Деление клеток

Деление клеток — это процесс, при котором родительская клетка делится на две дочерние клетки. [1] Деление клетки обычно происходит как часть более крупного клеточного цикла , в котором клетка растет и реплицирует свою хромосому (хромосомы) перед делением. У эукариот существует два различных типа деления клеток: вегетативное деление ( митоз ), производящее дочерние клетки, генетически идентичные родительской клетке, и клеточное деление, приводящее к образованию гаплоидных гамет для полового размножения ( мейоз ), уменьшающее количество хромосом от по два каждого типа в диплоидной родительской клетке и по одному каждого типа в дочерних клетках. [2] Митоз — это часть клеточного цикла , при которой реплицированные хромосомы разделяются на два новых ядра . Деление клеток приводит к образованию генетически идентичных клеток, в которых сохраняется общее число хромосом. Как правило, митозу (делению ядра) предшествует S-стадия интерфазы (во время которой происходит репликация ДНК ), за которой следуют телофаза и цитокинез ; который делит цитоплазму , органеллы и клеточную мембрану одной клетки на две новые клетки, содержащие примерно равные доли этих клеточных компонентов. Различные стадии митоза в совокупности определяют фазу М животных клеточного цикла — деление материнской клетки на две генетически идентичные дочерние клетки. [3] Чтобы обеспечить правильное продвижение по клеточному циклу, повреждения ДНК выявляются и устраняются на различных контрольных точках на протяжении всего цикла. Эти контрольные точки могут остановить развитие клеточного цикла, ингибируя определенные комплексы циклин-CDK . Мейоз претерпевает два деления, в результате чего образуются четыре гаплоидные дочерние клетки. Гомологичные хромосомы разделяются при первом делении мейоза, так что каждая дочерняя клетка имеет по одной копии каждой хромосомы. Эти хромосомы уже реплицированы и имеют две сестринские хроматиды, которые затем разделяются во время второго деления мейоза. [4] Оба этих цикла деления клеток используются в процессе полового размножения в определенный момент их жизненного цикла. Считается, что оба они присутствуют у последнего общего предка эукариот.
Прокариоты ( бактерии и археи ) обычно подвергаются вегетативному делению клеток, известному как бинарное деление , при котором их генетический материал разделяется поровну на две дочерние клетки, но наблюдаются и альтернативные способы деления, такие как почкование . Всем делениям клеток, независимо от организма, предшествует один раунд репликации ДНК.
Для простых одноклеточных микроорганизмов, таких как амеба , одно деление клетки эквивалентно размножению совершенно новый организм — создается . В более широком масштабе митотическое деление клеток может создавать потомство от многоклеточных организмов , таких как растения , вырастающие из черенков. Митотическое деление клеток позволяет организмам , размножающимся половым путем, развиваться из одноклеточной зиготы , которая сама образуется путем слияния двух гамет , каждая из которых образуется в результате мейотического деления клеток. [5] [6] После роста от зиготы к взрослой особи деление клеток путем митоза позволяет непрерывно строить и восстанавливать организм. [7] В организме человека происходит около 10 квадриллионов клеточных делений за жизнь . [8]
Основной задачей клеточного деления является поддержание исходного генома клетки . Прежде чем произойдет деление, геномная информация, хранящаяся в хромосомах, должна быть реплицирована, а дублированный геном должен быть четко разделен между клетками-потомками. [9] значительная часть клеточной инфраструктуры . В обеспечении согласованности геномной информации между поколениями задействована [10] [11] [12]
У бактерий
[ редактировать ]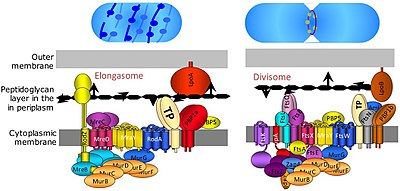
Деление бактериальных клеток происходит путем бинарного деления , а иногда и путем почкования . Дивисома представляет собой белковый комплекс бактерий, который отвечает за деление клеток, сужение внутренних и внешних мембран во время деления и ремоделирование пептидогликановой клеточной стенки в месте деления. Тубулиноподобный белок FtsZ играет решающую роль в формировании сократительного кольца для деления клеток. [14]
