Белковая нить
Эта статья может быть слишком технической для понимания большинства читателей . ( Октябрь 2023 г. ) |

В биологии представляет белковая нить собой длинную цепочку белковых мономеров, например, тех, что содержатся в волосах, мышцах или жгутиках . [1] Белковые нити формируются вместе и образуют цитоскелет клетки. Они часто объединяются вместе, чтобы обеспечить поддержку, прочность и жесткость клетки. Когда нити собраны вместе, они могут образовывать три разные клеточные части. Три основных класса белковых филаментов, составляющих цитоскелет, включают: актиновые филаменты , микротрубочки и промежуточные филаменты .
Типы клеток
[ редактировать ]Микрофиламенты
[ редактировать ]По сравнению с другими частями цитоскелета микрофиламенты содержат самые тонкие нити диаметром около 7 нм. Микрофиламенты являются частью цитоскелета, состоящего из белка актина . Две нити актина, переплетенные вместе, образуют нитевидную структуру, обеспечивающую движение моторных белков. Микрофиламенты могут встречаться либо в виде мономерного G-актина, либо в виде нитчатого F-актина. [2] Микрофиламенты важны, когда речь идет об общей организации плазматической мембраны. Актиновые нити считаются одновременно спиральными и гибкими. Они состоят из нескольких мономеров актина, соединенных вместе, что придает им гибкость. Они встречаются в нескольких местах тела, включая микроворсинки, сократительные кольца, стрессовые волокна, клеточную кору и т. д. В сократительном кольце актин обладает способностью способствовать клеточному делению, а в клеточной коре он может способствовать сохранению структурной целостности. клетки.
- Полимеризация микрофиламентов
микрофиламентов Полимеризация делится на три этапа. Стадия нуклеации является первой, лимитирующей и самой медленной стадией процесса. Следующим шагом в этом процессе является элонгация, представляющая собой быстрое добавление мономеров актина как на плюсовом, так и на минусовом конце микрофиламента. Последний этап – установившееся состояние. В этом состоянии добавление мономеров будет равносильно вычитанию мономеров, в результате чего микрофиламент больше не будет расти. Это известно как критическая концентрация актина. Известно несколько токсинов, которые ограничивают полимеризацию актина. Цитохалазин — это токсин, который связывается с полимером актина, поэтому он больше не может связываться с поступающими мономерами актина. Актин, первоначально прикрепленный к полимеру, все еще покидает микрофиламент, вызывая деполимеризацию. Фаллоидин — это токсин, который связывается с актином, фиксируя нить на месте. Мономеры не добавляют и не покидают этот полимер, что приводит к стабилизации молекулы. Латрункулин подобен цитохалазину, но представляет собой токсин, который связывается с мономерами актина, предотвращая его присоединение к полимеру актина. Это приведет к деполимеризации полимера актина в клетке. [ нужна ссылка ]
- Моторный белок на основе актина – миозин

Существует несколько различных белков, которые взаимодействуют с актином в организме. Однако одним из самых известных типов моторных белков является миозин . Миозин связывается с этими актинами, вызывая движение актина. Это движение миозина вдоль микрофиламента может вызывать сокращение мышц, ассоциацию мембран, эндоцитоз и транспорт органелл. Актиновое микрофиламент состоит из трех полос и одного диска. Полоса А — это часть актина, которая связывается с миозином во время мышечного сокращения. Группа I — это часть актина, которая не связана с миозином, но все равно будет двигаться во время мышечного сокращения. Зона H — это пространство между двумя соседними актинами, которое сжимается, когда мышца начинает сокращаться. Z-диск является частью микрофиламента, который характеризует общий конец каждой стороны саркомера , структурной единицы миофибриллы . [ нужна ссылка ]
- Белки, ограничивающие микрофиламенты
