Восточный африказавр
| Восточный африказавр Временной диапазон: поздняя юра , ( титон ) | |
|---|---|
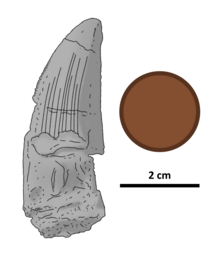 | |
| Иллюстрированный голотип зуба с британским пенни для масштаба. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Спинозавриды (?) |
| Род: | † Восточный Африказавр Бюффето , 2012 г. |
| Разновидность: | † О. crassiserratus |
| Биномиальное имя | |
| † Остафриказавр crassiserratus Бюффето, 2012 г. | |
Остафриказавр — род тероподовых в динозавров позднеюрского периода региона территории современного Линди . Танзании на Он известен только по ископаемым зубам, обнаруженным где-то между 1909 и 1912 годами во время экспедиции к формации Тендагуру Музеем естественной истории Берлина. Восемь зубов первоначально были отнесены к сомнительному роду динозавров Labrosaurus , а позже к Ceratosaurus , оба из которых известны из североамериканской формации Моррисон . Последующие исследования приписали два из этих зубов динозавру -спинозавриду а в 2012 году , Ostafrikasaurus crassiserratus назвал французский палеонтолог Эрик Бюффето , причем один зуб был голотипом , а другой относился к тому же виду. Родовое название происходит от немецкого слова « Немецкая Восточная Африка », прежнего названия колонии, в которой были найдены окаменелости, тогда как видовое название происходит от латинских слов, означающих «толстый» и « » , в отношении формы зазубренный зубы животного.
Остафриказавр предварительно оценивается в 8,4 метра (28 футов) в длину и весит 1,15 тонны (1,27 коротких тонны ; 1,13 длинных тонны ). Зуб голотипа имеет длину 46 миллиметров (1,8 дюйма), имеет изогнутый передний край и овальную форму в поперечном сечении. На зубе имеются зубцы, которые по стандартам спинозавров необычно велики, больше, чем у любого другого известного таксона . И передняя, и задняя режущие кромки имеют зазубрины, от двух до четырех зубчиков на мм (0,04 дюйма). Зуб также имеет продольные гребни с обеих сторон, а внешний слой эмали имеет морщинистую текстуру в областях между гребнями и без них.
Среди старейших известных окаменелостей спинозавридов остафриказавр может иметь важное значение для понимания эволюционного происхождения спинозавридов и их анатомических адаптаций . Сравнивая со своими более поздними родственниками, остафриказавр показывает, что зубы спинозавров стали более коническими и утратили зазубрины на протяжении всей эволюции. Возможно, это результат того, что они стали более специализированными к рыбоядной (поедающей рыбу) диете, как это было предложено для этого семейства на основе ископаемых данных и полуводных адаптаций, продемонстрированных многими видами. Также известно, что они питались птерозаврами и другими динозаврами. Остафриказавр жил в субтропической и тропической среде вместе со многими другими динозаврами, а также птерозаврами , крокодилообразными , рыбами, млекопитающими и беспозвоночными . Из-за своего возраста и местоположения Остафриказавр указывает на то, что спинозавриды могли быть распространены по всему миру до распада Пангеи .
История исследований
[ редактировать ]
Во времена Германской колониальной империи Музей естествознания (Museum für Naturkunde ) в Берлине организовал экспедицию в Германскую Восточную Африку (ныне Танзания ), которая проходила с 1909 по 1912 год и сейчас рассматривается учёными как одна из крупнейших экспедиции по палеонтологической истории. Большинство раскопок было проведено в юго-восточной части формации Тендагуру , богатой окаменелостями части бассейна Мандава, датируемой позднеюрским периодом . [1] [2] Среди множества окаменелостей динозавров, найденных на местах раскопок, было 230 образцов зубов теропод . [3] Одним из них был изолированный зуб, занесенный в каталог как MB R 1084, найденный либо неподалеку, либо на вершине холма Тендагуру в Верхней пачке динозавров . [4] Первоначально его отнесли к виду Labrosaurus ? stechowi в 1920 году немецким палеонтологом Вернером Яненшем на основе орнамента, сопоставимого с зубом, описанным как Labrosaurus sulcatus Отниэлем Чарльзом Маршем . [3] В подробной монографии Яненша, опубликованной в 1925 году, L. ? stechowi и разделил их на пять морфотипов (от а до е). [5]