У эукариотов
[ редактировать ]Деление клеток у эукариот происходит сложнее, чем у прокариот. Если число хромосом уменьшено, деление эукариотических клеток классифицируется как мейоз (редукционное деление). Если число хромосом не уменьшается, деление эукариотической клетки классифицируется как митоз (эквационное деление). Существует также примитивная форма деления клеток, называемая амитозом . Амитотические или митотические деления клеток более атипичны и разнообразны у различных групп организмов, таких как протисты (а именно диатомеи , динофлагелляты и др.) и грибы . [ нужна ссылка ]
 закрыто
закрыто
внутриядерный
плевромитоз закрыто
закрыто
внеядерный
плевромитоз закрыто
закрыто
ортомитоз полуоткрытый
полуоткрытый
плевромитоз полуоткрытый
полуоткрытый
ортомитоз открыть
открыть
ортомитоз
В митотической метафазе (см. ниже) обычно хромосомы (каждая из которых содержит 2 сестринские хроматиды, которые развились во время репликации в S-фазе интерфазы) выравниваются на метафазной пластинке. Затем сестринские хроматиды расщепляются и распределяются между двумя дочерними клетками. [ нужна ссылка ]
В мейозе I гомологичные хромосомы спариваются, а затем разделяются и распределяются между двумя дочерними клетками. С другой стороны, мейоз II похож на митоз. Хроматиды . разделяются и распределяются одинаково У человека, других высших животных и многих других организмов процесс мейоза называется гаметическим мейозом , во время которого мейоз производит четыре гаметы. Тогда как у ряда других групп организмов, особенно у растений (наблюдаемых во время мейоза у низших растений, но на рудиментарной стадии у высших растений), мейоз дает начало спорам , которые прорастают в гаплоидную вегетативную фазу (гаметофит). Такой вид мейоза называется споровым мейозом. [ нужна ссылка ]
Фазы деления эукариотических клеток
[ редактировать ]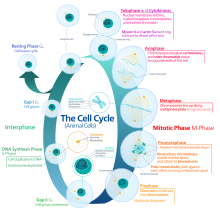
Интерфаза
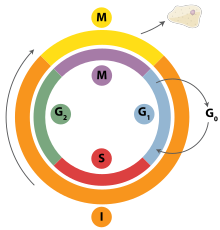
[ редактировать ]Интерфаза — это процесс, который клетка должна пройти перед митозом, мейозом и цитокинезом . [15] основных фаз: G1 трех , S и G2 Интерфаза состоит из . G 1 — это время роста клетки, когда происходят специализированные клеточные функции, необходимые для подготовки клетки к репликации ДНК. [16] Во время интерфазы существуют контрольные точки, которые позволяют клетке либо продвигаться вперед, либо останавливать дальнейшее развитие. Одна из контрольных точек находится между G 1 и S. Целью этой контрольной точки является проверка соответствующего размера клеток и любого повреждения ДНК . Вторая контрольная точка находится в фазе G 2. Эта контрольная точка также проверяет размер клеток, а также репликацию ДНК. Последняя контрольная точка расположена в месте метафазы, где она проверяет правильность соединения хромосом с митотическим веретеном. [17] В фазе S хромосомы реплицируются для сохранения генетического содержания. [18] Во время G 2 клетка проходит заключительные стадии роста, прежде чем она войдет в фазу М, где веретена синтезируются . Фаза М может быть либо митозом, либо мейозом в зависимости от типа клетки. Зародышевые клетки , или гаметы, подвергаются мейозу, а соматические клетки - митозу. После того, как клетка успешно проходит фазу М, она может подвергнуться клеточному делению посредством цитокинеза. Контроль каждой контрольной точки контролируется циклином и циклин-зависимыми киназами . Прогрессирование интерфазы является результатом увеличения количества циклина. По мере увеличения количества циклина все больше и больше циклин-зависимых киназ прикрепляются к циклину, сигнализируя клетке о дальнейшем переходе в интерфазу. На пике циклина, прикрепленного к циклин-зависимым киназам, эта система выталкивает клетку из интерфазы в фазу М, где происходят митоз, мейоз и цитокинез. [19] Перед входом в фазу М ячейка должна пройти три контрольных точки перехода. Наиболее важным из них является контрольная точка перехода G 1 -S. Если ячейка не проходит эту контрольную точку, это приводит к выходу ячейки из клеточного цикла. [20]
