Торозавр
| Торозавр Временной диапазон: поздний мел ( маастрихт ), | |
|---|---|
 | |
| Образец по прозвищу «Крошечный», Денверский музей природы и науки. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Цератопсия |
| Семья: | † Цератопсиды |
| Подсемейство: | † Хасмозаврины |
| Племя: | † Трицератопсины |
| Род: | † Торозавр Марш , 1891 г. |
| Типовой вид | |
| † Торозавр латус Марш, 1891 г. | |
| Другие виды | |
| |
| Синонимы | |
| |
Торозавр (что означает «перфорированная ящерица», в связи с большими отверстиями в его оборке) — это род травоядных цератопсовых динозавров -хазмозавров , живших в позднем маастрихтском периоде позднего мелового периода, между 68 и 66 миллионами лет назад. возможно, что ареал вида мог простираться еще на 69 миллионов лет назад. [1] Окаменелости были обнаружены по всей западной части Северной Америки, от севера до Саскачевана до юга до Техаса .
Торозавр обладал самыми большими черепами среди всех известных наземных животных: череп с оборками достигал 2,77 метра (9,1 фута) в длину. Считается, что торозавр был такого же размера, как современный трицератопс . [2] [3] [4] но отличается удлиненной оборкой с крупными отверстиями овальной формы, длинными чешуйчатыми косточками оборки с впадиной на их верхней поверхности и наличием пяти и более пар рожков на спинке оборки. [5] У Торозавра также не было длинного рога на носу, который был у Triceratops prorsus. Вместо этого он напоминал более раннего и более базального Triceratops horridus благодаря короткому носовому рогу. [5] На данный момент названы три вида: Torosaurus latus , T. Gladius и T. utahensis . Однако T.gladius больше не считается действительным видом.
В 2010 году достоверность существования Торозавра была оспорена. [6] Исследование гистологии ископаемых костей в сочетании с исследованием формы оборок пришло к выводу, что торозавр, вероятно, представлял собой зрелую форму трицератопса , при этом кости типичных экземпляров трицератопса все еще незрелые и демонстрируют признаки первого развития отчетливых отверстий для оборки торозавра . В процессе взросления оборка черепа сильно удлинилась бы и в ней появились бы отверстия. [7] [8] [9] Однако в 2011, 2012 и 2013 годах исследования внешних особенностей известных экземпляров показали, что морфологические различия между двумя родами исключают их синонимию . Основными проблемами являются отсутствие хороших переходных форм, очевидное существование настоящих полувзрослых особей торозавров , разные пропорции черепа, независимые от созревания, а также образование отверстий на взрослой стадии, не являющееся частью нормальной последовательности созревания цератопсов. [5] [10] [11] Следовательно, до сих пор активно обсуждается, действительно ли Торозавр является взрослым трицератопсом или отдельным родом, хотя большинство исследователей в настоящее время отдают предпочтение последней интерпретации.
Открытие и виды
[ редактировать ]В 1891 году, через два года после присвоения имени трицератопсу , пара черепов цератопсов с удлиненными оборками, несущими отверстия, была найдена Джоном Беллом Хэтчером в округе Ниобрара на юго-востоке Вайоминга . Работодатель Хэтчера, профессор-палеонтолог Отниэль Чарльз Марш , придумал для них род Torosaurus . [12]
Имя Торозавр часто переводится как «ящерица-бык» от латинского существительного «Таурус» или испанского слова «торо» , но, скорее всего, оно происходит от греческого глагола τορέω ( торео) , что означает «продырявить». [13] Намек делается на фенестры , «окнообразные» отверстия в удлиненном оборке, которые традиционно служили для отличия его от сплошного оборки трицератопса . Большая часть путаницы в этимологии названия возникает из-за того, что Марш никогда подробно не объяснял его в своих статьях.
два вида торозавров Были идентифицированы :
- T. latus Marsh , 1891 ( типовой вид ). Latus означает «широкий» на латыни в отношении оборки.
- T. utahensis ( Гилмор , 1946 ) Лоусон, 1976
Другой вид впоследствии был признан идентичным T. latus :
- Т. гладиус Марш , 1891 г. Гладиус на латыни означает «меч», что связано с удлиненной формой чешуйчатой кости .