В 2000 году американские палеонтологи Джеймс Мэдсен и Сэмюэл Уэллс назвали L. ? зубы stechowi у Ceratosaurus sp . (неопределенного вида), потому что они напоминали зубы предчелюстной и зубной челюстей цератозавра , теропода из североамериканской формации Моррисон . [6] В 2007 году американский палеонтолог Денвер Фаулер вместо этого предположил, что зубы принадлежали динозавру -спинозавриду , похожему на барионикса , что сделало бы его одним из старейших известных окаменелостей спинозаврид и, таким образом, одним из самых ранних свидетельств существования этой группы. [7] Этот анализ был поддержан французским палеонтологом Эриком Бюффето , который исследовал зубы в том же году, и в статье 2008 года отнес образец MB R 1084 к Spinosauridae. Бюфетаут обнаружил, что этот образец отличался от других зубов, ранее отнесенных к L. ? stechowi , и что другой изолированный зуб (MB R 1091) из средней группы динозавров может представлять то же животное. [8] Он также поставил под сомнение предварительное отнесение Яненшем зубов к сомнительному роду Labrosaurus , которое было основано на скудных останках из формации Моррисон, которые позже были отнесены к Allosaurus . [8] [9] Более того, Баффето отметил, L. что Зуб sulcatus , изображенный Маршем, теперь считается принадлежащим цератозавру . [4] Аналогично Л. , stechowi был отнесен к сомнительному цератозавру , родственному цератозавру . [6] [10]

В 2011 году немецкий палеонтолог Оливер Раухут считал, что зубы среднего динозавра приписывают L. ? stechowi как не имеющий диагностических признаков (уникальных производных признаков), что соответствует тому, что у этого вида сомнительное название. Раухут отметил, что их все еще можно отличить от других зубов теропод из формации Тендагуру, основываясь на их небольшом искривлении и уплощении вбок коронки зуба , а также широких гребнях на язычной стороне (которая обращена внутрь рта). [11] Он признал L. sulcatus сомнительным названием, поскольку зуб, о котором идет речь, был показан только на одной иллюстрации и не описан должным образом в тексте. Кроме того, первоначальные останки лаброзавров не содержали зубов, а появился дополнительный вид, Labrosaurusferox (теперь считающийся синонимом Allosaurus fragillis) . [9] ), был основан на зубной кости, несущей зубы, иные по морфологии, чем у L. sulcatus . Таким образом, Раухут пришел к выводу, что нет никаких оснований приписывать зубы Тендагуру Лаброзаврам . Он предварительно отнес их всех, кроме MB R 1084, к Ceratosaurus (под названием Ceratosaurus ? stechowi ), на основании анатомического сходства с зубами этого рода. По словам Раухута, различные особенности зубов Яненша типа b, c, e и d не представляют собой отдельные таксоны , а скорее представляют собой вариации в зубном ряду на челюстях животного. Раухут также приписал еще один зуб (MB R 1093), описанный Яненшем, но не относящийся к L. ? stechowi , к тому же таксону. [11]
Раухут также обнаружил, что тип а Яненша (MB R 1084) отличается по форме от остальных восьми зубов и, возможно, представляет собой другой таксон, тесно связанный с C. ? стехови . Он перечислил некоторые различия между ним и другими зубами, первоначально отнесенными к L. ? stechowi : MB R 1084 имеет больше язычных гребней (до одиннадцати) и три гребня и бороздки на губной стороне, обращенной к внешней стороне рта. Более того, некоторые гребни MB R 1084 ограничены основанием коронки и соединены более длинными гребнями, простирающимися почти по всей длине коронки, с областью высотой 5 мм (0,20 дюйма) на кончике зуба, лишенной каких-либо украшений. . Кроме того, почти на всех передних трех пятых кроны имеются гребни, тогда как задние две пятых гладкие. В передней части зуба ребристая часть отделена от киля (режущего края) участком, слегка вогнутым спереди назад. Единственное сходство между зубами MB R 1084 и зубами среднего динозавра заключается в их общей форме и плотности расположения. зубцы , так как все зубы имеют 10 зубчиков на 5 миллиметров (0,20 дюйма) на заднем киле и 13 зубчиков на 5 миллиметров (0,20 дюйма) на переднем киле. [11]
В статье 2012 года Баффетаут использовал MB R 1084 в качестве образца голотипа для нового рода и вида Ostafrikasaurus crassiserratus , описав его как раннего теропода-спинозаврида. Его родовое название происходит от немецкого названия колонии, в которой были найдены окаменелости, Deutsch-Ostafrika, что означает «Немецкая Восточная Африка», в сочетании с греческим σαῦρος ( sauros ), что означает «ящерица» или «рептилия». Видовое название происходит от латинского crassus , что означает «толстый»; и serratus , что означает «зазубренный», в отношении больших зазубрин его зубов. Из-за сходства с MB R 1084 Бюффето отнес MB R 1091 из группы средних динозавров к тому же виду. Оба зуба имеют изогнутый передний киль, не имеют кривизны из стороны в сторону и имеют сопоставимую форму в поперечном сечении. Их основные отличия включают в себя наличие у MB R 1091 пяти продольных гребней на язычной стороне по сравнению с десятью у MB R 1084, причем у первого гребни менее обширные. MB R 1091 также имела меньше морщин Зубная эмаль . Баффето отмечает, что эти различия можно объяснить индивидуальные вариации внутри таксона, но поскольку оба зуба произошли от разных членов формации Тендагуру, это отнесение является лишь предварительным. [4]