Профаза
[ редактировать ]Профаза – первая стадия деления. На этой стадии ядерная оболочка начинает разрушаться, длинные нити хроматина конденсируются, образуя более короткие и видимые нити, называемые хромосомами, ядрышко исчезает, и митотическое веретено начинает собираться из двух центросом. [21] Микротрубочки, связанные с выравниванием и разделением хромосом, называются веретеном и волокнами веретена. Хромосомы также будут видны под микроскопом и будут соединены в центромере. Во время этого периода конденсации и выравнивания в мейозе гомологичные хромосомы подвергаются разрыву своей двухцепочечной ДНК в тех же местах, за которым следует рекомбинация теперь фрагментированных цепей родительской ДНК в неродительские комбинации, известные как кроссинговер. [22] Доказано, что этот процесс в значительной степени вызван высококонсервативным белком Spo11 посредством механизма, аналогичного тому, который наблюдается с топоизомеразой при репликации и транскрипции ДНК. [23]
Прометафаза
[ редактировать ]Прометафаза – вторая стадия деления клеток. Эта стадия начинается с полного разрушения ядерной оболочки, в результате чего в цитоплазму открываются различные структуры. Этот разрыв затем позволяет веретенообразному аппарату, растущему из центросомы , прикрепиться к кинетохорам сестринских хроматид. Стабильное прикрепление веретенообразного аппарата к кинетохорам сестринских хроматид обеспечит безошибочную сегрегацию хромосом во время анафазы. [24] Прометафаза следует за профазой и предшествует метафазе.
Метафаза
[ редактировать ]В метафазе центромеры хромосом выравниваются на метафазной пластинке (или экваториальной пластинке ), воображаемой линии, которая находится на равных расстояниях от двух полюсов центросомы и удерживается вместе комплексами, известными как когезины . Хромосомы выстраиваются в середине клетки с помощью центров организации микротрубочек (MTOC), которые толкают и тянут центромеры обеих хроматид, тем самым заставляя хромосому двигаться к центру. На данный момент хромосомы все еще конденсируются и в настоящее время находятся в одном шаге от того, чтобы стать максимально свернутыми и конденсированными, а волокна веретена уже соединились с кинетохорами. [25] Во время этой фазы все микротрубочки, за исключением кинетохор, находятся в состоянии нестабильности, что способствует их переходу в анафазу. [26] В этот момент хромосомы готовы разделиться на противоположные полюса клетки по направлению к веретену, к которому они подключены. [27]
Анафаза
[ редактировать ]Анафаза — это очень короткая стадия клеточного цикла, которая возникает после выравнивания хромосом в митотической пластинке. Кинетохоры излучают сигналы ингибирования анафазы до тех пор, пока они не прикрепятся к митотическому веретену. Как только последняя хромосома правильно выровнена и прикреплена, последний сигнал рассеивается и вызывает резкий переход в анафазу. [26] Этот резкий сдвиг вызван активацией комплекса, способствующего анафазе , и его функцией по маркировке деградации белков, важных для перехода метафаза-анафаза. Одним из этих расщепляемых белков является секурин , который в результате своего распада высвобождает фермент сепаразу , которая расщепляет когезиновые кольца, удерживающие вместе сестринские хроматиды, что приводит к разделению хромосом. [28] После того как хромосомы выстроятся в середине клетки, волокна веретена раздвинут их. Хромосомы расщепляются, а сестринские хроматиды движутся к противоположным сторонам клетки. [29] Когда сестринские хроматиды раздвигаются, клетка и плазма удлиняются за счет некинетохорных микротрубочек. [30] Кроме того, на этой фазе активация комплекса, способствующего анафазе, посредством ассоциации с Cdh-1 начинает деградацию митотических циклинов. [31]
Телофаза
