Тираннозавриды
| Тираннозавриды Временной диапазон: поздний мел , | |
|---|---|
 | |
| Монтаж шести тираннозаврид, по часовой стрелке сверху слева: Тираннозавр , Дасплетозавр , Тарбозавр , Горгозавр , Чжученгтираннус и Алиорам. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Superfamily: | †Tyrannosauroidea |
| Clade: | †Pantyrannosauria |
| Clade: | †Eutyrannosauria |
| Family: | †Tyrannosauridae Osborn, 1906 |
| Type genus | |
| †Tyrannosaurus Osborn, 1905 | |
| Subgroups[1] | |
| Synonyms | |
Tyrannosaurides (или tyrannosaurids , что означает « ящерицы -тираны ») — семейство целурозавровых , -теропод динозавров включающее два подсемейства, содержащие до пятнадцати родов , включая одноименного Tyrannosaurus . Точное количество родов является спорным: некоторые эксперты признают всего три. Все эти животные жили ближе к концу мелового периода , и их окаменелости были найдены только в Северной Америке и Азии .
Хотя тираннозавриды произошли от более мелких предков , они почти всегда были крупнейшими хищниками в своих экосистемах , ставя их на вершину цепи пищевой . Самым крупным видом был тираннозавр рекс , самый массивный из известных наземных хищников, длина которого превышала 13 метров (43 фута). [2] и по самым современным оценкам до 8,87 метрических тонн (9,78 коротких тонн) в весе. [3] [4] Тираннозавриды были двуногими хищниками с массивными черепами , заполненными крупными зубами. Несмотря на большой размер, их ноги были длинными и приспособленными для быстрого передвижения. Напротив, их руки были очень маленькими и имели только два функциональных пальца .
В отличие от большинства других групп динозавров, у большинства известных тираннозаврид были обнаружены очень полные останки. Это позволило провести разнообразные исследования их биологии . на их онтогенезе , биомеханике и экологии Научные исследования были сосредоточены , среди прочего, .
History of discovery
[edit]
The first remains of tyrannosaurids were uncovered during expeditions led by the Geological Survey of Canada, which located numerous scattered teeth. These distinctive dinosaur teeth were given the name Deinodon ("terrible tooth") by Joseph Leidy in 1856. The first good specimens of a tyrannosaurid were found in the Horseshoe Canyon Formation of Alberta, and consisted of nearly complete skulls with partial skeletons. These remains were first studied by Edward Drinker Cope in 1876, who considered them a species of the eastern tyrannosauroid Dryptosaurus. In 1905, Henry Fairfield Osborn recognized that the Alberta remains differed considerably from Dryptosaurus, and coined a new name for them: Albertosaurus sarcophagus ("flesh-eating Alberta lizard").[5] Cope described more tyrannosaur material in 1892, in the form of isolated vertebrae, and gave this animal the name Manospondylus gigas. This discovery was mostly overlooked for over a century, and caused controversy in the early 2000s when it was discovered that this material actually belonged to, and had name priority over, Tyrannosaurus rex.[6]

In his 1905 paper naming Albertosaurus, Osborn described two additional tyrannosaur specimens that had been collected in Montana and Wyoming during a 1902 expedition of the American Museum of Natural History, led by Barnum Brown. Initially, Osborn considered these to be distinct species. The first, he named Dynamosaurus imperiosus ("emperor power lizard"), and the second, Tyrannosaurus rex ("king tyrant lizard"). A year later, Osborn recognized that these two specimens actually came from the same species. Despite the fact that Dynamosaurus had been found first, the name Tyrannosaurus had appeared one page earlier in his original article describing both specimens. Therefore, according to the International Code of Zoological Nomenclature (ICZN), the name Tyrannosaurus was used.[7]
Barnum Brown went on to collect several more tyrannosaurid specimens from Alberta, including the first to preserve the shortened, two-fingered forelimbs characteristic of the group (which Lawrence Lambe named Gorgosaurus libratus, "balanced fierce lizard", in 1914). A second significant find attributed to Gorgosaurus was made in 1942, in the form of a well-preserved, though unusually small, complete skull. The specimen waited until after the end of World War II to be studied by Charles W. Gilmore, who named it Gorgosaurus lancesnis.[5] This skull was re-studied by Robert T. Bakker, Phil Currie, and Michael Williams in 1988, and assigned to the new genus Nanotyrannus.[8] It was also in 1946 that paleontologists from the Soviet Union began expeditions into Mongolia, and uncovered the first tyrannosaur remains from Asia. Evgeny Maleev described new Mongolian species of Tyrannosaurus and Gorgosaurus in 1955, and one new genus: Tarbosaurus ("terrifying lizard"). Subsequent studies, however, showed that all of Maleev's tyrannosaur species were actually one species of Tarbosaurus at different stages of growth. A second species of Mongolian tyrannosaurid was found later, described by Sergei Kurzanov in 1976, and given the name Alioramus remotus ("remote different branch"), though its status as a true tyrannosaurid and not a more primitive tyrannosaur is still controversial.[9][5]
Description
[edit]
The tyrannosaurids were all large animals, with all species capable of weighing at least 1 metric ton.[10] A single specimen of Alioramus of an individual estimated at between 5 and 6 metres (16 and 20 ft) long has been discovered,[9] although it is considered by some experts to be a juvenile.[10][11] Albertosaurus, Gorgosaurus and Daspletosaurus all measured between 8 and 9 metres (26 and 30 ft) long,[12] while Tarbosaurus reached lengths of 11 metres (36 ft) from snout to tail.[13] The massive Tyrannosaurus reached 13 metres (43 ft) in the largest specimen, RSM P2523.8.[14]
Skull and dentition
[edit]



Tyrannosaurid skull anatomy is well understood, as complete skulls are known for all genera but Alioramus, which is known only from partial skull remains.[15] Tyrannosaurus, Tarbosaurus, and Daspletosaurus had skulls that exceeded 1 m (3.3 ft) in length.[12] Adult tyrannosaurids had tall, massive skulls, with many bones fused and reinforced for strength. At the same time, hollow chambers within many skull bones and large openings (fenestrae) between those bones helped to reduce skull weight. Many features of tyrannosaurid skulls were also found in their immediate ancestors, including tall premaxillae and fused nasal bones.[10]
Tyrannosaurid skulls had many unique characteristics, including fused parietal bones with a prominent sagittal crest, which ran longitudinally along the sagittal suture and separated the two supratemporal fenestrae on the skull roof. Behind these fenestrae, tyrannosaurids had a characteristically tall nuchal crest, which also arose from the parietals but ran along a transverse plane rather than longitudinally. The nuchal crest was especially well-developed in Tyrannosaurus, Tarbosaurus and Alioramus. Albertosaurus, Daspletosaurus and Gorgosaurus had tall crests in front of the eyes on the lacrimal bones, while Tarbosaurus and Tyrannosaurus had extremely thickened postorbital bones forming crescent-shaped crests behind the eyes. Alioramus had a row of six bony crests on top of its snout, arising from the nasal bones; lower crests have been reported on some specimens of Daspletosaurus and Tarbosaurus, as well as the more basal tyrannosauroid Appalachiosaurus.[11][16] The snout and other parts of the skull also sported numerous foramina. According to the 2017 study which described D. horneri, scaly integument as well as tactile sensitivity was correlated with the multiple rows of neurovascular foramina seen in crocodilians and tyrannosaurids.[17]


Tyrannosaurids, like their tyrannosauroid ancestors, were heterodonts, with premaxillary teeth D-shaped in cross section and smaller than the rest. Unlike earlier tyrannosauroids and most other theropods, the maxillary and mandibular teeth of mature tyrannosaurids are not blade-like but extremely thickened and often circular in cross-section, with some species having reduced serrations.[10] Tooth counts tend to be consistent within species, and larger species tend to have lower tooth counts than smaller ones. For example, Alioramus had 76 to 78 teeth in its jaws, while Tyrannosaurus had between 54 and 60.[18]
William Abler observed in 2001 that Albertosaurus tooth serrations resemble a crack in the tooth ending in a round void called an ampulla.[19] Tyrannosaurid teeth were used as holdfasts for pulling meat off a body, so when a tyrannosaur would have pulled back on a piece of meat, the tension could cause a purely crack-like serration to spread through the tooth.[19] However, the presence of the ampulla would have distributed these forces over a larger surface area, and lessened the risk of damage to the tooth under strain.[19] The presence of incisions ending in voids has parallels in human engineering. Guitar makers use incisions ending in voids to, as Abler describes, "impart alternating regions of flexibility and rigidity" to the wood they work with.[19] The use of a drill to create an "ampulla" of sorts and prevent the propagation of cracks through material is also used to protect airplane surfaces.[19] Abler demonstrated that a plexiglass bar with incisions called "kerfs" and drilled holes was more than 25% stronger than one with only regularly placed incisions.[19] Unlike tyrannosaurs and other theropods, ancient predators like phytosaurs and Dimetrodon had no adaptations to prevent the crack-like serrations of their teeth from spreading when subjected to the forces of feeding.[19]
Postcranial skeleton
[edit]
The skull was perched at the end of a thick, S-shaped neck, and a long, heavy tail acted as a counterweight to balance out the head and torso, with the center of mass over the hips. Tyrannosaurids are known for their proportionately very small two-fingered forelimbs, although remnants of a vestigial third digit are sometimes found.[10][20] Tarbosaurus had the shortest forelimbs compared to its body size, while Daspletosaurus had the longest.

Tyrannosaurids walked exclusively on their hindlimbs, so their leg bones were massive. In contrast to the forelimbs, the hindlimbs were longer compared to body size than almost any other theropods. Juveniles and even some smaller adults, like more basal tyrannosauroids, had longer tibiae than femora, a characteristic of cursorial (fast-running) dinosaurs like ornithomimids. Larger adults had leg proportions characteristic of slower-moving animals, but not to the extent seen in other large theropods like abelisaurids or carnosaurs. The third metatarsals of tyrannosaurids were pinched between the second and fourth metatarsals, forming a structure known as the arctometatarsus.[10] Tyrannosaurids also had large preserved leg muscle attachments and low rotational inertia relative to their body mass, indicating that they could turn more quickly than other large theropods.[21][22]
It is unclear when the arctometatarsus first evolved; it was not present in the earliest tyrannosauroids like Dilong,[23] but was found in the later Appalachiosaurus.[16] This structure also characterized troodontids, ornithomimids and caenagnathids,[24] but its absence in the earliest tyrannosauroids indicates that it was acquired by convergent evolution.[23]
Soft tissue reconstruction
[edit]A comparative analysis of the teeth suggests that tyrannosaurids, as well as the other large theropods, had lips that protected their teeth from external damage. This anatomical feature gave these animals more visual resemblance to lepidosaurs than to closely related crocodilians.[25]
Classification
[edit]The name Deinodontidae was coined by Edward Drinker Cope in 1866 for this family,[26] and continued to be used in place of the newer name Tyrannosauridae through the 1960s.[27] The type genus of the Deinodontidae is Deinodon, which was named after isolated teeth from Montana.[28] However, in a 1970 review of North American tyrannosaurs, Dale Russell concluded that Deinodon was not a valid taxon, and used the name Tyrannosauridae in place of Deinodontidae, stating that this was in accordance with ICZN rules.[12] Therefore, Tyrannosauridae is preferred by modern experts.[5]
Tyrannosaurus was named by Henry Fairfield Osborn in 1905, along with the family Tyrannosauridae.[29] The name is derived from the Ancient Greek words τυραννος (tyrannos) ('tyrant') and σαυρος (sauros) ('lizard'). The very common suffix -idae is normally appended to zoological family names and is derived from the Greek suffix -ιδαι -idai, which indicates a plural noun.[30]
Taxonomy
[edit]
Tyrannosauridae is a family in rank-based Linnaean taxonomy, within the superfamily Tyrannosauroidea and the suborder Theropoda.
