M. gigas . [5]
Обнаружение скелета и присвоение ему имени

Барнум Браун , помощник куратора Американского музея естественной истории , нашел первый частичный скелет тираннозавра в восточном Вайоминге в 1900 году. Браун нашел еще один частичный скелет в формации Хелл-Крик в Монтане в 1902 году, состоящий примерно из 34 окаменелых костей. [6] В то время Браун писал: «Карьер № 1 содержит бедренную кость, лобковую кость, плечевую кость, три позвонка и две кости неопределенного происхождения большого плотоядного динозавра, не описанного Маршем … Я никогда не видел ничего подобного из мелового периода ». [7] Генри Фэйрфилд Осборн , президент Американского музея естественной истории , назвал второй скелет тиранозавра в 1905 году. Родовое название происходит от греческих слов τύραννος ( tyrannos , что означает «тиран») и σαῦρος ( sauros , что означает «ящерица»). ). Осборн использовал латинское слово rex , что означает «король», для конкретного имени. Таким образом, полный бином переводится как «ящерица-тиран-король» или «ящерица-тиран-король», подчеркивая размер животного и предполагаемое доминирование над другими видами того времени. [6]

Осборн назвал другой экземпляр Dynamosaurus imperiosus в статье 1905 года. [6] В 1906 году Осборн признал, что эти два скелета принадлежали к одному и тому же виду, и выбрал Тираннозавр в качестве предпочтительного названия. [8] Оригинальный материал о динамозавре находится в коллекции Музея естественной истории в Лондоне. [9] В 1941 году экземпляр типа тираннозавра был продан Музею естественной истории Карнеги в Питтсбурге, штат Пенсильвания, за 7000 долларов. [7] Динамозавр позже будет удостоен чести в 2018 году, когда Эндрю Макдональд и его коллеги описали другой вид тираннозаврид, Dynamoterror dynastes , чье имя было выбрано в связи с названием 1905 года, поскольку оно было «фаворитом детства» Макдональдса. [10]
С 1910-х по конец 1950-х годов открытия Барнума оставались единственными экземплярами тираннозавра , поскольку Великая депрессия и войны не позволили многим палеонтологам заняться этой областью. [5]
Возрождающийся интерес

возобновился Начиная с 1960-х годов интерес к тираннозаврам , в результате чего в западной части Северной Америки было обнаружено 42 скелета (5–80% полных по количеству костей). [5] В 1967 году доктор Уильям Макманнис обнаружил и восстановил скелет под названием «MOR 008», который по подсчету костей завершен на 15%, а реконструированный череп выставлен в Музее Скалистых гор . В 1990-е годы было сделано множество открытий, почти в два раза больше находок, чем за все предыдущие годы, включая два наиболее полных скелета, найденных на сегодняшний день: Сью и Стэна . [5]
Сью Хендриксон , палеонтолог- любитель , обнаружила самый полный (примерно 85%) и самый крупный тираннозавра скелет в формации Хелл-Крик 12 августа 1990 года. Образец Сью, названный в честь первооткрывателя, стал объектом судебной тяжбы по поводу его права собственности. . В 1997 году судебный процесс был урегулирован в пользу Мориса Уильямса, первоначального владельца земли. Коллекция окаменелостей была куплена Филдовым музеем естественной истории на аукционе за 7,6 миллиона долларов, что сделало ее самым дорогим скелетом динозавра до продажи Стэна за 31,8 миллиона долларов в 2020 году. [11] С 1998 по 1999 год сотрудники Полевого музея естественной истории потратили более 25 000 часов на то, чтобы снять камень с костей. [12] Затем кости были отправлены в Нью-Джерси, где было построено крепление, а затем отправлены обратно в Чикаго для окончательной сборки. Установленный скелет был открыт для публики 17 мая 2000 года в Полевом музее естественной истории. Исследование окаменевших костей этого экземпляра показало, что Сью достигла полного размера в 19 лет и умерла в 28 лет, что является самой продолжительной предполагаемой жизнью среди известных тираннозавров. [13]

Другой тираннозавр , получивший прозвище Стэн (BHI 3033), в честь палеонтолога-любителя Стэна Сакрисона, был обнаружен в формации Хелл-Крик в 1992 году. Стэн является вторым наиболее полным найденным скелетом: 199 найденных костей составляют 70% от общего числа. [14] У этого тираннозавра также было множество костных патологий, в том числе сломанные и зажившие ребра, сломанная (и зажившая) шея и значительная дыра в затылке размером примерно с зуб тираннозавра . [15]
В 1998 году Баки Дерфлингер заметил выступающий над землей палец тиранозавра , что сделало Дерфлингера, которому на тот момент было 20 лет, самым молодым человеком, обнаружившим тираннозавра . Экземпляр, названный Баки в честь своего первооткрывателя, представлял собой молодого взрослого человека ростом 3,0 метра (10 футов) и длиной 11 метров (35 футов). Баки — первый тираннозавр найденный , у которого сохранилась вилка (поперечный рычаг). Баки постоянно экспонируется в Детском музее Индианаполиса . [16]

Летом 2000 года экипажи, организованные Джеком Хорнером, обнаружили пять скелетов тираннозавров возле водохранилища Форт-Пек . [17] обнаружила 50% полный скелет молодого тираннозавра В 2001 году в формации Хелл-Крик команда из Музея естественной истории Берпи . Находка, получившая название Джейн (BMRP 2002.4.1), считалась первым известным скелетом карликового тираннозавра Nanotyrannus , но последующие исследования показали, что это, скорее всего, молодой тираннозавр , и наиболее полный известный молодой экземпляр; [18] Джейн выставлена в Музее естественной истории Берпи. [19] В 2002 году скелет по имени Уайрекс, обнаруженный коллекционерами-любителями Дэном Уэллсом и Доном Уайриком, содержал 114 костей и был готов на 38%. Раскопки были завершены в течение трех недель в 2004 году Институтом Блэк-Хиллз, когда были проведены первые онлайн -раскопки тираннозавра , предоставляющие ежедневные отчеты, фотографии и видео. [5]
В 2006 году Государственный университет Монтаны обнаружил, что он обладает самым большим черепом тираннозавра , когда-либо обнаруженным (из экземпляра под названием MOR 008), длиной 5 футов (152 см). [20] Последующие сравнения показали, что самая длинная голова составляла 136,5 см (53,7 дюйма) (у экземпляра LACM 23844), а самая широкая голова составляла 90,2 см (35,5 дюйма) (от Сью). [21]
Следы

Два изолированных окаменелых следа были предварительно отнесены к тираннозаврам . Первый был обнаружен на ранчо Филмонт Скаут , штат Нью-Мексико, в 1983 году американским геологом Чарльзом Пиллмором. Первоначально считалось, что он принадлежал гадрозавриду , но исследование следа выявило большую «пятку», неизвестную в следах динозавров- орнитопод , а также следы того, что могло быть Hallux — четвертым пальцем стопы тираннозавра, похожим на прибылый коготь. След был опубликован под названием ichnogenus Tyrannosauripuspilmorei в 1994 году Мартином Локли и Адрианом Хантом . Локли и Хант предположили, что весьма вероятно, что след был оставлен тираннозавром , что делает его первым известным следом этого вида. Трасса проложена на территории, которая когда-то была заболоченной илистой равниной, покрытой растительностью. Его размеры 83 сантиметра (33 дюйма) в длину и 71 сантиметр (28 дюймов) в ширину. [22]
О втором следе, который мог оставить тираннозавр, впервые сообщил в 2007 году британский палеонтолог Фил Мэннинг из формации Хелл-Крик в Монтане. Длина этого второго следа составляет 72 сантиметра (28 дюймов), что короче, чем след, описанный Локли и Хантом. был ли след оставлен тираннозавром Неясно, , хотя тираннозавр — единственный крупный теропод, который, как известно, существовал в формации Хелл-Крик. [23] [24]
Набор следов в Гленроке, штат Вайоминг, датируемых маастрихтским этапом позднего мелового периода и происходящих из формации Лэнс, был описан Скоттом Персонсом, Филом Карри и коллегами в 2016 году и, как полагают, принадлежит либо молодому тираннозавра , либо сомнительный тираннозавр Nanotyrannus lancensis . Судя по измерениям и положению следов, животное двигалось со скоростью от 2,8 до 5 миль в час, а высота бедер оценивалась от 1,56 до 2,06 м (от 5,1 до 6,8 футов). [25] [26] [27] В 2017 году появился дополнительный документ, в котором оценки скорости увеличились на 50–80%. [28]
Описание
Размер
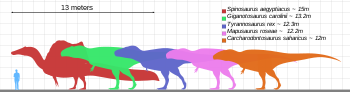
Ти-рекс был одним из крупнейших наземных хищников всех времен. Один из самых крупных и полных экземпляров по прозвищу Сью (FMNH PR2081) находится в Полевом музее естественной истории в Чикаго. Длина Сью составляла 12,3–12,4 м (40,4–40,7 футов). [29] [30] имел рост в бедрах 3,66–3,96 метра (12–13 футов), [31] [32] [33] и согласно самым последним исследованиям с использованием различных методов максимальная масса тела оценивается примерно в 8,4–8,46 метрических тонны (9,26–9,33 коротких тонны). [34] [35] Сообщается, что экземпляр по прозвищу Скотти (RSM P2523.8), находящийся в Королевском музее Саскачевана , имеет длину 13 м (43 фута). Используя метод оценки массы, который экстраполирует окружность бедренной кости, Скотти был оценен как самый крупный известный экземпляр с массой тела 8,87 метрических тонн (9,78 коротких тонн). [34] [36]
Не все найденные экземпляры взрослых тираннозавров такие большие. Исторически средние оценки массы взрослой особи на протяжении многих лет сильно различались: от всего лишь 4,5 метрических тонн (5,0 коротких тонн) до [37] [38] до более чем 7,2 метрических тонн (7,9 коротких тонн), [39] по самым современным оценкам от 5,4 до 8,0 метрических тонн (от 6,0 до 8,8 коротких тонн). [29] [40] [41] [42] [43]
Череп

Самые крупные из известных черепов тираннозавра имеют длину до 1,54 метра (5 футов). [20] [31] Большие фенестры (отверстия) в черепе уменьшали вес, как и у всех хищных теропод. В других отношениях череп тираннозавра значительно отличался от черепов крупных нетираннозавридных теропод . Он был чрезвычайно широким сзади, но имел узкую морду, обеспечивающую необычайно хорошее бинокулярное зрение . [44] [45] Кости черепа были массивными, а носовые и некоторые другие кости срослись, что препятствовало движению между ними; но многие из них были пневматизированы (содержали «соты» из крошечных воздушных пространств) и, следовательно, легче. Эти и другие особенности укрепления черепа являются частью тенденции тираннозаврид к более мощному укусу, который легко превзошел укус всех нетираннозаврид. [46] [47] [48] Кончик верхней челюсти имел U-образную форму (у большинства хищников, не являющихся тираннозавроидами, верхняя челюсть имела V-образную форму), что увеличивало количество тканей и костей, которые тираннозавр мог вырвать за один укус, хотя это также увеличивало нагрузку на переднюю часть. зубы. [49]

Зубы тираннозавра имели выраженную гетеродонтию (различия в форме). [50] [51] зубы Предчелюстные , по четыре с каждой стороны в передней части верхней челюсти, были плотно упакованы, D -образные в поперечном сечении, имели на задней поверхности укрепляющие гребни, были резцевидными (концы их представляли собой долотообразные лезвия) и загнутыми назад. D - образное поперечное сечение, усиливающие выступы и изгиб назад снижали риск того, что зубы сломались, когда тираннозавр кусал и тянул. Остальные зубы были крепкими, похожими на «смертоносные бананы», а не на кинжалы, располагались более широко и также имели укрепляющие гребни. [52] Те, что находятся в верхней челюсти, у взрослых особей по двенадцать с каждой стороны, [50] были крупнее своих собратьев нижней челюсти, за исключением задней части. По оценкам, самый крупный из найденных на данный момент зубов имел длину 30,5 см (12 дюймов), включая корень, когда животное было живо, что делает его самым большим зубом среди всех когда-либо найденных хищных динозавров. [53] Нижняя челюсть была крепкой. Его передняя зубная кость имела тринадцать зубов. За зубным рядом нижняя челюсть стала заметно выше. [50] Верхняя и нижняя челюсти тираннозавра , как и у многих динозавров, имели многочисленные отверстия или небольшие отверстия в кости. Для этих отверстий были предложены различные функции, например, сенсорная система, подобная крокодилу. [54] или признаки внеротовых структур, таких как чешуя или, возможно, губы, [55] [56] [57] с последующими исследованиями характера износа зубов теропод, подтверждающими такое предположение. [58]
Скелет

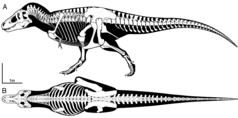
Позвоночный столб тираннозавра . состоял из десяти шейных позвонков, тринадцати задних позвонков и пяти крестцовых позвонков Число хвостовых позвонков неизвестно и вполне могло варьироваться у разных людей, но, вероятно, насчитывало не менее сорока. У Сью было сорок семь таких хвостовых позвонков. [50] Шея тираннозавра имела естественную S-образную форму, как и у других теропод. По сравнению с ними он был исключительно коротким, глубоким и мускулистым, чтобы поддерживать массивную голову. Второй позвонок, аксис, был особенно коротким. Остальные шейные позвонки были слабо опистоцелозными, т. е. с выпуклой передней частью тела позвонка и вогнутой задней. одиночные плевроцели — пневматические углубления, образованные воздушными мешками . Тела позвонков имели по бокам [50] Тела позвонков туловища были крепкими, но с узкой талией. Их нижняя сторона была килевидной. Передние борта имели вогнутую форму с глубоким вертикальным желобом. У них были крупные плевроцели. Их нервные отростки имели очень грубую переднюю и заднюю стороны для прикрепления сильных сухожилий. Крестцовые позвонки были слиты друг с другом как в телах позвонков, так и в нервных отростках. Они были пневматизированы. Они соединялись с тазом поперечными отростками и крестцовыми ребрами. Хвост был тяжелым и умеренно длинным, чтобы уравновешивать массивную голову и туловище и обеспечивать пространство для массивных опорно-двигательных мышц , прикрепленных к бедренным костям. Тринадцатый хвостовой позвонок образовывал точку перехода между глубоким основанием хвоста и средним хвостом, укрепленным довольно длинными передними сочленяющимися отростками. Нижняя часть туловища была покрыта восемнадцатью или девятнадцатью парами сегментированных брюшных ребер. [50]

Плечевой пояс был длиннее всей передней конечности. Лопатка имела узкое стержне, но была исключительно расширена на верхнем конце. Он соединялся длинным выступом вперед с клювовидным отростком , который был закругленным. Обе лопатки соединялись небольшой вилкой . Парные грудинные кости, возможно, состояли только из хряща . [50]
Передняя конечность или рука были очень короткими. Плечевая кость руки была короткой, но крепкой. У него был узкий верхний конец с исключительно закругленной головкой. Кости предплечья, локтевая и лучевая кости, представляли собой прямые элементы, значительно короче плечевой кости. Вторая пястная кость была длиннее и шире первой, тогда как в норме у теропод все наоборот. На передних конечностях было всего два когтистых пальца. [50] вместе с дополнительной шинообразной небольшой третьей пястной костью, представляющей собой остаток третьего пальца. [59]

Таз представлял собой крупную структуру. Его верхняя кость, подвздошная кость , была очень длинной и высокой, обеспечивая обширную зону прикрепления мышц задних конечностей. Передняя лобковая кость заканчивалась огромным лобковым сапогом, длиннее всего стержня элемента. Задняя седалищная кость была тонкой и прямой, направленной косо назад и вниз. [50]
В отличие от рук, задние конечности были одними из самых длинных по отношению к размеру тела среди всех теропод. На стопе плюсневая кость была «арктометатарзальной», то есть часть третьей плюсневой кости возле лодыжки была защемлена. Третья плюсневая кость также была исключительно извилистой. [50] Чтобы компенсировать огромную массу животного, многие кости по всему скелету были полыми, что уменьшило его вес без значительной потери прочности. [50]
Классификация

Tyrannosaurus — типовой род надсемейства Tyrannosauroidea , семейства Tyrannosauridae и подсемейства Tyrannosaurinae; другими словами, это стандарт, по которому палеонтологи решают, включать ли другие виды в ту же группу. Другие члены подсемейства тираннозавров включают североамериканского дасплетозавра и азиатского тарбозавра . [18] [60] оба из которых иногда были синонимами Тираннозавра . [61]
Тираннозаврид когда-то считалось потомками более ранних крупных теропод, таких как мегалозавры и карнозавры , хотя совсем недавно они были отнесены к более мелким целурозаврам . [49] Самой ранней группой тираннозавров были хохлатые процератозавриды, а более поздние и более производные представители принадлежат к Pantyrannosauria . Тираннозавры изначально были маленькими тероподами; однако, по крайней мере, некоторые из них стали крупнее к раннему меловому периоду .
Тираннозавроиды характеризуются сросшимися носовыми и зубными рядами. Пантираннозавры характеризуются уникальными особенностями бедер, а также увеличенным отверстием в квадратной кости, широкими заглазничными носами и носами в форме песочных часов. Некоторым из более продвинутых пантираннозавров не хватает носовой пневмонии, и у них меньшее соотношение плечевой и бедренной костей, а их руки начинают немного уменьшаться. У некоторых пантираннозавров начало развиваться арктометатарзус. У эвтираннозавров носовые кости шершавые, а нижнечелюстные отверстия уменьшены снаружи. У тираннозаврид отсутствуют кинетические черепа или специальные гребни на носовых костях, а также имеется слезная кость с характерным отростком на ней. У тираннозаврид также есть межфенестральная перемычка, которая вдвое меньше верхнечелюстного отверстия. [62]
Вполне вероятно, что тираннозавроиды приобрели известность после сокращения разнообразия аллозавроидов и мегалозавроидов, наблюдаемого на ранних стадиях позднего мела. Ниже представлена простая кладограмма общих взаимоотношений тираннозавроидов, обнаруженная после анализа, проведенного Ли и его коллегами в 2009 году. [63]
| Тираннозавроидея |
| |||||||||
Многие филогенетические анализы показали, что Tarbosaurus bataar является сестринским таксоном T. rex . [60] Открытие тираннозавра Lythronax также указывает на то, что Tarbosaurus и Tyrannosaurus тесно связаны, образуя кладу с другим азиатским тираннозавридом Zhuchengtyrannus , причем Lythronax является их сестринским таксоном. [64] [65] Дальнейшее исследование, проведенное в 2016 году Стивом Брусатте, Томасом Карром и его коллегами, также указывает на то, что Тираннозавр , возможно, был иммигрантом из Азии, а также возможным потомком Тарбозавра . [66]
Ниже представлена кладограмма тираннозаврид, основанная на филогенетическом анализе, проведенном Левеном и его коллегами в 2013 году. [64]
| Тираннозавриды |
| ||||||||||||||||||||||||
В своем описании Tyrannosaurus mcraeensis в 2024 году Далман и др . получили результаты, аналогичные предыдущим анализам, при этом тираннозавр был сестринским таксоном клады, образованной тарбозавром и жученгтиранносом , называемой тираннозавринами. Они также нашли подтверждение монофилетической кладе, содержащей Daspletosaurus и Thanatotheristes , обычно называемой Daspletosaurini . [67] [68]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Дополнительные виды