Эти микрофиламенты могут быть ограничены несколькими факторами или белками. Тропомодулин — это белок, который покрывает концы актиновых нитей, обеспечивая общую стабильность структуры. Небулин — еще один белок, который может связываться с сторонами актина, предотвращая прикрепление к ним миозина. Это вызывает стабилизацию актина, ограничивающего мышечное сокращение. Титин — еще один белок, но он связывается с миозином, а не с актиновыми микрофиламентами. Титин поможет стабилизировать сокращение и структуру миозина-актина. [ нужна ссылка ]
Микротрубочки
[ редактировать ]
Микротрубочки — это самый крупный тип нитей цитоскелета диаметром 25 нм. [3] Одна микротрубочка состоит из 13 линейных микрофиламентов. В отличие от микрофиламентов, микротрубочки состоят из белка тубулина. Тубулин состоит из димеров, называемых «αβ-тубулин» или «димеры тубулина», которые полимеризуются с образованием микротрубочек. [3] Эти микротрубочки структурно разделены на три основные группы: синглеты, дублеты и триплеты. Синглеты представляют собой структуры микротрубочек, которые, как известно, обнаруживаются в цитоплазме . Дублеты — это структуры, обнаруженные в ресничках и жгутиках . Триплеты встречаются в базальных тельцах и центриолях. Существует две основные популяции этих микротрубочек. Существуют нестабильные короткоживущие микротрубочки, которые быстро собираются и разбираются. Другая популяция представляет собой стабильные долгоживущие микротрубочки. Эти микротрубочки остаются полимеризованными в течение более длительных периодов времени и могут быть обнаружены в жгутиках, эритроцитах и нервных клетках. Микротрубочки обладают способностью играть значительную роль в организации органелл и везикул, биении ресничек и жгутиков, структуре нервов и эритроцитов, а также выравнивании/разделении хромосом во время митоза и мейоза. [ нужна ссылка ]
- Ориентация в клетках
Когда клетка находится в интерфазном процессе, все микротрубочки имеют тенденцию ориентироваться одинаково. Их отрицательно заряженный конец будет расположен близко к ядру клетки, а положительно – от тела клетки. Базальное тельце, находящееся внутри клетки, помогает микротрубочкам ориентироваться определенным образом. В митотических клетках они будут видеть аналогичную ориентацию, поскольку положительно заряженный конец будет ориентирован от клетки, а отрицательно заряженный конец будет направлен к центру организации микротрубочек (MTOC). Положительный конец этих микротрубочек прикрепится к кинетохору хромосомы, обеспечивая возможность клеточного деления, когда это возможно. Нервные клетки имеют тенденцию отличаться от этих двух других форм ориентации. В аксонной нервной клетке микротрубочки располагаются своим отрицательно заряженным концом к телу клетки, а положительно заряженным концом - вдали от тела клетки. Однако в дендритах микротрубочки могут иметь другую ориентацию. В дендритах положительно заряженный конец микротрубочек может быть обращен к телу клетки, но его отрицательно заряженный конец, скорее всего, будет находиться вдали от тела клетки. [ нужна ссылка ]
- Лекарства, разрушающие микротрубочки
Колхицин является примером препарата, который, как известно, используется в качестве ингибитора микротрубочек. Он связывается как с α-, так и с β- тубулином на димерах в микротрубочках. При низких концентрациях это может вызвать стабилизацию микротрубочек, но при высоких концентрациях может привести к деполимеризации микротрубочек. Таксол – еще один препарат, который часто используется для лечения рака молочной железы путем воздействия на микротрубочки. Таксол связывается с стенками канальцев и может привести к нарушению деления клеток. [ нужна ссылка ]
- Роль в подразделении сотовой связи

Существует три основных типа микротрубочек, участвующих в клеточном делении . Астральные микротрубочки — это те, которые выходят из центросомы к коре клетки. Они могут соединяться с плазматической мембраной через кортикальные ориентиры. Эти отложения определяются с помощью сигналов полярности, факторов роста и дифференциации или адгезионных контактов. Полярные микротрубочки будут простираться к середине клетки и перекрывать экватор, где клетка делится. Микротрубочки кинетохора будут расширяться и связываться с кинетохорами на хромосомах, помогая делению клетки. Эти микротрубочки прикрепляются к кинетохору на своем положительном конце. NDC80 — это белок, обнаруженный в этой точке связывания, который поможет стабилизировать это взаимодействие во время клеточного деления. В процессе клеточного деления общая длина микротрубочек не меняется. Однако это вызовет эффект беговой дорожки, который может вызвать разделение этих хромосом. [ нужна ссылка ]