Tyrannosauridae is uncontroversially divided into two subfamilies. Albertosaurinae comprises the North American genera Albertosaurus and Gorgosaurus, while Tyrannosaurinae includes Daspletosaurus, Teratophoneus, Bistahieversor, Tarbosaurus, Nanuqsaurus, Zhuchengtyrannus, and Tyrannosaurus itself.[31] Some authors include the species Gorgosaurus libratus in the genus Albertosaurus and Tarbosaurus bataar in the genus Tyrannosaurus,[16][5][32] while others prefer to retain Gorgosaurus and Tarbosaurus as separate genera.[10][11] Albertosaurines are characterized by more slender builds, lower skulls, and proportionately longer tibiae than tyrannosaurines.[10] In tyrannosaurines, the sagittal crest on the parietals continues forward onto the frontals.[11] In 2014, Lü Junchang et al. described the Alioramini as a tribe within the Tyrannosauridae containing the genera Alioramus and Qianzhousaurus. Their phylogenetic analysis indicated that the tribe was located at the base of the Tyrannosaurinae.[33][34] Some authors, such as George Olshevsky and Tracy Ford, have created other subdivisions or tribes for various combinations of tyrannosaurids within the subfamilies.[35][36] However, these have not been phylogenetically defined, and usually consisted of genera that are now considered synonymous with other genera or species.[18]
Additional subfamilies have been named for more fragmentary genera, including Aublysodontinae and Deinodontinae. However, the genera Aublysodon and Deinodon are usually considered nomina dubia, so they and their eponymous subfamilies are usually excluded from taxonomies of tyrannosaurids. An additional tyrannosaurid, Raptorex, was initially described as a more primitive tyrannosauroid, but likely represents a juvenile tyrannosaurine similar to Tarbosaurus. However, as it is known only from a juvenile specimen, it is also currently considered a nomen dubium.[37] However, Thomas Carr maintains its validity and finds it to be similar to tyrannosaurines.[38]
Phylogeny
[edit]With the advent of phylogenetic taxonomy in vertebrate paleontology, Tyrannosauridae has been given several explicit definitions. The original was produced by Paul Sereno in 1998, and included all tyrannosauroids closer to Tyrannosaurus than to either Alectrosaurus, Aublysodon or Nanotyrannus.[39] However, Nanotyrannus is often considered to be a juvenile Tyrannosaurus rex, while Aublysodon is usually regarded as a nomen dubium unsuitable for use in the definition of a clade.[10] Definitions since then have been based on more well-established genera.
In 2001, Thomas R. Holtz Jr. published a cladistic analysis of the Tyrannosauridae.[40] He concluded that there were two subfamilies: the more primitive Aublysodontinae, characterized by unserrated premaxillary teeth; and the Tyrannosaurinae.[40] The Aublysodontinae included Aublysodon, the "Kirtland Aublysodon", and Alectrosaurus.[40] Holtz also found that Siamotyrannus exhibited some of the synapomorphies of the tyrannosauridae, but lay "outside the [family] proper."[40]
Later in the same paper, he proposed that Tyrannosauridae be defined as "all descendants of the most recent common ancestor of Tyrannosaurus and Aublysodon".[40] He also criticized definitions previously proposed by other workers, like one proposed by Paul Sereno, that the Tyrannosauridae was "all taxa closer to "Tyrannosaurus" than to Alectrosaurus, Aublysodon, and Nanotyrannus".[40] Holtz observed that since Nanotyrannus was probably a misidentified T. rex juvenile, Sereno's proposed definition would have the family Tyrannosauridae as a subtaxon of the genus Tyrannosaurus.[40] Further, his proposed definition of the subfamily Tyrannosaurinae would also be limited to Tyrannosaurus.[40]
A 2003 attempt by Christopher Brochu included Albertosaurus, Alectrosaurus, Alioramus, Daspletosaurus, Gorgosaurus, Tarbosaurus and Tyrannosaurus in the definition.[41] Holtz redefined the clade in 2004 to use all of the above as specifiers except for Alioramus and Alectrosaurus, which his analysis could not place with certainty. However, in the same paper, Holtz also provided a completely different definition, including all theropods more closely related to Tyrannosaurus than to Eotyrannus.[10] The most recent definition is that of Sereno in 2005, which defined Tyrannosauridae as the least inclusive clade containing Albertosaurus, Gorgosaurus and Tyrannosaurus.[42]
Cladistic analyses of tyrannosaurid phylogeny often find Tarbosaurus and Tyrannosaurus to be sister taxa, with Daspletosaurus more basal than either. A close relationship between Tarbosaurus and Tyrannosaurus is supported by numerous skull features, including the pattern of sutures between certain bones, the presence of a crescent-shaped crest on the postorbital bone behind each eye, and a very deep maxilla with a noticeable downward curve on the lower edge, among others.[10][16] An alternative hypothesis was presented in a 2003 study by Phil Currie and colleagues, which found weak support for Daspletosaurus as a basal member of a clade also including Tarbosaurus and Alioramus, both from Asia, based on the absence of a bony prong connecting the nasal and lacrimal bones.[18] Alioramus was found to be the closest relative of Tarbosaurus in this study, based on a similar pattern of stress distribution in the skull.
A related study also noted a locking mechanism in the lower jaw shared between the two genera.[43] In a separate paper, Currie noted the possibility that Alioramus might represent a juvenile Tarbosaurus, but stated that the much higher tooth count and more prominent nasal crests in Alioramus suggest it is a distinct genus. Similarly, Currie uses the high tooth count of Nanotyrannus to suggest that it may be a distinct genus,[11] rather than a juvenile Tyrannosaurus as most other experts believe.[10][44] However, the discovery and description of Qianzhousaurus reveals that Alioramus is not a close relation to Tarbosaurus, instead belonging to a newly described tribe of tyrannosaurids; the Alioramini. Qianzhousaurus further reveals that similar long-snouted tyrannosaurids were widely distributed throughout Asia and would have shared the same environment while avoiding competition with larger and more robust tyrannosaurines by hunting different prey.[45]
|
|
|
Paleobiology
[edit]Growth
[edit]
Paleontologist Gregory Erickson and colleagues have studied the growth and life history of tyrannosaurids. Analysis of bone histology can determine the age of a specimen when it died. Growth rates can be examined when the age of various individuals are plotted against their size on a graph. Erickson has shown that after a long time as juveniles, tyrannosaurs underwent tremendous growth spurts for about four years midway through their lives. After the rapid growth phase ended with sexual maturity, growth slowed down considerably in adult animals. A tyrannosaurid growth curve is S-shaped, with the maximum growth rate of individuals around 14 years of age.[49]

The smallest known Tyrannosaurus rex individual (LACM 28471, the "Jordan theropod") is estimated to have weighed only 29.9 kilograms (66 lb) at only 2 years old, while the largest, such as FMNH PR2081 ("Sue"), most likely weighed about 5,654 kg (12,465 lb), estimated to have been 28 years old, an age which may have been close to the maximum for the species.[49] T. rex juveniles remained under 1,800 kg (4,000 lb) until approximately 14 years of age, when body size began to increase dramatically. During this rapid growth phase, a young T. rex would gain an average of 600 kg (1,300 lb) a year for the next four years. This slowed after 16 years, and at 18 years of age, the curve plateaus again, indicating that growth slowed dramatically.[50] For example, only 600 kg (1,300 lb) separated the 28-year-old "Sue" from a 22-year-old Canadian specimen (RTMP 81.12.1).[49] This sudden change in growth rate may indicate physical maturity, a hypothesis that is supported by the discovery of medullary tissue in the femur of an 18-year-old T. rex from Montana (MOR 1125, also known as "B-rex").[51] Medullary tissue is found only in female birds during ovulation, indicating that "B-rex" was of reproductive age.[52]
Other tyrannosaurids exhibit extremely similar growth curves, although with lower growth rates corresponding to their lower adult sizes.[53] Compared to albertosaurines, Daspletosaurus showed a faster growth rate during the rapid growth period due to its higher adult weight. The maximum growth rate in Daspletosaurus was 180 kilograms (400 lb) per year, based on a mass estimate of 1,800 kg (4,000 lb) in adults. Other authors have suggested higher adult weights for Daspletosaurus; this would change the magnitude of the growth rate, but not the overall pattern.[49] The youngest known Albertosaurus is a two-year-old discovered in the Dry Island bonebed, which would have weighed about 50 kg (110 lb) and measured slightly more than 2 metres (6.6 ft) in length. The 10-metre (33 ft) specimen from the same quarry is the oldest and largest known, at 28 years of age. The fastest growth rate is estimated to occur around 12–16 years of age, reaching 122 kg (269 lb) per year, based on a 1,300 kg (2,900 lb) adult, which is about a fifth of the rate for T.-rex. For Gorgosaurus, the calculated maximum growth rate is about 110 kilograms (240 lb) during the rapid growth phase, which is comparable to that of Albertosaurus.[49]
The discovery of an embryonic tyrannosaur of an as-yet-unknown genus suggests that tyrannosaurids developed their distinctive skeletal features while developing in the egg. Furthermore, the size of the specimen, a 1.1 in (2.8 cm) dentary from the lower jaw found in the Two Medicine Formation of Montana in 1983 and a foot claw found in the Horseshoe Canyon Formation in 2018 and described in 2020, suggests that neonate tyrannosaurids were born with skulls the size of a mouse or similarly sized rodents and may have been roughly the size of a small dog at birth. The jaw specimen is believed to have come from an animal roughly 2.5 ft (0.76 m) while the claw is believed to belong to a specimen measuring around 3 ft (0.91 m). While eggshells have not been found in association with either specimen, the location where these neonate tyrannosaurids were uncovered suggests these animals were using the same nest sites as other species they lived with and preyed upon.[54] The lack of eggshells associated with these specimens has also opened up speculation to the possibility that tyrannosaurids laid soft-shelled eggs as the genera Mussaurus and Protoceratops are believed to have done.[55]
Fossil footprints from the Wapiti Formation suggest that as tyrannosaurids grew, the feet became wider with thicker toes to support their weight. The broader feet suggest that adult tyrannosaurids were slower-moving than their offspring.[56][57]
Life history
[edit]
The end of the rapid growth phase suggests the onset of sexual maturity in Albertosaurus, although growth continued at a slower rate throughout the animals' lives.[49][53] Sexual maturation while still actively growing appears to be a shared trait among small[58] and large[59] dinosaurs as well as in large mammals, such as humans and elephants.[59] This pattern of relatively early sexual maturation differs strikingly from the pattern in birds, which delay their sexual maturity until after they have finished growing.[59][60]
By tabulating the number of specimens of each age group, Erickson and his colleagues were able to draw conclusions about life history in tyranosauridae populations. Their analysis showed that while juveniles were rare in the fossil record, subadults in the rapid growth phase and adults were far more common. Over half of the known T. rex specimens appear to have died within six years of reaching sexual maturity, a pattern that is also seen in other tyrannosaurs and in some large, long-lived birds and mammals today. These species are characterized by high infant mortality rates, followed by relatively low mortality among juveniles. Mortality increases again following sexual maturity, partly due to the stresses of reproduction. While this could be due to preservation or collection biases, Erickson hypothesized that the difference was due to low mortality among juveniles over a certain size, which is also seen in some modern large mammals, like elephants. This low mortality may have resulted from a lack of predation, since tyrannosaurs surpassed all contemporaneous predators in size by the age of two. Paleontologists have not found enough Daspletosaurus remains for a similar analysis, but Erickson notes that the same general trend seems to apply.[53]
The tyrannosaurids spent as much as half its life in the juvenile phase before ballooning up to near-maximum size in only a few years.[49] This, along with the complete lack of predators intermediate in size between huge adult tyrannosaurids and other small theropods, suggests these niches may have been filled by juvenile tyrannosaurids. This is seen in modern Komodo dragons, where hatchlings start off as tree-dwelling insectivores and slowly mature into massive apex predators capable of taking down large vertebrates.[10] For example, Albertosaurus have been found in aggregations that some have suggested to represent mixed-age packs.[61][62]
Locomotion
[edit]Locomotion abilities are best studied for Tyrannosaurus, and there are two main issues concerning this: how well it could turn; and what its maximum straight-line speed was likely to have been. Tyrannosaurus may have been slow to turn, possibly taking one to two seconds to turn only 45° – an amount that humans, being vertically oriented and tail-less, can spin in a fraction of a second.[63] The cause of the difficulty is rotational inertia, since much of Tyrannosaurus's mass was some distance from its center of gravity, like a human carrying a heavy timber.[64]
Scientists have produced a wide range of maximum speed estimates, mostly around 11 metres per second (25 mph), but a few as low as 5–11 metres per second (11–25 mph), and a few as high as 20 metres per second (45 mph). Researchers have to rely on various estimating techniques because, while there are many tracks of very large theropods walking, so far none have been found of very large theropods running—and this absence may indicate that they did not run.[65]