Бюффето подробно остановился на различиях между зубами Ostafrikasaurus crassiserratus и L. ? морфотипы стехови . Зубы морфотипа b (MB R 1083 и 1087) имели кривизну как спереди назад, так и из стороны в сторону, а также D-образное поперечное сечение. Морфотип c (MB R 1090) был изогнут из стороны в сторону, но не спереди назад, не был уплощен из стороны в сторону, имел закругленную переднюю часть без киля и имел пять сильных гребней на язычной стороне, но ни одного на губной стороне. . Морфотип е (MB R 1092) напоминал типичный зуб теропода. Он сильно уплощен из стороны в сторону, изогнут спереди назад, имеет 3 зубца на миллиметр (0,04 дюйма), его передний киль не доходит до основания коронки, никакой орнаментации, если не считать слабой бороздчатости на макушке. коронка и два зарождающихся гребня на язычной стороне. [4]
В 2020 году исследование зубов из формации Тендагуру и поздней юры Южной Америки снова позволило остафриказавра с цератозаврами идентифицировать . [12]
Описание
[ редактировать ]
В 2016 году испанские палеонтологи Молина-Перес и Ларраменди оценили Остафриказавра примерно в 8,4 метра (28 футов) в длину, 2,1 м (6,9 фута) в высоту в бедрах и вес 1,15 тонны (1,27 коротких тонны ; 1,13 длинных тонны ). [13] Однако без более полного материала, такого как череп или окаменелости тела, невозможно надежно рассчитать размер и вес тела фрагментарных таксонов спинозавров, особенно тех, которые известны только по зубам. Таким образом, оценки являются лишь предварительными. [14]
Зуб голотипа толстый, несколько сплюснутый с боков, его длина сверху вниз составляет 46 миллиметров (1,8 дюйма). Его кончик закруглен эрозией , основание сохранилось не полностью. Коронка зуба имеет четко выраженные кили (режущие края), при этом передний киль изогнут, а задний почти прямой. Имеется лишь небольшое искривление из стороны в сторону. Оба киля зубчатые, с закругленными зубчиками, перпендикулярными краю зуба. Между зубцами нет межзубных бороздок или борозд. Они выстилают передний киль от основания до кончика, а также, вероятно, и задний киль, основание которого сильно эродировано. К кончику зуба эти зубцы сильно стираются (особенно на переднем киле). На переднем киле имеется два зубца на мм (0,04 дюйма) возле кончика зуба и от трех до четырех на мм (0,04 дюйма), поскольку зубцы сжимаются к основанию коронки. По всему заднему килю имеется два зубца на мм (0,04 дюйма). Зубцы заметно крупнее, чем у всех других известных спинозавридов. [4]
Эмаль (самый внешний слой) зуба имеет на поверхности ряд гребней: 10 на язычной стороне и четыре более слабых и менее обширных на лабиальной стороне. Зазоры между выступами составляют не более 1 мм (0,039 дюйма). Ни один из гребней с обеих сторон не достигает кончика кроны. В передней части зуба с обеих сторон имеется область шириной 3 мм (0,12 дюйма) без гребней; аналогичная область на задней стороне зуба уменьшается в ширине с 8 до 4 мм (от 0,31 до 0,16 дюйма) по мере приближения к кончику коронки. С обеих сторон зуба, между гребнями и безгребневыми частями зубов, поверхность эмали мелкоморщинистая. [4]
Классификация
[ редактировать ]
Спинозавриды обычно разделяют на два подсемейства : Baryonychinae и Spinosaurinae . Что касается особенностей зубов, для барионихинов характерны слегка изогнутые, мелкозубчатые зубы с более овальным поперечным сечением, в то время как зубы спинозавров прямые, имеют сильно уменьшенные или полностью отсутствующие зубцы, имеют более круглые поперечные сечения и имеют выступающие канавки (продольные бороздки). ) на их эмали. [14] [15] В 2007 году Фаулер интерпретировал L. ? Зубы stechowi представляют собой возможную примитивную барионихину или предковую форму барионихинов, поскольку они имеют общие черты, такие как плотно упакованные зубцы , толстая форма, слегка уплощенные коронки зубов и гребни на язычной поверхности, обычно связанные с этой кладой . [7] В 2008 году Баффетаут выделил только морфотипы Яненса a (MB R 1084) и d (MB R 1091) как имеющие барионихиновые характеристики, к которым в первом относились общая форма, несколько уплощенное поперечное сечение, мелкоморщинистая эмаль, гребни, не доходящие до кончик зуба и имеет больше гребней на лингвальной, чем на губной стороне. У MB R 1091 только одна сторона зуба имеет гребни, что также можно наблюдать на зубах голотипического экземпляра барионикса . Более того, Баффето добавил, что единственное сходство, которое другие типы (b, c и e) имеют с типами Яненша a и d, - это гребни, покрывающие часть зубной коронки, тогда как во всех других аспектах, таких как их форма и поперечное сечение, они существенно разные. Согласно анализу Баффето, типы b, c и e, вероятно, имеют цератозавридное происхождение, тогда как тип a, вероятно, представляет собой раннего спинозаврида, который отличается от барионихинов раннего мела . [8]