[ редактировать ]Телофаза — последняя стадия клеточного цикла, на которой борозда расщепления разделяет цитоплазму клетки (цитокинез) и хроматин. Это происходит за счет синтеза новой ядерной оболочки, которая формируется вокруг хроматина, собранного на каждом полюсе. Ядрышко реформируется, когда хроматин возвращается в рыхлое состояние, в котором он находился во время интерфазы. [32] [33] Деление клеточного содержимого не всегда одинаковое и может варьироваться в зависимости от типа клеток, как это видно на примере образования ооцитов, когда одна из четырех дочерних клеток обладает большей частью утенка. [34]
Цитокинез
[ редактировать ]Последней стадией процесса деления клеток является цитокинез . На этой стадии происходит деление цитоплазмы, которое происходит в конце митоза или мейоза. На этом этапе происходит необратимое разделение, приводящее к образованию двух дочерних клеток. Деление клетки играет важную роль в определении судьбы клетки. Это связано с возможностью асимметричного разделения. В результате это приводит к цитокинезу, в результате которого образуются неравные дочерние клетки, содержащие совершенно разные количества или концентрации молекул, определяющих судьбу. [35]
У животных цитокинез заканчивается образованием сократительного кольца и последующим его расщеплением. Но у растений происходит иначе. Сначала формируется клеточная пластинка, а затем между двумя дочерними клетками развивается клеточная стенка. [36]
У делящихся дрожжей ( S. pombe ) цитокинез происходит в фазе G1. [37]
Варианты
[ редактировать ]
Клетки подразделяются на две основные категории: простые безъядерные прокариотические клетки и сложные ядросодержащие эукариотические клетки. Из-за структурных различий эукариотические и прокариотические клетки делятся неодинаково. Кроме того, механизм деления клеток, который превращает эукариотические стволовые клетки в гаметы ( сперматозоиды у мужчин или яйцеклетки у женщин), называемый мейозом, отличается от процесса деления соматических клеток в организме.
В 2022 году ученые обнаружили новый тип клеточного деления, названный асинтетическим делением, обнаруженный в плоских эпителиальных клетках эпидермиса молодых рыбок данио. Когда молодь рыбок данио растет, клетки кожи должны быстро покрывать быстро увеличивающуюся площадь поверхности рыбок данио. Эти клетки кожи делятся без дублирования своей ДНК (S-фаза митоза), в результате чего до 50% клеток имеют уменьшенный размер генома. Эти клетки позже заменяются клетками со стандартным количеством ДНК. Ученые ожидают обнаружить этот тип деления и у других позвоночных. [39]
Восстановление повреждений ДНК в клеточном цикле
[ редактировать ]Повреждения ДНК обнаруживаются и устраняются на различных этапах клеточного цикла. Контрольные точки G1/S, контрольные точки G2/M и контрольные точки между метафазой и анафазой контролируют повреждение ДНК и останавливают деление клеток путем ингибирования различных комплексов циклин-CDK. Белок -супрессор опухолей p53 играет решающую роль в контрольных точках G1/S и контрольных точках G2/M. Активированные белки р53 приводят к экспрессии многих белков, которые важны для остановки, восстановления и апоптоза клеточного цикла. В контрольной точке G1/S р53 гарантирует, что клетка готова к репликации ДНК, тогда как в контрольной точке G2/M р53 гарантирует, что клетки правильно продублировали свое содержимое перед вступлением в митоз. [40]
В частности, при наличии повреждения ДНК киназы ATM и ATR , активируя различные киназы контрольных точек. активируются [41] Эти киназы контрольных точек фосфорилируют р53, что стимулирует выработку различных ферментов, связанных с репарацией ДНК. [42] Активированный р53 также активирует р21 , который ингибирует различные комплексы циклин-cdk. Эти комплексы циклин-cdk фосфорилируют белок ретинобластомы (Rb) , супрессор опухоли, связанный с семейством транскрипционных факторов E2F. Связывание этого белка Rb гарантирует, что клетки не перейдут в S-фазу преждевременно; однако, если он не сможет фосфорилироваться этими комплексами циклин-cdk, белок останется, и клетка остановится в фазе G1 клеточного цикла. [43]