Jack Horner and Don Lessem argued in 1993 that Tyrannosaurus was slow and probably could not run (no airborne phase in mid-stride).[66] However, Holtz (1998) concluded that tyrannosaurids and their close relatives were the fastest large theropods.[67] Christiansen (1998) estimated that the leg bones of Tyrannosaurus were not significantly stronger than those of elephants, which are relatively limited in their top speed and never actually run (there is no airborne phase), and hence proposed that the dinosaur's maximum speed would have been about 11 metres per second (25 mph), which is about the speed of a human sprinter.[68] Farlow and colleagues (1995) have argued that a 6- to 8-ton Tyrannosaurus would have been critically or even fatally injured if it had fallen while moving quickly, since its torso would have slammed into the ground at a deceleration of 6 g (six times the acceleration due to gravity, or about 60 metres/s2) and its tiny arms could not have reduced the impact.[69][70] However, giraffes have been known to gallop at 50 km/h (31 mph), despite the risk that they might break a leg or worse, which can be fatal even in a "safe" environment such as a zoo.[71][72] Thus it is quite possible that Tyrannosaurus also moved fast when necessary and had to accept such risks; this scenario has been studied for Allosaurus too.[73][74] Most recent research on Tyrannosaurus locomotion does not narrow down speeds further than a range from 17 to 40 km/h (11 to 25 mph), i.e. from walking or slow running to moderate-speed running.[65][75][76] A computer model study in 2007 estimated running speeds, based on data taken directly from fossils, and claimed that T. rex had a top running speed of 8 metres per second (18 mph).[77][78] (Probably a juvenile individual.[79])
Studies by Eric Snively et al., published in 2019 indicate that tyrannosaurids such as Tarbosaurus and Tyrannosaurus itself were more manoeuvrable than allosauroids of comparable size due to low rotational inertia compared to their body mass combined with large leg muscles. As a result, it is hypothesized that tyrannosaurids were capable of making relatively quick turns and could likely pivot their bodies more quickly when close to their prey, or that while turning, they could "pirouette" on a single planted foot while the alternating leg was held out in a suspended swing during pursuit. The results of this study potentially could shed light on how agility could have contributed to the success of tyrannosaurid evolution.[80]
Additionally, a 2020 study indicates that tyrannosaurids were exceptionally efficient walkers. Studies by Dececchi et al., compared the leg proportions, body mass, and the gaits of more than 70 species of theropod dinosaurs including tyrannosaurids. The research team then applied a variety of methods to estimate each dinosaur's top speed when running as well as how much energy each dinosaur expended while moving at more relaxed speeds such as when walking. Among smaller to medium-sized species such as dromaeosaurids, longer legs appear to be an adaptation for faster running, in line with previous results by other researchers. But for theropods weighing over 1,000 kg (2,200 lb), top running speed is limited by body size, so longer legs instead were found to have correlated with low-energy walking. The results of the study further indicated that smaller theropods evolved long legs for speed as a means to both aid in hunting and escape from larger predators while larger predatory theropods that evolved long legs did so to reduce the energy costs and increase foraging efficiency, as they were freed from the demands of predation pressure due to their role as apex predators. Compared to more basal groups of theropods in the study, tyrannosaurids showed a marked increase in foraging efficiency due to reduced energy expenditures during hunting and scavenging. This likely resulted in tyrannosaurs having a reduced need for hunting forays and requiring less food to sustain themselves as a result. Additionally, the research, in conjunction with studies that show tyrannosaurs were more agile than other large-bodied theropods, indicates they were quite well-adapted to a long-distance stalking approach followed by a quick burst of speed to go for the kill. Analogies can be noted between tyrannosaurids and modern wolves as a result, supported by evidence that at least some tyrannosaurids such as Albertosaurus were hunting in group settings.[81][82]
Integument
[edit]
An ongoing debate in the paleontological community surrounds the extent and nature of tyrannosaurid integumentary covering. Long filamentous structures have been preserved along with skeletal remains of numerous coelurosaurs from the Early Cretaceous Yixian Formation and other nearby geological formations from Liaoning, China.[83] These filaments have usually been interpreted as "protofeathers," homologous with the branched feathers found in birds and some non-avian theropods,[84][85] although other hypotheses have been proposed.[86] A skeleton of Dilong was described in 2004 that included the first example of "protofeathers" in a tyrannosauroid. Similarly to down feathers of modern birds, the "protofeathers" found in Dilong were branched but not pennaceous, and may have been used for insulation.[23] The discovery and description of the 9-metre (30 ft) feathered tyrannosauroid Yutyrannus in 2012 indicates the possibility large tyrannosaurids were also feathered as adults.[87]

Based on the principle of phylogenetic bracketing, it was predicted that tyrannosaurids might also possess such feathering. However, a study in 2017 published by a team of researchers in Biology Letters described tyrannosaurid skin impressions collected in Alberta, Montana, and Mongolia, which came from five genera (Tyrannosaurus, Albertosaurus, Gorgosaurus, Daspletosaurus and Tarbosaurus).[88] Although the skin impressions are small, they are widely dispersed across the post-cranium, being collectively located on the abdomen, thoracic region, ilium, pelvis, tail, and neck. They show a tight pattern of fine, non-overlapping pebbly scales (which co-author Scott Persons compared to those seen on the flanks of a crocodile[89]) and preserve no hints of feathering. The basic texture is composed of tiny "basement scales" approximately 1 to 2 mm in diameter, with some impressions showing 7 mm "feature scales" interspersed between them. Additional scales can be seen in tyrannosaurid footprints.[90] Studies find that the facial integument of tyrannosaurids had scales on the dentary and maxilla, cornified epidermis and armor-like skin on the subordinate regions.[91][92]
Bell et al. performed an ancestral character reconstruction based on what is known about integument distribution in tyrannosauroids. Despite an 89% probability that tyrannosauroids started out with feathers, they determined that scaly tyrannosaurids have a 97% probability of being true. The data "provides compelling evidence of an entirely squamous covering in Tyrannosaurus," the team wrote, although they conceded that plumage may have still been present on the dorsal region where skin impressions haven't been found yet. Bell et al. hypothesizes that the scale impressions of tyrannosaurids are possibly reticula which are secondarily derived from feathers though evidence is needed to support this.[88] However, others argue that this is because of taphonomic bias in tyrannosaurids.[93]
It has yet to be determined why such an integumentary change might have occurred. A precedent for feather loss can be seen in other dinosaur groups such as ornithischians, in which filamentous structures were lost, and scales reappeared.[94] Although gigantism has been suggested as a mechanism, Phil R. Bell, who co-authored the study, noted that the feathered Yutyrannus overlapped in size with Gorgosaurus and Albertosaurus. "The problem here is that we have big tyrannosaurs, some with feathers, some without that live in pretty similar climates. So what's the reason for this difference? We really don't know."[95]
Vision
[edit]
Глазницы тираннозавра расположены так, что глаза направлены вперед, что дает им бинокулярное зрение немного лучше, чем у современных ястребов . В то время как хищные тероподы обычно имели бинокулярное зрение прямо перед черепом, у тираннозавров площадь перекрытия была значительно большей. Джек Хорнер также отметил, что линия тираннозавров имеет историю постоянного улучшения бинокулярного зрения. Трудно представить, как естественный отбор благоприятствовал бы этой долгосрочной тенденции, если бы тираннозавры были чистыми падальщиками, которым не требовалось бы развитое восприятие глубины , которое стереоскопическое зрение . обеспечивает [96][97] In modern animals, binocular vision is found mainly in predators (the principal exceptions are primates, which need it for leaping from branch to branch). Unlike Tyrannosaurus, Tarbosaurus had a narrower skull more typical of other tyrannosaurids in which the eyes faced primarily sideways. All of this suggests that Tarbosaurus relied more on its senses of smell and hearing than on its eyesight.[98] In Gorgosaurus specimens, the eye socket was circular rather than oval or keyhole-shaped as in other tyrannosaurid genera.[11] У дасплетозавра это был высокий овал, что-то среднее между круглой формой, наблюдаемой у горгозавра , и формой «замочной скважины» у тираннозавра . [10] [11] [44]
Чувствительность лица
[ редактировать ]На основе сравнения текстуры костей дасплетозавра Томасом с современными крокодилами подробное исследование, проведенное в 2017 году Д. Карром и соавт. были большие плоские чешуйки обнаружили, что у тираннозавров на мордах . [99] [100] В центре этих чешуек находились небольшие ороговевшие участки. У крокодилов такие участки покрывают пучки сенсорных нейронов , способных обнаруживать механические, термические и химические раздражители . [101] [102] Они предположили, что тираннозавры , вероятно, также имели пучки сенсорных нейронов под лицевой чешуей и, возможно, использовали их для идентификации объектов, измерения температуры в своих гнездах и осторожного сбора яиц и вылупившихся детенышей . [99]
Костлявые гребни
[ редактировать ]
Костные гребни встречаются на черепах многих теропод, в том числе многих тираннозаврид. Алиорамус , возможный тираннозаврид из Монголии, имеет один ряд из пяти выступающих костных выступов на носовых костях; аналогичный ряд значительно более низких бугорков имеется на черепе аппалачиозавра , а также некоторых экземпляров дасплетозавра , альбертозавра и тарбозавра . [16] У альбертозавра , горгозавра и дасплетозавра перед каждым глазом на слезной кости имеется выступающий рог. Слезный рог отсутствует у тарбозавра и тираннозавра , у которых вместо этого имеется серповидный гребень позади каждого глаза на заглазничной кости . Эти гребни на голове, возможно, использовались для демонстрации , возможно, для распознавания видов или ухаживания . поведения [10]
Терморегуляция
[ редактировать ]Долгое время считалось, что тираннозавр , как и большинство динозавров, обладает экзотермическим («хладнокровным») рептильным метаболизмом , но ему бросили вызов такие ученые, как Роберт Т. Баккер и Джон Остром , в первые годы « ренессанса динозавров », начиная с позднего периода. 1960-е годы. [103] [104] Утверждалось, что сам тираннозавр рекс был эндотермным («теплокровным»), что подразумевало очень активный образ жизни. [105] пытались определить способность тираннозавра регулировать С тех пор несколько палеонтологов своего тела температуру . Гистологические данные о высоких темпах роста молодых тираннозавров , сравнимых с темпами роста млекопитающих и птиц, могут поддерживать гипотезу о высоком метаболизме. Кривые роста показывают, что, как и у млекопитающих и птиц, рост T. rex был ограничен в основном неполовозрелыми животными, а не неопределенным ростом, наблюдаемым у большинства других позвоночных . [50] могла составлять не более 4–5 ° C (7–9 ° F) голени Было указано, что разница температур между позвонками туловища и большеберцовой костью Уильям Шоуерс утверждали, что этот небольшой температурный диапазон между сердцевиной тела и конечностями . Палеонтолог Риз Баррик и геохимик указывает на то, что тираннозавр поддерживает постоянную внутреннюю температуру тела ( гомеотермию ) и что его метаболизм находится где-то между экзотермическими рептилиями и эндотермными млекопитающими. [106] Позже они обнаружили аналогичные результаты у экземпляров гиганотозавров , которые жили на другом континенте и на десятки миллионов лет раньше. [107] Даже если тираннозавр рекс действительно демонстрирует признаки гомеотермии, это не обязательно означает, что он был эндотермным. Подобную терморегуляцию можно также объяснить гигантотермией , как у некоторых современных морских черепах . [108] [109] [110]
Палеоэкология
[ редактировать ]Сосуществование дасплетозавра и горгозавра
[ редактировать ]
В формации Парка Динозавров Горгозавр жил рядом с более редким видом тираннозавра Дасплетозавра . Это один из немногих примеров сосуществования двух родов тираннозавров. Хищники одинакового размера в современных гильдиях хищников разделены на разные экологические ниши из-за анатомических, поведенческих или географических различий, которые ограничивают конкуренцию. Дифференциация ниш между тираннозавридами Парка динозавров недостаточно изучена. [111] В 1970 году Дейл Рассел выдвинул гипотезу , что более распространённый горгозавр активно охотился на быстроногих гадрозавров , в то время как более редкие и доставляющие больше беспокойства цератопсы и анкилозавры (рогатые и тяжелобронированные динозавры ) были оставлены более тяжело сложённому дасплетозавру . [12] Однако экземпляр дасплетозавра ( OTM 200) из современной формации Ту Медисин в Монтане сохраняет переваренные останки молодого гадрозавра в области кишечника. [112] В отличие от некоторых других групп динозавров, ни один из этих родов не был более распространен на возвышенностях или понижениях, чем другой. [111] Однако горгозавр чаще встречается в северных формациях, таких как Парк динозавров, а виды дасплетозавра более распространены на юге. Та же картина наблюдается и у других групп динозавров. Цератопсы-хасмозавры и гадрозаврины-гадрозаврины также более распространены в формации Ту Медисин в Монтане и на юго-западе Северной Америки в кампанском периоде, в то время как центрозаврины и ламбеозаврины доминируют в северных широтах. Хольц предположил, что эта закономерность указывает на общие экологические предпочтения тираннозавров, хасмозавринов и гадрозавринов. В конце поздней маастрихтской стадии тираннозаврины, такие как Tyrannosaurus rex , гадрозаврины, такие как эдмонтозавр , и хасмозаврины, такие как трицератопс, были широко распространены по всей западной части Северной Америки, в то время как альбертозаврины и центрозаврины вымерли, а ламбеозаврины были редки. [10]
Социальное поведение
[ редактировать ]Имеются ограниченные свидетельства социального поведения тираннозаврид. Исследователи сообщили, что полувзрослый и молодой скелеты были найдены в том же карьере, что и экземпляр «Сью», который использовался для подтверждения гипотезы о том, что тираннозавры могли жить в каких-то социальных группах. [113] Хотя нет никаких свидетельств стадного поведения горгозавра . [61] [62] есть свидетельства некоторого поведения стаи у Альбертозавра и Дасплетозавра .