Раухут усомнился в этой интерпретации в 2011 году, заявив, что MB R 1084 имеет больше сходства, чем различий с Цератозавром ? зубы stechowi имеют округлое сечение, лишь краевую кривизну коронки, более выпуклую лингвальную, чем лабиальную сторону, сходные размеры и форму зубцов. Таким образом, согласно его анализу, только количество и распределение гребней остались уникальными для MB R 1084. Раухут отметил, что, хотя барионихины также имеют гребни по обе стороны зубов, они обычно наиболее развиты в задней части зуба, тогда как У MB R 1084 выступов на этой стороне нет. Он также утверждает, что морщины Бюффето, наблюдаемые на эмали MB R 1084, очень слабые и в основном ограничены язычной стороной, по сравнению с более заметной зернистой текстурой зубов Baryonyx . По словам Раухута, хотя MB R 1084 потенциально имеет спинозавридное происхождение, он имеет лишь общее сходство с барионихиновыми зубами и вместо этого, вероятно, представляет собой близкого родственника цератозавра ? Стехови . [11] В 2012 году, назвав Ostafrikasaurus Баффетотом , он поместил его в Spinosauridae, утверждая вместо этого, что зуб очень похож на зубы барионихин, включая барионикса . Среди их общих стоматологических особенностей он включил небольшое уплощение коронки вбок, мелкие морщины эмали и гребни с обеих сторон, которые не достигают кончика зуба и которые сильнее и многочисленнее на лингвальной, чем губной стороне. Если эта идентификация верна, Остафриказавр представляет собой одно из самых ранних известных свидетельств существования спинозавридов. [16]
Именование и различение новых динозавров исключительно по зубам часто считалось палеонтологами проблематичным, например, в связи со спорной идентичностью азиатского рода Siamosaurus . [14] [17] Баффето заявил, что при тщательном сравнении и анализе морфологических особенностей, таких как орнамент, зубы теропод могут быть достаточно диагностическими, чтобы вывести новые таксоны. [16] Более того, зубы спинозавридов имеют уникальную морфологию, очень отличающуюся от зубов других теропод. [16] [18] Тем не менее, точная идентификация Ostafrikasaurus до сих пор неясна, и исследователи указали на сходство в орнаменте его эмали с Ceratosaurus , Paronychodon , Zapsalis , Acheroraptor и Coelophys . [18]
Эволюция
[ редактировать ]
Фаулер в 2007 году выдвинул возможность того, что спинозавриды произошли от предков цератозавров , учитывая, что барионихиновые зубы имеют гребни на коронках, напоминающие те, что наблюдаются на предчелюстных и зубных зубах цератозавров . [7] В 2008 году Баффето отклонил это предложение, сославшись на то, что D-образное поперечное сечение зубов цератозавра отсутствует у барионихин. [8]
Основное отличие MB R 1084 от всех других известных зубов спинозаврид, как отмечал Бюффето, заключалось в большом размере зубчиков, несущих кили. Это привело его в 2008 году к выдвижению гипотезы о том, что эволюция зубов спинозаврид в значительной степени характеризовалась сокращением и возможной потерей зубцов. [8] В 2012 году Баффетаут предположил, что этот образец начнется с крупных зубцов у примитивных юрских таксонов, таких как Ostafrikasaurus (титонского возраста ) , которые напоминали таковые у типичных теропод аналогичного размера. Затем они развились в тонкие, редуцированные и более многочисленные зубцы раннемеловых барионихинов, таких как барионикс из баррема Европы и Зухомим из апта - альба Западной Африки . Барионикс , например, имел семь зубчиков на мм (0,04 дюйма) по сравнению с двумя-четырьмя зубцами у Остафриказавра . Наконец, зубчики полностью исчезнут у спинозавринов, таких как спинозавр, от альба до турона Северной Африки. [15] [16] Баффетот отмечает, что сильно уменьшенные зубцы у таких таксонов, как Siamosaurus из барремского периода Таиланда, по-видимому, представляют собой промежуточную форму. Он также отметил сходство между зубными рядами Остафриказавра и набором бариониксоподобных зубов из доаптской формации Кабао в Ливии, что может иметь важные биогеографические последствия. Эти зубы схожи по общей форме, овальному поперечному сечению и морщинистой поверхности коронки, но отличаются меньшими зубцами и канавками вместо гребней. [16]

В сочетании с исчезающими зубцами поперечные сечения зубов спинозавров изначально были лезвиевидными и линзовидными у более ранних таксонов, таких как Ostafrikasaurus и Baryonyx , и стали более круглыми у производных форм, таких как Spinosaurus . Однако орнаментация зубной эмали спинозавров развивалась более неравномерно, чем их зубцы и форма поперечного сечения. У большинства нептичьих теропод зубы обычно были гладкими, с минимальными морщинами. Зубы остафриказавра , напротив, имели сильные продольные гребни - особенность, также наблюдаемая у барионикса и его близких родственников, хотя их выступающее положение варьируется в зависимости от таксона. Зубы спинозавра , как и у типичных теропод, обычно были гладкими, а у азиатских форм, таких как сиамозавр, наблюдалось увеличение количества зубных гребней. Кроме того, коронка зуба была морщинистой у Ostafrikasaurus , Baryonyx иuchomimus , а также азиатских спинозавридов, но гладкой у Spinosaurus , и только у некоторых экземпляров последних наблюдались мелкие морщинки. [16]