В 1955 году советский палеонтолог Евгений Малеев назвал новый вид — Tyrannosaurus bataar из Монголии . [69] К 1965 году этот вид был переименован в отдельный род Tarbosaurus bataar . [70] Хотя большинство палеонтологов продолжают считать эти два рода отдельными родами, некоторые авторы, такие как Томас Хольц , Кеннет Карпентер и Томас Карр, утверждают, что эти два вида достаточно похожи, чтобы их можно было считать членами одного и того же рода, восстанавливая монгольского таксона исходный бином . имя . [49] [71] [54]
Некоторые экземпляры из позднемеловых отложений Китая были описаны как новые виды тираннозавров : T. lanpingensis, основанный на изолированном боковом зубе из красных отложений Юньнани в 1975 году; T. turpanensis из формации Субаши , Турфанский бассейн , Синьцзян в 1978 г.; и T. luanchuanensis из формации Киба , бассейн Тантоу , провинция Хэнань в 1979–1980 гг. [72] [73] [74] Все эти таксоны были опубликованы без подробных описаний и позже были приняты в качестве младших синонимов Tarbosaurus bataar в 2004 году. Хольцем [60] [73] [74]
ВГИ, нет. 231/3, большая фаланговая кость , отнесенная к Tyrannosaurus sp. Ярковым в 2000 году, был найден в нижнем маастрихте Береславки, Россия . В 2004 году Аверьянов и Ярков переосмыслили его как I пястную кость или I плюсневую кость , которая, возможно, принадлежит цератозавру . [75] В своем обзоре 2023 года Аверьянов и Лопатин упоминают этот экземпляр, а также один зуб из того же места, что и Theropoda indet. [76]
В 2001 году различные зубы и плюсневые кости тираннозаврид, обнаруженные в карьере недалеко от Чжучэна , Китай, были отнесены китайским палеонтологом Ху Чэнчжи к недавно выведенному виду Tyrannosaurus zhuchengensis . Однако на соседнем участке правая верхняя челюсть и левая челюстная кость были отнесены к недавно созданному роду тираннозавридов Zhuchengtyrannus в 2011 году. Вполне возможно, что zhuchengensis является синонимом Zhuchengtyrannus T. . В любом случае T. zhuchengensis считается nomen dubium, поскольку у голотипа отсутствуют диагностические признаки ниже уровня Tyrannosaurinae. [77]
В исследовании 2022 года Грегори С. Пол и его коллеги утверждали, что Tyrannosaurus rex в традиционном понимании на самом деле представляет три вида: типовой вид Tyrannosaurus rex и два новых вида: T. Emperor (что означает «тиран-ящерица-император») и T. Регина (что означает «королева-тиран-ящерица»). Голотипом первого ( T.imperator ) является экземпляр Сью , а голотипом второго ( T.regin ) — Wankel rex . Разделение на несколько видов было в первую очередь основано на наблюдении очень высокой степени изменчивости пропорций и прочности бедренной кости (и других элементов скелета) у каталогизированных экземпляров T. rex , в большей степени, чем у других теропод, признанных одним из видов. разновидность. В качестве доказательства также использовались различия в общих пропорциях тела, представляющие крепкие и изящные морфотипы, в дополнение к количеству маленьких, тонких резцеобразных зубов в зубной кости, судя по лункам зубов. В частности, описанный в статье тираннозавр отличался прочной анатомией, умеренным соотношением длины бедренной кости к окружности и наличием единственного тонкого резцевидного зубного зуба; T. imperator считался крепким человеком с небольшим соотношением длины бедра к окружности и двумя тонкими зубами; и T. regina представляла собой изящную форму с высоким соотношением бедер и одним тонким зубом. Было замечено, что вариации в пропорциях и устойчивости стали более выраженными выше в выборке в стратиграфическом плане . Это было интерпретировано как единая более ранняя популяция, T. imperator , разделившаяся на более чем один таксон, T. rex и T. retina . [78]
Однако несколько других ведущих палеонтологов, в том числе Стивен Брусатт , Томас Карр , Томас Хольц , Дэвид Хоун, Цзинмай О'Коннор и Линдсей Занно , раскритиковали исследование или выразили скептицизм по поводу его выводов, когда к ним обратились различные средства массовой информации за комментариями. [79] [80] [81] Их критика была впоследствии опубликована в техническом документе. [82] Хольц и Занно отметили, что вполне вероятно, что существовало более одного вида тираннозавров , но посчитали, что новое исследование недостаточно для подтверждения предложенного им вида. Хольц заметил, что, даже если Tyrannosaurus Emperor вид представлял собой отдельный от Tyrannosaurus rex , он может представлять тот же вид, что и Nanotyrannus lancensis , и его нужно было бы называть Tyrannosaurus lancensis . О'Коннор, куратор Полевого музея, где T. Emperor выставлен голотип Sue, считал, что новый вид слишком плохо поддерживается, чтобы оправдать изменение вывесок на выставке. Брусатте, Карр и О'Коннор считали, что предложенные между видами отличительные черты отражают естественные вариации внутри вида. И Карр, и О'Коннор выразили обеспокоенность по поводу неспособности исследования определить, к какому из предложенных видов принадлежали несколько хорошо сохранившихся экземпляров. Другой палеонтолог, Филип Дж. Карри , первоначально был соавтором исследования, но отказался от него, поскольку не хотел участвовать в названии нового вида. [79]
Пол отверг возражения критиков, настаивая на том, что они не желают считать, что тираннозавр может представлять более чем один вид. [83] В последующей статье, ожидающей публикации, Пол подтвердил вывод, что тираннозавр состоит из трех видов. Он отметил, что критика исследования, назвавшего T.mperator и T.regina, сосредоточилась только на двух признаках, используемых для различения двух новых видов (количество мелких резцевидных зубов и прочность бедренных костей), в то время как в первоначальном исследовании также сравнивались прочность других костей (верхней кости, зубной кости, плечевой кости, подвздошной кости и плюсневых костей). Более того, Пол утверждал, что тираннозавра можно разделить на три разных вида по форме бугорков («заглазничных выступов») за глазами. Пол также утверждал, что прошлые исследования, приведшие к выводу, что тираннозавр состоит только из одного вида ( T. rex ), просто предполагали, что все скелеты тираннозавров представляют собой один вид, и что многие новые виды динозавров были названы на основе меньшего количества различий, чем он и его коллеги использовали при предложении T. Emperor и T. regina . [84]
Тираннозавр mcraensis

В 2024 году Далман и его коллеги описали останки тираннозавра, обнаруженные в 1983 году в формации озера Холл в кампане и начале маастрихта в Нью-Мексико. , хранящийся в Музее естественной истории и науки Нью-Мексико Ископаемый материал (NMMNH P-3698) , состоит из правой заглазничной кости , правой чешуйчатой кости , левой небной кости и неполной верхней челюсти черепа, левой зубной кости , правой селезеночной кости , правой предсуставной кости. , правый угловой и правый суставной от нижних челюстей, изолированные зубы и шевроны . [67] Некоторые кости были кратко упомянуты в 1984 году как принадлежащие тираннозаврам . [85] и описан в 1986 г. [86]
Леман и Карпентер (1990) предположили, что NMMNH P-3698 принадлежал к новому роду тираннозаврид. [87] в то время как Карр и Уильямсон (2000) не согласились с их утверждением. [88] Салливан и Лукас (2015) утверждали, что существует мало доказательств того, что NMMNH P-3698 является образцом тираннозавра рекса , поэтому они предварительно классифицировали его как ср. Тираннозавр вид; они также считали, что тираннозавр Макрея жил до Лансиана (до 67 миллионов лет назад), основываясь на его сосуществовании с Аламозавром . [89]
Далман и др . (2024) предложили новое название Tyrannosaurus mcraeensis для голотипа (NMMNH P-3698), отсылая к группе Макрея, слоям горных пород, к которым принадлежит формация Холл-Лейк. Возраст этих слоев горных пород оценивается от 72,7 до 70,9 млн лет назад, что соответствует позднему кампану или самому раннему маастрихту. [67] от позднего кампана до раннего маастрихта . Оценки возраста циркона U-Pb, проведенные Шанцем и Амато (2024), также подтверждают возраст формации Холл-Лейк [90] Это примерно за 5–7 миллионов лет до тираннозавра , существовавшего в конце маастрихта. Длина T. mcraeensis оценивалась в 12 метров (39 футов), что соответствует размеру взрослого тираннозавра . Эти двое отличаются характером черепа. Среди них зубная кость T. mcraeensis пропорционально длиннее и имеет менее выступающий подбородок, а нижняя челюсть более мелкая, чем у T. rex , что предполагает более слабый прикус. Зубы также более тупые и более сжаты с боков, а посторбитальные гребни менее выражены. Аналогичным образом, анатомия скелета демонстрирует общие характеристики с Тарбозавром и Чжученгтиранусом . [67] [91]
Нанотиран

Другие окаменелости тираннозаврид, обнаруженные в тех же формациях, что и T. rex, первоначально были классифицированы как отдельные таксоны, включая Aublysodon и Albertosaurus megagracilis , [61] последний в 1995 году был назван Dinotyrannus megagracilis . [92] Сейчас повсеместно считается, что эти окаменелости принадлежат молодым тираннозаврам . [93] Исключением может быть небольшой, но почти полный череп из Монтаны длиной 60 сантиметров (2,0 фута). Этот череп, CMNH в 1946 году как вид горгозавра ( G. lancensis ) 7541, первоначально был классифицирован Чарльзом У. Гилмором . [94] В 1988 году образец был повторно описан Робертом Т. Баккером , Филом Карри и Майклом Уильямсом, в то время куратором палеонтологии в Кливлендском музее естественной истории, где оригинальный образец находился и сейчас выставлен на обозрение. Их первоначальные исследования показали, что кости черепа срослись и, следовательно, представляют собой взрослый экземпляр. В свете этого Баккер и его коллеги отнесли череп к новому роду под названием Nanotyrannus (что означает «карликовый тиран» из-за его явно небольшого размера для взрослого человека). По оценкам, на момент смерти экземпляр имел длину около 5,2 метра (17 футов). [95] Однако в 1999 году подробный анализ, проведенный Томасом Карром, показал, что это молодой экземпляр, что побудило Карра и многих других палеонтологов считать его молодым особью тираннозавра . [96] [97]

В 2001 году был обнаружен более полный молодой тираннозавр (по прозвищу « Джейн », каталожный номер BMRP 2002.4.1), принадлежащий к тому же виду, что и исходный экземпляр Nanotyrannus . посвященную вопросам достоверности Nanotyrannus в Музее естественной истории Берпи Это открытие побудило провести в 2005 году . конференцию по тираннозаврам , Джейн» как подтверждение того, что Нанотиранус на самом деле был молодым тираннозавром . [98] [99] [100] Питер Ларсон продолжал поддерживать гипотезу о том, Н. что lancensis был отдельным, но близкородственным видом, судя по особенностям черепа, таким как наличие на два зуба в обеих челюстях больше, чем у T. rex ; а также пропорционально большие руки с фалангами на третьей пястной кости и другую анатомию поперечных рычагов у неописанного экземпляра. Он также утверждал, что Стигивенатор , которого обычно считают молодым тираннозавром , может быть более молодым экземпляром Nanotyrannus . [101] [102] Более поздние исследования показали, что у других тираннозаврид, таких как горгозавр, также наблюдалось уменьшение количества зубов во время роста. [96] и учитывая несоответствие в количестве зубов между особями одной и той же возрастной группы этого рода и тираннозавра , эта особенность также может быть связана с индивидуальными вариациями . [97] В 2013 году Карр отметил, что все различия, которые, как утверждается, поддерживают Nanotyrannus, оказались индивидуально или онтогенетически изменчивыми особенностями или продуктами искажения костей . [103]

В 2016 году анализ пропорций конечностей, проведенный Персонсом и Карри, показал, что образцы Nanotyrannus имеют разные уровни беглости, что потенциально отличает его от T. rex . [104] Однако палеонтолог Манабу Сакомото отметил, что на этот вывод может повлиять небольшой размер выборки , и расхождение не обязательно отражает таксономические различия. [105] В 2016 году Джошуа Шмерге приводил доводы в пользу достоверности Nanotyrannus , основываясь на особенностях черепа, включая зубную борозду на черепе BMRP 2002.4.1. По мнению Шмерге, поскольку эта особенность отсутствует у T. rex и встречается только у Dryptosaurus и albertosaurines, это предполагает, что Nanotyrannus является отдельным таксоном среди Albertosaurinae. [106] В том же году Карр и его коллеги отметили, что этого недостаточно, чтобы прояснить обоснованность или классификацию Nanotyrannus , поскольку это распространенная и онтогенетически изменчивая особенность среди тираннозавроидов. [107]
Исследование, проведенное в 2020 году Холли Вудворд и ее коллегами, показало, что все экземпляры, относящиеся к Nanotyrannus, были онтогенетически незрелыми, и обнаружило вероятность того, что эти экземпляры принадлежали T. rex . [108] В том же году Карр опубликовал статью Т. о rex , обнаружив, что CMNH 7541 соответствует ожидаемой онтогенетической вариации таксона и демонстрирует ювенильные характеристики, обнаруженные у других образцов. Он был классифицирован как подросток в возрасте до 13 лет с черепом менее 80 см (31 дюйм). Никаких существенных половых или филогенетических различий не было обнаружено ни у одного из 44 изученных экземпляров, при этом Карр заявил, что признаки потенциальной филогенетической важности уменьшаются с возрастом с той же скоростью, что и рост. [109] Обсуждая результаты статьи, Карр описал, как все экземпляры Nanotyrannus образуют непрерывный переходный период роста между самыми маленькими молодыми особями и взрослыми особями, в отличие от того, чего можно было бы ожидать, если бы это был отдельный таксон, где экземпляры группировались бы, исключая тираннозавра . Карр пришел к выводу, что «наноморфы» не так уж похожи друг на друга и вместо этого образуют важный мост в серии роста тираннозавра , который фиксирует начало глубоких изменений от мелкого черепа молодых особей к глубокому черепу, который наблюдается у тираннозавра. наблюдается у полностью развитых взрослых». [110]
Однако в статье 2024 года, опубликованной Ником Лонгричем и Эваном Томасом Сайттой, голотип был повторно исследован и упомянуты образцы Nanotyrannus . Основываясь на нескольких факторах, включая различия в морфологии, онтогенезе и филогении, Лонгрич и Саитта предполагают, что Nanotyrannus является отдельным таксоном, который может выходить за рамки Tyrannosauridae , основываясь на некоторых их филогенетических анализах. [111]
Палеобиология
История жизни

Идентификация нескольких экземпляров как молодых тираннозавров позволила ученым задокументировать онтогенетические изменения вида, оценить продолжительность жизни и определить, насколько быстро животные могли бы вырасти. Самый маленький из известных особей ( LACM 28471, «иорданский теропод»), по оценкам, весил всего 30 кг (66 фунтов), тогда как самые крупные взрослые особи, такие как FMNH PR2081 (Сью), скорее всего, весили около 5650 кг (12 460 фунтов). Гистологический анализ костей тиранозавра показал, что LACM 28471 на момент смерти имел возраст всего 2 года, в то время как Сью было 28 лет, возраст, который, возможно, был близок к максимальному для этого вида. [40]

Гистология также позволила определить возраст других образцов. Кривые роста можно построить, нанеся на график возраст различных экземпляров вместе с их массой. Кривая роста тираннозавра . имеет S-образную форму: молодые особи остаются ниже 1800 кг (4000 фунтов) примерно до 14 лет, когда размер тела начал резко увеличиваться Во время этой фазы быстрого роста молодой тираннозавр будет набирать в среднем 600 кг (1300 фунтов) в год в течение следующих четырех лет. В 18 лет кривая снова выравнивается, что указывает на резкое замедление роста. Например, всего 600 кг (1300 фунтов) отделяли 28-летнюю Сью от 22-летней канадской особи ( RTMP 81.12.1). [40] Гистологическое исследование, проведенное в 2004 году разными исследователями, подтверждает эти результаты: обнаружено, что быстрый рост начал замедляться примерно в 16-летнем возрасте. [112]
Исследование Хатчинсона и его коллег, проведенное в 2011 году, в целом подтвердило предыдущие методы оценки, но их оценка пиковых темпов роста значительно выше; выяснилось, что «максимальные темпы роста тираннозавра на экспоненциальной стадии составляют 1790 кг/год». [29] Хотя эти результаты были намного выше предыдущих оценок, авторы отметили, что эти результаты значительно снизили большую разницу между фактической скоростью роста и той, которую можно было бы ожидать от животного такого размера. [29] Внезапное изменение скорости роста в конце скачка роста может указывать на физическую зрелость, гипотеза, которая подтверждается открытием медуллярной ткани в бедренной кости 16-20-летнего тираннозавра из Монтаны ( MOR 1125, также известный как B-rex ). Медуллярная ткань обнаруживается только у самок птиц во время овуляции, что указывает на репродуктивный возраст B-rex. [113] Дальнейшие исследования показывают, что этому экземпляру 18 лет. [114] В 2016 году Мэри Хигби Швейцер, Линдси Занно и коллеги наконец подтвердили, что мягкие ткани бедренной кости MOR 1125 представляют собой мозговую ткань. Это также подтвердило принадлежность экземпляра к женщине. Обнаружение медуллярной костной ткани у тираннозавра может оказаться полезным для определения пола других видов динозавров в будущих исследованиях, поскольку химический состав медуллярной ткани безошибочен. [115] Другие тираннозавриды демонстрируют чрезвычайно похожие кривые роста, хотя и с более низкими темпами роста, что соответствует их меньшим размерам взрослых особей. [116]
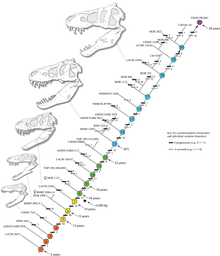
Дополнительное исследование, опубликованное в 2020 году Вудвордом и его коллегами для журнала Science Advances, показывает, что во время своего роста от юного животного к взрослому тираннозавр был способен замедлять свой рост, чтобы противостоять таким факторам окружающей среды, как нехватка еды. Исследование, сосредоточенное на двух молодых особях в возрасте от 13 до 15 лет, хранящихся в Музее Берпи в Иллинойсе, показывает, что скорость созревания тираннозавра зависела от изобилия ресурсов. Это исследование также показывает, что в таких меняющихся условиях тираннозавр был особенно хорошо приспособлен к среде, которая ежегодно менялась в отношении изобилия ресурсов, намекая на то, что другим хищникам среднего размера, возможно, было трудно выжить в таких суровых условиях, и объясняя разделение ниш между молодыми и молодыми особями. взрослые тираннозавры. Исследование также показывает, что тираннозавр и сомнительный род Nanotyrannus являются синонимами, что следует из анализа годичных колец в костях двух изученных экземпляров. [117] [118]
Более половины известных экземпляров тиранозавров, по-видимому, умерли в течение шести лет после достижения половой зрелости, и эта закономерность сегодня также наблюдается у других тираннозавров и у некоторых крупных долгоживущих птиц и млекопитающих. Для этих видов характерен высокий уровень младенческой смертности, за которым следует относительно низкая смертность среди молоди. Смертность снова возрастает после достижения половой зрелости, отчасти из-за стресса воспроизводства. Одно исследование предполагает, что редкость окаменелостей молодых тираннозавров отчасти объясняется низким уровнем смертности молоди; в этом возрасте животные не умирали в больших количествах и поэтому не часто превращались в окаменелости. Эта редкость также может быть связана с неполнотой летописи окаменелостей или с предвзятостью коллекционеров окаменелостей к более крупным и впечатляющим экземплярам. [116] В лекции 2013 года Томас Хольц-младший предположил, что динозавры «жили быстро и умирали молодыми», потому что они быстро размножались, тогда как млекопитающие имеют большую продолжительность жизни, потому что им требуется больше времени для размножения. [119] Грегори С. Пол также пишет, что тираннозавр быстро размножался и умер молодым, но объясняет свою короткую продолжительность жизни опасной жизнью, которую они прожили. [120]
Кожа и возможно нитевидное оперение.