У молодого экземпляра вида Daspletosaurus из Парка динозавров ( TMP 94.143.1) на лице видны следы укусов, нанесенных другим тираннозавром. Следы укусов зажили, что указывает на то, что животное пережило укус. У взрослого дасплетозавра из парка динозавров (TMP 85.62.1) также наблюдаются следы укусов тираннозавра, что показывает, что нападения на лицо не ограничивались молодыми животными. Хотя возможно, что укусы были приписаны другим видам, внутривидовая агрессия, в том числе укусы лица, очень распространена среди хищников. Укусы лица наблюдаются у других тираннозавров, таких как Gorgosaurus и Tyrannosaurus , а также у других родов теропод, таких как Sinraptor и Saurornitholestes . Даррен Танке и Фил Карри предполагают, что укусы происходят из-за внутривидовой конкуренции за территорию или ресурсы или за доминирование внутри социальной группы. [61]

Доказательства того, что дасплетозавр жил социальными группами, получены из костного ложа, найденного в формации Ту-Медицина в Монтане. Костное ложе включает останки трех дасплетозавров , в том числе крупного взрослого человека, маленького подростка и еще одной особи среднего размера. В том же месте сохранилось как минимум пять гадрозавров. Геологические данные указывают на то, что останки не были собраны вместе речными течениями , а все животные были похоронены одновременно в одном месте. Останки гадрозавров разбросаны и имеют множество следов от зубов тираннозавров, что указывает на то, что дасплетозавр питался гадрозаврами во время смерти. Причина смерти неизвестна. Карри предполагает, что дасплетозавры образовывали стаю , хотя утверждать это с уверенностью нельзя. [62] Другие ученые скептически относятся к свидетельствам существования социальных групп у дасплетозавра и других крупных теропод; [114] Брайан Роуч и Дэниел Бринкман предположили, что социальное взаимодействие дасплетозавра больше напоминало бы современного дракона Комодо , где отказывающиеся от сотрудничества особи нападают на трупы, часто нападая и даже поедая друг друга в процессе. [115]
Костное ложе Сухого острова, обнаруженное Барнумом Брауном и его командой, содержит останки 22 альбертозавров , самого большого количества особей, обнаруженных в одном местонахождении среди всех теропод мелового периода, и второго по величине среди всех крупных динозавров-тероподов после группы аллозавров в динозавре Кливленда-Ллойда. Карьер в штате Юта . Группа, кажется, состоит из одного очень пожилого человека; восемь взрослых в возрасте от 17 до 23 лет; семь подростков, находящихся в фазе быстрого роста в возрасте от 12 до 16 лет; и шесть подростков в возрасте от 2 до 11 лет, которые еще не достигли фазы роста. [53] Практически полное отсутствие останков травоядных животных и одинаковая степень сохранности многих особей в карьере, где находился костяной пласт альбертозавра, привели Фила Карри к выводу, что это место не является ловушкой для хищников, как смоляные ямы Ла-Бреа в Калифорнии , и что все сохранившиеся останки травоядных животных животные погибли одновременно. Карри утверждает, что это свидетельство поведения стаи. [116] Другие ученые настроены скептически, отмечая, что животные могли быть согнаны вместе из-за засухи, наводнения или по другим причинам. [53] [114] [117]
Хотя в целом это остается спорным, существуют доказательства, подтверждающие теорию о том, что по крайней мере некоторые тираннозавриды были социальными. В в Британской Колумбии местный формации Вапити Ричард МакКри и др аутфиттер по имени Аарон Фредлунд обнаружил следы, состоящие из следов трех отдельных тираннозаврид (названных ихногеном Bellatoripes fredlundi), и описал их в журнале PLOS One . . Исследование тропы не обнаружило никаких свидетельств того, что одна тропа осталась после того, как была проложена другая, что еще раз подтверждает гипотезу о том, что три отдельных тираннозавра путешествовали вместе как группа. Дальнейшие исследования показали, что животные двигались со скоростью от 3,9 до 5,2 миль в час (от 6,3 до 8,4 км/ч), а высота бедер, вероятно, составляла от 7 до 9 футов. Поскольку из формации известны три разных рода тираннозаврид ( Горгозавр , Дасплетозавр и Альбертозавр соответственно), неизвестно, какой род был создателем тропы. [118] [119] [120] Дополнительные свидетельства в виде костного пласта из карьера Радуги и Единорогов в формации Кайпаровитс в Южной Юте, описанного в 2021 году и приписываемого Тератофонею, позволяют предположить, что другие тираннозавриды также были социальными животными. Окаменелости, состоящие из четырех или, возможно, пяти различных животных в возрасте от 4 до 22 лет, предполагают массовую смертность, возможно, вызванную наводнением или, что менее вероятно, цианобактериальным токсикозом , пожаром или засухой. Тот факт, что все сохранившиеся животные, похоже, погибли в течение короткого промежутка времени, еще больше усиливает аргумент в пользу стадного поведения тираннозаврид, поскольку костные пласты таких родов, как Teratophoneus , Albertosaurus , Tyrannosaurus и Daspletosaurus, демонстрируют предполагаемое социальное поведение, возможно, широко распространенное среди тираннозавриды вообще. [121] [122] [123]
Кормление
[ редактировать ]Следы зубов тираннозавров — наиболее часто сохранившиеся следы питания хищных динозавров. [124] О них сообщалось от цератопсов , гадрозавров и других тираннозавров. [124] Кости тираннозаврид со следами зубов составляют около 2% известных окаменелостей с сохранившимися следами зубов. [124] Зубы тираннозаврид использовались в качестве опор для отрыва мяса от тела, а не в качестве режущих функций, подобных ножу . [125] Характер износа зубов намекает на то, что сложное покачивание головой могло быть связано с кормлением тираннозавров. [125]

Предположения о охотничьих привычках альбертозавра были высказаны несколькими исследователями, которые предполагают, что более молодые члены стаи, возможно, были ответственны за то, что подгоняли свою добычу к взрослым особям, которые были крупнее и мощнее, но также медленнее. [116] Молодые особи, возможно, также вели образ жизни, отличный от взрослых, заполняя ниши хищников между огромными взрослыми особями и более мелкими современными тероподами, самые крупные из которых были на два порядка меньше, чем взрослый альбертозавр по массе. [10] Однако, поскольку сохранение поведения в летописи окаменелостей встречается крайне редко, эти идеи нелегко проверить. Фил Карри предполагает, что дасплетозавры формировали стаи для охоты , хотя это нельзя утверждать с уверенностью. [62] Нет никаких свидетельств такого стадного поведения у горгозавра . [61] [62]
Споры о том, ли Тираннозавр был хищником или чистым падальщиком, так же стары, как и споры о его передвижении. Ламбе (1917) описал хороший скелет горгозавра тираннозавра близкого родственника и что он, а следовательно, и тираннозавр, были чистыми падальщиками, поскольку зубы горгозавра пришел к выводу , почти не изнашивались. [126] Этот аргумент уже не воспринимается всерьез, поскольку у теропод довольно быстро сменились зубы. С момента первого открытия тираннозавра большинство ученых согласились с тем, что это был хищник, хотя, как и современные крупные хищники, он был бы рад добыть или украсть добычу другого хищника, если бы у него была такая возможность. [127] [128]
Известный по гадрозаврам эксперт Джек Хорнер в настоящее время является основным сторонником идеи о том, что тираннозавр был исключительно падальщиком и вообще не занимался активной охотой. [66] [129] [130] Хорнер представил несколько аргументов в поддержку гипотезы чистого мусорщика. Наличие крупных обонятельных луковиц и обонятельных нервов предполагает высокоразвитое обоняние, позволяющее вынюхивать трупы на больших расстояниях. Зубы могли раздробить кость и, следовательно, извлечь как можно больше пищи ( костного мозга ) из остатков туши, обычно наименее питательных частей. По крайней мере, некоторые из его потенциальных жертв могли быстро передвигаться, хотя данные свидетельствуют о том, что Тираннозавр шел, а не бежал. [129] [131]
Другие данные свидетельствуют о охотничьем поведении тираннозавра . Глазницы тираннозавров расположены так, что глаза направлены вперед, что дает им бинокулярное зрение немного лучше, чем у современных ястребов . Повреждения, нанесенные тираннозаврами, были обнаружены на скелетах гадрозавров и трицератопсов, которые, по-видимому, пережили первоначальные нападения. [132] [133] [134] Некоторые исследователи утверждают, что если бы тираннозавр был падальщиком, то главным хищником верхнего мела Америки должен был быть другой динозавр. Главной добычей были более крупные маргиноцефалы и орнитоподы . Другие тираннозавриды имеют настолько много общих характеристик с тираннозавром , что только небольшие дромеозавры остаются возможными высшими хищниками. В этом свете сторонники гипотезы падальщиков предположили, что размер и сила тираннозавров позволяли им воровать добычу у более мелких хищников. [131]

было зарегистрировано появление молодого горгозавра с содержимым желудка in situ , содержащим двух неповрежденных молодых особей Citipes возрастом около года В 2023 году в формации Парк динозавров . На момент смерти этому подростку было 5-7 лет, его длина составляла около 4 метров (13 футов), а вес - около 335 килограммов (739 фунтов). Он намного крупнее двух молодых особей Citipes , которые весят около 9–12 кг (20–26 фунтов), что противоречит предположению, что тираннозавриды питались добычей своего размера, когда они достигли веса 16–32 кг (35–71 фунт), что указывает на что молодые тираннозавриды все еще потребляли гораздо меньшую добычу после превышения определенного порога размера. Обнаружение этого экземпляра показало, что тираннозавриды, вероятно, не охотились стаями, состоящими из нескольких поколений, поскольку размер их добычи слишком мал, чтобы их можно было разделить с сородичами. Это также прямое диетическое свидетельство, которое подкрепляет теорию «онтогенетического сдвига в рационе» тираннозаврид, ранее выведенную с помощью экологического моделирования и анатомических особенностей среди разных возрастных групп. Лишь остатки задних конечностей и хвостовых позвонков молоди. Цитипы присутствовали в полости желудка тираннозаврид, что позволяет предположить, что молодой горгозавр , возможно, преимущественно потреблял мускулистые задние конечности. [135]
Каннибализм
[ редактировать ]Имеющиеся данные также убедительно свидетельствуют о том, что тираннозавриды, по крайней мере, иногда были каннибалами. У самого тираннозавра есть веские доказательства, указывающие на то, что он был каннибалом, по крайней мере, в плане способности питаться мусором, о чем свидетельствуют следы зубов на костях стопы, плечевой кости и плюсневых костях одного экземпляра. [136] Окаменелости из формации Фрутленд , формации Киртланд (обе по возрасту кампана) и формации Оджо Аламо маастихтского возраста позволяют предположить, что каннибализм присутствовал в различных родах тираннозаврид в бассейне Сан-Хуан. Данные, собранные на основе образцов, позволяют предположить оппортунистическое пищевое поведение тираннозаврид, которые поедали представителей своего собственного вида. [137]
Распределение
[ редактировать ]
Самые ранние известные останки тиранозавра произошли между 129,4 и 125 миллионами лет назад в Исикаве, Япония, около 36,166668 широты и 136,633331 широты. Найденные Маршем в 1881 г. и идентифицированные Г.Ф. Осборном в 1906 г., возраст останков был определен Х. Мацуокой и др. в 2002 году.