В 2016 году испанский палеонтолог Алеандро Серрано-Мартинес и его коллеги описали возможный зуб спинозаврид, занесенный в каталог как MUPE HB-87, из сланцев Ирхазер в Нигере. Найденный вместе со скелетом динозавра зауропода - Spinophorosaurus , зуб, вероятно, датируется батским этапом средней юры , за 14 миллионов лет до Остафриказавра . Если эта идентификация верна, MUPE HB-87 представляет собой старейшее известное свидетельство существования спинозаврид в летописи окаменелостей. На основе этого экземпляра авторы предложили новую эволюционную модель перехода от «нормальных» зубов теропод к зубам спинозаврид. Зуб овального сечения, загнут назад, имеет небольшие зубцы, а также сильную бороздку, не доходящую до кончика коронки. [18] Однако другие исследователи отметили, что этот экземпляр обладает многими чертами, не относящимися к спинозавридам, и вместо этого эти исследователи отдают предпочтение идентификации мегалозаврид. [19] [12]
Палеобиология
[ редактировать ]не было обнаружено материала черепа Хотя у остафриказавра , известно, что черепа спинозаврид напоминали черепа крокодилов; они были длинными, низкими, узкими и расширенными на передних концах, придавая конечную розеткообразную форму, с прочным вторичным небом на нёбе, что делало их более устойчивыми к нагрузкам и изгибу. Напротив, примитивным и типичным состоянием теропод была высокая, широкая и клиновидная морда с менее развитым вторичным небом. Адаптации черепа спинозаврид совпадали с черепами крокодилов ; ранние представители последней группы имели черепа, похожие на типичные нептичьи (или нептичьи) тероподы, позже у них появились удлиненные морды, конические зубы и вторичное небо. Эта адаптация могла быть результатом перехода от наземной добычи к рыбе. [20] [21] В 2012 году Баффетаут предположил, что уменьшение зазубрин на зубах спинозаврид, проиллюстрированное остафриказавром, может представлять собой переходный период во время этого изменения в рационе. [16] У большинства динозавров-тероподов были изогнутые, похожие на лезвия зубы с зазубренными килями для разрезания плоти, тогда как зубы спинозаврид стали более прямыми, более коническими и имели небольшие или отсутствующие зубцы. Такой зубной ряд наблюдается у ныне живущих рыбоядных хищников, таких как гавиалы , поскольку он лучше подходит для прокалывания и удержания скользкой водной добычи, поэтому ее можно проглотить целиком, а не разорвать на части. [14] [20] [22]
Палеосреда и палеобиогеография
[ редактировать ]
Верхняя пачка динозавров формации Тендагуру сложена преимущественно алевролитами , известковистыми песчаниками и аргиллитов пластами . Эти породы, вероятно, относятся к титонскому этапу позднеюрского периода, примерно от 152,1 до 145 миллионов лет назад. [2] [23] Однако точная хронологическая граница между ранним мелом и поздней юрой формации Тендагуру до сих пор неясна. [4] Среда обитания Остафриказавра была от субтропической до тропической , с периодическими дождями и ярко выраженными засушливыми сезонами. В формации Тендагуру присутствовало три типа палеосред: первый представлял собой мелководную морскую обстановку с условиями, напоминающими лагуны , защищенными отмелями ооидных пород, очевидно , и кремнеобломочных подверженными приливам и штормам. Второй представлял собой прибрежную среду с приливными отмелями , состоящую из солоноватых озер, прудов и речных каналов . В этой экосистеме было мало растительной жизни, которой могли бы питаться динозавры-зауроподы, и большинство динозавров, вероятно, приходили в этот район только во время засухи. В третьей и наиболее внутренней среде обитания преобладали хвойные растения на хорошо заросшей территории, что представляло собой большую кормовую площадку для зауроподов. [24]
Формация Тендагуру была домом для множества разнообразных организмов. беспозвоночные , такие как двустворчатые моллюски , брюхоногие моллюски , устрицы , иглокожие , членистоногие , брахиоподы , кораллы и многие микрофауны , включая остракоды , фораминиферы , харофиты и палиноморфы . Из отложений известны [25] [2] В регионе были широко распространены динозавры-зауроподы, представленные Giraffatitan Branchai , Dicraeosaurus hansemanni и D. sattleri , Australodocus bohetii , Janenschia Robusta Tornieria africana , Tendaguria tanzaniensis , Wamweracaudia keranjei , а также неопределенными диплодоцидами , flagellicaudatans и туриазаврами. , [26] [27] [28] Они бы сосуществовали с малопросматривающими птицетазовыми животными, такими как орнитопод Dysalotosaurus lettowvorbecki и стегозавр Kentrosaurus aethiopicus . [2] [29] Тероподы, помимо Ostafrikasaurus, включали кархародонтозаврид Veterupristisaurus milneri и ноазавридов Elaphrosaurus bambergi . Фрагментарные окаменелости также указывают на присутствие базальных цератозавридов ( Ceratosaurus ? stechowi ) и тетануранов , неопознанных абелизавроидов , а также возможных абелизавридов , кархародонтозавридов и мегалозавроидов . [11]