Открытие пернатых динозавров привело к спорам о том, мог ли тираннозавр быть пернатым и если да, то в какой степени. [121] [122] Нитевидные структуры, которые обычно считаются предшественниками перьев , были обнаружены у мелкого базального тираннозавра Dilong paradoxus из раннемеловой формации Исянь в Китае в 2004 году. [123] Поскольку на покровных отпечатках более крупных тираннозавроидов, известных в то время, были обнаружены признаки чешуи , исследователи, изучавшие Дилонга, предположили, что изолирующие перья могли быть потеряны более крупными видами из-за меньшего соотношения их поверхности к объему. [123] Последующее открытие гигантского вида Yutyrannus huali , также из Исяня, показало, что даже у некоторых крупных тираннозавроидов перья покрывали большую часть их тел, что ставит под сомнение гипотезу о том, что они были особенностью, связанной с размером. [124] В исследовании 2017 года были рассмотрены известные отпечатки кожи тираннозаврид, в том числе экземпляра тираннозавра по прозвищу «Вайрекс» (BHI 6230), у которого сохранились участки мозаичной чешуи на хвосте, бедре и шее. [5] Исследование пришло к выводу, что перьевой покров крупных тираннозаврид, таких как тираннозавр , если и присутствовал, то ограничивался верхней стороной туловища. [121]
В тезисах конференции, опубликованных в 2016 году, утверждалось, что у теропод, таких как тираннозавр, верхние зубы были покрыты губами, а не голыми, как у крокодилов . Это было основано на наличии эмали , которая, согласно исследованию, должна оставаться гидратированной — проблема, с которой не сталкиваются водные животные, такие как крокодилы. [56] Тем не менее, была критика в пользу идеи губ: аналитическое исследование 2017 года показало, что у тираннозаврид были большие плоские чешуйки на морде, а не на губах, как у современных крокодилов. [54] [125] Но у крокодилов скорее потрескавшаяся ороговевшая кожа, а не плоская чешуя; Наблюдая за бугристой морщинистой поверхностью тираннозаврид и сравнивая ее с современными ящерицами, исследователи обнаружили, что тираннозавриды имели чешуйчатую чешую, а не кожу, как у крокодила. [126] [127]
In 2023, Cullen and colleagues supported the idea that theropods like tyrannosaurids had lips based on anatomical patterns, such as those of the foramina on their face and jaws, more similar to those of modern squamates such as monitor lizards or marine iguanas than those of modern crocodilians like alligators. By comparatively analysing the dentition of Daspletosaurus and the American alligator, it was shown that the enamel of tyrannosaurids had no significant wear, while that of modern crocodilians had erosion on the labial side and substantial wear. This suggests that it is likely that theropod teeth existed under hydrated conditions (i.e. extraoral tissues). On the basis of the relationship between hydration and wear resistance, the authors argued that it is unlikely that the teeth of theropods including tyrannosaurids would have remained unworn when exposed for a long time, as it would have been difficult to maintain hydration. The authors also performed regression analyses to demonstrate the relationship between tooth height and skull length, and found that varanids like the crocodile monitor had substantially greater tooth height–to–skull length ratios than Tyrannosaurus, indicating that the teeth of theropods were not too big to be covered by extraoral tissues when the mouth was closed.[58]
Sexual dimorphism

As the number of known specimens increased, scientists began to analyze the variation between individuals and discovered what appeared to be two distinct body types, or morphs, similar to some other theropod species. As one of these morphs was more solidly built, it was termed the 'robust' morph while the other was termed 'gracile'. Several morphological differences associated with the two morphs were used to analyze sexual dimorphism in T. rex, with the 'robust' morph usually suggested to be female. For example, the pelvis of several 'robust' specimens seemed to be wider, perhaps to allow the passage of eggs.[128] It was also thought that the 'robust' morphology correlated with a reduced chevron on the first tail vertebra, also ostensibly to allow eggs to pass out of the reproductive tract, as had been erroneously reported for crocodiles.[129]
In recent years, evidence for sexual dimorphism has been weakened. A 2005 study reported that previous claims of sexual dimorphism in crocodile chevron anatomy were in error, casting doubt on the existence of similar dimorphism between T. rex sexes.[130] A full-sized chevron was discovered on the first tail vertebra of Sue, an extremely robust individual, indicating that this feature could not be used to differentiate the two morphs anyway. As T. rex specimens have been found from Saskatchewan to New Mexico, differences between individuals may be indicative of geographic variation rather than sexual dimorphism. The differences could also be age-related, with 'robust' individuals being older animals.[50]
Only a single Tyrannosaurus specimen has been conclusively shown to belong to a specific sex. Examination of B-rex demonstrated the preservation of soft tissue within several bones. Some of this tissue has been identified as a medullary tissue, a specialized tissue grown only in modern birds as a source of calcium for the production of eggshell during ovulation. As only female birds lay eggs, medullary tissue is only found naturally in females, although males are capable of producing it when injected with female reproductive hormones like estrogen. This strongly suggests that B-rex was female and that she died during ovulation.[113] Recent research has shown that medullary tissue is never found in crocodiles, which are thought to be the closest living relatives of dinosaurs. The shared presence of medullary tissue in birds and other theropod dinosaurs is further evidence of the close evolutionary relationship between the two.[131]
Posture

Like many bipedal dinosaurs, T. rex was historically depicted as a 'living tripod', with the body at 45 degrees or less from the vertical and the tail dragging along the ground, similar to a kangaroo. This concept dates from Joseph Leidy's 1865 reconstruction of Hadrosaurus, the first to depict a dinosaur in a bipedal posture.[132] In 1915, convinced that the creature stood upright, Henry Fairfield Osborn, former president of the American Museum of Natural History, further reinforced the notion in unveiling the first complete T. rex skeleton arranged this way. It stood in an upright pose for 77 years, until it was dismantled in 1992.[133]
By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the dislocation or weakening of several joints, including the hips and the articulation between the head and the spinal column.[134] The inaccurate AMNH mount inspired similar depictions in many films and paintings (such as Rudolph Zallinger's famous mural The Age of Reptiles in Yale University's Peabody Museum of Natural History)[135] until the 1990s, when films such as Jurassic Park introduced a more accurate posture to the general public.[136] Modern representations in museums, art, and film show T. rex with its body approximately parallel to the ground with the tail extended behind the body to balance the head.[137]
To sit down, Tyrannosaurus may have settled its weight backwards and rested its weight on a pubic boot, the wide expansion at the end of the pubis in some dinosaurs. With its weight rested on the pelvis, it may have been free to move the hindlimbs. Getting back up again might have involved some stabilization from the diminutive forelimbs.[138][134] The latter known as Newman's pushup theory has been debated. Nonetheless, Tyrannosaurus was probably able to get up if it fell, which only would have required placing the limbs below the center of gravity, with the tail as an effective counterbalance. Healed stress fractures in the forelimbs have been put forward both as evidence that the arms cannot have been very useful[139][140] and as evidence that they were indeed used and acquired wounds,[141] like the rest of the body.
Arms

When T. rex was first discovered, the humerus was the only element of the forelimb known.[6] For the initial mounted skeleton as seen by the public in 1915, Osborn substituted longer, three-fingered forelimbs like those of Allosaurus.[4] A year earlier, Lawrence Lambe described the short, two-fingered forelimbs of the closely related Gorgosaurus.[142] This strongly suggested that T. rex had similar forelimbs, but this hypothesis was not confirmed until the first complete T. rex forelimbs were identified in 1989, belonging to MOR 555 (the "Wankel rex").[143][144] The remains of Sue also include complete forelimbs.[50] T. rex arms are very small relative to overall body size, measuring only 1 meter (3.3 ft) long, and some scholars have labelled them as vestigial. However, the bones show large areas for muscle attachment, indicating considerable strength. This was recognized as early as 1906 by Osborn, who speculated that the forelimbs may have been used to grasp a mate during copulation.[8] Newman (1970) suggested that the forelimbs were used to assist Tyrannosaurus in rising from a prone position.[134] Since then, other functions have been proposed, although some scholars find them implausible.[140] Padian (2022) argued that the reduction of the arms in tyrannosaurids did not serve a particular function but was a secondary adaptation, stating that as tyrannosaurids developed larger and more powerful skulls and jaws, the arms got smaller to avoid being bitten or torn by other individuals, particularly during group feedings.[140]
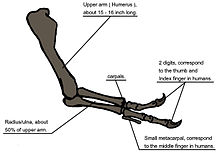
Another possibility is that the forelimbs held struggling prey while it was killed by the tyrannosaur's enormous jaws. This hypothesis may be supported by biomechanical analysis. T. rex forelimb bones exhibit extremely thick cortical bone, which has been interpreted as evidence that they were developed to withstand heavy loads. The biceps brachii muscle of an adult T. rex was capable of lifting 199 kilograms (439 lb) by itself; other muscles such as the brachialis would work along with the biceps to make elbow flexion even more powerful. The M. biceps muscle of T. rex was 3.5 times as powerful as the human equivalent. A T. rex forearm had a limited range of motion, with the shoulder and elbow joints allowing only 40 and 45 degrees of motion, respectively. In contrast, the same two joints in Deinonychus allow up to 88 and 130 degrees of motion, respectively, while a human arm can rotate 360 degrees at the shoulder and move through 165 degrees at the elbow. The heavy build of the arm bones, strength of the muscles, and limited range of motion may indicate a system evolved to hold fast despite the stresses of a struggling prey animal. In the first detailed scientific description of Tyrannosaurus forelimbs, paleontologists Kenneth Carpenter and Matt Smith dismissed notions that the forelimbs were useless or that Tyrannosaurus was an obligate scavenger.[145]
The idea that the arms served as weapons when hunting prey have also been proposed by Steven M. Stanley, who suggested that the arms were used for slashing prey, especially by using the claws to rapidly inflict long, deep gashes to its prey.[146] This was dismissed by Padian, who argued that Stanley based his conclusion on incorrectly estimated forelimb size and range of motion.[140]
Thermoregulation

Tyrannosaurus, like most dinosaurs, was long thought to have an ectothermic ("cold-blooded") reptilian metabolism. The idea of dinosaur ectothermy was challenged by scientists like Robert T. Bakker and John Ostrom in the early years of the "Dinosaur Renaissance", beginning in the late 1960s.[147][148] T. rex itself was claimed to have been endothermic ("warm-blooded"), implying a very active lifestyle.[38] Since then, several paleontologists have sought to determine the ability of Tyrannosaurus to regulate its body temperature. Histological evidence of high growth rates in young T. rex, comparable to those of mammals and birds, may support the hypothesis of a high metabolism. Growth curves indicate that, as in mammals and birds, T. rex growth was limited mostly to immature animals, rather than the indeterminate growth seen in most other vertebrates.[112]
Oxygen isotope ratios in fossilized bone are sometimes used to determine the temperature at which the bone was deposited, as the ratio between certain isotopes correlates with temperature. In one specimen, the isotope ratios in bones from different parts of the body indicated a temperature difference of no more than 4 to 5 °C (7 to 9 °F) between the vertebrae of the torso and the tibia of the lower leg. This small temperature range between the body core and the extremities was claimed by paleontologist Reese Barrick and geochemist William Showers to indicate that T. rex maintained a constant internal body temperature (homeothermy) and that it enjoyed a metabolism somewhere between ectothermic reptiles and endothermic mammals.[149] Other scientists have pointed out that the ratio of oxygen isotopes in the fossils today does not necessarily represent the same ratio in the distant past, and may have been altered during or after fossilization (diagenesis).[150] Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus).[151] Ornithischian dinosaurs also showed evidence of homeothermy, while varanid lizards from the same formation did not.[152] In 2022, Wiemann and colleagues used a different approach—the spectroscopy of lipoxidation signals, which are byproducts of oxidative phosphorylation and correlate with metabolic rates—to show that various dinosaur genera including Tyrannosaurus had endothermic metabolisms, on par with that of modern birds and higher than that of mammals. They also suggested that such a metabolism was ancestrally common to all dinosaurs.[153]
Even if T. rex does exhibit evidence of homeothermy, it does not necessarily mean that it was endothermic. Such thermoregulation may also be explained by gigantothermy, as in some living sea turtles.[154][155][156] Similar to contemporary crocodilians, openings (dorsotemporal fenestrae) in the skull roofs of Tyrannosaurus may have aided thermoregulation.[157]
Soft tissue
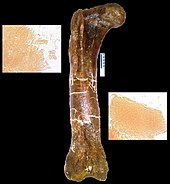
In the March 2005 issue of Science, Mary Higby Schweitzer of North Carolina State University and colleagues announced the recovery of soft tissue from the marrow cavity of a fossilized leg bone from a T. rex. The bone had been intentionally, though reluctantly, broken for shipping and then not preserved in the normal manner, specifically because Schweitzer was hoping to test it for soft tissue.[158] Designated as the Museum of the Rockies specimen 1125, or MOR 1125, the dinosaur was previously excavated from the Hell Creek Formation. Flexible, bifurcating blood vessels and fibrous but elastic bone matrix tissue were recognized. In addition, microstructures resembling blood cells were found inside the matrix and vessels. The structures bear resemblance to ostrich blood cells and vessels. Whether an unknown process, distinct from normal fossilization, preserved the material, or the material is original, the researchers do not know, and they are careful not to make any claims about preservation.[159] If it is found to be original material, any surviving proteins may be used as a means of indirectly guessing some of the DNA content of the dinosaurs involved, because each protein is typically created by a specific gene. The absence of previous finds may be the result of people assuming preserved tissue was impossible, therefore not looking. Since the first, two more tyrannosaurs and a hadrosaur have also been found to have such tissue-like structures.[158] Research on some of the tissues involved has suggested that birds are closer relatives to tyrannosaurs than other modern animals.[160]
In studies reported in Science in April 2007, Asara and colleagues concluded that seven traces of collagen proteins detected in purified T. rex bone most closely match those reported in chickens, followed by frogs and newts. The discovery of proteins from a creature tens of millions of years old, along with similar traces the team found in a mastodon bone at least 160,000 years old, upends the conventional view of fossils and may shift paleontologists' focus from bone hunting to biochemistry. Until these finds, most scientists presumed that fossilization replaced all living tissue with inert minerals. Paleontologist Hans Larsson of McGill University in Montreal, who was not part of the studies, called the finds "a milestone", and suggested that dinosaurs could "enter the field of molecular biology and really slingshot paleontology into the modern world".[161]
The presumed soft tissue was called into question by Thomas Kaye of the University of Washington and his co-authors in 2008. They contend that what was really inside the tyrannosaur bone was slimy biofilm created by bacteria that coated the voids once occupied by blood vessels and cells.[162] The researchers found that what previously had been identified as remnants of blood cells, because of the presence of iron, were actually framboids, microscopic mineral spheres bearing iron. They found similar spheres in a variety of other fossils from various periods, including an ammonite. In the ammonite, they found the spheres in a place where the iron they contain could not have had any relationship to the presence of blood.[163] Schweitzer has strongly criticized Kaye's claims and argues that there is no reported evidence that biofilms can produce branching, hollow tubes like those noted in her study.[164] San Antonio, Schweitzer and colleagues published an analysis in 2011 of what parts of the collagen had been recovered, finding that it was the inner parts of the collagen coil that had been preserved, as would have been expected from a long period of protein degradation.[165] Other research challenges the identification of soft tissue as biofilm and confirms finding "branching, vessel-like structures" from within fossilized bone.[166]
Speed

Scientists have produced a wide range of possible maximum running speeds for Tyrannosaurus: mostly around 9 meters per second (32 km/h; 20 mph), but as low as 4.5–6.8 meters per second (16–24 km/h; 10–15 mph) and as high as 20 meters per second (72 km/h; 45 mph), though it running this speed is very unlikely. Tyrannosaurus was a bulky and heavy carnivore so it is unlikely to run very fast at all compared to other theropods like Carnotaurus or Giganotosaurus.[167] Researchers have relied on various estimating techniques because, while there are many tracks of large theropods walking, none showed evidence of running.[168]
A 2002 report used a mathematical model (validated by applying it to three living animals: alligators, chickens, and humans; and eight more species, including emus and ostriches[168]) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph).[167] Scientists who think that Tyrannosaurus was able to run point out that hollow bones and other features that would have lightened its body may have kept adult weight to a mere 4.5 metric tons (5.0 short tons) or so, or that other animals like ostriches and horses with long, flexible legs are able to achieve high speeds through slower but longer strides.[168] Proposed top speeds exceeded 40 kilometers per hour (25 mph) for Tyrannosaurus, but were deemed infeasible because they would require exceptional leg muscles of approximately 40–86% of total body mass. Even moderately fast speeds would have required large leg muscles. If the muscle mass was less, only 18 kilometers per hour (11 mph) for walking or jogging would have been possible.[167] Holtz noted that tyrannosaurids and some closely related groups had significantly longer distal hindlimb components (shin plus foot plus toes) relative to the femur length than most other theropods, and that tyrannosaurids and their close relatives had a tightly interlocked metatarsus (foot bones).[169] The third metatarsal was squeezed between the second and fourth metatarsals to form a single unit called an arctometatarsus. This ankle feature may have helped the animal to run more efficiently.[170] Together, these leg features allowed Tyrannosaurus to transmit locomotory forces from the foot to the lower leg more effectively than in earlier theropods.[169]

Additionally, a 2020 study indicates that Tyrannosaurus and other tyrannosaurids were exceptionally efficient walkers. Studies by Dececchi et al., compared the leg proportions, body mass, and the gaits of more than 70 species of theropod dinosaurs including Tyrannosaurus and its relatives. The research team then applied a variety of methods to estimate each dinosaur's top speed when running as well as how much energy each dinosaur expended while moving at more relaxed speeds such as when walking. Among smaller to medium-sized species such as dromaeosaurids, longer legs appear to be an adaptation for faster running, in line with previous results by other researchers. But for theropods weighing over 1,000 kg (2,200 lb), top running speed is limited by body size, so longer legs instead were found to have correlated with low-energy walking. The results further indicate that smaller theropods evolved long legs as a means to both aid in hunting and escape from larger predators while larger theropods that evolved long legs did so to reduce the energy costs and increase foraging efficiency, as they were freed from the demands of predation pressure due to their role as apex predators. Compared to more basal groups of theropods in the study, tyrannosaurs like Tyrannosaurus itself showed a marked increase in foraging efficiency due to reduced energy expenditures during hunting or scavenging. This in turn likely resulted in tyrannosaurs having a reduced need for hunting forays and requiring less food to sustain themselves as a result. Additionally, the research, in conjunction with studies that show tyrannosaurs were more agile than other large-bodied theropods, indicates they were quite well-adapted to a long-distance stalking approach followed by a quick burst of speed to go for the kill. Analogies can be noted between tyrannosaurids and modern wolves as a result, supported by evidence that at least some tyrannosaurids were hunting in group settings.[171][172]
A study published in 2021 by Pasha van Bijlert et al., calculated the preferred walking speed of Tyrannosaurus, reporting a speed of 1.28 meters per second (4.6 km/h; 2.9 mph). While walking, animals reduce their energy expenditure by choosing certain step rhythms at which their body parts resonate. The same would have been true for dinosaurs, but previous studies did not fully account for the impact the tail had on their walking speeds. According to the authors, when a dinosaur walked, its tail would slightly sway up and down with each step as a result of the interspinous ligaments suspending the tail. Like rubber bands, these ligaments stored energy when they are stretched due to the swaying of the tail. Using a 3-D model of Tyrannosaurus specimen Trix, muscles and ligaments were reconstructed to simulate the tail movements. This results in a rhythmic, energy-efficient walking speed for Tyrannosaurus similar to that seen in living animals such as humans, ostriches and giraffes.[173]
A 2017 study estimated the top running speed of Tyrannosaurus as 17 mph (27 km/h), speculating that Tyrannosaurus exhausted its energy reserves long before reaching top speed, resulting in a parabola-like relationship between size and speed.[174][175] Another 2017 study hypothesized that an adult Tyrannosaurus was incapable of running due to high skeletal loads. Using a calculated weight estimate of 7 tons, the model showed that speeds above 11 mph (18 km/h) would have probably shattered the leg bones of Tyrannosaurus. The finding may mean that running was also not possible for other giant theropod dinosaurs like Giganotosaurus, Mapusaurus and Acrocanthosaurus.[176]However, studies by Eric Snively and colleagues, published in 2019 indicate that Tyrannosaurus and other tyrannosaurids were more maneuverable than allosauroids and other theropods of comparable size due to low rotational inertia compared to their body mass combined with large leg muscles. As a result, it is hypothesized that Tyrannosaurus was capable of making relatively quick turns and could likely pivot its body more quickly when close to its prey, or that while turning, the theropod could "pirouette" on a single planted foot while the alternating leg was held out in a suspended swing during a pursuit. The results of this study potentially could shed light on how agility could have contributed to the success of tyrannosaurid evolution.[177]
Possible footprints

Rare fossil footprints and trackways found in New Mexico and Wyoming that are assigned to the ichnogenus Tyrannosauripus have been attributed to being made by Tyrannosaurus, based on the stratigraphic age of the rocks they are preserved in. The first specimen, found in 1994 was described by Lockley and Hunt and consists of a single, large footprint. Another pair of ichnofossils, described in 2021, show a large tyrannosaurid rising from a prone position by rising up using its elbows in conjunction with the pads on their feet to stand. These two unique sets of fossils were found in Ludlow, Colorado and Cimarron, New Mexico.[178] Another ichnofossil described in 2018, perhaps belonging to a juvenile Tyrannosaurus or the dubious genus Nanotyrannus was uncovered in the Lance Formation of Wyoming. The trackway itself offers a rare glimpse into the walking speed of tyrannosaurids, and the trackmaker is estimated to have been moving at a speed of 4.5–8.0 kilometers per hour (2.8–5.0 mph), significantly faster than previously assumed for estimations of walking speed in tyrannosaurids.[179][180]
Brain and senses