В то время как более ранние тираннозавроиды встречаются на всех трех северных континентах, окаменелости тираннозаврид известны только из Северной Америки и Азии. Иногда фрагментарные останки, обнаруженные в Южном полушарии, называют «тираннозавридами Южного полушария», хотя, похоже, это были ошибочно идентифицированные окаменелости абелизаврид. [138]
Останки тираннозаврид никогда не были обнаружены в восточной части Северной Америки, в то время как более базальные тираннозавриды, такие как дриптозавр и аппалачиозавр , сохранялись там до конца мелового периода, что указывает на то, что тираннозавриды, должно быть, эволюционировали или расселились по западной части Северной Америки после того, как континент был разделен пополам. у Западного внутреннего морского пути в середине мелового периода. [16] Окаменелости тираннозаврид были найдены на Аляске , что, возможно, послужило путем распространения между Северной Америкой и Азией. [139] В одном кладистическом анализе обнаружено, что Алиорамус и Тарбозавр связаны между собой, образуя уникальную азиатскую ветвь семейства. [18] Позже это было опровергнуто открытием Цяньчжоузавра и описанием семейства тираннозавров Алиорамини. Зубы тираннозаврид крупного вида неизвестной разновидности были обнаружены на полуострове Нагасаки исследователями из Музея динозавров префектуры Фукуи, что еще больше расширило ареал группы. Возраст зубов оценивается в 81 миллион лет ( кампанский век). [140]

Из двух подсемейств тираннозавры, по-видимому, были более распространены. Альбертозаврины неизвестны в Азии, где обитали тираннозавры, такие как Тарбозавр и Чжученгтираннус , а также Цяньчжоузавр и Алиорам из Алиорамини . Подсемейства как Tyrannosaurinae, так и Albertosaurinae присутствовали на кампанском и раннем маастрихтском этапах Северной Америки, при этом тираннозавры, такие как Daspletosaurus, обитали по всей западной части континента, тогда как альбертозаврины Albertosaurus и Gorgosaurus в настоящее время известны только из северо-западной части континента. [141]
К концу маастрихта альбертозаврины, похоже, вымерли, а тираннозавр тираннозавр кочевал из Саскачевана в Техас . Эта закономерность отражена и у других таксонов динозавров Северной Америки. В кампане и раннем маастрихте ламбеозаврины- гадрозавры и центрозаврины- цератопсы были распространены на северо-западе, тогда как гадрозаврины и хасмозаврины были более распространены на юге. К концу мелового периода центрозаврины неизвестны, а ламбеозаврины редки, тогда как гадрозаврины и хасмозаврины были распространены по всей территории Западных внутренних районов. [10] Исследование, опубликованное в журнале Scientific Reports 2 февраля 2016 года Стивом Брусаттом, Томасом Карром и др . указывает на то, что в позднем маастрихте сам тираннозавр мог быть частично ответственен за вымирание других тираннозавридов на большей части западной части Северной Америки. Исследование показывает, что тираннозавр мог быть иммигрантом из Азии, а не эволюционировать в Северной Америке (возможно, потомком близкородственного тарбозавра ), который вытеснил и вытеснил других тираннозаврид. Эта теория дополнительно подтверждается тем фактом, что в пределах известного ареала тираннозавров практически не встречается других типов тираннозаврид . [142]
Хронология родов
[ редактировать ]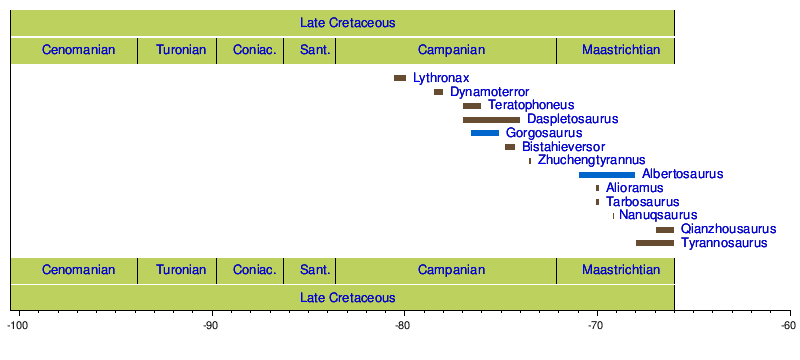
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хольц, Томас Р. младший (2012) Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, зима 2011 г., Приложение. Архивировано 12 августа 2017 г. в Wayback Machine.
- ^ Статья о Скотти на Osel.cz (на чешском языке)
- ^ Лица, SW; Карри, Пи Джей; Эриксон, генеральный директор (2019). «Старый и исключительно крупный взрослый экземпляр тираннозавра рекса » . Анатомическая запись . 303 (4): 656–672. дои : 10.1002/ar.24118 . ISSN 1932-8486 . ПМИД 30897281 .
- ^ Лайл, А. (22 марта 2019 г.). «Палеонтологи идентифицируют самого большого из когда-либо обнаруженных тираннозавров » . Фолио, Университет Альберты . Проверено 25 марта 2019 г.
- ^ Перейти обратно: а б с д и Карпентер, Кен. (1992). «Тираннозавриды (Динозаврии) Азии и Северной Америки». В Матее, Найл Дж.; Чэнь Пейджи (ред.). Аспекты неморской меловой геологии . Пекин: China Ocean Press. стр. 250–268.
- ^ Брайтаупт, Британская Колумбия; Саутвелл, штат Эйха; Мэтьюз, Северная Каролина (18 октября 2005 г.). «В честь 100-летия тираннозавра рекса : Manospondylus gigas , Ornithomimus grandis и Dynamosaurus imperiosus , самых ранних открытий тираннозавра рекса на Западе» . Рефераты с программами . 2005 Ежегодное собрание в Солт-Лейк-Сити . Том. 37. Геологическое общество Америки . п. 406. Архивировано из оригинала 22 октября 2019 года . Проверено 8 октября 2008 г.
- ^ Брайтауп, Британская Колумбия; Саутвелл EH; Мэтьюз Н.А. (2006). « Dynamosaurus imperiosus и самые ранние открытия тираннозавра рекса в Вайоминге и на Западе». Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 257–258.
- ^ Баккер РТ; Уильямс М.; Карри П. (1988). « Nanotyrannus , новый род карликовых тираннозавров из позднего мелового периода Монтаны». Хантерия . 1 :1–30.
- ^ Перейти обратно: а б Курзанов Сергей М. «Новый карнозавр из позднего мела Ногон-Цава, Монголия». Труды совместной советско-монгольской палеонтологической экспедиции . 3 : 93–104.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р Хольц, Томас Р. (2004). «Тираннозавроида». В Вейшампеле, Дэвид Б .; Додсон, Питер ; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 111–136. ISBN 978-0-520-24209-8 .
- ^ Перейти обратно: а б с д и ж г Карри, Филип Дж. (2003). «Краниальная анатомия тираннозаврид позднего мела Альберты» (PDF) . Acta Palaeontologica Polonica . 48 (2): 191–226. Архивировано (PDF) из оригинала 19 августа 2018 г. Проверено 12 июня 2010 г.
- ^ Перейти обратно: а б с д Рассел, Дейл А. (1970). «Тираннозавры из позднего мела западной Канады». Публикации Национального музея естественных наук по палеонтологии . 1 :1–34.
- ^ Мортимер, Микки (21 июля 2003 г.). «И самый большой теропод...» Список рассылки по динозаврам. Архивировано из оригинала 25 марта 2010 года . Проверено 8 сентября 2007 г.
- ^ Статья о Скотти на Osel.cz (на чешском языке)
- ^ Карри, Филип Дж. (2000). «Тероподы мела Монголии». Эпоха динозавров в России и Монголии . Кембридж: Издательство Кембриджского университета. стр. 434–455. ISBN 978-0-521-54582-2 .
- ^ Перейти обратно: а б с д и ж Карр, Томас Д.; Уильямсон, Томас Э.; Швиммер, Дэвид Р. (2005). «Новый род и вид тираннозавроидов из позднемеловой (средний кампан) формации Демополис в Алабаме». Журнал палеонтологии позвоночных . 25 (1): 119–143. doi : 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2 . ISSN 0272-4634 . S2CID 86243316 .
- ^ Карр, Томас Д.; Варриккио, Дэвид Дж.; Седлмайр, Джейк К.; Робертс, Эрик М.; Мур, Джейсон Р. (30 марта 2017 г.). «Новый тираннозавр с доказательствами анагенеза и сенсорной системой лица, подобной крокодиловой» . Научные отчеты . 7 (1): 44942. Бибкод : 2017NatSR...744942C . дои : 10.1038/srep44942 . ISSN 2045-2322 . ПМЦ 5372470 . ПМИД 28358353 .
- ^ Перейти обратно: а б с д Карри, Филип Дж .; Хурум, Йорн Х; Сабат, Кароль. (2003). «Строение черепа и эволюция в филогении тираннозаврид» (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234. Архивировано (PDF) из оригинала 31 октября 2008 г. Проверено 2 октября 2010 г.
- ^ Перейти обратно: а б с д и ж г Аблер, В.Л. 2001. Модель зубцов тираннозавра, сделанная с помощью пропила и сверла. п. 84–89. В: Жизнь мезозойских позвоночных . Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Индианского университета.
- ^ Куинлан, Элизабет Д.; Дерстлер, Крейг; Миллер, Мерседес М. (2007). «Анатомия и функция III пальца тираннозавра рекса » . Ежегодное собрание Геологического общества Америки — Тезисы с программами : 77. Архивировано из оригинала 24 февраля 2008 г. Проверено 15 января 2008 г. [только аннотация]
- ^ Снайвли, Эрик; О'Брайен, Хейли; Хендерсон, Дональд М.; Мэллисон, Генрих; Сюрринг, Лара А.; Бернс, Майкл Э.; Хольц, Томас Р.; Рассел, Энтони П.; Уитмер, Лоуренс М.; Карри, Филип Дж.; Хартман, Скотт А.; Коттон, Джон Р. (21 февраля 2019 г.). «Меньшая вращательная инерция и большие мышцы ног указывают на более быстрые повороты у тираннозаврид, чем у других крупных теропод» . ПерДж . 7 : е6432. дои : 10.7717/peerj.6432 . ISSN 2167-8359 . ПМК 6387760 . ПМИД 30809441 .
- ^ Хартман, Скотт (21 февраля 2019 г.). «Меньшая вращательная инерция и большие мышцы ног указывают на более быстрые повороты у тираннозаврид, чем у других крупных теропод» . Скотт Хартман — доктор философии . Проверено 29 декабря 2023 г.
- ^ Перейти обратно: а б с Сюй Син, X ; Норелл, Марк А.; Куанг Сюэнь; Ван Сяолинь; Чжао Ци; Цзя Чэнкай. (2004). «Базальные тираннозавроиды из Китая и свидетельства существования протоперьев у тираннозавроидов» (PDF ) Природа 431 (7009): 680–684. Бибкод : 2004Nature.431..680X . дои : 10.1038/nature02855 . ПМИД 15470426 . S2CID 4381777 .
- ^ Хольц, Томас Р. (1994). «Филогенетическое положение тираннозаврид: значение для систематики теропод». Журнал палеонтологии . 68 (5): 1100–1117. Бибкод : 1994JPal...68.1100H . дои : 10.1017/S0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Томас М. Каллен, Дерек Уильям Ларсон, Марк П. Уиттон, Дайан Скотт, Теа Махо, Кирстин С. Бринк, Дэвид С. Эванс, Роберт Рейс (30 марта 2023 г.). «Реконструкция лица динозавра-теропода и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Бибкод : 2023Sci...379.1348C . дои : 10.1126/science.abo7877 . ПМИД 36996202 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Коуп ЭД (1866). «Открытие гигантского динозавра в меловом периоде Нью-Джерси». Труды Академии естественных наук Филадельфии . 18 : 275–279.