T. latus был основан на голотипе YPM 1830 , представляющем собой частичный череп. Голотипом T.gladius был экземпляр YPM 1831, череп которого был еще больше. Обе окаменелости были найдены в формации Ланс позднего маастрихта . Подобные экземпляры, найденные в Вайоминге , Монтане , Южной Дакоте , Северной Дакоте , Колорадо , Юте и Саскачеване , с тех пор были отнесены к торозавру . К числу тех, кого можно идентифицировать с некоторой уверенностью, относятся ANSP 15192 (меньшая особь в Южной Дакоте , обнаруженная Эдвином Харрисом Колбертом в 1944 году), [14] MPM VP6841 (частичный скелет с черепом, установленный в Общественном музее Милуоки ), SMM P97.6.1 (череп без морды) и два частичных черепа из формации Хелл-Крик , о которых сообщалось в 2002 году: MOR 981 (обнаружен в 1998 году) и МОР 1122 (с 2001 г.). [15] Фрагментарные останки, которые можно было бы идентифицировать как торозавра, были найдены в районе Биг-Бенд в Техасе и в бассейне Сан-Хуан в Нью-Мексико . [16] Палеонтологи заметили, что экземпляры торозавров встречаются редко в летописи окаменелостей, а экземпляры трицератопса гораздо более многочисленны.
Torosaurus utahensis был первоначально описан как Arrinoceratops utahensis Чарльзом Уитни Гилмором в 1946 году на основе экземпляра USNM 15583 (фрагмент оборки из округа Эмери , штат Юта ). [17] переименовал его в Torosaurus utahensis В 1976 году Дуглас Лоусон . [18] Обзор Роберта Салливана и др. в 2005 году [19] оставил его как Torosaurus utahensis и несколько старше Т. latus . В 2008 году Ребекка Хант сообщила об этом виде значительный дополнительный материал. [20] Еще не опубликовано исследование о том, T. utahensis следует ли рассматривать как новый род или, как было предложено в отношении T. latus , как зрелую стадию роста одного из видов трицератопсов . [7]
Описание
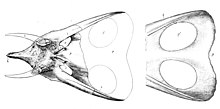
[ редактировать ]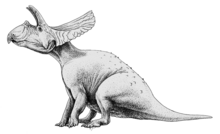
Все особи, отнесенные к торозавру , крупные, сравнимые с крупнейшими экземплярами трицератопсов , их длина составляет от 7,5 до 9 м (от 25 до 30 футов), а вес - от 4 до 6 т (от 8 800 до 13 200 фунтов). За счет удлиненной оборки длина черепа особенно значительна. Хэтчер оценил череп YPM 1830 в 2,2 м (7,2 фута), а череп YPM 1831 - в 2,35 м (7,7 фута). [21] В 1933 году Ричард Суонн Лулл увеличил это значение до 2,4 м (7,9 фута) и 2,57 м (8,4 фута) соответственно. [22] На основании этого Торозавра считалось, что у самый длинный череп среди всех известных наземных животных. Однако в 1998 году Томас Леман заявил, что экземпляр пентацератопса обладал частичным черепом, длина которого при жизни составляла бы 2,9 м (9,5 футов). [23] В этом снова усомнился Николас Лонгрич , который в 2011 году выделил этот экземпляр в отдельный род Titanoceratops и пришел к выводу, что его череп был реконструирован как слишком длинный. [24] Более того, в 2006 году Эндрю Фарк отметил, что описанные им новые черепа в среднем даже длиннее, чем два оригинальных черепа Хэтчера. MOR 1122 имеет длину 2,52 м (8,3 фута), а MOR 981 — 2,77 м (9,1 фута). [25] Хорошо сохранившийся череп экземпляра торозавра по прозвищу «Адам», впервые выставленный на обозрение в 2023 году, является самым крупным из известных черепов, его длина составляет 3 м (9,8 фута). [26] [27]
В 2006 году Фарке установил некоторые диагностические признаки торозавра . Оборка очень длинная по сравнению с остальной частью черепа. Задний край оборки несет десять и более эпипериеталей, или треугольных остеодерм . Срединная треугольная остеодерма отсутствует. Аналогично, остеодерма не пересекает теменно-чешуйчатую границу. Теменная кость тонкая, пронизана теменными окнами в виде круглых или поперечно-овальных отверстий. Теменная кость примерно на 20% шире своей длины. Фарке выявил единственный признак, по которому T. latus отличался как от Triceratops horridus, так и от T. utahensis . Чешуйчатая кость имела по краю заметный гребень, теменная часть которого сочеталась с параллельной ей глубокой продольной впадиной. [25]

Фарке отметил, что известные экземпляры торозавров довольно разнообразны. Орбитальные «бровные» рога иногда большие и изогнуты вперед, как у MOR 981, а иногда короткие и прямые, как показано MOR 1122 и ANSP 15191. Кроме того, положение этих рогов различается, поскольку они часто расположены прямо над глазницей. Однако у YPM 1831 они возникают на заднем краю орбиты. Также существует вариация формы носового рога. YPM 1831 и, в меньшей степени, YPM 1830 имеют прямой, вертикальный носовой рог, но MOR 981, ANSP 15192 и особенно MOR 1122 обладают максимум низким выступом. Оборка также отличается: у ANSP 15192 и YPM 1830 имеется щиток, загнутый вверх сзади, а у YPM 1831 оборка почти плоская, хотя это может быть артефактом реставрации. Оборка YPM 1831 также имеет сердцевидную форму с четким вырезом по средней линии, тогда как у остальных экземпляров задний край прямой. Пропорции оборок весьма разнообразны. У YPM 1831 соотношение длины и ширины составляет 1,26, а у MOR 981 щиток в 2,28 раза длиннее, чем его ширина. Количество треугольных остеодерм оценить сложно, поскольку большинство окаменелостей, по-видимому, утратили их. MOR 981 и MOR 1122 имеют десять и двенадцать эпипариеталей соответственно. YPM 1831 был восстановлен с родничком в крыше черепа, который, возможно, является подлинным. Фарке также пришел к выводу, что степень изменчивости не превышает таковую у родственных родов. [25]