Среди других позвоночных, населявших эту среду, были птерозавры, такие как Tendaguripterus Recki , а также неопределенный джунгариптероид , аждархид и, возможно, археоптеродактилоид . Был также карликовый крокодилообразный Bernissartia sp., [30] [31] лисамфибийные амфибии, парамацеллодидные ящерицы и различные мелкие млекопитающие , [2] в том числе Brancatherulum tendaguense , [32] Allostaffia enigmatica , Tendagurodon janenschi , Tendagurutherium Dietrichi и множество неопознанных симметродонтов . [2] Окаменелости актиноптеригиев Lepidotes tenguriensis и L. sp. , скат такие Engaibatis schultzei и акулы -гибодонты, как Hybodus sp ., Lonchidion sp. и Sphenodus sp. [25] [33] Флора араукариев формации Тендагуру была одинаково разнообразна: растительность состояла , , кипарисов , саговников гинкговых тисов , хейролепидиевых , из и празинофитов . Встречались также динофлагелляты и Zygnemataceae водоросли , а также многочисленные таксоны пыльцы и спор . [33] [34] [35]
В 2007 году Фаулер отметил, что барионихиновая идентификация L. ? Зубы stechowi соответствовали бы биогеографическим моделям, предложенным в то время для эволюции и распространения спинозавров, которые предполагали происхождение группы на южном суперконтиненте Гондвана с последующим распространением и диверсификацией в Европе. [7] В 2012 году Баффетот отметил, что присутствие такого базального рода, как Ostafrikasaurus, в Африке не обязательно указывает на то, что там произошли спинозавриды. Вместо этого группа, возможно, была широко распространена на ранних стадиях своей эволюционной истории, когда другие таксоны населяли Северную Америку, Европу и остальную часть Гондваны одновременно с Остафриказавром . Баффето считал это особенно вероятным из-за открытия окаменелостей спинозаврид в Азии, которая, вероятно, была отделена от других континентов на протяжении большей части позднеюрского и раннего мелового периода, поскольку Пангея продолжала распадаться, поэтому до сих пор неясно, как группа прибыла в Азию. [4] Раннее глобальное распространение группы также считалось вероятным такие авторы, как Стивен Брусатт и его коллеги в 2010 году. [36] и Ронан Аллен и его коллеги в 2012 году, причем последний предположил, что такое распространение могло произойти раньше по Пангее, до ее распада, начавшегося в поздней юре. [37] Палеобиогеография спинозавридов остается плохо изученной и, вероятно, была очень сложной, учитывая открытия в Азии и, возможно, Австралии, а также сходство между некоторыми европейскими и азиатскими таксонами. [18] [38] [39]
Ссылки
[ редактировать ]- ^ Тамборини, Марко; Веннен, Марейке (5 июня 2017 г.). «Нарушения и изменение привычек: случай экспедиции Тендагуру». Журнал истории музея . 10 (2): 183–199. дои : 10.1080/19369816.2017.1328872 . ISSN 1936-9816 . S2CID 164402709 .
- ^ Перейти обратно: а б с д и ж Буссерт, Роберт; Генрих, Вольф-Дитер; Аберхан, Мартин (1 августа 2009 г.). «Формация Тендагуру (от поздней юры до раннего мела, южная Танзания): определение, палеосреда и стратиграфия последовательностей» . Ископаемый рекорд . 12 (2): 141–174. дои : 10.1002/mmng.200900004 .
- ^ Перейти обратно: а б В. Яненш, 1920, «О Elaphrosaurus bambergi и мегалозаврах из слоев Тендагуру Германской Восточной Африки», труды Общества друзей естественных исследований в Берлине, 1920 : 225–235.
- ^ Перейти обратно: а б с д и ж г час Баффето, Эрик (2012). «Ранний спинозавридный динозавр из поздней юры Тендагуру (Танзания) и эволюция зубного ряда спинозаврид» (PDF) . Ориктос . 10 : 1–8.
- ^ Яненш, В., 1925, «Целурозавры и тероподы слоев Тендагуру в Германской Восточной Африке», Palaeontographica 7: 1–99. Приложение
- ^ Перейти обратно: а б Мэдсен, Джеймс Х.; Уэллс, Сэмюэл П. (2000). Цератозавр (Dinosauria, Theropoda), пересмотренная остеология . Разные публикации, 00-2 . Геологическая служба Юты.
- ^ Перейти обратно: а б с д Фаулер, Д.В. (2007). «Недавно открытые барионихиновые зубы (Dinosauria: Theropoda): новые морфологические данные, расширение ареала и сходство с цератозавром » . Журнал палеонтологии позвоночных . 27 (3): 3.
- ^ Перейти обратно: а б с д и Баффето, Эрик (2008). «Зубы спинозаврид из поздней юры Тендагуру, Танзания, с замечаниями по эволюционной и биогеографической истории спинозавридов» . Документы геологических лабораторий Лиона . 164 : 26–28.
- ^ Перейти обратно: а б Хольц, Томас Р. мл. ; Мольнар, Ральф Э.; Карри, Филип Дж. (2004). «Базальная столбняк». В Вейшампеле Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 71–110. ISBN 978-0-520-24209-8 .
- ^ Тыкоски, Рональд С.; и Роу, Тимоти. (2004). «Цератозаврия» в «Динозаврии » (2-й). 47–70.
- ^ Перейти обратно: а б с д и Раухут, Оливер В.М. (2011). «Тероподовые динозавры поздней юры Тендагуру (Танзания)» . Специальные статьи по палеонтологии . 86 : 195–239.