- ^ Малеев Е.А. (1955). «Новые хищные динозавры из верхнемеловой Монголии». Доклады Академии наук СССР . 104 (5): 779–783.
- ^ Лейди, Джозеф (1856). «Уведомление об останках вымерших рептилий и рыб, обнаруженных доктором Ф. В. Хайденом в бесплодных землях реки Джудит, территория Небраски». Труды Академии естественных наук Филадельфии . 8 : 72–73.
- ^ Осборн, Генри Ф. (1905). « Тираннозавр и другие хищные динозавры мелового периода». Бюллетень Американского музея естественной истории . 21 (3): 259–265. дои : 10.1111/j.1468-5965.2007.00735_17.x . hdl : 2246/1464 .
- ^ Лидделл, Генри Г .; Скотт, Роберт (1980). Греко-английский лексикон (сокращенное изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-910207-5 .
- ^ Фиорилло, Арканзас ; Тыкоски, Р.С. (2014). Додсон, Питер (ред.). «Маленький новый тираннозавр с вершины мира» . ПЛОС ОДИН . 9 (3): e91287. Бибкод : 2014PLoSO...991287F . дои : 10.1371/journal.pone.0091287 . ПМЦ 3951350 . ПМИД 24621577 .
- ^ Пол, Грегори С. (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 464стр . ISBN 978-0-671-61946-6 .
- ^ Лю, Цзюньчан; Йи, Лайпин; Брусатте, Стивен Л.; Ян, Линг; Ли, Хуа; Чен, Лю (2014). «Новая клада азиатских длиннорылых тираннозаврид позднего мела» . Природные коммуникации . 5 : 3788. Бибкод : 2014NatCo...5.3788L . дои : 10.1038/ncomms4788 . ПМИД 24807588 .
- ^ «В Азии обнаружен длиннорылый тираннозавр Пиноккио Рекс» . 7 мая 2014 г. Архивировано из оригинала 2 февраля 2019 г. . Проверено 8 мая 2014 г.
- ^ Ольшевский, Георгий (1995). «Происхождение и эволюция тираннозаврид». Кёрюгаку Сайзенсен [Dino Frontline] . 9–10 : 92–119.
- ^ Ольшевский Г.; Форд ТЛ (1995). «Происхождение и эволюция тираннозаврид, часть 2 [на японском языке]». Дино Фронтлайн . 6 : 75–99.
- ^ Фаулер, Д.В.; Вудворд, Х.Н.; Фридман, Э.А.; Ларсон, Польша; Хорнер, младший (2011). «Повторный анализ «Raptorex kriegsteini»: молодого динозавра-тираннозавра из Монголии» . ПЛОС ОДИН . 6 (6): e21376. Бибкод : 2011PLoSO...621376F . дои : 10.1371/journal.pone.0021376 . ПМК 3126816 . ПМИД 21738646 .
- ^ Карр, Томас Д. (25 ноября 2022 г.). «Переоценка окаменелостей тираннозавроидов из формации Ирен-Дабасу (коньяк-кампан), Внутренняя Монголия, Китайская Народная Республика» . Журнал палеонтологии позвоночных . 42 (5). Бибкод : 2022JVPal..42E9817C . дои : 10.1080/02724634.2023.2199817 . ISSN 0272-4634 .
- ^ Серено, Пол К. (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Новогодний ежегодник по геологии и палеонтологии, трактаты (на немецком языке). 210 : 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Перейти обратно: а б с д и ж г час Хольц, Т.Р.-младший (2001), Филогения и таксономия тираннозаврид в книге К. Карпентера и Д. Танке [ред.], Жизнь мезозойских позвоночных. Университет Индианы. Пресс, стр. 64-83.
- ^ Брошу, Кристофер Р. (2003). «Остеология тираннозавра рекса : данные по почти полному скелету и компьютерному томографическому анализу черепа с высоким разрешением». Мемуары Общества палеонтологии позвоночных . 7 : 1–138. дои : 10.2307/3889334 . JSTOR 3889334 .
- ^ Серено, Пол К. (7 ноября 2005 г.). «Стеблевая архозаврия — TaxonSearch» . Архивировано из оригинала 19 февраля 2007 года . Проверено 14 января 2008 г.
- ^ Хурум, Йорн Х.; Сабат, Кароль. (2003). «Гигантские динозавры-тероподы из Азии и Северной Америки: сравнение черепов Tarbosaurus bataar и Tyrannosaurus rex » (аннотация) . Acta Palaeontologica Polonica . 48 (2): 161–190. Архивировано из оригинала 8 марта 2012 г. Проверено 10 сентября 2008 г.
- ^ Перейти обратно: а б Карр, Томас Д. (1999). «Черепно-лицевой онтогенез тираннозаврид (Dinosauria, Coelurosauria)» . Журнал палеонтологии позвоночных . 19 (3): 497–520. Бибкод : 1999JVPal..19..497C . дои : 10.1080/02724634.1999.10011161 . S2CID 83744433 . Архивировано из оригинала 20 февраля 2020 г. Проверено 2 декабря 2019 г.
- ^ «Недавно найденный динозавр — длинноносый родственник тираннозавра рекса» . ScienceDaily . Архивировано из оригинала 17 марта 2018 г. Проверено 9 марта 2018 г.
- ^ CITEREFBrusatte_&_Carr2016
- ^ CITEREFFiorillo_&_Tykoski2014
- ^ Лоуэн, Массачусетс ; Ирмис, РБ ; Сертич, JJW ; Карри, Пи Джей ; Сэмпсон, SD (2013). Эванс, Дэвид С. (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение позднемеловых океанов» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- ^ Перейти обратно: а б с д и ж г час Эриксон, Грегори М., генеральный директор; Маковицкий, Питер Дж.; Карри, Филип Дж .; Норелл, Марк А.; Йерби, Скотт А.; Брошу, Кристофер А. (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 .
Выборочная продолжительность жизни тираннозавра колебалась от 2 до 28 лет
(ошибка: дои : 10.1038/nature16487 , PMID 26675726 , Часы втягивания ) и соответствующие оценки массы тела варьировались от 29,9 до 5654 кг. - ^ Перейти обратно: а б Хорнер, младший и Падиан, К. (2004). «Возраст и динамика роста тираннозавра рекса» . Труды Лондонского королевского общества Б. 271 (1551): 1875–1880. дои : 10.1098/rspb.2004.2829 . ПМК 1691809 . ПМИД 15347508 .
- ^ Ли, Эндрю Х.; Вернинг, Сара (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . Труды Национальной академии наук . 105 (2): 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ПМК 2206579 . ПМИД 18195356 .
- ^ Швейцер, М. Х., Виттмайер, Дж. Л., и Хорнер, младший (2005). «Полоспецифическая репродуктивная ткань бескилевых и тираннозавра рекса ». Наука . 308 (5727): 1456–1460. Бибкод : 2005Sci...308.1456S . дои : 10.1126/science.1112158 . ПМИД 15933198 . S2CID 30264554 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с д и Эриксон, ГМ, Карри, П.Дж., Иноуе, Б.Д., и Винн, А.А. (2006). «Таблицы жизни тираннозавров: пример биологии популяции нептичьих динозавров». Наука . 313 (5784): 213–217. Бибкод : 2006Sci...313..213E . дои : 10.1126/science.1125721 . ПМИД 16840697 . S2CID 34191607 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Обнаружены первые окаменелости эмбрионов тираннозавров» . Наука . 19 октября 2020 года. Архивировано из оригинала 5 декабря 2020 года . Проверено 23 ноября 2020 г.
- ^ Геггель, Лаура (15 октября 2020 г.). «Обнаружены первые в своем роде окаменелости детенышей тираннозавра» . www.livscience.com . Архивировано из оригинала 17 октября 2020 года . Проверено 17 октября 2020 г.
- ^ Энрикес, Натан Джеймс; Кампионе, Николас; Белл, Фил (21 апреля 2021 г.). «Толстногие родители тираннозавров не могли угнаться за своим более худым потомством, как показывают ископаемые следы» . Разговор . Архивировано из оригинала 26 апреля 2021 г. Проверено 28 апреля 2021 г.
- ^ Энрикес, Натан Дж.; Кампионе, Николас Э.; Бруэм, Том; Фанти, Федерико; Уайт, Мэтт А.; Сиссонс, Робин Л.; Салливан, Корвин; Ваврек, Мэтью Дж.; Белл, Фил Р. (21 апреля 2021 г.). «Изучение возможных онтогенетических траекторий тираннозаврид с использованием следов из формации Вапити (верхний кампан) в Альберте, Канада». Журнал палеонтологии позвоночных . 40 (6): e1878201. дои : 10.1080/02724634.2021.1878201 . S2CID 234814620 .
- ^ Эриксон, Грегори М.; Карри Роджерс, Кристи; Варриккио, Дэвид Дж.; Норелл, Марк; Сюй, Син (2007). «Модели роста высиживающих динозавров показывают сроки половой зрелости нептичьих динозавров и происхождение птичьего состояния» . Письма по биологии . 3 (5): 558–561. дои : 10.1098/rsbl.2007.0254 . ПМК 2396186 . ПМИД 17638674 .
- ^ Перейти обратно: а б с Ли, Эндрю Х.; Вернинг, Сара (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . ПНАС . 105 (2): 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ПМК 2206579 . ПМИД 18195356 .
- ^ Риклефс, Роберт Э. (2007). «Старение тираннозавра» . Письма по биологии . 3 (2): 214–217. дои : 10.1098/rsbl.2006.0597 . ПМЦ 2375931 . ПМИД 17284406 .
- ^ Перейти обратно: а б с д Танке, Даррен Х.; Карри, Филип Дж . (1998). «Кусание головы динозавров-теропод: палеопатологические данные» (PDF) . Гайя . 15 : 167–184. Архивировано (PDF) из оригинала 27 февраля 2008 г. Проверено 13 сентября 2008 г. [не печаталось до 2000 г.]
- ^ Перейти обратно: а б с д и Карри, Филип Дж .; Трекслер, Дэвид; Коппельхус, Ева Б .; Уикс, Келли; Мерфи, Нейт (2005). «Необычное костное ложе тираннозаврид, состоящее из нескольких особей, в формации Ту Медисин (поздний мел, кампан) в Монтане (США)». В Карпентер, Кеннет (ред.). Плотоядные динозавры . Блумингтон: Издательство Университета Индианы. стр. 313–324. ISBN 978-0-253-34539-4 .
- ^ «У Тираннозавра был плохой радиус поворота» . Архивировано из оригинала 7 мая 2012 года . Проверено 25 сентября 2007 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Кэрриер, Д.Р., Уолтер, Р.М. и Ли, Д.В. (15 ноября 2001 г.). «Влияние инерции вращения на поворотные способности динозавров-теропод: подсказки людей с повышенной инерцией вращения» . Журнал экспериментальной биологии . 204 (Часть 22): 3917–3926. дои : 10.1242/jeb.204.22.3917 . ПМИД 11807109 . Архивировано из оригинала 6 января 2008 года . Проверено 12 октября 2008 г.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Хатчинсон, младший (2004). «Биомеханическое моделирование и анализ чувствительности способности к двуногому бегу. II. Вымершие таксоны» (PDF) . Журнал морфологии . 262 (1): 441–461. дои : 10.1002/jmor.10240 . ПМИД 15352202 . S2CID 15677774 . Архивировано из оригинала (PDF) 31 октября 2008 г.
- ^ Перейти обратно: а б Хорнер, Джон Р .; Дон Лессем (1993). Полный Ти-рекс . Нью-Йорк : Саймон и Шустер . стр. 120 . ISBN 978-0-671-74185-3 .
- ^ Хольц, Томас Р. (1 мая 1996 г.). «Филогенетическая систематика целурозавров (Dinosauria; Theropoda)» . Журнал палеонтологии . 70 (3): 536–538. Бибкод : 1996JPal...70..536H . дои : 10.1017/S0022336000038506 . S2CID 87599102 . Архивировано из оригинала 26 октября 2008 года . Проверено 3 октября 2008 г.
- ^ Кристиансен, П. (1998). «Значения показателей силы длинных костей теропод с комментариями к пропорциям конечностей и беговому потенциалу» (PDF) . Гайя . 15 : 241–255. ISSN 0871-5424 . Архивировано (PDF) из оригинала 31 октября 2008 г. Проверено 12 октября 2008 г.