Фарке подчеркнул, что, кроме оборки, никаких систематических различий между торозавром и трицератопсом обнаружить не удалось . Все экземпляры торозавров схожи тем, что у них нет по-настоящему длинного носового рога и горизонтальной артериальной бороздки в переднем основании этого рога, но окаменелости трицератопсов с такой же комбинацией признаков не являются редкостью. [25] В 2008 году Хант пришел к выводу, что T. utahensis , в отличие от T. latus , но схожего с трицератопсом , обладает срединной эпипариетальной линией. [20]
Классификация
[ редактировать ]

Ниже представлена кладограмма цератопсид, основанная на филогенетическом анализе, проведенном Sampson et al. в 2010 году: [28]
В 1891 году Марш поместил Торозавра в Ceratopsidae семейство Ceratopsia (греч. «рогатые лица»). [12] группа травоядных динозавров с клювами, похожими на попугаи , которые процветали в Северной Америке и Азии в юрский и меловой периоды.
Торозавр с его длинным воротником традиционно относился к Chasmosaurinae . Его считали поздним представителем линии, происходящей от Анчицератопса или Арриноцератопса . Таким образом, его поместили в другую ветвь трицератопса , который вплоть до 1980-х годов считался представителем Centrosaurinae из-за его короткого воротника. Однако в 1990-е годы точный кладистический анализ показал, что оба рода были хасмозаврами. Недавние анализы неизменно показывают тесную связь между торозавром и трицератопсом . [25] [28]
Возможная синонимия с трицератопсом.
[ редактировать ]
Споры разгорелись по поводу возможности того, что Торозавр может быть идентичен Трицератопсу . В маастрихтскую эпоху ларамидии два близкородственных хасмозаврина жили в одной и той же среде обитания . Единственным заметным различием между ними была форма оборки. неизвестны Молодые особи торозавров значительное количество молодых особей трицератопсов , но обнаружено . Трицератопс отличается от других хасмозавринов сохранением во взрослом возрасте ювенильных признаков. У взрослых сохраняются короткие чешуйчатые кости, что является случаем педоморфоза . В 2009 году Джон Сканнелла, исследуя онтогенез динозавров в формации Хелл-Крик в Монтане , пришел к выводу, что эту ситуацию лучше всего можно объяснить гипотезой о том, что трицератопс и торозавр были стадиями роста одного рода. Экземпляры торозавров были бы полностью зрелыми особями трицератопса . Торозавр будет младшим синонимом трицератопса , причем последнее имя имеет приоритет. [29]


В 2010 году Сканелла и Джек Хорнер , наставник Сканнеллы в Университете штата Монтана , опубликовали исследование закономерностей роста тридцати восьми образцов черепов (двадцать девять трицератопсов , девять торозавров ) из формации Хелл-Крик. Они пришли к выводу, что Торозавр действительно представляет собой зрелую форму трицератопса . [7] Хорнер подчеркивал, что оборка черепов цератопсов состояла из метапластической кости . Характерной чертой метапластической кости является то, что она может со временем удлиняться и укорачиваться, растягиваясь и резорбируясь, образуя новые формы. Значительное развитие наблюдается даже в тех черепах, которые уже идентифицированы как трицератопсы , заметил Хорнер, «где ориентация рогов назад у молодых и вперед у взрослых». Примерно 50% всех черепов подростков трицератопса имеют две тонкие области в оборках, которые соответствуют расположению «отверстий» в оборках черепа торозавров , окруженных зрелой зернистой костью, что позволяет предположить, что они развились, чтобы компенсировать вес, который в противном случае пришлось бы иметь. были добавлены по мере того, как у взрослеющих особей трицератопса появлялись более длинные оборки. Хорнер сделал это частью более широкого аргумента о том, что в целом многие предполагаемые виды динозавров могли быть стадиями роста других известных видов. [30] [31] У старых особей трицератопса оборка начала значительно удлиняться, в результате чего она уплощалась и расширялась на заднем крае. В то же время должны были появиться теменные отверстия , что привело к типичной форме оборки хасмозаврина. [7]