A study conducted by Lawrence Witmer and Ryan Ridgely of Ohio University found that Tyrannosaurus shared the heightened sensory abilities of other coelurosaurs, highlighting relatively rapid and coordinated eye and head movements; an enhanced ability to sense low frequency sounds, which would allow tyrannosaurs to track prey movements from long distances; and an enhanced sense of smell.[181] A study published by Kent Stevens concluded that Tyrannosaurus had keen vision. By applying modified perimetry to facial reconstructions of several dinosaurs including Tyrannosaurus, the study found that Tyrannosaurus had a binocular range of 55 degrees, surpassing that of modern hawks. Stevens estimated that Tyrannosaurus had 13 times the visual acuity of a human and surpassed the visual acuity of an eagle, which is 3.6 times that of a person. Stevens estimated a limiting far point (that is, the distance at which an object can be seen as separate from the horizon) as far as 6 km (3.7 mi) away, which is greater than the 1.6 km (1 mi) that a human can see.[44][45][182]
Thomas Holtz Jr. would note that high depth perception of Tyrannosaurus may have been due to the prey it had to hunt, noting that it had to hunt ceratopsians such as Triceratops, ankylosaurs such as Ankylosaurus, and hadrosaurs. He would suggest that this made precision more crucial for Tyrannosaurus enabling it to, "get in, get that blow in and take it down." In contrast, Acrocanthosaurus had limited depth perception because they hunted large sauropods, which were relatively rare during the time of Tyrannosaurus.[119]
Though no Tyrannosaurus sclerotic ring has been found, Kenneth Carpenter estimated its size based on that of Gorgosaurus. The inferred sclerotic ring for the Stan specimen is ~7 cm (2.8 in) in diameter with an internal aperture diameter of ~3.5 cm (1.4 in). Based on eye proportions in living reptiles, this implies a pupil diameter of about 2.5 cm (0.98 in), an iris diameter about that of the sclerotic ring, and an eyeball diameter of 11–12 cm (4.3–4.7 in). Carpenter also estimated an eyeball depth of ~7.7–9.6 cm (3.0–3.8 in). Based on these calculations, the f-number for Stan's eye is 3–3.8; since diurnal animals have f-numbers of 2.1 or higher, this would indicate that Tyrannosaurus had poor low-light vision and hunted during the day.[183]
Tyrannosaurus had very large olfactory bulbs and olfactory nerves relative to their brain size, the organs responsible for a heightened sense of smell. This suggests that the sense of smell was highly developed, and implies that tyrannosaurs could detect carcasses by scent alone across great distances. The sense of smell in tyrannosaurs may have been comparable to modern vultures, which use scent to track carcasses for scavenging. Research on the olfactory bulbs has shown that T. rex had the most highly developed sense of smell of 21 sampled non-avian dinosaur species.[184]

Somewhat unusually among theropods, T. rex had a very long cochlea. The length of the cochlea is often related to hearing acuity, or at least the importance of hearing in behavior, implying that hearing was a particularly important sense to tyrannosaurs. Specifically, data suggests that T. rex heard best in the low-frequency range, and that low-frequency sounds were an important part of tyrannosaur behavior.[181] A 2017 study by Thomas Carr and colleagues found that the snout of tyrannosaurids was highly sensitive, based on a high number of small openings in the facial bones of the related Daspletosaurus that contained sensory neurons. The study speculated that tyrannosaurs might have used their sensitive snouts to measure the temperature of their nests and to gently pick up eggs and hatchlings, as seen in modern crocodylians.[54] Another study published in 2021 further suggests that Tyrannosaurus had an acute sense of touch, based on neurovascular canals in the front of its jaws, which it could utilize to better detect and consume prey. The study, published by Kawabe and Hittori et al., suggests that Tyrannosaurus could also accurately sense slight differences in material and movement, allowing it to utilize different feeding strategies on different parts of its prey's carcasses depending on the situation. The sensitive neurovascular canals of Tyrannosaurus also likely were adapted to performing fine movements and behaviors such as nest building, parental care, and other social behavior such as intraspecific communication. The results of this study also align with results made in studying the related tyrannosaurid Daspletosaurus horneri and the allosauroid Neovenator, which have similar neurovascular adaptations, suggesting that the faces of theropods were highly sensitive to pressure and touch.[185][186] However, a more recent study reviewing the evolution of the trigeminal canals among sauropsids notes that a much denser network of neurovascular canals in the snout and lower jaw is more commonly encountered in aquatic or semiaquatic taxa (e.g., Spinosaurus, Halszkaraptor, Plesiosaurus), and taxa that developed a rhamphotheca (e.g., Caenagnathasia), while the network of canals in Tyrannosaurus appears simpler, though still more derived than in most ornithischians, and overall terrestrial taxa such as tyrannosaurids and Neovenator may have had average facial sensitivity for non-edentulous terrestrial theropods, although further research is needed. The neurovascular canals in Tyrannosaurus may instead have supported soft tissue structures for thermoregulation or social signaling, the latter of which could be confirmed by the fact that the neurovascular network of canals may have changed during ontogeny.[187]
A study by Grant R. Hurlburt, Ryan C. Ridgely and Lawrence Witmer obtained estimates for Encephalization Quotients (EQs), based on reptiles and birds, as well as estimates for the ratio of cerebrum to brain mass. The study concluded that Tyrannosaurus had the relatively largest brain of all adult non-avian dinosaurs with the exception of certain small maniraptoriforms (Bambiraptor, Troodon and Ornithomimus). The study found that Tyrannosaurus's relative brain size was still within the range of modern reptiles, being at most 2 standard deviations above the mean of non-avian reptile EQs. The estimates for the ratio of cerebrum mass to brain mass would range from 47.5 to 49.53 percent. According to the study, this is more than the lowest estimates for extant birds (44.6 percent), but still close to the typical ratios of the smallest sexually mature alligators which range from 45.9–47.9 percent.[188] Other studies, such as those by Steve Brusatte, indicate the encephalization quotient of Tyrannosaurus was similar in range (2.0–2.4) to a chimpanzee (2.2–2.5), though this may be debatable as reptilian and mammalian encephalization quotients are not equivalent.[189]
Social behavior

Philip J. Currie suggested that Tyrannosaurus may have been pack hunters, comparing T. rex to related species Tarbosaurus bataar and Albertosaurus sarcophagus, citing fossil evidence that may indicate gregarious (describing animals that travel in herds or packs) behavior.[190] A find in South Dakota where three T. rex skeletons were in close proximity may suggest the formation of a pack.[191][192] Cooperative pack hunting may have been an effective strategy for subduing prey with advanced anti-predator adaptations which pose potential lethality such as Triceratops and Ankylosaurus.[190]
Currie's pack-hunting T. rex hypothesis has been criticized for not having been peer-reviewed, but rather was discussed in a television interview and book called Dino Gangs.[193] The Currie theory for pack hunting by T. rex is based mainly by analogy to a different species, Tarbosaurus bataar. Evidence of gregariousness in T. bataar itself has not been peer-reviewed, and to Currie's own admission, can only be interpreted with reference to evidence in other closely related species. According to Currie gregariousness in Albertosaurus sarcophagus is supported by the discovery of 26 individuals with varied ages in the Dry Island bonebed. He ruled out the possibility of a predator trap due to the similar preservation state of individuals and the near absence of herbivores.[193][194]
Additional support of tyrannosaurid gregariousness can be found in fossilized trackways from the Upper Cretaceous Wapiti Formation of northeastern British Columbia, Canada, left by three tyrannosaurids traveling in the same direction.[195][196] According to scientists assessing the Dino Gangs program, the evidence for pack hunting in Tarbosaurus and Albertosaurus is weak and based on group skeletal remains for which alternate explanations may apply (such as drought or a flood forcing dinosaurs to die together in one place).[193] Others researchers have speculated that instead of large theropod social groups, some of these finds represent behavior more akin to Komodo dragon-like mobbing of carcasses, even going as far as to say true pack-hunting behavior may not exist in any non-avian dinosaurs due to its rarity in modern predators.[197]
Evidence of intraspecific attack was found by Joseph Peterson and his colleagues in the juvenile Tyrannosaurus nicknamed Jane. Peterson and his team found that Jane's skull showed healed puncture wounds on the upper jaw and snout which they believe came from another juvenile Tyrannosaurus. Subsequent CT scans of Jane's skull would further confirm the team's hypothesis, showing that the puncture wounds came from a traumatic injury and that there was subsequent healing.[198] The team would also state that Jane's injuries were structurally different from the parasite-induced lesions found in Sue and that Jane's injuries were on its face whereas the parasite that infected Sue caused lesions to the lower jaw.[199]
Feeding strategies


Most paleontologists accept that Tyrannosaurus was both an active predator and a scavenger like most large carnivores.[200] By far the largest carnivore in its environment, T. rex was most likely an apex predator, preying upon hadrosaurs, armored herbivores like ceratopsians and ankylosaurs, and possibly sauropods.[201] A study in 2012 by Karl Bates and Peter Falkingham found that Tyrannosaurus had the most powerful bite of any terrestrial animal that has ever lived, finding an adult Tyrannosaurus could have exerted 35,000 to 57,000 N (7,868 to 12,814 lbf) of force in the back teeth.[202][203][204] Even higher estimates were made by Mason B. Meers in 2003.[47] This allowed it to crush bones during repetitive biting and fully consume the carcasses of large dinosaurs.[21] Stephan Lautenschlager and colleagues calculated that Tyrannosaurus was capable of a maximum jaw gape of around 80 degrees, a necessary adaptation for a wide range of jaw angles to power the creature's strong bite.[205][206]
A debate exists, however, about whether Tyrannosaurus was primarily a predator or a pure scavenger. The debate originated in a 1917 study by Lambe which argued that large theropods were pure scavengers because Gorgosaurus teeth showed hardly any wear.[207] This argument disregarded the fact that theropods replaced their teeth quite rapidly. Ever since the first discovery of Tyrannosaurus most scientists have speculated that it was a predator; like modern large predators it would readily scavenge or steal another predator's kill if it had the opportunity.[208]
Paleontologist Jack Horner has been a major proponent of the view that Tyrannosaurus was not a predator at all but instead was exclusively a scavenger.[143][209][210] He has put forward arguments in the popular literature to support the pure scavenger hypothesis:
- Tyrannosaur arms are short when compared to other known predators. Horner argues that the arms were too short to make the necessary gripping force to hold on to prey.[211] Other paleontologists such as Thomas Holtz Jr. argued that there are plenty of modern-day predators that do not use their forelimbs to hunt such as wolves, hyenas, and secretary birds as well as other extinct animals thought to be predators that would not have used their forelimbs such as phorusrhacids.[212][213]
- Tyrannosaurs had large olfactory bulbs and olfactory nerves (relative to their brain size). These suggest a highly developed sense of smell which could sniff out carcasses over great distances, as modern vultures do. Research on the olfactory bulbs of dinosaurs has shown that Tyrannosaurus had the most highly developed sense of smell of 21 sampled dinosaurs.[184]
- Tyrannosaur teeth could crush bone, and therefore could extract as much food (bone marrow) as possible from carcass remnants, usually the least nutritious parts. Karen Chin and colleagues have found bone fragments in coprolites (fossilized feces) that they attribute to tyrannosaurs, but point out that a tyrannosaur's teeth were not well adapted to systematically chewing bone like hyenas do to extract marrow.[214]
- Since at least some of Tyrannosaurus's potential prey could move quickly, evidence that it walked instead of ran could indicate that it was a scavenger.[209] On the other hand, recent analyses suggest that Tyrannosaurus, while slower than large modern terrestrial predators, may well have been fast enough to prey on large hadrosaurs and ceratopsians.[167][24]
Other evidence suggests hunting behavior in Tyrannosaurus. The eye sockets of tyrannosaurs are positioned so that the eyes would point forward, giving them binocular vision slightly better than that of modern hawks. It is not obvious why natural selection would have favored this long-term trend if tyrannosaurs had been pure scavengers, which would not have needed the advanced depth perception that stereoscopic vision provides.[44][45] In modern animals, binocular vision is found mainly in predators.

A skeleton of the hadrosaurid Edmontosaurus annectens has been described from Montana with healed tyrannosaur-inflicted damage on its tail vertebrae. The fact that the damage seems to have healed suggests that the Edmontosaurus survived a tyrannosaur's attack on a living target, i.e. the tyrannosaur had attempted active predation.[215] Despite the consensus that the tail bites were caused by Tyrannosaurus, there has been some evidence to show that they might have been created by other factors. For example, a 2014 study suggested that the tail injuries might have been due to Edmontosaurus individuals stepping on each other,[216] while another study in 2020 backs up the hypothesis that biomechanical stress is the cause for the tail injuries.[217] There is also evidence for an aggressive interaction between a Triceratops and a Tyrannosaurus in the form of partially healed tyrannosaur tooth marks on a Triceratops brow horn and squamosal (a bone of the neck frill); the bitten horn is also broken, with new bone growth after the break. It is not known what the exact nature of the interaction was, though: either animal could have been the aggressor.[218] Since the Triceratops wounds healed, it is most likely that the Triceratops survived the encounter and managed to overcome the Tyrannosaurus. In a battle against a bull Triceratops, the Triceratops would likely defend itself by inflicting fatal wounds to the Tyrannosaurus using its sharp horns.[219] Studies of Sue found a broken and healed fibula and tail vertebrae, scarred facial bones and a tooth from another Tyrannosaurus embedded in a neck vertebra, providing evidence for aggressive behavior.[220] Studies on hadrosaur vertebrae from the Hell Creek Formation that were punctured by the teeth of what appears to be a late-stage juvenile Tyrannosaurus indicate that despite lacking the bone-crushing adaptations of the adults, young individuals were still capable of using the same bone-puncturing feeding technique as their adult counterparts.[221]
Tyrannosaurus may have had infectious saliva used to kill its prey, as proposed by William Abler in 1992. Abler observed that the serrations (tiny protuberances) on the cutting edges of the teeth are closely spaced, enclosing little chambers. These chambers might have trapped pieces of carcass with bacteria, giving Tyrannosaurus a deadly, infectious bite much like the Komodo dragon was thought to have.[222][223] Jack Horner and Don Lessem, in a 1993 popular book, questioned Abler's hypothesis, arguing that Tyrannosaurus's tooth serrations as more like cubes in shape than the serrations on a Komodo monitor's teeth, which are rounded.[143]: 214–215
Tyrannosaurus, and most other theropods, probably primarily processed carcasses with lateral shakes of the head, like crocodilians. The head was not as maneuverable as the skulls of allosauroids, due to flat joints of the neck vertebrae.[224]
Cannibalism
Evidence also strongly suggests that tyrannosaurs were at least occasionally cannibalistic. Tyrannosaurus itself has strong evidence pointing towards it having been cannibalistic in at least a scavenging capacity based on tooth marks on the foot bones, humerus, and metatarsals of one specimen.[225] Fossils from the Fruitland Formation, Kirtland Formation (both Campanian in age) and the Maastrichtian aged Ojo Alamo Formation suggest that cannibalism was present in various tyrannosaurid genera of the San Juan Basin. The evidence gathered from the specimens suggests opportunistic feeding behavior in tyrannosaurids that cannibalized members of their own species.[226] A study from Currie, Horner, Erickson and Longrich in 2010 has been put forward as evidence of cannibalism in the genus Tyrannosaurus.[225] They studied some Tyrannosaurus specimens with tooth marks in the bones, attributable to the same genus. The tooth marks were identified in the humerus, foot bones and metatarsals, and this was seen as evidence for opportunistic scavenging, rather than wounds caused by intraspecific combat. In a fight, they proposed it would be difficult to reach down to bite in the feet of a rival, making it more likely that the bitemarks were made in a carcass. As the bitemarks were made in body parts with relatively scantly amounts of flesh, it is suggested that the Tyrannosaurus was feeding on a cadaver in which the more fleshy parts already had been consumed. They were also open to the possibility that other tyrannosaurids practiced cannibalism.[225]
Parenting
While there is no direct evidence of Tyrannosaurus raising their young (the rarity of juvenile and nest Tyrannosaur fossils has left researchers guessing), it has been suggested by some that like its closest living relatives, modern archosaurs (birds and crocodiles) Tyrannosaurus may have protected and fed its young. Crocodilians and birds are often suggested by some paleontologists to be modern analogues for dinosaur parenting.[227] Direct evidence of parental behavior exists in other dinosaurs such as Maiasaura peeblesorum, the first dinosaur to have been discovered to raise its young, as well as more closely related Oviraptorids, the latter suggesting parental behavior in theropods.[228][229][230][231][232]
Pathology

In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. Of the 81 Tyrannosaurus foot bones examined in the study, one was found to have a stress fracture, while none of the 10 hand bones were found to have stress fractures. The researchers found tendon avulsions only among Tyrannosaurus and Allosaurus. An avulsion injury left a divot on the humerus of Sue the T. rex, apparently located at the origin of the deltoid or teres major muscles. The presence of avulsion injuries being limited to the forelimb and shoulder in both Tyrannosaurus and Allosaurus suggests that theropods may have had a musculature more complex than and functionally different from those of birds. The researchers concluded that Sue's tendon avulsion was probably obtained from struggling prey. The presence of stress fractures and tendon avulsions, in general, provides evidence for a "very active" predation-based diet rather than obligate scavenging.[233]
A 2009 study showed that smooth-edged holes in the skulls of several specimens might have been caused by Trichomonas-like parasites that commonly infect birds. According to the study, seriously infected individuals, including "Sue" and MOR 980 ("Peck's Rex"), might therefore have died from starvation after feeding became increasingly difficult. Previously, these holes had been explained by the bacterious bone infection Actinomycosis or by intraspecific attacks.[234] A subsequent study found that while trichomoniasis has many attributes of the model proposed (osteolytic, intra oral) several features make the assumption that it was the cause of death less supportable by evidence. For example, the observed sharp margins with little reactive bone shown by the radiographs of Trichomonas-infected birds are dissimilar to the reactive bone seen in the affected T. rex specimens. Also, trichomoniasis can be very rapidly fatal in birds (14 days or less) albeit in its milder form, and this suggests that if a Trichomonas-like protozoan is the culprit, trichomoniasis was less acute in its non-avian dinosaur form during the Late Cretaceous. Finally, the relative size of this type of lesions is much larger in small bird throats, and may not have been enough to choke a T. rex.[235] A more recent study examining the pathologies concluded that the osseous alteration observed most closely resembles those around healing human cranial trepanations and healing fractures in the Triassic reptile Stagonolepis, in the absence of infection. The possible cause may instead have been intraspecific combat.[236]
One study of Tyrannosaurus specimens with tooth marks in the bones attributable to the same genus was presented as evidence of cannibalism.[225] Tooth marks in the humerus, foot bones and metatarsals, may indicate opportunistic scavenging, rather than wounds caused by combat with another T. rex.[225][237] Other tyrannosaurids may also have practiced cannibalism.[225]
Paleoecology

Tyrannosaurus lived during what is referred to as the Lancian faunal stage (Maastrichtian age) at the end of the Late Cretaceous. Tyrannosaurus ranged from Canada in the north to at least New Mexico in the south of Laramidia.[5] During this time Triceratops was the major herbivore in the northern portion of its range, while the titanosaurian sauropod Alamosaurus "dominated" its southern range. Tyrannosaurus remains have been discovered in different ecosystems, including inland and coastal subtropical, and semi-arid plains.