- ^ Перейти обратно: а б Сото, Матиас; Турино, Пол; Переа, Дэниел (01 ноября 2020 г.). « Зубы цератозавра (Theropoda, Ceratosauria) из формации Такуарембо (поздняя юра, Уругвай)» . Журнал южноамериканских наук о Земле . 103 : 102781. Бибкод : 2020JSAES.10302781S . doi : 10.1016/j.jsames.2020.102781 . ISSN 0895-9811 . S2CID 224842133 .
- ^ Молина-Перес и Ларраменди (2016). Записи и курьезы динозавров Тероподы и другие динозавроморфы . Барселона, Испания: Ларусс. п. 275. ИСБН 9780565094973 .
- ^ Перейти обратно: а б с д Хоун, Дэвид Уильям Эллиотт; Хольц, Томас Ричард (июнь 2017 г.). «Век спинозавров – обзор и пересмотр спинозавридов с комментариями по их экологии» . Acta Geologica Sinica — английское издание . 91 (3): 1120–1132. дои : 10.1111/1755-6724.13328 . ISSN 1000-9515 . S2CID 90952478 .
- ^ Перейти обратно: а б Бюффето, Э.; Сутитхорн, В.; Тонг, Х.; Амиот, Р. (2008). «Раннемеловой теропод спинозавра из южного Китая». Геологический журнал . 145 (5): 745–748. Бибкод : 2008GeoM..145..745B . дои : 10.1017/S0016756808005360 . S2CID 129921019 .
- ^ Перейти обратно: а б с д и ж г Баффето, Эрик (2012). «Ранний спинозавридный динозавр из поздней юры Тендагуру (Танзания) и эволюция зубного ряда спинозаврид» (PDF) . Ориктос . 10 : 1–8.
- ^ Продажи, Маркос АФ; Шульц, Сезар Л. (06 ноября 2017 г.). «Таксономия спинозавров и эволюция краниодентальных особенностей: данные из Бразилии» . ПЛОС ОДИН . 12 (11): e0187070. Бибкод : 2017PLoSO..1287070S . дои : 10.1371/journal.pone.0187070 . ISSN 1932-6203 . ПМЦ 5673194 . ПМИД 29107966 .
- ^ Перейти обратно: а б с д Серрано-Мартинес, Алехандро; Видаль, Дэниел; Шишио, Лара; Ортега, Франциско; Нолл, Фабьен (01 июня 2016 г.). «Изолированные зубы теропод из средней юры Нигера и ранняя стоматологическая эволюция спинозавридов» . Acta Palaeontologica Polonica . 61 (2): 403–415. дои : 10.4202/app.00101.2014 . hdl : 10261/152148 . ISSN 0567-7920 . S2CID 53331040 .
- ^ Хендрикс, Кристоф; Мэтью, Октавио; Араужо, Рикардо; Шуаньер, Иона (2019). «Распределение особенностей зубов у нептичьих динозавров-теропод: таксономический потенциал, степень гомоплазии и основные эволюционные тенденции» . Электронная палеонтология . дои : 10.26879/820 . hdl : 11336/146011 . ISSN 1935-3952 .
- ^ Перейти обратно: а б Хольц-младший, TR (1998). «Спинозавры как подражатели крокодилам». Наука . 282 (5392): 1276–1277. дои : 10.1126/science.282.5392.1276 . S2CID 16701711 .
- ^ Ибрагим, Н.; Серено, ПК; Даль Сассо, К.; Магануко, С.; Фабри, М.; Мартилл, DM; Зухри, С.; Мирвольд, Н.; Лурино, Д.А. (2014). «Полуводные адаптации гигантского хищного динозавра» . Наука . 345 (6204): 1613–1616. Бибкод : 2014Sci...345.1613I . дои : 10.1126/science.1258750 . ПМИД 25213375 . S2CID 34421257 . Дополнительная информация
- ^ Кафф, Эндрю Р.; Рэйфилд, Эмили Дж. (2013). «Механика питания спинозавровых теропод и современных крокодилов» . ПЛОС ОДИН . 8 (5): e65295. Бибкод : 2013PLoSO...865295C . дои : 10.1371/journal.pone.0065295 . ISSN 1932-6203 . ПМЦ 3665537 . ПМИД 23724135 .
- ^ «ICS – График/Шкала времени» . www.stratigraphy.org . Проверено 13 июля 2018 г.
- ^ Аберхан, Мартин; Буссерт, Р.; Генрих, Вольф-Дитер; Шранк, Э; Шультка, Стефан; Самес, Бенджамин; Кривет, Юрген; Капилима, С (1 января 2002 г.). «Палеоэкология и условия осадконакопления слоев Тендагуру (от поздней юры до раннего мела, Танзания)» . Ископаемый рекорд . 5 : 19–44. doi : 10.5194/fr-5-19-2002 .
- ^ Перейти обратно: а б Арратия, Глория; Кривет, Юрген; Генрих, Вольф-Дитер (2002). «Селачи и актиноптериги из верхней юры Тендагуру, Танзания» . Ископаемый рекорд . 5 (1): 207–230. дои : 10.1002/mmng.20020050112 . ISSN 1860-1014 .
- ^ Ремес, Кристиан (2009). «Таксономия позднеюрских диплодоцидных зауроподов из Тендагуру (Танзания)» . Ископаемый рекорд . 12 (1): 23–46. дои : 10.1002/mmng.200800008 . ISSN 1860-1014 .