Сканелла и Хорнер признали, что не все данные можно легко объяснить с помощью их гипотезы. Для этого они выдвинули вспомогательные гипотезы . Одна из проблем заключалась в том, что если бы торозавр был нормальной последней фазой созревания трицератопса , которую они назвали «фазой тороморфа», можно было бы ожидать, что окаменелости торозавров были довольно распространены, тогда как на самом деле они довольно редки. Они объяснили это высокой смертностью молодых особей и возможностью того, что старые животные преимущественно жили на высотах, где эрозия предотвращала окаменение . Вторая проблема заключалась в диапазоне размеров экземпляров торозавров , что, по-видимому, предполагает существование подлинных полувзрослых особей торозавров . Из них они утверждали, что структура костей указывает на полностью зрелый возраст, а разница в размерах является очевидным результатом индивидуальных различий. Третьим возможным возражением было кажущееся отсутствие переходных форм между особями с теменными отверстиями и без них. Эти окна всегда имеют идеальную форму, а не зарождающиеся перфорации. Чтобы противостоять этому, они указали на экземпляр USNM 2412, голотип спорного Недоцератопс , как пример именно такой переходной формы. Проблемные черты этого рода просто отражают его нахождение на первых стадиях превращения в «тороморфа». Последнюю проблему представляло количество остеодерм по краю оборки. У трицератопса обычно имеется пять эпипариетальных костей, включая срединную остеодерму. У Torosaurus их десять или двенадцать, срединная эпипариетальная линия отсутствует. Различно также количество эписквамозальных пластинок на боковом крае оборки (пять у трицератопса , шесть или семь у торозавра ). Это объяснили предположением, что число эпокципиталей увеличивается по мере созревания. Также было отмечено, что как количество, так и положение остеодерм у трицератопса вариабельны . Это показано на экземпляре MOR 2923, у которого есть шесть эпипариетальных костей, но отсутствует срединная. [7]
Выводы Сканнеллы и Хорнера не были приняты единогласно. Некоторые эксперты, хотя и допускали вероятность того, что гипотеза «тороморфа» верна, отрицали ее вероятность. Гипотеза была напрямую оспорена в статье Эндрю Фарка 2011 года и статье Николаса Лонгрича 2012 года. В 2011 году Фарке переописал проблемного Nedoceratops выводка как старую или больную особь своего собственного рода. Сканнелла и Хорнер приводили доводы в пользу его идентификации с трицератопсом . Фарке отметил, что отверстия неправильной формы в оборке недоцератопса не только не проникали в истонченную кость, но и были окружены толстыми вздутиями. Фарке далее пришел к выводу, что некоторые факты трудно совместить с предполагаемым развитием трицератопса в торозавра . В целом число эпокципиталей у цератопсов по мере роста оборки не увеличивается. Несмотря на то, что количество эписквамозальных особей часто варьируется, похоже, нет никакой связи с размером, поскольку у некоторых молодых особей уже наблюдается максимальное количество. По-видимому, это вопрос индивидуальной изменчивости, а не онтогенеза. Так же и у Ceratopia вообще образование отверстий в оборке не связано с возрастом, так как даже у самых молодых особей часто имеются теменные фенестры . Фарке объяснил, что тонкие костные участки на оборке трицератопса , предполагаемое расположение зарождающихся отверстий, являются местами прикрепления мышц. Не было бы последовательной связи между отверстиями и зернистой структурой кости. Многие экземпляры трицератопсов имеют оборки с глубокими прожилками на поверхности, что указывает на значительный возраст. Кость их оборок должна была быть омоложена, а затем снова гранулироваться, чтобы началось образование отверстий, что Фарке считал маловероятным. Наконец, Фарке отметил, что экземпляр YPM 1831, несмотря на свой огромный размер, по-видимому, еще не полностью вырос, о чем свидетельствуют его несросшиеся швы и гладкая текстура кости. Таким образом, он, казалось, представлял собой настоящего несовершеннолетнего Торозавра . [10]

В том же году Сканелла и Хорнер ответили на некоторые критические замечания Фарке. Они признали, что USNM 2412 ввиду своих патологий не является идеальным кандидатом на переходную форму, но подчеркнули, что, помимо вздутий, отверстия в его оборке также окаймлены зернистой и истонченной костью. Приняв во внимание все доказательства, они сочли гораздо более вероятным, что Недоцератопс представляет собой больную особь трицератопса , а не отдельный род. Они также указали на экземпляры трицератопсов, демонстрирующие точную комбинацию прожилковой, зернистой и молодой поперечно-полосатой кости, которую Фарке считал невероятной. Идея о том, что тонкие участки на оборках трицератопса являются местами прикрепления мышц, была ими отвергнута, поскольку кость в этих местах не имела