Several notable Tyrannosaurus remains have been found in the Hell Creek Formation. During the Maastrichtian this area was subtropical, with a warm and humid climate. The flora consisted mostly of angiosperms, but also included trees like dawn redwood (Metasequoia) and Araucaria. Tyrannosaurus shared this ecosystem with ceratopsians Leptoceratops, Torosaurus, and Triceratops, the hadrosaurid Edmontosaurus annectens, the parksosaurid Thescelosaurus, the ankylosaurs Ankylosaurus and Denversaurus, the pachycephalosaurs Pachycephalosaurus and Sphaerotholus, and the theropods Ornithomimus, Struthiomimus, Acheroraptor, Dakotaraptor, Pectinodon and Anzu.[238]
Another formation with Tyrannosaurus remains is the Lance Formation of Wyoming. This has been interpreted as a bayou environment similar to today's Gulf Coast. The fauna was very similar to Hell Creek, but with Struthiomimus replacing its relative Ornithomimus. The small ceratopsian Leptoceratops also lived in the area.[239]
In its southern range, specifically based on remains discovered from the North Horn Formation of Utah, Tyrannosaurus rex lived alongside the titanosaur Alamosaurus, the ceratopsid Torosaurus and the indeterminate troodontids and hadrosaurids.[240][241] Tyrannosaurus mcraeensis from the McRae Group of New Mexico coexisted with the ceratopsid Sierraceratops and possibly the titanosaur Alamosaurus.[67] Potential remains identified as cf. Tyrannosaurus have also been discovered from the Javelina Formation of Texas,[67] where the remains of the titanosaur Alamosaurus, the ceratopsid Bravoceratops, the pterosaurs Quetzalcoatlus and Wellnhopterus, and possible species of troodontids and hadrosaurids are found.[242][243][244] Its southern range is thought to have been dominated by semi-arid inland plains, following the probable retreat of the Western Interior Seaway as global sea levels fell.[245]
Tyrannosaurus may have also inhabited Mexico's Lomas Coloradas Formation in Sonora. Though skeletal evidence is lacking, six shed and broken teeth from the fossil bed have been thoroughly compared with other theropod genera and appear to be identical to those of Tyrannosaurus. If true, the evidence indicates the range of Tyrannosaurus was possibly more extensive than previously believed.[246] It is possible that tyrannosaurs were originally Asian species, migrating to North America before the end of the Cretaceous period.[247]
Population estimates
According to studies published in 2021 by Charles Marshall et al., the total population of adult Tyrannosaurus at any given time was perhaps 20,000 individuals, with computer estimations also suggesting a total population no lower than 1,300 and no higher than 328,000. The authors themselves suggest that the estimate of 20,000 individuals is probably lower than what should be expected, especially when factoring in that disease pandemics could easily wipe out such a small population. Over the span of the genus' existence, it is estimated that there were about 127,000 generations and that this added up to a total of roughly 2.5 billion animals until their extinction.[248][249]
In the same paper, it is suggested that in a population of Tyrannosaurus adults numbering 20,000, the number of individuals living in an area the size of California could be as high as 3,800 animals, while an area the size of Washington D.C. could support a population of only two adult Tyrannosaurus. The study does not take into account the number of juvenile animals in the genus present in this population estimate due to their occupation of a different niche than the adults, and thus it is likely the total population was much higher when accounting for this factor. Simultaneously, studies of living carnivores suggest that some predator populations are higher in density than others of similar weight (such as jaguars and hyenas, which are similar in weight but have vastly differing population densities). Lastly, the study suggests that in most cases, only one in 80 million Tyrannosaurus would become fossilized, while the chances were likely as high as one in every 16,000 of an individual becoming fossilized in areas that had more dense populations.[248][249]
Meiri (2022) questioned the reliability of the estimates, citing uncertainty in metabolic rate, body size, sex and age-specific survival rates, habitat requirements and range size variability as shortcomings Marshall et al. did not take into account.[250] The authors of the original publication replied that while they agree that their reported uncertainties were probably too small, their framework is flexible enough to accommodate uncerainty in physiology, and that their calculations do not depend on short-term changes in population density and geographic range, but rather on their long-term averages. Finally, they remark that they did estimate the range of reasonable survivorship curves and that they did include uncertainty in the time of onset of sexual maturity and in the growth curve by incorporating theuncertainty in the maximum body mass.[251]
Cultural significance
Since it was first described in 1905, T. rex has become the most widely recognized dinosaur species in popular culture. It is the only dinosaur that is commonly known to the general public by its full scientific name (binomial name) and the scientific abbreviation T. rex has also come into wide usage.[50] Robert T. Bakker notes this in The Dinosaur Heresies and explains that, "a name like 'T. rex' is just irresistible to the tongue."[38]
См. также
- История палеонтологии
- Сью (динозавр) (FMNH-PR-2081)
- Тираннозавриды
Примечания
- ^ букв . ' ящерица-тиран ' ; от древнегреческого τύραννος ( túrannos ) «тиран» и σαῦρος ( saûros ) «ящерица»
Ссылки
- ^ «Стенограмма Колорадо». 8 июля 1874 г. - через www.coloradohistoricnewspapers.org.
- ^ Jump up to: а б Брайтаупт, Британская Колумбия; Саутвелл, штат Эйха; Мэтьюз, Северная Каролина (15 октября 2005 г.). «В честь 100-летия тираннозавра рекса : Manospondylus gigas , Ornithomimus grandis и Dynamosaurus imperiosus , самых ранних открытий тираннозавра рекса на Западе» . Рефераты с программами; 2005 Ежегодное собрание в Солт-Лейк-Сити . 37 (7). Геологическое общество Америки : 406. ISSN 0016-7592 . Архивировано из оригинала 30 мая 2012 года . Проверено 8 октября 2008 г.
- ^ Хэтчер, Дж. Б. (1907). «Цератопсия» . Монографии Геологической службы США . 49 : 113–114. ISSN 0886-7550 .
- ^ Jump up to: а б Осборн, HF (1917). «Скелетные адаптации орнитолестов , струтиомимов , тираннозавров ». Бюллетень Американского музея естественной истории . 35 (43): 733–771. hdl : 2246/1334 .
- ^ Jump up to: а б с д и ж г Ларсон, Нидерланды (2008). «Сто лет тираннозавра рекса : скелеты». В Ларсоне, П.; Карпентер, К. (ред.). Тираннозавр рекс , Король-Тиран . Блумингтон, Индиана: Издательство Университета Индианы. стр. 1 –55. ISBN 978-0-253-35087-9 .
- ^ Jump up to: а б с д Осборн, HF (1905). « Тираннозавр и другие хищные динозавры мелового периода». Бюллетень АМНХ . 21 (14): 259–265. hdl : 2246/1464 . Проверено 6 октября 2008 г.
- ^ Jump up to: а б Дингус, Л.; Норелл, М. (3 мая 2010 г.). Барнум Браун: Человек, открывший тираннозавра рекса . Издательство Калифорнийского университета. стр. 90 , 124. ISBN. 978-0-520-94552-4 .
- ^ Jump up to: а б Осборн, Флорида ; Браун, Б. (1906). « Тираннозавр , плотоядный динозавр верхнего мела». Бюллетень АМНХ . 22 (16): 281–296. hdl : 2246/1473 .
- ^ Брайтаупт, Британская Колумбия; Саутвелл, штат Эйха; Мэтьюз, Северная Каролина (2006). Лукас, СГ; Салливан, Р.М. (ред.). « Dynamosaurus imperiosus и самые ранние открытия тираннозавра рекса в Вайоминге и на Западе» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 258.
Оригинальный скелет Dynamosaurus imperiosus (AMNH 5866/BM R7995) вместе с другими материалами тираннозавра (включая части AMNH 973, 5027 и 5881) был продан Британскому музею естественной истории (ныне The Музей естественной истории) в 1960 году. Этот материал был использован в интересной «полумонтовой» экспозиции этого динозавра в Лондоне. В настоящее время материал находится в научных коллекциях.
- ^ Макдональд, AT; Вульф, генеральный директор; Дули, AC младший (2018). «Новый тираннозаврид (Dinosauria: Theropoda) из верхнемеловой формации Менефи в Нью-Мексико» . ПерДж . 6 :6:e5749. дои : 10.7717/peerj.5749 . ПМК 6183510 . ПМИД 30324024 .
- ^ Смолл, Закари (7 октября 2020 г.). «Скелет Ти Рекса принес на аукционе Christie's 31,8 миллиона долларов» . Нью-Йорк Таймс . Архивировано из оригинала 7 октября 2020 года . Проверено 5 мая 2021 г.
- ^ «Подготовка костей Сью» . Сью в Полевом музее . Полевой музей. 2007 . Проверено 24 октября 2014 г.
- ^ Эриксон, Г.; Маковицкий, П.Дж.; Карри, Пи Джей; Норелл, М.; Йерби, С.; Брошу, Калифорния (26 мая 2004 г.). «Гигантизм и параметры истории жизни динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 .
- ^ «Стэн» . Манчестерский университет . 18 сентября 2010 г. Архивировано из оригинала 18 сентября 2010 г.
- ^ Файффер, С. (2000). «Юрский фарс». Тираннозавр Сью . WH Freeman and Company, Нью-Йорк. стр. 121–122 . ISBN 978-0-7167-4017-9 .
- ^ «Знакомьтесь, Баки, тираннозавр-подросток» . Детский музей Индианаполиса . 7 июля 2014. Архивировано из оригинала 27 декабря 2014 года . Проверено 2 декабря 2019 г.
- ^ «Раскопки выявили пять экземпляров тираннозавра» . Новости Би-би-си . 10 октября 2000 г. Проверено 13 декабря 2008 г.
- ^ Jump up to: а б Карри, Пи Джей; Хурум, Дж. Х.; Сабат, К. (2003). «Строение черепа и эволюция динозавров-тираннозаврид» (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234 . Проверено 8 октября 2008 г.
- ^ Блэк, Райли (28 октября 2015 г.). «Крошечный ужас: спорный вид динозавров — это всего лишь неуклюжий подросток -тираннозавр » . Смитсоновский журнал . Проверено 10 декабря 2018 г.
- ^ Jump up to: а б «Музей представляет самый большой в мире череп тиранозавра » . 2006. Архивировано из оригинала 14 апреля 2006 года . Проверено 7 апреля 2006 г.
- ^ Jump up to: а б Жиньяк, премьер-министр; Эриксон, генеральный директор (2017). «Биомеханика крайней остеофагии у тираннозавра рекса » . Научные отчеты . 7 (1): 2012. Бибкод : 2017НацСР...7.2012Г . дои : 10.1038/s41598-017-02161-w . ПМЦ 5435714 . ПМИД 28515439 .
- ^ Локли, МГ; Хант, AP (1994). «След гигантского динозавра-теропода тираннозавра недалеко от границы мелового и третичного периодов, север Нью-Мексико». Ихнос . 3 (3): 213–218. Бибкод : 1994Ично...3..213Л . дои : 10.1080/10420949409386390 .
- ^ «Вероятный след тираннозаврид из формации Хелл-Крик (верхний мел), Монтана, США» . Новости Национального исторического музея . 2007. Архивировано из оригинала 14 декабря 2007 года . Проверено 18 декабря 2007 г.
- ^ Jump up to: а б Мэннинг, Польша; Отт, К.; Фолкингем, Польша (2009 г.). «Первый след тираннозаврид из формации Хелл-Крик (поздний мел), Монтана, США». ПАЛЕОС . 23 (10): 645–647. Бибкод : 2008Палай..23..645М . дои : 10.2110/palo.2008.p08-030r . S2CID 129985735 .
- ^ Смит, С.Д.; Лица, WS; Син, Л. (2016). «Тропа «Тираннозавра» в Гленроке, Ланс-Формейшн (Маастрихт), Вайоминг» . Меловые исследования . 61 (1): 1–4. Бибкод : 2016CrRes..61....1S . дои : 10.1016/j.cretres.2015.12.020 .
- ^ Перкинс, С. (2016). «Возможно, вы могли бы обогнать тираннозавра » . Палеонтология . дои : 10.1126/science.aae0270 .
- ^ Уолтон, Т. (2016). «Забудьте все, что вы знали из «Парка Юрского периода»: по скорости тираннозавр превосходит велоцираптора » . США сегодня . Проверено 13 марта 2016 г.
- ^ Руис, Дж. (2017). «Комментарии к «Следу тираннозавра в Гленроке, формация Лэнс (Маастрихт), Вайоминг» (Смит и др. , Cretaceous Research , т. 61, стр. 1–4, 2016)». Меловые исследования . 82 : 81–82. дои : 10.1016/j.cretres.2017.05.033 .
- ^ Jump up to: а б с д Хатчинсон-младший; Бейтс, КТ; Молнар, Дж.; Аллен, В.; Маковицкий, П.Дж. (2011). «Вычислительный анализ размеров конечностей и тела тираннозавра рекса с последствиями для передвижения, онтогенеза и роста» . ПЛОС ОДИН . 6 (10): e26037. Бибкод : 2011PLoSO...626037H . дои : 10.1371/journal.pone.0026037 . ПМК 3192160 . ПМИД 22022500 .
- ^ Хольц, ТР (2011). «Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, приложение, зима 2011 г.» (PDF) . Проверено 13 января 2012 г.
- ^ Jump up to: а б «Информационный бюллетень о Сью» (PDF) . Сью в Полевом музее . Полевой музей естественной истории . Архивировано из оригинала (PDF) 18 августа 2016 г.
- ^ «Насколько хорошо ты знаешь Сью?» . Полевой музей естественной истории. 11 августа 2016 года . Проверено 31 декабря 2018 г.
- ^ «Сью на Ти Рекса» . Полевой музей . 5 февраля 2018 г. . Проверено 20 июля 2018 г.
- ^ Jump up to: а б Лица, SW; Карри, Пи Джей; Эриксон, генеральный директор (2019). «Старый и исключительно крупный взрослый экземпляр тираннозавра рекса » . Анатомическая запись . 303 (4): 656–672. дои : 10.1002/ar.24118 . ISSN 1932-8486 . ПМИД 30897281 .
- ^ Хартман, Скотт (7 июля 2013 г.). «Массовые оценки: сокращение Севера и Юга» . Skeletal Drawing.com Скотта Хартмана. Архивировано из оригинала 12 октября 2013 года . Проверено 24 августа 2013 г.
- ^ Лайл, А. (22 марта 2019 г.). «Палеонтологи определили самый крупный из когда-либо обнаруженных тираннозавров » . Фолио, Университет Альберты . Проверено 25 марта 2019 г.
- ^ Андерсон, Дж. Ф.; Холл-Мартин, AJ; Рассел, Д. (1985). «Окружность и вес длинных костей у млекопитающих, птиц и динозавров». Журнал зоологии . 207 (1): 53–61. дои : 10.1111/j.1469-7998.1985.tb04915.x .
- ^ Jump up to: а б с Баккер, RT (1986). Ересь динозавров . Нью-Йорк: Издательство Кенсингтон. п. 241 . ISBN 978-0-688-04287-5 . OCLC 13699558 .
- ^ Хендерсон, DM (1 января 1999 г.). «Оценка массы и центров масс вымерших животных методом трехмерного математического нарезки» . Палеобиология . 25 (1): 88–106.
- ^ Jump up to: а б с Эриксон, генеральный директор; Маковицкий, П.Дж.; Карри, Пи Джей ; Норелл, Массачусетс; Йерби, ЮАР; Брочу, Калифорния (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 .
- ^ Фарлоу, Джо; Смит, МБ; Робинсон, Дж. М. (1995). «Масса тела, «индикатор прочности» костей и беговой потенциал тираннозавра рекса » . Журнал палеонтологии позвоночных . 15 (4): 713–725. Бибкод : 1995JVPal..15..713F . дои : 10.1080/02724634.1995.10011257 . Архивировано из оригинала 23 октября 2008 года.
- ^ Сибахер, Ф. (2001). «Новый метод расчета аллометрических отношений длины и массы динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. CiteSeerX 10.1.1.462.255 . doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 . S2CID 53446536 .
- ^ Кристиансен, П.; Фаринья, РА (2004). «Массовое предсказание динозавров-теропод». Историческая биология . 16 (2–4): 85–92. Бибкод : 2004HBio...16...85C . дои : 10.1080/08912960412331284313 . S2CID 84322349 .
- ^ Jump up to: а б с Стивенс, Кент А. (июнь 2006 г.). «Бинокулярное зрение тероподовых динозавров». Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 . S2CID 85694979 .
- ^ Jump up to: а б с Яффе, Э. (1 июля 2006 г.). «Зрение для глаз Саура: зрение тираннозавра было одним из лучших в природе» . Новости науки . 170 (1): 3–4. дои : 10.2307/4017288 . JSTOR 4017288 . Архивировано из оригинала 29 сентября 2012 года . Проверено 6 октября 2008 г.
- ^ Снайвли, Э.; Хендерсон, DM; Филлипс, Д.С. (2006). «Сросшиеся и сводчатые носовые кости динозавров-тираннозаврид: влияние на силу черепа и механику питания» (PDF) . Acta Palaeontologica Polonica . 51 (3): 435–454 . Проверено 8 октября 2008 г.
- ^ Jump up to: а б Меерс, МБ (август 2003 г.). «Максимальная сила укуса и размер добычи тираннозавра рекса и их связь с выводами о пищевом поведении». Историческая биология . 16 (1): 1–12. дои : 10.1080/0891296021000050755 . S2CID 86782853 .
- ^ Эриксон, генеральный директор; Ван Кирк, SD; Су, Дж.; Левенстон, Мэн; Калер, МЫ; Картер, доктор медицинских наук (1996). «Оценка силы укуса тираннозавра рекса по костям с отметинами зубов» . Природа . 382 (6593): 706–708. Бибкод : 1996Natur.382..706E . дои : 10.1038/382706a0 . S2CID 4325859 .
- ^ Jump up to: а б с Хольц, Т.Р. (1994). «Филогенетическое положение тираннозаврид: значение для систематики теропод». Журнал палеонтологии . 68 (5): 1100–1117. Бибкод : 1994JPal...68.1100H . дои : 10.1017/S0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Брошу, ЧР (2003). «Остеология тираннозавра рекса : данные по почти полному скелету и компьютерному томографическому анализу черепа с высоким разрешением». Мемуары Общества палеонтологии позвоночных . 7 : 1–138. дои : 10.2307/3889334 . JSTOR 3889334 .