- ^ Фарлоу, Джеймс О .; Смит, Мэтью Б.; Робинсон, Джон М. (1995). «Масса тела, «показатель прочности» костей и беговой потенциал тираннозавра рекса » . Журнал палеонтологии позвоночных . 15 (4): 713–725. Бибкод : 1995JVPal..15..713F . дои : 10.1080/02724634.1995.10011257 . Архивировано из оригинала 23 октября 2008 г.
- ^ «Чем больше они становятся, тем сильнее они падают». Архивировано 31 мая 2015 г. в Wayback Machine New Scientist, 7 октября 1995 г., стр. 18.
- ^ «Жираф» . WildlifeSafari.info. Архивировано из оригинала 12 июня 2010 года . Проверено 29 апреля 2006 г.
- ^ «История зоопарка Вудленд-Парк — Глава 4» . Архивировано из оригинала 2 июня 2007 года . Проверено 29 апреля 2006 г.
- ^ Александр, РМ (7 августа 2006 г.). «Биомеханика динозавров» . Учебник по биологическим наукам . 273 (1596): 1849–1855. дои : 10.1098/rspb.2006.3532 . ПМЦ 1634776 . ПМИД 16822743 .
- ^ Ханна, Ребекка Р. (2002). «Множественные травмы и инфекции у подростков-тероподов ( Allosaurus fragilis ) в сравнении с патологией аллозавров в коллекции карьеров динозавров Кливленда-Ллойда». Журнал палеонтологии позвоночных . 22 (1): 76–90. doi : 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2 . ISSN 0272-4634 . S2CID 85654858 . каталогизирует травмы аллозавра , известного как « Большой Эл » — по крайней мере, одна из них была приписана падению.
- ^ Хатчинсон, Джон Р.; Гарсия, М. (28 февраля 2002 г.). « Тираннозавр не был быстрым бегуном» . Природа . 415 (6875): 1018–1021. Бибкод : 2002Natur.415.1018H . дои : 10.1038/4151018a . ПМИД 11875567 . S2CID 4389633 .
- ^ Хайдул, Р. (1997). Сухожилия. Карточки с динозаврами . ООО «Орбис Паблишинг» D36044311.
- ^ Селлерс, Висконсин и Мэннинг, Польша (июль 2007 г.). «Оценка максимальной скорости бега динозавров с помощью эволюционной робототехники» . Учеб. Р. Сок. Б. 274 (1626): 2711–6. дои : 10.1098/rspb.2007.0846 . ПМК 2279215 . ПМИД 17711833 . Архивировано из оригинала 7 февраля 2011 г. Проверено 12 октября 2008 г. Это может быть предварительная версия Селлерса, Висконсин, Мэннинга, П.Л., Кромптона, Р.Х. и Кодда, младшего. (2007), «Изучение эффектов накопления упругой энергии при двуногом передвижении с использованием эволюционной робототехники», Журнал биомеханики , в обзоре.
- ^ Лиз Сьюард: «Ти-рекс 'обгонит футболиста'». Архивировано 7 января 2012 г. в Wayback Machine . Веб-сайт BBC News , вторник, 21 августа 2007 г. В статье цитируется доктор Билл Селлерс из Манчестерского университета, соавтор статьи, опубликованной в журнале Proceedings of the Royal Society B. Проверено 22 августа 2007 г.
- ^ Каллисон, Г.; Его Величество Куимби (1984). «Крошечные динозавры: они выросли?». Журнал палеонтологии позвоночных . 3 (4): 200–209. Бибкод : 1984JVPal...3..200C . дои : 10.1080/02724634.1984.10011975 .
- ^ Снайвли, Эрик; О'Брайен, Хейли; Хендерсон, Дональд М.; Мэллисон, Генрих; Сюрринг, Лара А.; Бернс, Майкл Э.; младший, Томас Р. Хольц; Рассел, Энтони П.; Уитмер, Лоуренс М.; Карри, Филип Дж.; Хартман, Скотт А.; Коттон, Джон Р. (21 февраля 2019 г.). «Меньшая вращательная инерция и большие мышцы ног указывают на более быстрые повороты у тираннозаврид, чем у других крупных теропод» . ПерДж . 7 : е6432. дои : 10.7717/peerj.6432 . ПМК 6387760 . ПМИД 30809441 .
- ^ Дечекки, Т. Александр; Млошевская, Александра М.; Хольц, Томас Р.; Хабиб, Майкл Б.; Ларссон, Ханс CE (2020). «Быстрый и бережливый: различные стратегии локомоторики приводят к удлинению конечностей у тероподных динозавров» . ПЛОС ОДИН . 15 (5): e0223698. Бибкод : 2020PLoSO..1523698D . дои : 10.1371/journal.pone.0223698 . ПМК 7220109 . ПМИД 32401793 .
- ^ «Ти-рекс был чемпионом по ходьбе, сверхэффективным на низких скоростях» . ЭврекАлерт! . Архивировано из оригинала 27 июня 2020 г. Проверено 27 июня 2020 г.
- ^ Чжоу Чжунхэ, Z; Барретт, Пол М.; Хилтон, Джейсон. (2003). «Исключительно сохранившаяся экосистема нижнего мела». Природа . 421 (6925): 807–814. Бибкод : 2003Natur.421..807Z . дои : 10.1038/nature01420 . ПМИД 12594504 . S2CID 4412725 .
- ^ Чэнь Пейджи, Пэй-цзи; Донг Чжимин ; Чжэнь Шуонань. (1998). «Исключительно хорошо сохранившийся динозавр-теропод из формации Исянь в Китае» (PDF) . Природа . 391 (6663): 147–152. Бибкод : 1998Natur.391..147C . дои : 10.1038/34356 . S2CID 4430927 .
- ^ Сюй Син, X ; Чжоу Чжунхэ; Прум, Ричард А. (2003). «Разветвленные покровные структуры у синорнитозавра и происхождение перьев». Природа . 410 (6825): 200–204. Бибкод : 2001Natur.410..200X . дои : 10.1038/35065589 . ПМИД 11242078 . S2CID 4426803 .
- ^ Лингем-Солиар, Теагартен; Федучча, Алан ; Ван, Сяолинь (2007). «Новый китайский образец указывает на то, что «протоперья» раннемелового тероподного динозавра Sinosauropteryx представляют собой деградировавшие коллагеновые волокна» . Труды Лондонского королевского общества. Серия Б, Биологические науки . 274 (1620): 1823–1829. дои : 10.1098/rspb.2007.0352 . ПМК 2270928 . ПМИД 17521978 .
- ^ «Архивная копия» (PDF) . Архивировано из оригинала (PDF) 17 апреля 2012 г. Проверено 4 апреля 2012 г.
{{cite web}}: CS1 maint: архивная копия в заголовке ( ссылка ) - ^ Перейти обратно: а б Белл, PR; Кампионе, Невада; Лица, WS; Карри, Пи Джей; Ларсон, Польша; Танке, Д.Х.; Баккер, RT (2017). «Покровы тираннозавроидов демонстрируют противоречивые закономерности гигантизма и эволюции перьев» . Письма по биологии . 13 (6): 20170092. doi : 10.1098/rsbl.2017.0092 . ПМЦ 5493735 . ПМИД 28592520 .
- ^ http://globalnews.ca/news/3506400/tyrannosaurus-rex-fossil-skin-scales-university-of-alberta/. Архивировано 17 июня 2017 г. в Wayback Machine. Что чувствует тираннозавр? Палеонтологи Университета Австралии узнали об открытии новой окаменелости
- ^ Карри, П.Дж., Бадамгарав, Д., Коппельхус, Э.Б. 2003. Первые позднемеловые следы из местонахождения Немегт в Гоби, Монголия. Ихнос. Том 10:1-12. Карри, П.Дж., Бадамгарав, Д., Коппельхус, Э.Б. 2003.
- ^ Карр, Т.Д., Варриккио, DJ, Седлмайр, JC, Робертс, EM, и Мур, JR (2017). Новый тираннозавр с доказательствами анагенеза и сенсорной системой лица, напоминающей крокодиловую. Научные отчеты , 7.
- ^ Виттон, Марк; Хоун, Дэвид (сентябрь 2018 г.). «Тероподы-тираннозавриды: они когда-нибудь улыбались, как крокодилы?» (PDF) . Ежегодный симпозиум по палеонтологии и сравнительной анатомии позвоночных .
- ^ Сайтта, Эван Т.; Флетчер, Ян; Мартин, Питер; Питтман, Майкл; Кэй, Томас Г.; Верно, Лоуренс Д.; Норелл, Марк А.; Эбботт, Джеффри Д.; Вызов, Роджер Э.; Пенкман, Кирсти; Винтер, Якоб (01 ноября 2018 г.). «Сохранение волокон перьев позднемелового динозавра Shuvuuia Deserti вызывает обеспокоенность по поводу иммуногистохимического анализа окаменелостей» (PDF) . Органическая геохимия . 125 : 142–151. Бибкод : 2018OrGeo.125..142S . doi : 10.1016/j.orggeochem.2018.09.008 . ISSN 0146-6380 . S2CID 105753275 .
- ^ блоггер, Гость. «Десять вещей, которые мы не знаем о тираннозаврах» . Архивировано из оригинала 30 сентября 2020 г. Проверено 05 января 2021 г.
- ^ Гуарино, Бен (27 октября 2021 г.). «У тираннозавра рекса была чешуйчатая кожа, и он не был покрыт перьями, говорится в новом исследовании» . Вашингтон Пост . ISSN 0190-8286 . Проверено 7 августа 2023 г.
- ^ Стивенс, Кент А. (июнь 2006 г.). «Бинокулярное зрение динозавров-теропод» (PDF) . Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 . ISSN 0272-4634 . S2CID 85694979 . Архивировано из оригинала (PDF) 23 марта 2012 г.
- ^ Яффе, Эрик (1 июля 2006 г.). «Зрение для глаз Саура: зрение тираннозавра было одним из лучших в природе» . Новости науки . 170 (1): 3–4. дои : 10.2307/4017288 . JSTOR 4017288 . Архивировано из оригинала 29 сентября 2012 года . Проверено 6 октября 2008 г.
- ^ Савельев Сергей Владимирович; Алифанов, Владимир Р. (2005). «Новое исследование мозга хищного динозавра Tarbosaurus bataar (Theropoda, Tyrannosauridae)». Палеонтологический журнал . 41 (3): 281–289. дои : 10.1134/S0031030107030070 . S2CID 53529148 .
- ^ Перейти обратно: а б Карр, Томас Д.; Варриккио, Дэвид Дж.; Седлмайр, Джейк К.; Робертс, Эрик М.; Мур, Джейсон Р. (30 марта 2017 г.). «Новый тираннозавр с доказательствами анагенеза и сенсорной системой лица, подобной крокодиловой» . Научные отчеты . 7 : 44942. Бибкод : 2017NatSR...744942C . дои : 10.1038/srep44942 . ISSN 2045-2322 . ПМЦ 5372470 . ПМИД 28358353 .
- ^ Баркер, Крис; Нэйш, Даррен; Кацаменис, Орестис; Дайк, Гарет (16 июня 2017 г.). «Сложная нейроанатомия рострума теропода Neovenator salerii с острова Уайт» . Научные отчеты . 7 (1): 3749. Бибкод : 2017НатСР...7.3749Б . дои : 10.1038/s41598-017-03671-3 . ПМК 5473926 . ПМИД 28623335 .
- ^ Лейтч, Дункан Б.; Катания, Кеннет К. (01 декабря 2012 г.). «Строение, иннервация и ответные свойства покровных органов чувств крокодилов» . Журнал экспериментальной биологии . 215 (23): 4217–4230. дои : 10.1242/jeb.076836 . ISSN 0022-0949 . ПМК 4074209 . ПМИД 23136155 .
- ^ Ди-Пои, Николя; Милинкович, Мишель К. (2 июля 2013 г.). «У крокодилов развились рассеянные мультисенсорные микроорганизмы» . ЭвоДево . 4 (1): 19. дои : 10.1186/2041-9139-4-19 . ISSN 2041-9139 . ПМЦ 3711810 . ПМИД 23819918 .
- ^ Баккер, Роберт Т. (1968). «Превосходство динозавров». Открытие . 3 (2): 11–12.