морщинистой поверхности, типичной для такого прикрепления. Разнице в количестве эпокципиталей они предложили два дополнительных объяснения. Кончики остеодермы у старых людей могли разрушиться в течение жизни таким образом, что каждая остеодерма создавала впечатление присутствия двух остеодерм. Таким образом, обычное количество пять или шесть Эпипариетальных трицератопсов могло удвоиться до десяти или двенадцати — именно столько, сколько наблюдалось у экземпляров торозавров . С другой стороны, экземпляры Torosaurus latus , обнаруженные в более старых слоях, могут в процессе анагенеза представлять собой раннюю стадию эволюции трицератопса . Самый старый экземпляр, который можно датировать, MOR 1122, имеет двенадцать эпипариеталей, а более молодой MOR 981 - десять, что, по-видимому, указывает на эволюционную последовательность, в которой количество эпипариеталей постепенно уменьшалось. [32]

В 2012 году Лонгрич исследовал проблему, применив принцип фальсификации . Из любой достоверной научной гипотезы прогнозы можно сделать , с помощью которых можно ее проверить . Лонгрич утверждал, что гипотеза «тороморфа» предполагает три таких предсказания. Во-первых, если бы торозавр был идентичен трицератопсу , их окаменелости должны были быть найдены в тех же местах. На самом деле их географические ареалы не совсем совпадают. На самом севере окаменелости Torosaurus не обнаружены, а на юге только Torosaurus utahensis известен . Однако такая ситуация может быть результатом относительной нехватки останков торозавров и несовершенства выборки. Поэтому Лонгрич пришел к выводу, что гипотеза подтверждается первым предсказанием. Во-вторых, гипотеза предсказывала, что все экземпляры торозавров будут взрослыми, в то время как ни один экземпляр трицератопса не будет очень старым. По мнению Лонгрича, этот последний момент еще не был установлен. Следует признать, что в 2011 году Хорнер опубликовал гистологическое исследование, показывающее, что все Исследованные экземпляры трицератопсов имели полувзрослую костную структуру. [33] но выборка была слишком маленькой, чтобы можно было сделать достоверное обобщение на все окаменелости трицератопса . Чтобы лучше проверить предсказание, Лонгрич предложил список из двадцати четырех внешних черт черепа, с помощью которых можно было проверить образцы на предмет уровня слияния элементов черепа и их созревания. С применением этих критериев было исследовано тридцать шесть образцов. Выяснилось, что слияние обычно происходило в определенной последовательности, что давало дополнительную информацию об их возрасте. Действительно, по этим критериям большинство экземпляров торозавров были очень старыми. Однако было два исключения. Небольшая особь ANSP 15192 была относительно молодой взрослой особью, о чем свидетельствует отсутствие сращения костей морды. Самым молодым экземпляром был YPM 1831 с несросшимися мордой, надскуловой костью и затылочным мыщелком . Более того, он потерял все свои остеодермы жабо, потому что они, по-видимому, еще не срослись, а край жабо имел внешний вид растущей молодой кости. С другой стороны, Лонгрич обнаружил, что десять из Исследованные черепа трицератопсов достигли того же уровня зрелости, что и самые старые экземпляры торозавров . Лонгрич пришел к выводу, что проверка второго предсказания опровергла гипотезу. Третье предсказание заключалось в том, что между торозавром и трицератопсом можно найти переходные формы . Лонгрич считал утверждение о том, что тонкие участки оборок трицератопса были предшественниками теменных окон , самым убедительным доказательством переходной фазы. Однако он отметил, что эти структуры различаются по положению. Впадины трицератопса частично расположены на чешуйчатой кости, а отверстия торозавра полностью окружены теменной. Кроме того, впадины окаймлены гораздо более толстой костью, а отверстия торозавра окружены тонкой костью. Лонгрич пришел к выводу, что гипотеза не удалась в отношении третьего предсказания. Будучи опровергнутой в двух из трёх предсказаний, гипотеза должна быть отвергнута. [5]
Лонгрич также выдвинул некоторые дополнительные возражения против гипотезы «тороморфов». Переходных форм по количеству эпипариеталей не известно. Кроме того, трудно представить, как их число могло увеличиться, поскольку они занимали весь край оборки и в процессе метапластического роста просто увеличили бы свой размер и остальную часть оборки. Предполагаемое расщепление остеодермы в результате эрозии было установлено только для эписквамозных костей, но не для эпипариетальных костей. У Торозавра чешуйчатая кость утолщена на внутренней стороне и вогнута на внешней поверхности, тогда как у трицератопса чешуйчатая кость вогнута на внутренней стороне и плоская сверху. Переходные формы неизвестны. удлинен . Чешуйчатый торозавр