- ^ Смит, Дж. Б. (1 декабря 2005 г.). «Гетеродонтия у тираннозавра рекса : последствия для таксономической и систематической полезности зубных рядов теропод». Журнал палеонтологии позвоночных . 25 (4): 865–887. doi : 10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2 . S2CID 86184190 .
- ^ Дуглас, К.; Янг, С. (1998). «Детективы о динозаврах» . Новый учёный . Проверено 16 октября 2008 г.
Один палеонтолог незабываемо описал огромные изогнутые зубы тираннозавра как «смертельные бананы».
- ^ «Жизненная статистика Сью» . Сью в Полевом музее . Полевой музей естественной истории . Архивировано из оригинала 29 сентября 2007 года . Проверено 15 сентября 2007 г.
- ^ Jump up to: а б с д Карр, Т.Д.; Варриккио, диджей; Седлмайр, Дж. К.; Робертс, EM; Мур-младший (30 марта 2017 г.). «Новый тираннозавр с доказательствами анагенеза и сенсорной системой лица, подобной крокодиловой» . Научные отчеты . 7 : 44942. Бибкод : 2017NatSR...744942C . дои : 10.1038/srep44942 . ISSN 2045-2322 . ПМК 5372470 . ПМИД 28358353 .
- ^ Морхардт, Эшли (2009). Улыбка динозавра: обеспечивают ли текстура и морфология предчелюстной, верхней и зубной костей зауропсидов остеологические корреляты, позволяющие надежно сделать вывод о внеротовых структурах у динозавров? (Магистерская диссертация). Университет Западного Иллинойса.
- ^ Jump up to: а б Рейс, РР; Ларсон, Д. (2016). «Анатомия зубов и соотношение длины черепа и размера зубов подтверждают гипотезу о том, что у динозавров-теропод были губы» . 4-е ежегодное собрание, 2016 г., Канадское общество палеонтологии позвоночных . ISSN 2292-1389 .
- ^ МОРФОЛОГИЯ, ТАКСОНОМИЯ И ФИЛОГЕНЕТИЧЕСКИЕ СВЯЗИ МОНТЕВИАЛЬНЫХ КРОКОДИЛОВ (ОЛИГОЦЕН, ИТАЛИЯ) . 2018. с. 67 . Проверено 9 октября 2020 г.
- ^ Jump up to: а б Каллен, Томас М.; Ларсон, Дерек В.; Виттон, Марк П.; Скотт, Дайан; Махо, Чай; Бринк, Кирстин С.; Эванс, Дэвид С.; Рейс, Роберт (31 марта 2023 г.). «Реконструкция лица динозавра-теропода и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Бибкод : 2023Sci...379.1348C . дои : 10.1126/science.abo7877 . ISSN 0036-8075 . ПМИД 36996202 . S2CID 257836765 .
- ^ Липкин, К.; Карпентер, К. (2008). «Снова глядя на переднюю конечность тираннозавра рекса ». В Карпентере, К.; Ларсон, ЧП (ред.). Тираннозавр рекс , Король-Тиран . Блумингтон: Издательство Университета Индианы. стр. 167–190 . ISBN 978-0-253-35087-9 .
- ^ Jump up to: а б с Хольц, Т.Р. младший (2004). «Тираннозавроидея». Вейшампель , DB ; Додсон, П .; Осмольска, Х. (ред.). Динозаврия . Беркли: Издательство Калифорнийского университета. стр. 111–136 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б Пол, Грегори С. (1988). Хищные динозавры мира: полный иллюстрированный справочник . Нью-Йорк: Саймон и Шустер. п. 228 . ISBN 978-0-671-61946-6 . OCLC 18350868 .
- ^ Брусатте, Стивен Л.; Норелл, Марк А.; Карр, Томас Д.; Эриксон, Грегори М.; Хатчинсон, Джон Р.; Баланофф, Эми М.; Бевер, Гейб С.; Шуаньер, Иона Н.; Маковицкий, Питер Дж.; Сюй, Син (17 сентября 2010 г.). «Палеобиология тираннозавров: новые исследования древних образцовых организмов» . Наука . 329 (5998): 1481–1485. Бибкод : 2010Sci...329.1481B . дои : 10.1126/science.1193304 . ISSN 0036-8075 . ПМИД 20847260 .
- ^ Ли, Дацин; Норелл, Марк А.; Гао, Кэ-Цинь; Смит, Натан Д.; Маковицкий, Питер Дж. (2009). «Лонгиростровый тираннозавроид из раннего мела Китая» . Учебник по биологическим наукам . 277 (1679): 183–190. дои : 10.1098/rspb.2009.0249 . ПМЦ 2842666 . ПМИД 19386654 .
- ^ Jump up to: а б Лоуэн, Массачусетс; Ирмис, РБ; Сертич, JJW; Карри, Пи Джей ; Сэмпсон, SD (2013). Эванс, округ Колумбия (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение позднемеловых океанов» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- ^ Вергано, Д. (7 ноября 2013 г.). «Новообретенный динозавр, «король крови», правил до Ти Рекса» . Нэшнл Географик . Архивировано из оригинала 8 ноября 2013 года . Проверено 10 ноября 2017 г.
- ^ Геггель, Л. (29 февраля 2016 г.). « Ти-Рекс, вероятно, был инвазивным видом» . Живая наука . Проверено 10 ноября 2017 г.
- ^ Jump up to: а б с д и ж Далман, Себастьян Г.; Лоуэн, Марк А.; Пайрон, Р. Александр; Ясински, Стивен Э.; Малинзак, Д. Эдвард; Лукас, Спенсер Г.; Фиорилло, Энтони Р.; Карри, Филип Дж.; Лонгрич, Николас Р. (11 января 2024 г.). «Гигантский тираннозавр из кампана-маастрихта на юге Северной Америки и эволюция гигантизма тираннозаврид» . Научные отчеты . 13 (1): 22124. doi : 10.1038/s41598-023-47011-0 . ISSN 2045-2322 . ПМЦ 10784284 . ПМИД 38212342 .
- ^ Шерер, Чарли Роджер; Войкулеску-Холвад, Кристиан (2024). «Повторный анализ набора данных опровергает утверждения об анагенезе у тираннозавров линии Tyrannosaurus (Theropoda, Tyrannosauridae)» . Меловые исследования . 155 . 105780. Бибкод : 2024CrRes.15505780S . дои : 10.1016/j.cretres.2023.105780 . ISSN 0195-6671 .
- ^ Малеев, Э. А. (1955). «(название на русском языке)» [Гигантские хищные динозавры Монголии] (PDF) . Доклады Академии наук СССР . 104 (4). перевод Ф. Дж. Алкока: 634–637.
- ^ Рождественский, А.К. (1965). «Изменения роста азиатских динозавров и некоторые проблемы их систематики». Палеонтологический журнал . 3 : 95–109.
- ^ Карпентер, К. (1992). «Тираннозавриды (Динозавры) Азии и Северной Америки». В Матере, штат Нью-Джерси; Чен, П.-дж. (ред.). Аспекты неморской меловой геологии . Пекин: China Ocean Press. стр. 250–268. ISBN 978-7-5027-1463-5 . ОСЛК 28260578 .
- ^ Хурум Дж. Х., Сабат К. (2003). «Гигантские динозавры-тероподы из Азии и Северной Америки: сравнение черепов Tarbosaurus bataar и Tyrannosaurus rex » (PDF) . Acta Palaeontologica Polonica . 48 (2): 161–190. Архивировано из оригинала (PDF) 17 мая 2023 года.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Ли Фэн, Би Шуньдун, Майкл Питтман, Стивен Л. Брусатте, Сюй Син (2016). «Новый экземпляр тираннозавра (Theropoda: Tyrannosauroidea) с отверстиями насекомых из верхнемеловой формации Хунлишан на северо-западе Китая» . Меловые исследования . 66 : 155–162. Бибкод : 2016CrRes..66..155L . дои : 10.1016/j.cretres.2016.06.002 . hdl : 20.500.11820/a49b0878-3ba2-4c3e-b4b1-70fe282e43ea .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Себастьян Г. Далман, Спенсер Г. Лукас (январь 2018 г.). «Динозавры-тираннозавриды (Theropoda: Tyrannosauridae) из верхнего мела (ранний кампан) члена Эллисона формации Менефи, Нью-Мексико: значение для происхождения тираннозаврид в Северной Америке» . Ископаемый рекорд . Бюллетень Музея естественной истории и науки Нью-Мексико. 6 (79): 99–112.
- ^ А.О. Аверьянов, А.А. Ярков (2004). «Плотоядные динозавры (Saurischia, Theropoda) из маастрихта Волго-Донского междуречья, Россия» (PDF) . Палеонтологический журнал . 38 (1): 78–82. Архивировано (PDF) из оригинала 5 марта 2024 г.
- ^ A. O. Averianov, A. V. Lopatin (2023). "Динозавры России: обзор местонахождений [Dinosaurs of Russia: Overview of locations]" (PDF) . Vestnik Rossiiskoi Akademii Nauk (in Russian). 93 (4): 342–354. doi : 10.31857/S0869587323040023 . Archived (PDF) from the original on February 16, 2024.
- ^ Хон, DWE; Ван, К.; Салливан, К.; Чжао, X.; Чен, С.; Ли, Д.; Джи, С.; Джи, К.; Сюй, X. (2011). «Новый крупный теропод-тираннозавр из верхнего мела Китая». Меловые исследования . 32 (4): 495–503. Бибкод : 2011CrRes..32..495H . дои : 10.1016/j.cretres.2011.03.005 .
- ^ Пол, Грегори С.; Лица IV, В. Скотт; ван Раалте, Джей (2022). «Король, королева и император-тиран-ящерица: многочисленные линии морфологических и стратиграфических данных подтверждают тонкую эволюцию и вероятное видообразование в пределах североамериканского рода тираннозавров» . Эволюционная биология . 49 (2): 156–179. Бибкод : 2022EvBio..49..156P . дои : 10.1007/s11692-022-09561-5 . S2CID 247200214 .
- ^ Jump up to: а б Эльбейн, Ашер (28 февраля 2022 г.). «Они хотят разделить тираннозавра на 3 вида. Другие палеонтологи недовольны» . Нью-Йорк Таймс . Архивировано из оригинала 1 марта 2022 года . Проверено 1 марта 2022 г.
- ^ Хант, Кэти (1 марта 2022 г.). «Тираннозавра рекса, возможно, неправильно поняли» . CNN .
- ^ Грешко, Михаил (1 марта 2022 г.). «Призыв разделить тираннозавра на три вида вызывает ожесточенные дебаты» . Нэшнл Географик . Архивировано из оригинала 1 марта 2022 года.
- ^ Карр, Т.Д.; Наполи, JG; Брусатте, СЛ; Хольц, ТР; Хон, DWE; Уильямсон, TE; Занно, LE (2022). «Недостаточно доказательств существования нескольких видов тираннозавров в последнем меловом периоде Северной Америки: комментарий к книге «Король, королева и император ящеров-тиранов: многочисленные линии морфологических и стратиграфических данных подтверждают тонкую эволюцию и вероятное видообразование в пределах североамериканского рода тираннозавров »» . Эволюционная биология . 49 (3): 314–341. Бибкод : 2022EvBio..49..327C . дои : 10.1007/s11692-022-09573-1 .
- ^ Осборн М. «Исследование опровергает противоречивые исследования, которые разделили тираннозавра на три вида» . Журнал Smithsonian, 27 июля 2022 г.
- ^ Пол, Грегори С. (5 августа 2022 г.). «Наблюдения за определением палеовидов, с дополнительными данными о тираннозавре , включая его сильно различающиеся виды, специфические надглазничные украшения, которые придают тираннозаврам новый и уникальный жизненный облик». bioRxiv (Препринт). дои : 10.1101/2022.08.02.502517 . S2CID 251474294 .
- ^ Лозинский, Ричард П.; Хант, Адриан П.; Вольберг, Дональд Л.; Лукас, Спенсер Г. (1984). «Позднемеловые (ланкийские) динозавры из формации Макрей, округ Сьерра, Нью-Мексико» (PDF) . Геология Нью-Мексико . 6 (4): 72–77. дои : 10.58799/NMG-v6n4.72 . ISSN 0196-948X . S2CID 237011797 .
- ^ Джилетт, Дэвид Д.; Вольберг, Дональд Л.; Хант, Адриан П. (1986). « Тираннозавр рекс из формации Макрей (Лансиан, верхний мел), водохранилище Элефант-Бьютт, округ Сьерра, Нью-Мексико» (PDF) . Путеводитель геологического общества Нью-Мексико . 37 : 235–238. дои : 10.56577/FFC-37.235 . S2CID 251985284 .
- ^ Леман, Томас М.; Карпентер, Кеннет (1990). «Частичный скелет динозавра-тираннозавра Aublysodon из верхнего мела Нью-Мексико» . Журнал палеонтологии . 64 (6): 1026–1032. Бибкод : 1990JPal...64.1026L . дои : 10.1017/S0022336000019843 . JSTOR 1305741 . S2CID 132662000 .
- ^ Карр, Томас Д.; Уильямсон, Томас Э. (2000). «Обзор тираннозаврид (Dinosauria: Coelurosauria) из Нью-Мексико» . Бюллетень . 17 . Музей естественной истории и науки Нью-Мексико : 113–145.
- ^ Салливан, Роберт М.; Лукас, Спенсер Г. (2015). «Меловые позвоночные Нью-Мексико» . Бюллетень Музея естественной истории и науки Нью-Мексико . 68 .
- ^ Шанц, Эмма Э.; Амато, Джеффри М. (2024). «Возрастные ограничения позднемеловых динозавров: геохронологический и стратиграфический анализ формации Макрей в бассейне ранчо Ларамид Лав, Нью-Мексико». Геологическое общество Нью-Мексико . дои : 10.56577/SM-2024.3002 .
- ^ Эльбейн, Ашер (11 января 2024 г.). «Новая история происхождения тираннозавра рекса , предложенная ископаемыми. Исследователи говорят, что вид, который они назвали Tyrannosaurus mcraeensis, предшествовал великому хищнику эпохи динозавров» . Нью-Йорк Таймс . Архивировано из оригинала 12 января 2024 года . Проверено 12 января 2024 г.
- ^ Ольшевский, Г. (1995). «Происхождение и эволюция тираннозаврид». Кёрюгаку Сайзенсен [Dino Frontline] . 9–10 : 92–119.
- ^ Карр, Т.Д.; Уильямсон, Т.Э. (2004). «Разнообразие позднемаастрихтских тираннозаврид (Dinosauria: Theropoda) из западной части Северной Америки» . Зоологический журнал Линнеевского общества . 142 (4): 479–523. дои : 10.1111/j.1096-3642.2004.00130.x .
- ^ Гилмор, CW (1946). «Новый плотоядный динозавр из формации Ланс в Монтане». Разные коллекции Смитсоновского института . 106 : 1–19.
- ^ Баккер, RT; Уильямс, М.; Карри, Пи Джей (1988). « Nanotyrannus , новый род карликовых тираннозавров из позднего мелового периода Монтаны». Хантерия . 1 :1–30.
- ^ Jump up to: а б Карр, Т.Д. (1999). «Черепно-лицевой онтогенез тираннозаврид (Dinosauria, Coelurosauria)» . Журнал палеонтологии позвоночных . 19 (3): 497–520. Бибкод : 1999JVPal..19..497C . дои : 10.1080/02724634.1999.10011161 . S2CID 83744433 .
- ^ Jump up to: а б Цуихиджи, Т.; Ватабе, М.; Цогтбаатар, К.; Цубамото, Т.; Барсболд, Р.; Сузуки, С.; Ли, АХ; Риджли, Колорадо; Кавахара, Ю.; Витмер, LM (2011). «Краниальная остеология молодого экземпляра Tarbosaurus bataar из нэмегтинской свиты (верхний мел) Бугин Цав, Монголия». Журнал палеонтологии позвоночных . 31 (3): 497–517. Бибкод : 2011JVPal..31..497T . дои : 10.1080/02724634.2011.557116 . S2CID 15369707 .
- ^ Карри, Пи Джей (2003a). «Краниальная анатомия динозавров-тираннозаврид из позднего мела Альберты, Канада». Acta Palaeontologica Polonica . 48 : 191–226.
- ^ Карри, Хендерсон, Хорнер и Уильямс (2005). «О зубах тираннозавра, положении зубов и таксономическом статусе Nanotyrannus lancensis ». Симпозиум «Происхождение, систематика и палеобиология тираннозаврид», организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Хендерсон (2005). «Nano No More: Смерть карликового тирана». Симпозиум «Происхождение, систематика и палеобиология тираннозаврид», организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Ларсон (2005). «Дело Нанотирануса ». Симпозиум «Происхождение, систематика и палеобиология тираннозаврид», организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Ларсон П. (2013), «Достоверность Nanotyrannus Lancensis (Theropoda, Lancian – верхний маастрихт Северной Америки)», Общество палеонтологии позвоночных: 73-е ежегодное собрание, Рефераты с программами , стр. 159.
- ^ Д. Карр, Томас (15 сентября 2013 г.). «Нанотиранус на самом деле не настоящий» . Тираннозавроида Центральная . Проверено 28 мая 2019 г.
- ^ Лица, WS; Карри, Пи Джей (2016). «Подход к оценке пропорций конечностей хищных динозавров и попытка объяснить аллометрию» . Научные отчеты . 6 . 19828. Бибкод : 2016НатСР...619828П . дои : 10.1038/srep19828 . ПМЦ 4728391 . ПМИД 26813782 .
- ^ «Пропорции задних конечностей не подтверждают достоверность Нанотирануса» . mambobob-raptorsnest.blogspot.com .
- ^ Шмерге, Джошуа Д.; Ротшильд, Брюс М. (2016). «Распределение зубной борозды динозавров-теропод: значение для филогении теропод и достоверность рода Nanotyrannus Bakker et al., 1988». Меловые исследования . 61 : 26–33. Бибкод : 2016CrRes..61...26S . doi : 10.1016/J.CRETRES.2015.12.016 . S2CID 85736236 .
- ^ Брусатте, Стивен Л.; Карр, Томас Д.; Уильямсон, Томас Э.; Хольц, Томас Р.; Хоун, Дэвид МЫ; Уильямс, Скотт А. (2016). «Морфология зубных бороздок не позволяет отличить Nanotyrannus как действительный таксон тираннозавроидных динозавров. Комментарий к: «Распределение зубных бороздок тероподных динозавров: значение для филогении теропод и достоверность рода Nanotyrannus Bakker et al., 1988 » (PDF) . Меловые исследования . 65 : 232–237. Бибкод : 2016CrRes..65..232B . doi : 10.1016/J.CRETRES.2016.02.007 . hdl : 20.500.11820/f1e76074-47eb-4c25-b4c1-a3782551fd5a . S2CID 56090258 .
- ^ Вудворд, Холли Н.; Тремейн, Кэти; Уильямс, Скотт А.; Занно, Линдси Э.; Хорнер, Джон Р.; Мирвольд, Натан (2020). «Выращивание тираннозавра рекса: остеогистология опровергает карликового «нанотирануса» и поддерживает онтогенетическое разделение ниш у молодых тираннозавров » . Достижения науки . 6 (1): eaax6250. Бибкод : 2020SciA....6.6250W . дои : 10.1126/sciadv.aax6250 . ISSN 2375-2548 . ПМК 6938697 . ПМИД 31911944 .
- ^ Карр, ТД (2020). «Серия роста тираннозавра рекса в высоком разрешении , полученная на основе множества доказательств» . ПерДж . 8 : е9192. дои : 10.7717/peerj.9192 .
- ^ Карр, Т.Д. (5 июня 2020 г.). «Серия роста тираннозавра рекса с высоким разрешением, полученная на основе множества доказательств – автор доктор Томас Д. Карр обсуждает свое новое исследование» . ПирДжблог . Проверено 10 июня 2020 г.
- ^ Лонгрич, Николас Р.; Сайтта, Эван Т. (1 марта 2024 г.). «Таксономический статус Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea) — особого таксона мелких тираннозавров» . Исследования окаменелостей . 2 (1): 1–65. doi : 10.3390/fossils2010001 . eISSN 2813-6284 .
- ^ Jump up to: а б Хорнер, младший; Падиан, К. (2004). «Возраст и динамика роста тираннозавра рекса» . Труды: Биологические науки . 271 (1551): 1875–80. дои : 10.1098/rspb.2004.2829 . ПМК 1691809 . ПМИД 15347508 .
- ^ Jump up to: а б Швейцер, Миннесота; Виттмейер, Дж.Л.; Хорнер, младший (2005). «Полоспецифичная репродуктивная ткань бескилевых и тираннозавра рекса » (PDF) . Наука . 308 (5727): 1456–60. Бибкод : 2005Sci...308.1456S . дои : 10.1126/science.1112158 . ПМИД 15933198 . S2CID 30264554 .
- ^ Ли, АХ; Вернинг, С. (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . Труды Национальной академии наук . 105 (2): 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ПМК 2206579 . ПМИД 18195356 .