- ^ Баккер, Роберт Т. (1972). «Анатомические и экологические доказательства эндотермии у динозавров». Природа . 238 (5359): 81–85. Бибкод : 1972Natur.238...81B . дои : 10.1038/238081a0 . S2CID 4176132 .
- ^ Баккер, Роберт Т. (1986). Ересь динозавров . Нью-Йорк: Издательство Кенсингтон. ISBN 978-0-688-04287-5 . OCLC 13699558 . [ нужна страница ]
- ^ Баррик, Риз Э.; Уильям Дж. Шауэрс (июль 1994 г.). «Термофизиология тираннозавра рекса: данные по изотопам кислорода». Наука . 265 (5169). Нью-Йорк : 222–224. Бибкод : 1994Sci...265..222B . дои : 10.1126/science.265.5169.222 . ПМИД 17750663 . S2CID 39392327 .
- ^ Баррик, Риз Э.; Уильям Дж. Шауэрс (октябрь 1999 г.). «Термофизиология и биология гиганотозавра : сравнение с тираннозавром » . Электронная палеонтология . 2 (2). Архивировано из оригинала 17 мая 2011 г. Проверено 12 октября 2008 г.
- ^ Паладино, Фрэнк В.; Спотила, Джеймс Р.; Додсон, Питер (1999). «Проект гигантов: моделирование физиологии крупных динозавров». У Джеймса О. Фарлоу ; М.К. Бретт-Сурман (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 491–504. ISBN 978-0-253-21313-6 .
- ^ Чинсами, Анусуя; Виллем Дж. Хиллениус (2004). «Физиология нептичьих динозавров». У Дэвида Б. Вейшампеля; Питер Додсон; Гальшка Осмольска (ред.). Динозаврия . Беркли: Издательство Калифорнийского университета. стр. 643–659. ISBN 978-0-520-24209-8 .
- ^ Сеймур, Роджер С. (5 июля 2013 г.). «Максимальная аэробная и анаэробная выработка энергии у крупных крокодилов по сравнению с млекопитающими: последствия гигантотермии динозавров» . ПЛОС ОДИН . 8 (7): e69361. Бибкод : 2013PLoSO...869361S . дои : 10.1371/journal.pone.0069361 . ISSN 1932-6203 . ПМК 3702618 . ПМИД 23861968 .
- ^ Перейти обратно: а б Фарлоу, Джеймс О .; Пианка, Эрик Р. (2002). «Перекрытие размеров тела, разделение среды обитания и требования к жизненному пространству наземных позвоночных хищников: последствия для палеоэкологии крупных динозавров-теропод» (PDF) . Историческая биология . 16 (1): 21–40. Бибкод : 2002HBio...16...21F . дои : 10.1080/0891296031000154687 . S2CID 18114585 .
- ^ Варриккио, Дэвид Дж. (2001). «Содержимое кишечника мелового тираннозаврид: значение для пищеварительного тракта тероподных динозавров» (PDF) . Журнал палеонтологии . 75 (2): 401–406. doi : 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2 . ISSN 0022-3360 . S2CID 86113170 .
- ^ Guinness World Records Ltd. (2003). Книга рекордов Гиннеса 2003 года . стр 90.
- ^ Перейти обратно: а б Эберт, Дэвид А.; МакКри, Ричард Т. (2001). «Были ли крупные тероподы стадными?». Журнал палеонтологии позвоночных . 21 (Дополнение к номеру 3): 46А. дои : 10.1080/02724634.2001.10010852 . S2CID 220414868 .
- ^ Роуч, Брайан Т.; Бринкман, Дэниел Л. (2007). «Переоценка совместной охоты и стадности Deinonychus antirhopus и других нептичьих динозавров-тероподов». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- ^ Перейти обратно: а б Карри, Филип Дж . (1998). «Возможные доказательства стадного поведения тираннозаврид» (PDF) . Гайя . 15 : 271–277. Архивировано (PDF) из оригинала 26 марта 2009 г. Проверено 13 сентября 2008 г. (не печаталось до 2000 г.)
- ^ Роуч, Брайан Т.; Бринкман, Дэниел Т. (2007). «Переоценка совместной охоты и стадности Deinonychus antirhopus и других нептичьих динозавров-тероподов». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- ^ МакКри, Ричард Т.; Бакли, Лиза Г.; Фарлоу, Джеймс О.; Локли, Мартин Г.; Карри, Филип Дж.; Мэтьюз, Неффра А.; Пембертон, С. Джордж (2014). «Ужас тираннозавров»: первые следы тираннозаврид и свидетельства стадности и патологии тираннозаврид» . ПЛОС ОДИН . 9 (7): е103613. Бибкод : 2014PLoSO...9j3613M . дои : 10.1371/journal.pone.0103613 . ПМК 4108409 . ПМИД 25054328 .
- ^ «Следы тираннозавра показывают походку динозавра» . 23 июля 2014 г. Архивировано из оригинала 30 сентября 2014 г. Проверено 16 января 2015 г.
- ^ «Следы намекают на социальную жизнь динозавров-тиранов» . 23 июля 2014 г. Архивировано из оригинала 8 января 2015 г. Проверено 16 января 2015 г.
- ^ Титус, Алан Л.; Нолл, Катя; Сертич, Джозеф JW; Ямамура, Дайго; Суарес, Селина А.; Гласспул, Ян Дж.; Жинувес, Джонатан Э.; Лукачич, Эбигейл К.; Робертс, Эрик М. (19 апреля 2021 г.). «Геология и тафономия уникального костного пласта тираннозаврид из формации Кайпаровиц верхнего кампана на юге Юты: последствия для стадности тираннозаврид» . ПерДж . 9 : e11013. дои : 10.7717/peerj.11013 . ПМК 8061582 . ПМИД 33976955 .
- ^ Эйльперин, Джульетта (19 апреля 2021 г.). «Тираннозавры, скорее всего, охотились стаями, а не в одиночку, считают ученые» . Вашингтон Пост . Архивировано из оригинала 20 апреля 2021 года . Проверено 27 сентября 2021 г.
- ^ «Тираннозавр Рекс «охотился стаями» » . 22 июня 2011 г. Архивировано из оригинала 27 сентября 2021 г. Проверено 27 сентября 2021 г.
- ^ Перейти обратно: а б с Якобсен, А.Р. 2001. Маленькая кость теропода с отмеченными зубами: чрезвычайно редкий след. п. 58-63. В: Жизнь мезозойских позвоночных. Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Индианского университета.
- ^ Перейти обратно: а б Аблер, В.Л. 2001. Модель зубцов тираннозавра, сделанная с помощью пропила и сверла. п. 84-89. В: Жизнь мезозойских позвоночных . Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Индианского университета.
- ^ Ламбе, Л.Б. (1917). «Меловой тероподный динозавр Горгозавр » . Мемуары Геологической службы Канады . 100 : 1–84. дои : 10.4095/101672 . Архивировано из оригинала 9 сентября 2021 г. Проверено 23 сентября 2021 г.
- ^ Фарлоу, Джо; Хольц, Т.Р. (2002). Ковалевски, М.; Келли, PH (ред.). «Ископаемые записи хищничества» (PDF) . Документы Палеонтологического общества. стр. 251–266. Архивировано из оригинала (PDF) 31 октября 2008 г.
- ^ Дори, М. (1997). Тиранозавр. Карточки с динозаврами . ООО «Орбис Паблишинг» D36045907.
- ^ Перейти обратно: а б Хорнер, младший (1994). «Ножи для стейка, глаза-бусинки и крохотные ручки (портрет тираннозавра -падальщика)». Специальное издание Палеонтологического общества . 7 : 157–164. дои : 10.1017/S2475262200009497 .
- ^ Амос, Дж. (31 июля 2003 г.). «Ти-рекс предстанет перед судом» . Би-би-си. Архивировано из оригинала 13 марта 2012 года . Проверено 12 октября 2008 г.
- ^ Перейти обратно: а б Уолтерс М., Пакер Дж. (1995). Словарь доисторической жизни. Книги Клермонта. ISBN 1-85471-648-4 .
- ^ Эриксон ГМ; Олсон К.Х. (1996). «Следы укусов тираннозавра рекса : предварительное описание и последствия» . Журнал палеонтологии позвоночных . 16 (1): 175–178. Бибкод : 1996JVPal..16..175E . дои : 10.1080/02724634.1996.10011297 . Архивировано из оригинала 19 октября 2021 г. Проверено 12 сентября 2020 г.
- ^ Карпентер, К. (1998). «Свидетельства хищнического поведения динозавров-теропод» . Гайя . 15 : 135–144. Архивировано из оригинала 17 ноября 2007 года . Проверено 5 декабря 2007 г.
- ^ Фаулер, Денвер В.; Роберт М. Салливан (2006). «Таз цератопсида со следами зубов из формации Киртланд верхнего мела, Нью-Мексико: свидетельства пищевого поведения тираннозаврид позднего кампана». Бюллетень Музея естественной истории и науки Нью-Мексико . 35 : 127–130.
- ^ Терриен, Ф.; Зеленицкий, ДК; Танака, К.; Ворис, Дж. Т.; Эриксон, генеральный директор; Карри, Пи Джей; ДеБур, CL; Кобаяши, Ю. (2023). «Исключительно сохранившееся содержимое желудка молодого тираннозавра указывает на онтогенетические изменения в рационе культового вымершего хищника» . Достижения науки . 9 (49): eadi0505. Бибкод : 2023SciA....9I.505T . дои : 10.1126/sciadv.adi0505 . ПМЦ 10846869 . ПМИД 38064561 .
- ^ Лонгрич, Северная Каролина; Хорнер, младший; Эриксон, генеральный директор; Карри, Пи Джей (2010). «Каннибализм у тираннозавра рекса » . ПЛОС ОДИН . 5 (10): е13419. Бибкод : 2010PLoSO...513419L . дои : 10.1371/journal.pone.0013419 . ПМЦ 2955550 . ПМИД 20976177 .
- ^ «НОВЫЕ СВИДЕТЕЛЬСТВА КАННИБАЛИЗМА У ТИРАННОЗАВРИДОВ ДИНОЗАВРОВ ИЗ ВЕРХНЕГО МЕЛА (КАМПАН/МААСТРИХТ) БАССЕЙНА САН-ХУАН В НЬЮ-МЕКСИКЕ» . Исследовательские ворота . Архивировано из оригинала 10 ноября 2021 г. Проверено 6 января 2021 г.
- ^ Додсон, Питер; Бритт, Брукс; Карпентер, Кеннет; Форстер, Кэтрин А.; Джилетт, Дэвид Д.; Норелл, Марк А.; Ольшевский, Георгий; Пэрриш, Дж. Майкл и Вейшампель, Дэвид Б. (1993). «Абелизавр». Эпоха динозавров . Публикации Интернешнл, ООО. п. 105. ИСБН 978-0-7853-0443-2 .
- ^ Фиорилло, Энтони Р.; Ганглофф, Роланд А. (2000). «Зубы теропода из формации Принс-Крик (мел) на севере Аляски, с размышлениями о палеоэкологии арктических динозавров». Журнал палеонтологии позвоночных . 20 (4): 675–682. doi : 10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2 . ISSN 0272-4634 . S2CID 130766946 .
- ^ «Эти зубы возрастом 81 миллион лет — первое свидетельство существования самого большого динозавра в истории Японии | Irish Examiner» . Архивировано из оригинала 12 апреля 2016 г. Проверено 10 марта 2016 г.
- ^ Томас Р. Хольц младший (1994). «Филогенетическое положение тираннозаврид: значение для систематики теропод». Журнал палеонтологии . 68 (5): 1100–1117. Бибкод : 1994JPal...68.1100H . дои : 10.1017/S0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Февраль 2016 г., Лаура Геггель 29 (29 февраля 2016 г.). «Ти-Рекс, вероятно, был инвазивным видом» . www.livscience.com . Архивировано из оригинала 14 июля 2019 года . Проверено 5 января 2021 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- Список образцов тираннозаврид и обсуждение в базе данных Theropod















- ^ Иероним, Тобин; Уитмер, Лоуренс; Танке, Даррен; Карри, Филип (26 августа 2009 г.). «Лицевые покровы центрозавровых цератопсид: морфологические и гистологические корреляты новых структур кожи» . Анатомическая запись . 292 (9): 1370–1396. дои : 10.1002/ar.20985 . ПМИД 19711467 .