также, независимо от абсолютных размеров, гораздо более Лонгрич отметил, что когда образцы торозавров и трицератопсов объединялись для создания единой последовательности роста, как это сделали Сканелла и Хорнер, образцы торозавров ANSP 15192 и YPM 1831 были выдающимися по сравнению с другими образцами. линия регрессии , потому что их чешуйчатые кости были более удлиненными, чем можно было объяснить с помощью аллометрии . Лонгрич признал, что тот факт, что Хорнер в своем гистологическом исследовании смог обнаружить только полувзрослых особей трицератопса, был наводящим на размышления, но предложил альтернативное объяснение, согласно которому трицератопс отличался от своих родственников сохранением относительно молодой костной структуры до старости. С другой стороны, ремоделирование кости не является надежным показателем зрелости ввиду экспериментальных исследований, демонстрирующих, что различия в условиях механического напряжения различных костей могут существенно изменить скорость или степень такого ремоделирования и могут создать иллюзию старой костной ткани. Лонгрич предвидел, что Сканелла и Хорнер отреагируют на его вторую проверку своей гипотезы, заявив, что ее результаты были вызваны индивидуальными вариациями. Однако, по мнению Лонгрича, важность этого фактора была ограничена: например, разницу в размерах между ANSP 15192 и YPM 1831 лучше было бы объяснить половым диморфизмом. , первая, возможно, является молодой взрослой женщиной, а вторая - несовершеннолетним мужчиной. [5]

В 2013 году Фарке и Леонардо Майорино опубликовали морфометрическое исследование, статистический анализ морфопространства ( пространства форм), описывающий вариации черепов Torosaurus , Triceratops horridus , Triceratops prorsus и Nedoceratops , коррелирующие с взрослением. Они пришли к выводу, что черепа Torosaurus latus на протяжении всего периода созревания сохраняли форму, отличную от черепов T. horridus и T. prorsus , причем последние два вида демонстрируют перекрывающиеся пропорции. Это справедливо даже в том случае, если не учитывать форму оборки. Недоцератопс , за исключением размера, оказался неправдоподобной переходной формой между Torosaurus и Triceratops horridus . Фарке и Майорино признали, что небольшое количество экземпляров торозавров снижает надежность этих результатов, но пришли к выводу, что торозавр и трицератопс были отдельными таксонами, хотя и допускали возможность анагенеза, то есть нескольких таксонов, образующих единую хроновидовую линию происхождения, учитывая отсутствие хороших стратиграфических данных. [11]
Гипотеза о том, что образцы Torosaurus latus могут представлять собой «тороморфную» фазу созревания трицератопса , подняла вопрос о том, является ли второй вид Torosaurus , Torosaurus utahensis , также «тороморфом». Эта проблема осложняется отсутствием качественного ископаемого материала, поскольку большинство образцов состоят из изолированных костей. T. utahensis в основном относили к Torosaurus из-за удлиненных чешуйчатых костей, указывающих на длинную оборку. Количество эпипариетальных окон, а также размер, расположение и даже наличие теменных окон неизвестны. Исследователи заявили, что отдельные молодые особи торозавров были раскопаны в костном пласте в формации Джавелина в национальном парке Биг-Бенд , основываясь на их идентификации как Torosaurus cf. utahensis по их близости к взрослой особи с характерной теменной чертой Torosaurus . [20] Сканелла и Хорнер пришли к выводу, что только будущие находки смогут решить эту проблему. Они предположили, что этот таксон, который расширяет ареал торозавров к югу от трицератопса , может представлять собой отдельный род хасмозавров или третий вид трицератопсов . Морфометрическое исследование Фарке в 2013 году не дало окончательных результатов по этому вопросу: морфопространство T. utahensis находится между трицератопсом и Torosaurus latus и недостаточно отделено ни от одного из них. [11]
В 2022 году Мэллон и др. утверждали, что два экземпляра, найденные в канадских формациях Френчмен и Сколлард , EM P16.1. (в Историческом музее Истенда в Саскачеване) и UALVP 1646 (в Университете Альберты ) являются несовершеннолетними и могут быть отнесены к торозавру , что указывает на то, что это действительный таксон. В том же исследовании также отмечается, что торозавр действительно жил в позднем маастрихте . [34]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ * Хикс, Дж. Ф., Джонсон, К. Р., Обрадович, Дж. Д., Миггинс, Д. П. и Токс, Л. 2003. Магнитостратиграфия от верхнего мела (маастрихта) до нижнего эоцена в бассейне Денвера, Колорадо. В книге К.Р. Джонсона, Р.Г. Рейнольдса и М.Л. Рейнольдса (редакторы), «Палеонтология и стратиграфия слоев ларамида в бассейне Денвера», Pt. II., Геология Скалистых гор 38: 1-27.