- ^ Мэтьюз, О.В.; Мэннион, Пенсильвания; Апчерч, П. (2014). « Zby atlanticus , новый туриазавровый зауропод (Dinosauria, Eusauropoda) из поздней юры Португалии» Журнал палеонтологии позвоночных . 34 (3):618.doi : 10.1080 /02724634.2013.822875 . S2CID 59387149 .
- ^ Филип Д. Мэннион, Пол Апчерч, Даниэла Шварц, Оливер Уингс, 2019, «Таксономическое сходство предполагаемых титанозавров из позднеюрской формации Тендагуру в Танзании: филогенетические и биогеографические последствия для эволюции динозавров-эузавропод», Зоологический журнал Линнеевского общества , zly068 , https://doi.org/10.1093/zoolinnean/zly068
- ^ Том Р. Хюбнер и Оливер В.М. Раухут (2010). «Молодой череп Dysalotosaurus lettowvorbecki (Ornithischia: Iguanodontia) и его значение для краниального онтогенеза, филогении и таксономии динозавров-орнитопод» . Зоологический журнал Линнеевского общества . 160 (2): 366–396. дои : 10.1111/j.1096-3642.2010.00620.x .
- ^ Коста, Фабиана Р.; Келлнер, Александр В.А. (2009). «На двух плечевых птерозаврах из слоев Тендагуру (верхняя юра, Танзания)» . Анналы Бразильской академии наук . 81 (4): 813–818. дои : 10.1590/S0001-37652009000400017 . ISSN 0001-3765 . ПМИД 19893904 .
- ^ Барретт, премьер-министр; Батлер, Р.Дж.; Эдвардс, Северная Каролина; Милнер., Арканзас (2008). «Распределение птерозавров во времени и пространстве: атлас, стр. 61–107, в журнале Flugsaurier: статьи о птерозаврах в честь Питера Велнхофера – Хоне, DWE, и Баффетаута, Э. (ред.)» (PDF) . Зиттелиана . 28 : 1–264 — в epub.
- ^ Аверьянов, АО; Мартин, Т. (2015). «Онтогенез и систематика Paurodon valens (Mammalia, Cladotheria) из верхнеюрской формации Моррисон США» . Труды Зоологического института РАН . 319 (3): 326–340. дои : 10.31610/трудизин/2015.319.3.326 . S2CID 91551517 .
- ^ Перейти обратно: а б Аберхан, Мартин; Буссерт, Роберт; Генрих, Вольф-Дитер; Шранк, Экхарт; Шультка, Стефан; Самес, Бенджамин; Кривет, Юрген; Капилима, Саиди (2002). «Палеоэкология и условия осадконакопления слоев Тендагуру (от поздней юры до раннего мела, Танзания)» . Ископаемый рекорд . 5 (1): 19–44. дои : 10.1002/mmng.20020050103 . ISSN 1860-1014 .
- ^ Калерт, Эберхард; Шультка, Стефан; Сладкий, Герберт (1999). «Мезофитная флора отложения динозавров Тендагуру (Танзания). Первые результаты» . Fossil Record (на немецком языке). 2 (1): 185–199. дои : 10.1002/mmng.1999.4860020114 . ISSN 1860-1014 .
- ^ Шранк, Э. (1 января 1999 г.). «Палинология отложений динозавров Тендагуру (Танзания) — предварительные результаты» . Ископаемый рекорд . 2 (1): 171–183. doi : 10.5194/fr-2-171-1999 . ISSN 2193-0074 .
- ^ Брусатте, Стивен; Би Джей Бенсон, R; Сюй, Син (10 декабря 2010 г.). «Эволюция крупнотелых динозавров в мезозое в Азии» . Журнал иберийской геологии . 36 (2): 275–296. дои : 10.5209/rev_JIGE.2010.v36.n2.12 .
- ^ Аллен, Р.; Ксайсанавонг, Т.; Ричир, П.; Хентавонг, Б. (2012). «Первый окончательный азиатский спинозаврид (Dinosauria: Theropoda) из раннего мела Лаоса». Naturwissenschaften . 99 (5): 369–377. Бибкод : 2012NW.....99..369A . дои : 10.1007/s00114-012-0911-7 . ПМИД 22528021 . S2CID 2647367 .
- ^ Мэтью, О.; Араужо, Р.; Натарио, К.; Кастанинья, Р. (2011). «Новый экземпляр тероподного динозавра барионикса из раннего мела Португалии и таксономическая достоверность Сухозавра » (PDF) . Зоотакса . 2827. 2827 : 54–68. дои : 10.11646/zootaxa.2827.1.3 .
- ^ Элизабет Малафайя; Хосе Майкл Гасулла; Фердинанд Скарс; Иван Нарваез; Хосе Луис Санс; Франсиско Ортега (2019). «Новый теропод спинозаврид (Dinosauria: Megalosauroidea) из позднего баррема Валлибоны, Испания: значение для разнообразия спинозавридов в раннем мелу Пиренейского полуострова». Меловые исследования. в печати: 104221. два : 10.1016/j.cretres.2019.104221 .