- ^ Швейцер, Миннесота; Чжэн, В.; Занно, Л.; Вернинг, С.; Сугияма, Т. (2016). «Химические исследования подтверждают идентификацию репродуктивной ткани тираннозавра рекса с учетом пола» . Научные отчеты . 6 (23099): 23099. Бибкод : 2016NatSR...623099S . дои : 10.1038/srep23099 . ПМЦ 4791554 . ПМИД 26975806 .
- ^ Jump up to: а б Эриксон, генеральный директор; Карри, Пи Джей; Иноуе, Б.Д.; Винн, А.А. (2006). «Таблицы жизни тираннозавров: пример биологии популяции нептичьих динозавров». Наука . 313 (5784): 213–7. Бибкод : 2006Sci...313..213E . дои : 10.1126/science.1125721 . ПМИД 16840697 . S2CID 34191607 .
- ^ Вудворд, Холли Н; Тремейн, Кэти; Уильямс, Скотт А; Занно, Линдси Э; Хорнер, Джон Р.; Мирвольд, Натан (1 января 2020 г.). «Выращивание тираннозавра рекса: остеогистология опровергает карликового «нанотирануса» и поддерживает онтогенетическое разделение ниш у молодых тираннозавров» . Достижения науки . 6 (1): eaax6250. Бибкод : 2020SciA....6.6250W . дои : 10.1126/sciadv.aax6250 . ПМК 6938697 . ПМИД 31911944 .
- ^ Грешко Михаил (1 января 2020 г.). «Эти гладкие хищные динозавры на самом деле являются ти-рексами-подростками» . Нэшнл Географик . Архивировано из оригинала 1 января 2020 года . Проверено 2 января 2020 г.
- ^ Jump up to: а б Хольц, Т. Р. младший (19 марта 2013 г.) [лекция состоялась 8 марта 2013 г.]. Жизнь и времена тираннозавра рекса с доктором Томасом Хольцем (лекция). Сиэтл, Вашингтон: Музей естественной истории и культуры Берка . Архивировано из оригинала 11 декабря 2021 года . Проверено 12 октября 2013 г.
- ^ Пол, GS (2008). «Глава 18: Экстремальный образ жизни и привычки гигантских тираннозаврид-суперхищников мелового периода Северной Америки и Азии» . В Ларсоне, Польша; Карпентер, К. (ред.). Тираннозавр , Король-Тиран . Издательство Университета Индианы. стр. 307–345. ISBN 978-0-253-35087-9 . Проверено 14 сентября 2013 г.
- ^ Jump up to: а б Белл, PR; Кампионе, Невада; Лица IV, WS; Карри, Пи Джей; Ларсон, Польша; Танке, DH; Баккер, RT (2017). «Покровы тираннозавроидов демонстрируют противоречивые закономерности гигантизма и эволюции перьев» . Письма по биологии . 13 (6): 20170092. doi : 10.1098/rsbl.2017.0092 . ПМЦ 5493735 . ПМИД 28592520 .
- ^ Фараго, Дж. (7 марта 2019 г.). «Ти-Рекс, каким вы его не видели: с перьями» . Нью-Йорк Таймс . Архивировано из оригинала 8 марта 2019 года . Проверено 7 марта 2019 г.
- ^ Jump up to: а б Син, X .; Норелл, Массачусетс; Куанг, X.; Ван, X.; Чжао, К.; Цзя, К. (7 октября 2004 г.). «Базальные тираннозавроиды из Китая и свидетельства существования протоперьев у тираннозавроидов» (PDF) . Природа . 431 (7009): 680–684. Бибкод : 2004Natur.431..680X . дои : 10.1038/nature02855 . ПМИД 15470426 . S2CID 4381777 .
- ^ Син, X .; Ван, К.; Чжан; Ма, Кью; Син, Л.; Салливан, К.; Ху, Д.; Ченг, С.; Ван, С. (5 апреля 2012 г.). «Гигантский пернатый динозавр из нижнего мела Китая» (PDF) . Природа . 484 (7392): 92–95. Бибкод : 2012Natur.484...92X . дои : 10.1038/nature10906 . ПМИД 22481363 . S2CID 29689629 . Архивировано из оригинала (PDF) 17 апреля 2012 г.
- ^ Нэйш, Д. «Чувствительная морда большого хищного динозавра» . Зоология четвероногих . Сеть блогов Scientific American . Проверено 5 декабря 2018 г.
- ^ "МОРФОЛОГИЯ, ТАКСОНОМИЯ И ФИЛОГЕНЕТИЧЕСКИЕ СВЯЗИ МОНТЕВИАЛЬНЫХ КРОКОДИЛОВ (ОЛИГОЦЕН, ИТАЛИЯ). стр. 67" . Ежегодный симпозиум по палеонтологии и сравнительной анатомии позвоночных . Проверено 9 октября 2020 г.
- ^ Милинкович, Мишель; Манукян, Лиана; Дебри, Адриан; Ди-Пои, Николас; Мартин, Сэмюэл; Сингх, Далджит; Ламберт, Доминик; Цвикер, Матиас (4 января 2013 г.). «Чешуйки крокодиловой головы не являются единицами развития, а возникают в результате физического растрескивания» . Наука . 339 (6115): 78–81. Бибкод : 2013Sci...339...78M . дои : 10.1126/science.1226265 . ПМИД 23196908 . S2CID 6859452 .
- ^ Карпентер, К. (1992). «Вариация тираннозавра рекса ». В Карпентере, К.; Карри, Пи Джей (ред.). Систематика динозавров: подходы и перспективы . Кембридж : Издательство Кембриджского университета . стр. 141–145. ISBN 978-0-521-43810-0 .
- ^ Ларсон, Польша (1994). « Секс тираннозавра ». В Розенберге, Грузия; Вольберг, Д.Л. (ред.). Дино Фест . Том. 7. Специальные публикации Палеонтологического общества. стр. 139–155.
- ^ Эриксон, генеральный директор; Кристофер, Луизиана; Ларсон, П. (2005). «Андрогинный рекс - использование шевронов для определения пола крокодилов и нептичьих динозавров» (PDF) . Зоология (Йена, Германия) . 108 (4): 277–86. Бибкод : 2005Zool..108..277E . дои : 10.1016/j.zool.2005.08.001 . ПМИД 16351976 .
- ^ Швейцер, Миннесота; Элси, РМ; Даке, К.Г.; Хорнер, младший; Ламм, ET (2007). «Образуют ли яйцекладущие крокодилы ( Alligator Mississippiensis ) архозавры медуллярную кость?». Кость . 40 (4): 1152–8. дои : 10.1016/j.bone.2006.10.029 . ПМИД 17223615 .
- ^ Лейди, Дж. (1865). «Мемуары о вымерших рептилиях меловых формаций США». Смитсоновский вклад в знания . 14 : 1–135.
- ^ « Тираннозавр » . Американский музей естественной истории . Архивировано из оригинала 8 декабря 2008 года . Проверено 16 октября 2008 г.
- ^ Jump up to: а б с Ньюман, Б.Х. (1970). «Стойка и походка плотоядного тираннозавра » . Биологический журнал Линнеевского общества . 2 (2): 119–123. дои : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ Фреска «Эпоха рептилий» . Йельский университет. 2008. Архивировано из оригинала 29 сентября 2018 года . Проверено 16 октября 2008 г.
- ^ Росс, РМ; Дагган-Хаас, Д.; Олмон, WD (2013). «Поза тираннозавра рекса : почему взгляды студентов отстают от науки?». Журнал геонаучного образования . 61 (1): 145–160. Бибкод : 2013JGeEd..61..145R . дои : 10.5408/11-259.1 . S2CID 162343784 .
- ^ « Тираннозавр Рекс : Больше не штатив» . Американская ассоциация содействия развитию науки . 2 апреля 2013 г.
- ^ Стивенс, Кент А. (2011). «Тираннозавр рекс – «Рекс, сядь» » . ix.cs.uoregon.edu . Проверено 26 июля 2020 г.
- ^ «Если Ти-рекс упал, как он поднялся, учитывая его крошечные руки и низкий центр тяжести?» . Научный американец . Проверено 26 июля 2020 г.
- ^ Jump up to: а б с д Падиан К. (2022). «Почему передние конечности тираннозаврид были такими короткими: интегративная гипотеза» . Acta Palaeontologica Polonica 67 (1): с. 63-76
- ^ Стивенс К.А., Ларсон П., Уиллис Э.Д. и Андерсон А. «Рекс, сиди: цифровое моделирование тираннозавра в состоянии покоя». В Ларсон П. и Карпентер К. (ред.). Тираннозавр рекс, король-тиран (Indiana University Press, 2008). п. 192-203
- ^ Ламбе, LM (1914). «О новом роде и виде хищных динозавров из формации Белли-Ривер в Альберте, с описанием черепа Stephanosaurus Marginatus из того же горизонта». Оттавский натуралист . 27 : 129–135.
- ^ Jump up to: а б с Хорнер, младший ; Лессем, Д. (1993). Полный Ти-рекс . Нью-Йорк : Саймон и Шустер . ISBN 978-0-671-74185-3 .
- ^ «Новый взгляд на тираннозавра | Смитсоновский национальный музей естественной истории» . 13 апреля 2020 года. Архивировано из оригинала 13 апреля 2020 года . Проверено 13 апреля 2020 г.
- ^ Карпентер, К .; Смит, М. (2001). «Остеология и биомеханика передних конечностей тираннозавра рекса » . Танке , DH ; Карпентер, К. (ред.). Мезозойская жизнь позвоночных . Блумингтон: Издательство Университета Индианы. стр. 90–116 . ISBN 978-0-253-33907-2 .
- ^ Пикрелл, Дж. (2 ноября 2017 г.). «Крошечные руки тираннозавра могли быть жестоким оружием» . Нэшнл Географик . Архивировано из оригинала 2 ноября 2017 года . Проверено 10 декабря 2018 г.
- ^ Баккер, RT (1968). «Превосходство динозавров» (PDF) . Открытие . 3 (2): 11–12. Архивировано из оригинала (PDF) 9 сентября 2006 г. Проверено 7 октября 2008 г.
- ^ Баккер, RT (1972). «Анатомические и экологические доказательства эндотермии у динозавров» (PDF) . Природа . 238 (5359): 81–85. Бибкод : 1972Natur.238...81B . дои : 10.1038/238081a0 . S2CID 4176132 . Архивировано из оригинала (PDF) 9 сентября 2006 г. Проверено 7 октября 2008 г.
- ^ Баррик, RE; Души, WJ (1994). «Термофизиология тираннозавра рекса : данные по изотопам кислорода». Наука . 265 (5169): 222–224. Бибкод : 1994Sci...265..222B . дои : 10.1126/science.265.5169.222 . ПМИД 17750663 . S2CID 39392327 .
- ^ Труман, К.; Ченери, К.; Эберт, Д.А.; Спиро, Б. (2003). «Диагенетическое влияние на изотопный состав кислорода костей динозавров и других позвоночных животных, извлеченных из наземных и морских отложений» (PDF) . Журнал Геологического общества . 160 (6): 895–901. Бибкод : 2003JGSoc.160..895T . дои : 10.1144/0016-764903-019 . S2CID 130658189 .
- ^ Баррик, RE; Души, WJ (1999). «Термофизиология и биология гиганотозавра : сравнение с тираннозавром » . Электронная палеонтология . 2 (2). Архивировано из оригинала 17 мая 2011 года . Проверено 7 октября 2008 г.
- ^ Баррик, RE; Стоскопф, МК; Души, WJ (1999). «Изотопы кислорода в костях динозавров». В Фарлоу, Джо; Бретт-Сурман, депутат Кнессета (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 474–490. ISBN 978-0-253-21313-6 .
- ^ Виманн, Дж.; Менендес, И.; Кроуфорд, Дж. М.; Фаббри, М.; Готье, Дж.А.; Халл, премьер-министр; Норелл, Массачусетс; Бриггс, DEG (2022). «Ископаемые биомолекулы обнаруживают птичий метаболизм у предков динозавров» . Природа . 606 (7914): 522–526. Бибкод : 2022Natur.606..522W . дои : 10.1038/s41586-022-04770-6 . ПМИД 35614213 . S2CID 249064466 .
- ^ Паладино, ФВ; Спотила, младший; Додсон, П. (1999). «Проект гигантов: моделирование физиологии крупных динозавров». В Фарлоу, Джо; Бретт-Сурман, депутат Кнессета (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 491–504. ISBN 978-0-253-21313-6 .
- ^ Чинсами, А.; Хиллениус, WJ (2004). «Физиология нептичьих динозавров». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия . Беркли: Издательство Калифорнийского университета. стр. 643–659 . ISBN 978-0-520-24209-8 .
- ^ Сеймур, Р.С. (5 июля 2013 г.). «Максимальная аэробная и анаэробная выработка энергии у крупных крокодилов по сравнению с млекопитающими: последствия гигантотермии динозавров» . ПЛОС ОДИН . 8 (7): e69361. Бибкод : 2013PLoSO...869361S . дои : 10.1371/journal.pone.0069361 . ISSN 1932-6203 . ПМК 3702618 . ПМИД 23861968 .
- ^ Холлидей, СМ; Портер, WR; Вилет, Калифорния; Витмер, LM (2019). «Лобно-теменная ямка и дорсотемпоральное отверстие архозавров и их значение для интерпретации сосудистой и мышечной анатомии динозавров» . Анатомическая запись . 303 (4): 1060–1074. дои : 10.1002/ar.24218 . ПМИД 31260177 . S2CID 195756776 .
- ^ Jump up to: а б Филдс, Х. (2006). «Шокер динозавров» . Смитсоновский журнал . Архивировано из оригинала 14 октября 2008 года . Проверено 2 октября 2008 г.
- ^ Швейцер, Миннесота ; Виттмейер, Дж.Л.; Хорнер, младший; Топорски, Дж. К. (2005). «Сосуды мягких тканей и сохранение клеток у тираннозавра рекса ». Наука . 307 (5717): 1952–5. Бибкод : 2005Sci...307.1952S . дои : 10.1126/science.1108397 . ПМИД 15790853 . S2CID 30456613 .
- ^ Ринкон, П. (12 апреля 2007 г.). «Белок связывает тираннозавра с курами» . Новости Би-би-си . Проверено 2 октября 2008 г.
- ^ Вергано, Д. (13 апреля 2007 г.). «Вчерашний Ти-Рекс — сегодняшняя курица» . США сегодня . Проверено 8 октября 2008 г.
- ^ Кэй, Т.Г.; Гоглер, Г.; Савлович, З. (2008). Степанова, А. (ред.). «Мягкие ткани динозавров, интерпретируемые как бактериальные биопленки» . ПЛОС ОДИН . 3 (7): e2808. Бибкод : 2008PLoSO...3.2808K . дои : 10.1371/journal.pone.0002808 . ПМЦ 2483347 . ПМИД 18665236 .
- ^ «Новые исследования бросают вызов представлению о том, что мягкие ткани динозавров все еще выживают» (пресс-релиз). Новости. 24 июля 2008 года . Проверено 8 октября 2008 г.
- ^ «Дебаты исследователей: это консервированная ткань динозавра или бактериальная слизь?» (Пресс-релиз). Обнаружить. 30 июля 2008 года. Архивировано из оригинала 7 августа 2008 года . Проверено 4 сентября 2008 г.
- ^ Сан-Антонио, JD; Швейцер, Миннесота; Дженсен, СТ; Каллури, Р.; Бакли, М.; Оргель, JPRO (2011). Ван Вин, HW (ред.). «Пептиды динозавров предполагают механизмы выживания белков» . ПЛОС ОДИН . 6 (6): e20381. Бибкод : 2011PLoSO...620381S . дои : 10.1371/journal.pone.0020381 . ПМК 3110760 . ПМИД 21687667 .
- ^ Петерсон, Дж. Э.; Ленчевский, МЭ; Шерер, Р.П. (12 октября 2010 г.). «Влияние микробных биопленок на сохранность первичных мягких тканей у ископаемых и современных архозавров» . ПЛОС ОДИН . 5 (10): е13334. Бибкод : 2010PLoSO...513334P . дои : 10.1371/journal.pone.0013334 . ПМЦ 2953520 . ПМИД 20967227 .
[Т] интерпретация сохранившихся органических остатков как микробной биопленки [крайне маловероятна]
- ^ Jump up to: а б с д Хатчинсон-младший; Гарсия, М. (2002). « Тираннозавр не был быстрым бегуном» . Природа . 415 (6875): 1018–21. Бибкод : 2002Natur.415.1018H . дои : 10.1038/4151018a . ПМИД 11875567 . S2CID 4389633 .
- ^ Jump up to: а б с Хатчинсон, младший (2004). «Биомеханическое моделирование и анализ чувствительности способности к двуногому бегу. II. Вымершие таксоны» (PDF) . Журнал морфологии . 262 (1): 441–461. дои : 10.1002/jmor.10240 . ПМИД 15352202 . S2CID 15677774 . Архивировано из оригинала (PDF) 31 октября 2008 г.
- ^ Jump up to: а б Хольц, Т.Р. (1 мая 1996 г.). «Филогенетическая систематика целурозавров (Dinosauria; Theropoda)» . Журнал палеонтологии . 70 (3): 536–538. Бибкод : 1996JPal...70..536H . дои : 10.1017/S0022336000038506 . S2CID 87599102 . Проверено 3 октября 2008 г.
- ^ Бентон, М. (2014). Палеонтология позвоночных (4-е изд.). Джон Уайли и сыновья. п. 193. ИСБН 978-1-118-40755-4 .
- ^ Дечекки, Т. Александр; Млошевская, Александра М.; Хольц, Томас Р. младший; Хабиб, Майкл Б.; Ларссон, Ханс CE (13 мая 2020 г.). «Быстро и экономно: различные стратегии локомоторики способствуют удлинению конечностей у тероподных динозавров» . ПЛОС ОДИН . 15 (5): e0223698. Бибкод : 2020PLoSO..1523698D . дои : 10.1371/journal.pone.0223698 . ПМК 7220109 . ПМИД 32401793 .
- ^ «Ти-рекс был чемпионом по ходьбе, сверхэффективным на низких скоростях» . ЭврекАлерт! . 13 мая 2020 г. Проверено 16 августа 2023 г.
- ^ ван Байлерт, Пенсильвания; ван Зост, AJK; Шульп, А.С. (2021). «Метод естественной частоты: оценка предпочтительной скорости ходьбы тираннозавра рекса на основе естественной частоты хвоста» . Королевское общество открытой науки . 8 (4): 201441. Цифровой код : 2021RSOS....801441V . дои : 10.1098/rsos.201441 . ПМЦ 8059583 . ПМИД 33996115 . S2CID 233312053 .
- ^ «Почему Тираннозавр бегал медленно и почему самые большие не всегда самые быстрые» . ScienceDaily . 17 июля 2017 г. Проверено 10 ноября 2017 г.
- ^ Хирт, MR; Джетц, В.; Ралл, Британская Колумбия; Броуз, У. (2017). «Общий закон масштабирования показывает, почему самые крупные животные не являются самыми быстрыми». Экология и эволюция природы . 1 (8): 1116–1122. Бибкод : 2017NatEE...1.1116H . дои : 10.1038/s41559-017-0241-4 . ПМИД 29046579 . S2CID 425473 .
- ^ Селлерс, Висконсин; Пруд, Южная Каролина; Брасси, Калифорния; Мэннинг, Польша; Бейтс, КТ (18 июля 2017 г.). «Исследование беговых способностей тираннозавра рекса с использованием динамического анализа нескольких тел с ограничениями по напряжению» . ПерДж . 5 : е3420. дои : 10.7717/peerj.3420 . ISSN 2167-8359 . ПМК 5518979 . ПМИД 28740745 .
- ^ Коттон, младший; Хартман, SA; Карри, Пи Джей; Уитмер, Л.М.; Рассел, AP; Хольц, Т.Р. младший; Бернс, Мэн; Сурринг, Луизиана; Мэллисон, Х.; Хендерсон, DM; О'Брайен, Х.; Снивели, Э. (21 февраля 2019 г.). «Меньшая вращательная инерция и большие мышцы ног указывают на более быстрые повороты у тираннозаврид, чем у других крупных теропод» . ПерДж . 7 : е6432. дои : 10.7717/peerj.6432 . ПМК 6387760 . ПМИД 30809441 .
- ^ Канир, Т.; Молкестад, Т.; Лукас, СГ (2021). «СЛЕДЫ В ВЕРХНЕМ МЕЛУ БАССЕЙНА РАТОН, ВОЗМОЖНО, ПОКАЗЫВАЮТ ПОДЪЕМ ТИРАННОЗАВРИДА ИЗ ПОЛОЖЕНИЯ ЛЕЖА» . Музей естественной истории и науки Нью-Мексико : 29–37.
- ^ Смит, С.Д.; Лица, WS; Син, Лида (2016). «Тропа тираннозавра в Гленроке, Ланс-Формейшн (Маастрихт), Вайоминг» . Меловые исследования . 61 : 1–4. Бибкод : 2016CrRes..61....1S . дои : 10.1016/j.cretres.2015.12.020 .
- ^ Де Пастино, Б. (2016). «Следы тираннозавра, обнаруженные в Вайоминге, показывают скорость динозавра» . Западные раскопки . 61 : 1–4. Бибкод : 2016CrRes..61....1S . дои : 10.1016/j.cretres.2015.12.020 . Архивировано из оригинала 18 января 2021 года . Проверено 3 января 2021 г.
- ^ Jump up to: а б Уитмер, Л.М.; Риджли, Р.К. (2009). «Новые сведения о мозге, черепной коробке и области ушей тираннозавров (динозавров, теропод) с последствиями для сенсорной организации и поведения» . Анатомическая запись . 292 (9): 1266–1296. дои : 10.1002/ar.20983 . ПМИД 19711459 . S2CID 17978731 .
- ^ Стивенс, штат Калифорния (1 апреля 2011 г.). «Бинокулярное зрение теропод-динозавров» . Проверено 19 июля 2013 г.
- ^ Музей естественной истории Берпи; Университет Северного Иллинойса, ред. (2013). Палеобиология тираннозаврид . Жизнь прошлого. Блумингтон (Индиана): Издательство Университета Индианы. стр. 267–268. ISBN 978-0-253-00930-2 .