- ^ Пол, GS; Кристиансен, П. (2000). «Положение передних конечностей у неоцератопсических динозавров: значение для походки и передвижения» (PDF) . Палеобиология . 26 (3): 450–465. doi : 10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2 . JSTOR 2666120 . S2CID 85280946 .
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. стр. 265–267 . ISBN 978-0-691-13720-9 .
- ^ Хольц, Томас Р. младший (2012). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов (PDF) .
Зима 2011 г. Приложение
- ^ Перейти обратно: а б с д и Лонгрич, НР и Филд, ди-джей (2012). « Торозавр - это не трицератопс : онтогенез хасмозавровых цератопсид как пример систематики динозавров» . ПЛОС ОДИН . 7 (2): e32623. Бибкод : 2012PLoSO...732623L . дои : 10.1371/journal.pone.0032623 . ПМК 3290593 . ПМИД 22393425 .
- ^ «Морфозавры: Как динозавры-оборотни обманули нас – жизнь – 28 июля 2010 г.» . Новый учёный. дои : 10.1080/02724634.2010.483632 . S2CID 86767957 . Проверено 3 августа 2010 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Перейти обратно: а б с д и Сканнелла Дж. и Хорнер-младший (2010). « Torosaurus Marsh, 1891 — это Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): синонимия через онтогенез». Журнал палеонтологии позвоночных , 30 (4): 1157–1168. дои : 10.1080/02724634.2010.483632
- ^ Суитек, Брайан. «Новое исследование говорит, что торозавр = трицератопс» . Отслеживание динозавров . Смитсоновский институт.com. Архивировано из оригинала 13 ноября 2013 года . Проверено 2 марта 2011 г.
- ^ Хорнер, Джек (ноябрь 2011 г.). Динозавры, меняющие форму . Выступления TEDX . Проверено 20 ноября 2012 г.
- ^ Перейти обратно: а б Фарке, Эндрю А. (20 января 2011 г.). «Анатомия и таксономический статус хасмозаврового цератопсида Nedoceratops инкубатора из верхнемеловой формации Ланс в Вайоминге, США» . ПЛОС ОДИН . 6 (1): e16196. Бибкод : 2011PLoSO...616196F . дои : 10.1371/journal.pone.0016196 . ISSN 1932-6203 . ПМК 3024410 . ПМИД 21283763 .
- ^ Перейти обратно: а б с Майорино Л., Фарке А.А., Коцакис Т., Пирас П., 2013, «Является ли торозавр трицератопсом ? Геометрические морфометрические свидетельства позднемаастрихтских цератопсидных динозавров», PLoS ONE 8 (11): e81608. doi:10.1371/journal.pone.0081608
- ^ Перейти обратно: а б О. К. Марш, 1891, «Уведомление о новых окаменелостях позвоночных», Американский журнал науки, серия 3, 42 : 265–269.
- ^ Додсон, П. Рогатые динозавры . Принстонский университет. Пресс (Принстон), 1996.
- ^ Колберт, Э. Х. и Дж. Д. Бамп, 1947, «Череп торозавра из Южной Дакоты и пересмотр рода: Труды Академии естественных наук Филадельфии , 99 : 93–106».
- ^ Фарке, А., 2002, «Обзор образцов торозавров (Dinosauria: Ceratopsidae) из Техаса и Нью-Мексико», Журнал палеонтологии позвоночных , 22 : 52A.
- ^ Лукас, С.Г., Мак, Г.Х., Эстеп, Г.В., 1998, «Цератопсовый динозавр Торозавр из формации Макрей верхнего мела, округ Сьерра, Нью-Мексико», Путеводитель геологического общества Нью-Мексико, 49-я полевая конференция, округ Лас-Крусес II
- ^ CW Гилмор, 1946, «Рептилийная фауна формации Норт-Хорн в центральной Юте», Профессиональный документ Министерства внутренних дел США по геологической службе 210 -C: 29-53.