Если ДНК повреждена, клетка также может изменить путь Akt, при котором BAD фосфорилируется и диссоциируется от Bcl2, тем самым ингибируя апоптоз. Если этот путь изменен мутацией потери функции в Akt или Bcl2, то клетка с поврежденной ДНК будет вынуждена подвергнуться апоптозу. [44] Если повреждение ДНК не может быть восстановлено, активированный р53 может вызвать гибель клеток путем апоптоза . Это можно сделать путем активации модулятора апоптоза с повышенной регуляцией р53 (PUMA) . PUMA представляет собой проапоптотический белок, который быстро индуцирует апоптоз путем ингибирования антиапоптотических членов семейства Bcl-2 . [45]
Деградация
[ редактировать ]Многоклеточные организмы заменяют изношенные клетки путем клеточного деления. Однако у некоторых животных деление клеток со временем прекращается. У людей это происходит в среднем после 52 делений, известных как предел Хейфлика . Клетка тогда называется стареющей . клеток С каждым делением теломеры , защитные последовательности ДНК на конце хромосомы , которые предотвращают деградацию хромосомной ДНК, укорачиваются . Это сокращение коррелирует с такими негативными последствиями, как возрастные заболевания и сокращение продолжительности жизни людей. [46] [47] С другой стороны, считается, что раковые клетки не деградируют таким образом, если вообще деградируют. Ферментный . комплекс под названием теломераза , присутствующий в больших количествах в раковых клетках, восстанавливает теломеры посредством синтеза теломерных повторов ДНК, позволяя делению продолжаться бесконечно [48]
История
[ редактировать ]
Деление клеток под микроскопом было впервые обнаружено немецким ботаником Гуго фон Молем в 1835 году, когда он работал над зеленой водорослью Cladophora glomerata . [49]
В 1943 году впервые было снято на видео деление клеток. [50] Курт Мишель с использованием фазово-контрастного микроскопа . [51]
См. также
[ редактировать ]- Слияние клеток
- Рост клеток
- Циклинзависимая киназа
- Лабильные клетки – клетки, которые постоянно делятся.
- Митотическая катастрофа
Ссылки
[ редактировать ]- ^ Мартин Э.А., Хайн Р. (2020). Биологический словарь (6-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 9780199204625 . OCLC 176818780 .
- ^ Гриффитс Эй Джей (2012). Введение в генетический анализ (10-е изд.). WH Freeman and Co. Нью-Йорк: ISBN 9781429229432 . OCLC 698085201 .
- ^ «10.2 Клеточный цикл – Биология 2e | OpenStax» . openstax.org . 28 марта 2018 года . Проверено 24 ноября 2020 г.
- ^ Гилберт, Скотт Ф. (2000), «Мейоз» , Биология развития. 6-е издание , Sinauer Associates , получено 8 сентября 2023 г.
- ^ Гилберт С.Ф. (2000). «Сперматогенез» . Биология развития (6-е изд.). Синауэр Ассошиэйтс.
- ^ Гилберт С.Ф. (2000). «Оогенез» . Биология развития (6-е изд.). Синауэр Ассошиэйтс.
- ^ Матон, Антея (1997). Клетки: строительные блоки жизни (3-е изд.). Река Аппер-Сэддл, Нью-Джерси: Прентис-Холл. стр. 70–74 . ISBN 978-0134234762 . ОСЛК 37049921 .
- ^ Кваммен Д. (апрель 2008 г.). «Заразный рак» . Журнал Харпера . ISSN 0017-789X . Проверено 14 апреля 2019 г.
- ^ Голицын Юрий Н.; Крылов, Михаил СС (2010). Деление клеток: теория, варианты и деградация . Нью-Йорк: Издательство Nova Science. п. 137. ИСБН 9781611225938 . OCLC 669515286 .
- ^ Флетчер, Дэниел А.; Маллинз, Р. Дайч (28 января 2010 г.). «Механика клетки и цитоскелет» . Природа . 463 (7280): 485–492. Бибкод : 2010Natur.463..485F . дои : 10.1038/nature08908 . ISSN 0028-0836 . ПМЦ 2851742 . ПМИД 20110992 .
- ^ Ли, Шаньвэй; Солнце, Тяньтянь; Рен, Хайюнь (27 апреля 2015 г.). «Функции цитоскелета и связанных с ним белков при митозе и цитокинезе в растительных клетках» . Границы в науке о растениях . 6 : 282. дои : 10.3389/fpls.2015.00282 . ISSN 1664-462X . ПМК 4410512 . ПМИД 25964792 .
- ^ Хоманн, Тим; Дегани, Фарамарз (18 апреля 2019 г.). «Цитоскелет — сложная взаимодействующая сеть» . Клетки . 8 (4): 362. doi : 10.3390/cells8040362 . ISSN 2073-4409 . ПМК 6523135 . ПМИД 31003495 .
- ^ Хьюгонне Ж.Э., Менжен-Лекрелькс Д., Монтон А., ден Блаувен Т., Карбоннелле Е., Векерле С. и др. (октябрь 2016 г.). «Эшерихия коли» . электронная жизнь . 5 . дои : 10.7554/elife.19469 . ПМК 5089857 . ПМИД 27767957 .