- ^ Jump up to: а б « Исследование мозга тиранозавра выявило утонченный «нос» » . Калгари Геральд . 28 октября 2008 года . Проверено 29 октября 2008 г.
- ^ Кавабе, Соитиро; Хаттори, Соки (2021). «Сложная сосудисто-нервная система в зубной кости тираннозавра» . Историческая биология . 34 (7): 1137–1145. дои : 10.1080/08912963.2021.1965137 .
- ^ «На челюсти тираннозавра были датчики, которые делали его еще более грозным хищником» . физ.орг .
- ^ Бенуа, Флориан Буабделла, Эмили Лесснер и Жюльен (20 января 2022 г.). «Ростральная нервно-сосудистая система тираннозавра рекса» . Электронная палеонтология . 25 (1): 1–20. дои : 10.26879/1178 . ISSN 1094-8074 . S2CID 246204236 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Херлберт, Г.С.; Риджли, Колорадо; Уитмер, LM (5 июля 2013 г.). «Относительный размер головного мозга и головного мозга у динозавров-тираннозаврид: анализ с использованием количественных соотношений мозг-эндокаст у современных аллигаторов» . В Пэррише, MJ; Мольнар, Р.Э.; Карри, Пи Джей; Коппельхус, Э.Б. (ред.). Палеобиология тираннозаврид . Издательство Университета Индианы. стр. 134–154. ISBN 978-0-253-00947-0 . Проверено 20 октября 2013 г.
- ^ Брусаттен, Стив (2018). Взлет и падение динозавров . Нью-Йорк, Нью-Йорк: Издательство HarperCollins. п. 219. ИСБН 978-0-06-249043-8 .
- ^ Jump up to: а б «Банды Дино» . Канал Дискавери . 22 июня 2011. Архивировано из оригинала 19 января 2012 года . Проверено 19 января 2012 г.
- ^ Коллинз, Н. (22 июня 2011 г.). «Тираннозавр Рекс «охотился стаями» » . Телеграф . Архивировано из оригинала 10 января 2022 года . Проверено 23 марта 2014 г.
- ^ Уоллис, П. (11 июня 2012 г.). «Комментарий: Охотники за стаями тираннозавров? Страшно, но, скорее всего, это правда» . Digitaljournal.com . Проверено 23 декабря 2015 г.
- ^ Jump up to: а б с Свитек, Б. (25 июля 2011 г.). «Куча костей не делает банду кровожадных тираннозавров» . Хранитель . Проверено 21 июня 2015 г.
- ^ Карри, Филип Дж . (1998). «Возможные доказательства стадного поведения тираннозаврид» (PDF) . Гайя . 15 : 271–277. Архивировано из оригинала (PDF) 26 марта 2009 г. Проверено 3 мая 2009 г. (не печаталось до 2000 г.)
- ^ Образец, И. (23 июля 2014 г.). «Исследователи обнаружили первые признаки того, что тираннозавры охотились стаями» . Хранитель . Проверено 28 июля 2014 г.
- ^ МакКри, RT (2014). «Ужас тираннозавров»: первые следы тираннозаврид и свидетельства стадности и патологии тираннозаврид» . ПЛОС ОДИН . 9 (7): е103613. Бибкод : 2014PLoSO...9j3613M . дои : 10.1371/journal.pone.0103613 . ПМК 4108409 . ПМИД 25054328 .
- ^ Роуч, Брайан Т.; Бринкман, Дэниел Т. (2007). «Переоценка совместной охоты и стадности Deinonychus antirhopus и других нептичьих динозавров-тероподов». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- ^ Петерсон, Дж. Э.; Хендерсон, доктор медицины; Шерер, Р.П.; Витторе, CP (2009). «Укусы лица молодых тираннозаврид и поведенческие последствия» . ПАЛЕОС . 24 (11): 780–784. Бибкод : 2009Палай..24..780P . дои : 10.2110/palo.2009.p09-056r . S2CID 85602478 . Архивировано из оригинала 11 августа 2013 года.
- ^ Паризи, Т. (2 ноября 2009 г.). «Ужасные подростки тираннозавров учёных НИУ: Молодые тираннозавры устроили серьёзную битву друг против друга» . Университет Северного Иллинойса. Архивировано из оригинала 14 августа 2013 года . Проверено 10 августа 2013 г.
- ^ «Время убить тираннозавра- мусорщика «Дебаты» » . Нэшнл Географик . 16 июля 2013 г. Архивировано из оригинала 12 июля 2018 г.
- ^ Блэк, Райли (13 апреля 2012 г.). «Когда тираннозавр грыз зауроподов» . Смитсоновский журнал . 25 . Smithsonian Media: 469. doi : 10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2 . S2CID 131583311 . Проверено 24 августа 2013 г.
- ^ Блэк, Райли (2012). « тираннозавра рекса Опасный и смертельный укус » . Смитсоновский журнал . Смитсоновский институт.
- ^ Бейтс, КТ; Фолкингем, Польша (29 февраля 2012 г.). «Оценка максимальной эффективности укуса тираннозавра рекса с использованием динамики нескольких тел» . Биологические письма . 8 (4): 660–664. дои : 10.1098/rsbl.2012.0056 . ПМЦ 3391458 . ПМИД 22378742 .
- ^ Скалли, К. (2002). Оксфордский справочник по прикладным стоматологическим наукам . Издательство Оксфордского университета. п. 156 . ISBN 978-0-19-851096-3 .
- ^ Лаутеншлагер, Стефан (4 ноября 2015 г.). «Оценка краниальных скелетно-мышечных ограничений у тероподовых динозавров» . Королевское общество открытой науки . 2 (11): 150495. Бибкод : 2015RSOS....250495L . дои : 10.1098/rsos.150495 . ПМК 4680622 . ПМИД 26716007 .
- ^ «Чем лучше тебя есть? Как челюсти динозавров повлияли на рацион» . Наука Дейли . 3 ноября 2015 года. Архивировано из оригинала 4 ноября 2015 года . Проверено 14 сентября 2018 г.
- ^ Ламбе, Л.Б. (1917). «Меловой тероподный динозавр Горгозавр » . Мемуары Геологической службы Канады . 100 : 1–84. дои : 10.4095/101672 .
- ^ Фарлоу, Джо; Хольц (2002). «Ископаемая летопись хищничества динозавров» (PDF) . В Ковалевском, М.; Келли, PH (ред.). Ископаемая летопись хищничества . Документы Палеонтологического общества. Том. 8. ТР-младший, стр. 251–266. Архивировано из оригинала (PDF) 31 октября 2008 г.
- ^ Jump up to: а б Хорнер, младший (1994). «Ножи для стейка, глаза-бусинки и крохотные ручки (портрет тираннозавра в образе мусорщика)». Специальное издание Палеонтологического общества . 7 : 157–164. дои : 10.1017/S2475262200009497 .
- ^ Амос, Дж. (31 июля 2003 г.). «Наука/Природа: Ти-рекс предстанет перед судом» . Новости Би-би-си . Проверено 23 декабря 2015 г.
- ^ Амос, Дж. (31 июля 2003 г.). «Наука/Природа: Ти-рекс предстанет перед судом» . Новости Би-би-си . Проверено 23 декабря 2015 г.
- ^ Фарлоу, Джо; Хольц, Т.Р. (2002). Ковалевски, М.; Келли, PH (ред.). «Ископаемые записи хищничества динозавров» (PDF) . Документы Палеонтологического общества . 8 : 251–266. дои : 10.1017/S108933260000111X . Архивировано из оригинала (PDF) 31 октября 2008 г.
- ^ Хольц, Томас Р. (2008). «Глава 20: Критическая переоценка гипотезы обязательного падальщика тираннозавра рекса и других динозавров-тиранов» . В Ларсоне, Питер; Карпентер, Кеннет (ред.). Тираннозавр рекс: Король-тиран . Книжные издательства. стр. 371–394. ISBN 978-0-253-35087-9 .
- ^ Чин, К.; Токарык, ТТ; Эриксон, генеральный директор; Калк, LC (18 июня 1998 г.). «Копролит-теропод королевских размеров» . Природа . 393 (6686): 680–682. Бибкод : 1998Natur.393..680C . дои : 10.1038/31461 . S2CID 4343329 . Резюме на Монастерский Р. (20 июня 1998 г.). «Получение совок из фекалий тираннозавра » . Новости науки . 153 (25): 391. дои : 10.2307/4010364 . JSTOR 4010364 . Архивировано из оригинала 11 мая 2013 года.
- ^ Карпентер, К. (1998). «Свидетельства хищнического поведения динозавров-теропод» . Гайя . 15 : 135–144. Архивировано из оригинала 17 ноября 2007 года . Проверено 5 декабря 2007 г.
- ^ Эберт, Дэвид А.; Эванс, Дэвид С., ред. (2015). «Палеопатология позднемеловых Hadrosauridae из Альберты, Канада». Гадрозавры . Издательство Университета Индианы. стр. 540–571. ISBN 978-0-253-01390-3 . препринт
- ^ Сивьеро, АРТ В.; Брэнд, Леонард Р.; Купер, Аллен М.; Хейс, Уильям К.; Рега, Элизабет; Сивьеро, Бетания, Коннектикут (2020). «Скелетная травма, повлиявшая на внутрихвостовую подвижность у Edmontosaurus Annectens из монодоминантного костного ложа, формация Ланс (Маастрихт), Вайоминг, США» . ПАЛЕОС . 35 (4): 201–214. Бибкод : 2020Палай..35..201S . дои : 10.2110/palo.2019.079 . S2CID 218503493 .
- ^ Хэпп, Дж.; Карпентер, К. (2008). «Анализ поведения хищника и жертвы при столкновении тираннозавра рекса и трицератопса ». В Карпентере, К.; Ларсон, ЧП (ред.). Тираннозавр рекс, Король-Тиран (Жизнь прошлого) . Блумингтон: Издательство Университета Индианы. стр. 355–368. ISBN 978-0-253-35087-9 .
- ^ Додсон, П. (1996). Рогатые динозавры . Издательство Принстонского университета. п. 19 .
- ^ Танке, DH ; Карри, Пи Джей (1998). «Кусание головы динозавров-теропод: палеопатологические данные» (PDF) . Гайя (15): 167–184. ISSN 0871-5424 . Архивировано из оригинала (PDF) 27 февраля 2008 г.
- ^ Петерсон, Дж. Э.; Даус, КНЦ (4 марта 2019 г.). «Следы питания, относящиеся к молодому тираннозавру рексу, дают представление об онтогенетических тенденциях в питании» . ПерДж . 7 : е6573. дои : 10.7717/peerj.6573 . ISSN 2167-8359 . ПМК 6404657 . ПМИД 30863686 .
- ^ Аблер, В.Л. (1992). «Зазубренные зубы динозавров-тираннозаврид и кусающие структуры других животных». Палеобиология . 18 (2): 161–183. Бибкод : 1992Pbio...18..161A . дои : 10.1017/S0094837300013956 . S2CID 88238481 .
- ^ Гольдштейн, ЕЕК; Тиррелл, КЛ; Цитрон, ДМ; Кокс, ЧР; Реккьо, ИМ; Окимото, Б.; Брия, Дж.; Фрай, Б.Г. (1 июня 2013 г.). «Анаэробная и аэробная бактериология слюны и десен 16 содержащихся в неволе драконов Комодо ( Varanus komodoensis ): новые последствия для модели «бактерии как яд»». Журнал медицины зоопарков и дикой природы . 44 (2): 262–272. дои : 10.1638/2012-0022R.1 . ISSN 1042-7260 . ПМИД 23805543 . S2CID 9932073 .
- ^ Снайвли, Э.; Коттон, младший; Риджли, Р.; Витмер, LM (2013). «Многотельная динамическая модель функции головы и шеи у аллозавра (Dinosauria, Theropoda)» . Электронная палеонтология . 16 (2). дои : 10.26879/338 .
- ^ Jump up to: а б с д и ж Лонгрич, Северная Каролина; Хорнер, младший; Эриксон, генеральный директор; Карри, Пи Джей (2010). «Каннибализм у тираннозавра рекса » . ПЛОС ОДИН . 5 (10): е13419. Бибкод : 2010PLoSO...513419L . дои : 10.1371/journal.pone.0013419 . ПМЦ 2955550 . ПМИД 20976177 .
- ^ «НОВЫЕ СВИДЕТЕЛЬСТВА КАННИБАЛИЗМА У ТИРАННОЗАВРИДОВ ДИНОЗАВРОВ ИЗ ВЕРХНЕГО МЕЛА (КАМПАН/МААСТРИХТ) БАССЕЙНА САН-ХУАН В НЬЮ-МЕКСИКЕ» . Исследовательские ворота .
- ^ Фарлоу, Джеймс Орвилл (1989). Палеобиология динозавров . Геологическое общество Америки. ISBN 978-0-8137-2238-2 .
- ^ «Майазавр», Додсон и др. (1994); страницы 116-117
- ^ Хорнер, младший; Макела, Р. (1979). «Гнездо молоди является свидетельством семейной структуры динозавров». Природа . 282 (5736): 296–298. Бибкод : 1979Natur.282..296H . дои : 10.1038/282296a0 . S2CID 4370793 .
- ^ « Лучшая из всех матерей «Maiasaura peeblesorum» . bioweb.uwlax.edu/ . Университет Висконсина-Ла-Кросс . Проверено 22 марта 2021 г.
- ^ Норелл, Марк А.; Кларк, Джеймс М.; Кьяппе, Луис М.; Дашевег, Демберелин (1995). «Гнездящийся динозавр» . Природа . 378 (6559): 774–776. Бибкод : 1995Natur.378..774N . дои : 10.1038/378774a0 . ISSN 1476-4687 . S2CID 4245228 .
- ^ Ватанабэ, Мирна Э. (1 марта 2009 г.). «Развивающиеся идеи о происхождении родительской заботы». Бионаука . 59 (3): 272. doi : 10.1525/bio.2009.59.3.17 . ISSN 0006-3568 . S2CID 85066992 .
- ^ Ротшильд, Б.; Танке, DH; Форд, ТЛ (2001). «Стрессовые переломы теропод и разрывы сухожилий как ключ к активности». В Танке, DH; Карпентер, К. (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. стр. 331–336.
- ^ Вольф, ЭДС; Солсбери, Юго-Запад; Хорнер, младший; Варрикки, диджей (2009). Хансен, DM (ред.). «Обычная птичья инфекция поразила динозавров-тиранов» . ПЛОС ОДИН . 4 (9): е7288. Бибкод : 2009PLoSO...4.7288W . дои : 10.1371/journal.pone.0007288 . ПМЦ 2748709 . ПМИД 19789646 .
- ^ Рега, Э. (2012). «Болезни динозавров». В Бретт-Сурман, М.; Хольц, Т.; Фарлоу, Дж. (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы.
- ^ Ротшильд, Брюс; О'Коннор, Цзинмай; Лосадо, Мария Сесилия (1 декабря 2022 г.). «Более тщательное изучение не подтверждает, что инфекция является причиной загадочных патологий нижней челюсти у тираннозавра рекса» . Меловые исследования . 140 : 105353. Бибкод : 2022CrRes.14005353R . дои : 10.1016/j.cretres.2022.105353 . ISSN 0195-6671 . S2CID 252055157 .
- ^ Перкинс, С. (29 октября 2015 г.). «Тираннозавры, вероятно, были каннибалами (Комментарий)» . Наука . Проверено 2 ноября 2015 г.
- ^ Эстес, Р.; Бербериан, П. (1970). «Палеоэкология позднемелового сообщества позвоночных из Монтаны». Бревиора . 343 : 1–35.
- ^ Дерстлер, К. (1994). «Динозавры формации Ланс в восточном Вайоминге». В Нельсоне, GE (ред.). Динозавры Вайоминга . Путеводитель геологической ассоциации Вайоминга, 44-я ежегодная полевая конференция. Геологическая ассоциация Вайоминга. стр. 127–146.
- ^ Сэмпсон, Скотт Д.; Лоевон, Марк А. (27 июня 2005 г.). « Тираннозавр рекс из верхнемеловой (маастрихтской) формации Северного Рога в штате Юта: биогеографические и палеоэкологические последствия» . Журнал палеонтологии позвоночных . 25 (2): 469–472. doi : 10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2 . JSTOR 4524461 . S2CID 131583311 .
- ^ Чифелли, Ричард Л.; Нидам, Рэндалл Л.; Итон, Джеффри Г.; Гарднер, Джеймс Д.; Киркланд, Джеймс И. (1999). «Фауны позвоночных формации Норт-Хорн (верхний мел – нижний палеоцен), округа Эмери и Санпит, штат Юта». В Gillette, Дэвид Д. (ред.). Палеонтология позвоночных в Юте . Солт-Лейк-Сити: Геологическая служба Юты. стр. 377–388. ISBN 1-55791-634-9 .
- ^ Уик, Стивен Л.; Леман, Томас М. (1 июля 2013 г.). «Новый цератопсовый динозавр из формации Джавелина (Маастрихт) Западного Техаса и его значение для филогении хасмозаврин» . Naturwissenschaften . 100 (7): 667–682. Бибкод : 2013NW....100..667W . дои : 10.1007/s00114-013-1063-0 . ПМИД 23728202 . S2CID 16048008 . Проверено 27 ноября 2020 г.
- ^ Андрес, Б.; Лэнгстон, В. младший (2021). «Морфология и систематика Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)» . Журнал палеонтологии позвоночных . 41 (sup1): 142. Бибкод : 2021JVPal..41S..46A . дои : 10.1080/02724634.2021.1907587 . ISSN 0272-4634 . S2CID 245125409 .
- ^ Твит, Дж.С.; Сантуччи, В.Л. (2018). «Опись нептичьих динозавров из зон обслуживания национальных парков» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 79 : 703–730.
- ^ Ясински, SE; Салливан, РМ; Лукас, СГ (2011). «Таксономический состав местной фауны Аламо-Уош из верхнемеловой формации Охо Аламо (член Наашойбито) бассейна Сан-Хуан, Нью-Мексико». Бюллетень . 53 : 216–271.
- ^ Серрано-Браньяс, CI; Торрес-Родригеса, Э.; Луна, ПЦР; Гонсалес, И.; Гонсалес-Леон, К. (2014). «Зубы тираннозаврид из формации Ломас-Колорадас, группа Кабуллона (верхний мел) Сонора, Мексика». Меловые исследования . 49 : 163–171. Бибкод : 2014CrRes..49..163S . дои : 10.1016/j.cretres.2014.02.018 .
- ^ Брусатте, СЛ; Карр, ТД (2016). «Филогения и история эволюции динозавров-тираннозавроидов» . Научные отчеты . 6 : 20252. Бибкод : 2016NatSR...620252B . дои : 10.1038/srep20252 . ПМЦ 4735739 . ПМИД 26830019 .
- ^ Jump up to: а б Чанг, Кеннет (15 апреля 2021 г.). «Сколько тираннозавров-рексов когда-либо жило на Земле? Вот новая подсказка» . Нью-Йорк Таймс . Архивировано из оригинала 28 декабря 2021 года.
- ^ Jump up to: а б Маршалл, Чарльз Р.; Латорре, Дэниел В.; Уилсон, Коннор Дж.; Фрэнк, Таннер М.; Магулик, Кэтрин М.; Зиммт, Джошуа Б.; Пауст, Эшли В. (16 апреля 2021 г.). «Абсолютная численность и степень сохранности тираннозавра рекса» . Наука . 372 (6539): 284–287. Бибкод : 2021Sci...372..284M . дои : 10.1126/science.abc8300 . ПМИД 33859033 .
- ^ Мейри, Шай (2022). «Размеры популяции тираннозавра не могут быть точно оценены» . Границы биогеографии . 14 (2). дои : 10.21425/F5FBG53781 . S2CID 245288933 .
- ^ Маршалл, Чарльз Р.; Латорре, Дэниел В.; Уилсон, Коннор Дж.; Фрэнк, Таннер М.; Магулик, Кэтрин М.; Зиммт, Джошуа П.; Пауст, Эшли В. (2022). «С какой точностью можно оценить численность популяции тираннозавра рекса? Ответ Мейри» . Границы биогеографии . 14 (2). дои : 10.21425/F5FBG55042 . hdl : 10852/101238 . S2CID 245314491 .
 Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Дальнейшее чтение
- Фарлоу, Джо; Гейтси, С.М.; Хольц, Т.Р. младший; Хатчинсон-младший; Робинсон, Дж. М. (2000). «Передвижение тероподов» . Американский зоолог . 40 (4): 640–663. дои : 10.1093/icb/40.4.640 . JSTOR 3884284 .
Внешние ссылки
- Лекция доктора Стивена Брусатта в Эдинбургском университете «Открытия тираннозавров», 20 февраля 2015 г.
- 28 видов генеалогического древа тираннозавров, когда и где они жили Стивен Брусатт Томас Карр 2016
- Ответ Австралии тираннозаврам , Государственная библиотека Квинсленда
Экспонаты















| Базы данных органов управления : Национальные |
|---|
- Тиранозавр
- неологизмы 1900-х годов
- Высшие хищники
- Ископаемые таксоны описаны в 1905 году.
- Фауна Хелл-Крик
- Ланс фауна
- Формация Ларами
- Позднемеловые динозавры Северной Америки.
- Маастрихтское вымирание родов
- Первые появления маастрихтского рода
- Маастрихтская жизнь
- Палеонтология в Колорадо
- Палеонтология в Монтане
- Палеонтология в Южной Дакоте
- Палеонтология в Вайоминге
- Фауна Сколларда
- Таксоны, названные Генри Фэйрфилдом Осборном
- Палеонтология в Альберте