- ^ Д. А. Лоусон, 1976, « Тираннозавр и торозавр , маастрихтские динозавры из Транс-Пекос, Техас», Journal of Paleontology 50 (1): 158-164.
- ^ Салливан, RM, AC Boer и SG Лукас. 2005. Переописание цератопсидного динозавра Torosaurus utahensis (Gilmore, 1946) и пересмотр рода. Журнал Палеонтологии 79:564-582.
- ^ Перейти обратно: а б с Хант, Ребекка К. и Томас М. Леман. 2008. Атрибуты цератопсового динозавра Torosaurus и новый материал из формации Джавелина (Маастрихт) в Техасе. Журнал палеонтологии 82 (6): 1127–1138.
- ^ Хэтчер, Дж. Б., Марш О. К. и Лулл, Р. С., 1907, Цератопсия , Монографии Геологической службы США 49 : 1-198.
- ^ Луллий, RS, 1933, Пересмотр Ceratopsia или рогатых динозавров , Мемуары Музея естественной истории Пибоди 3 (3): 1-175
- ^ Леман, Т.М., 1998, «Гигантский череп и скелет рогатого динозавра Pentaceratops sternbergi из Нью-Мексико: Журнал Палеонтологии, 72 (5): 894-906.
- ^ Николас Р. Лонгрич, 2011, « Titanoceratops Ouranous , гигантский рогатый динозавр из позднего кампана Нью-Мексико», Cretaceous Research , 32 : 264-276.
- ^ Перейти обратно: а б с д и Фарке А.А. «Краниальная остеология и филогенетические взаимоотношения хасмозавринового цератопсида Torosaurus latus », стр. 235-257. В К. Карпентере (ред.). Рога и клювы: цератопсы и динозавры-орнитоподы . Университет Индианы. Пресс (Блумингтон), 2006.
- ^ «Этот скелет торозавра — самый крупный рогатый динозавр, когда-либо обнаруженный в мире» . www.museumofevolution.com . Проверено 20 июня 2023 г.
- ^ «Самый большой череп динозавра, когда-либо найденный на выставке в Дании» . Государственный журнал Висконсина. 13 июня 2023 г. . Проверено 1 июля 2023 г.
- ^ Перейти обратно: а б Скотт Д. Сэмпсон; Марк А. Лоуэн; Эндрю А. Фарк; Эрик М. Робертс; Кэтрин А. Форстер; Джошуа А. Смит и Алан А. Титус (2010). «Новорогие динозавры из Юты служат доказательством внутриконтинентального эндимизма динозавров» . ПЛОС ОДИН . 5. 5 (9): е12292. Бибкод : 2010PLoSO...512292S . дои : 10.1371/journal.pone.0012292 . ПМЦ 2929175 . ПМИД 20877459 .
- ^ Scannella J., 2009, «И потом было одно: последствия синонимии краниального онтогенеза трицератопса », Журнал палеонтологии позвоночных 29 : 177A
- ^ «Новые исследования роста динозавров могут уничтожить треть видов» . Sciencedaily.com. 31 октября 2009 года . Проверено 3 августа 2010 г.
- ^ Хорнер, Джон Р.; Гудвин, Марк Б. (2009). «Чрезвычайная краниальная онтогенез верхнемелового динозавра пахицефалозавра» . ПЛОС ОДИН . 4 (10): е7626. Бибкод : 2009PLoSO...4.7626H . дои : 10.1371/journal.pone.0007626 . ПМК 2762616 . ПМИД 19859556 .
- ^ Сканнелла, Дж. Б. и Хорнер, младший, 2011, «Недоцератопс: пример переходной морфологии», PLoS ONE 6 (12): DOI: 10.1371/journal.pone.0028705
- ^ Хорнер, младший, Ламм, ET., 2011, «Онтогенез теменной оборки трицератопса : предварительный гистологический анализ», Comptes Rendus de l'Academie des Sciences Paris série D 10 : 439–452.
- ^ Мэллон, Джордан С; Холмс, Роберт Б; Бэмфорт, Эмили Л; Шуман, Дирк (7 мая 2022 г.). «Записи о торозаврах (Ornithischia: Ceratopsidae) в Канаде и их таксономические значения» . Зоологический журнал Линнеевского общества . 195 (1): 157–171. doi : 10.1093/zoolinnean/zlab120 .
- Додсон, П. (1996). Рогатые динозавры . Издательство Принстонского университета, Принстон, Нью-Джерси, стр. xiv-346.