Промежуточные нити
[ редактировать ]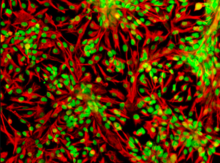
Промежуточные филаменты являются частью структуры цитоскелета, обнаруженной в большинстве эукариотических клеток. Примером промежуточной нити является нейрофиламент . Они обеспечивают поддержку структуры аксона и являются основной частью цитоскелета. Промежуточные филаменты имеют средний диаметр 10 нм, что меньше, чем у микротрубочек, но больше, чем у микрофиламентов. [4] Эти нити размером 10 нм состоят из полипептидных цепей, принадлежащих к тому же семейству, что и промежуточные нити. Промежуточные филаменты не участвуют в прямом движении клеток в отличие от микротрубочек и микрофиламентов. Промежуточные нити могут играть роль в клеточной коммуникации в процессе, известном как перекрестные помехи. Эти перекрестные помехи могут помочь в механоощущении. Это механочувствительность может помочь защитить клетку во время клеточной миграции внутри организма. Они также могут помочь в связывании актина и микротрубочек с цитоскелетом, что в конечном итоге приведет к движению и делению клеток. Наконец, эти промежуточные нити обладают способностью улучшать проницаемость сосудов за счет организации непрерывных слипчивых соединений посредством сшивки плектина. [5]
- Классификация промежуточных филаментов
Промежуточные филаменты состоят из нескольких белков, в отличие от микрофиламентов и микротрубочек, которые состоят в основном из актина и тубулина. Эти белки были разделены на 6 основных категорий на основе их схожих характеристик. Промежуточные нити типа 1 и 2 состоят из кератина и в основном встречаются в эпителиальных клетках. Промежуточные филаменты 3-го типа содержат виментин. Их можно найти в различных клетках, включая гладкомышечные клетки, фибробласты и лейкоциты. Промежуточные филаменты 4-го типа — это нейрофиламенты, обнаруженные в нейронах. Их можно найти во многих различных моторных аксонах, поддерживающих эти клетки. Промежуточные филаменты типа 5 состоят из ядерных ламинов, которые можно найти в ядерной оболочке многих эукариотических клеток. Они помогут собрать в этих клетках ортогональную сеть ядерной мембраны. Промежуточные филаменты 6-го типа связаны с нестином, который взаимодействует со стволовыми клетками центральной нервной системы. [6]
Ссылки
[ редактировать ]- ^ Шпеер Б., Ваггонер Б. (13 августа 1995 г.). «Нитья» . Глоссарий UCMP: Клеточная биология . Беркли, Калифорния: Музей палеонтологии Калифорнийского университета . Проверено 2 ноября 2011 г.
- ^ Хоманн Т., Дегани Ф. (апрель 2019 г.). «Цитоскелет — сложная взаимодействующая сеть» . Клетки . 8 (4): 362. doi : 10.3390/cells8040362 . ПМК 6523135 . ПМИД 31003495 .
- ^ Jump up to: а б Гудсон Х.В., Джонассон Э.М. (июнь 2018 г.). «Микротрубочки и белки, ассоциированные с микротрубочками» . Перспективы Колд-Спринг-Харбор в биологии . 10 (6): а022608. doi : 10.1101/cshperspect.a022608 . ПМЦ 5983186 . ПМИД 29858272 .
- ^ Херрманн Х, Эби У (ноябрь 2016 г.). «Промежуточные нити: структура и сборка» . Перспективы Колд-Спринг-Харбор в биологии . 8 (11): а018242. doi : 10.1101/cshperspect.a018242 . ПМК 5088526 . ПМИД 27803112 .
- ^ Ндиай, Энн-Бетти; Кендеринк, Гейсье Х. ; Шемеш, Михал (2022). «Промежуточные филаменты клеточной механореактивности: опосредование цитоскелетных перекрестных помех от мембраны к ядру и обратно» . Границы клеточной биологии и биологии развития . 10 : 882037. doi : 10.3389/fcell.2022.882037 . ISSN 2296-634X . ПМЦ 9035595 . ПМИД 35478961 .
- ^ Купер, Джеффри М. (2000). «Промежуточные нити» . Клетка: молекулярный подход. 2-е издание .