- ^ Отделение ячеек: Цикл кольца, Лоуренс Ротфилд и Шерил Джастис, CELL , DOI
- ^ Мариб Э.Н. (2000). Основы анатомии и физиологии человека (6-е изд.). Сан-Франциско: Бенджамин Каммингс. ISBN 978-0805349405 . OCLC 41266267 .
- ^ Парди AB (ноябрь 1989 г.). «События G1 и регуляция пролиферации клеток». Наука . 246 (4930): 603–8. Бибкод : 1989Sci...246..603P . дои : 10.1126/science.2683075 . ПМИД 2683075 .
- ^ Молинари М. (октябрь 2000 г.). «Контрольные точки клеточного цикла и их инактивация при раке человека» . Пролиферация клеток . 33 (5): 261–74. дои : 10.1046/j.1365-2184.2000.00191.x . ПМК 6496592 . ПМИД 11063129 .
- ^ Морган Д.О. (2007). Клеточный цикл: принципы управления . Лондон: New Science Press. ISBN 9780199206100 . ОСЛК 70173205 .
- ^ Линдквист А., ван Зон В., Карлссон Розенталь С., Волтуис Р.М. (май 2007 г.). «Активация циклина B1-Cdk1 продолжается после разделения центросом, чтобы контролировать митотическое прогрессирование» . ПЛОС Биология . 5 (5): е123. doi : 10.1371/journal.pbio.0050123 . ПМК 1858714 . ПМИД 17472438 .
- ^ Паулович А.Г., Точиски Д.П., Хартвелл Л.Х. (февраль 1997 г.). «Когда контрольно-пропускные пункты выходят из строя» . Клетка . 88 (3): 315–21. doi : 10.1016/S0092-8674(00)81870-X . ПМИД 9039258 . S2CID 5530166 .
- ^ Шермелле Л., Карлтон П.М., Хаасе С., Шао Л., Виното Л., Кнер П. и др. (июнь 2008 г.). «Субдифракционная многоцветная визуализация ядерной периферии с помощью трехмерной микроскопии со структурированным освещением» . Наука . 320 (5881): 1332–6. Бибкод : 2008Sci...320.1332S . дои : 10.1126/science.1156947 . ПМЦ 2916659 . ПМИД 18535242 .
- ^ Левонтин Р.К., Миллер Дж.Х., Гелбарт В.М., Гриффитс А.Дж. (1999). «Механизм кроссинговера» . Современный генетический анализ .
- ^ Кини С. (2001). Механизм и контроль инициации мейотической рекомбинации . Текущие темы биологии развития. Том. 52. Эльзевир. стр. 1–53 . дои : 10.1016/s0070-2153(01)52008-6 . ISBN 9780121531522 . ПМИД 11529427 .
- ^ «Прометафаза – обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 21 ноября 2023 г.
- ^ «Исследователи пролили свет на сокращение хромосом» . ScienceDaily . Проверено 14 апреля 2019 г.
- ^ Перейти обратно: а б Уолтер П., Робертс К., Рафф М., Льюис Дж., Джонсон А., Альбертс Б. (2002). "Митоз". Молекулярная биология клетки (4-е изд.). Гирляндная наука.
- ^ Элрод С (2010). Очерки Шаума: генетика (5-е изд.). Нью-Йорк: Макгроу-Хилл. п. 8. ISBN 9780071625036 . OCLC 473440643 .
- ^ Брукер А.С., Берковиц К.М. (2014). «Роль когезинов в митозе, мейозе, здоровье и заболеваниях человека». Контроль клеточного цикла . Методы молекулярной биологии. Том. 1170. Нью-Йорк: Спрингер. стр. 229–66. дои : 10.1007/978-1-4939-0888-2_11 . ISBN 9781493908875 . ПМЦ 4495907 . ПМИД 24906316 .
- ^ «Клеточный цикл» . www.biology-pages.info . Проверено 14 апреля 2019 г.
- ^ Урри Л.А., Кейн М.Л., Джексон Р.Б., Вассерман С.А., Минорский П.В., Рис Дж.Б. (2014). Кэмпбелл Биология в фокусе . Бостон (Массачусетс): Пирсон. ISBN 978-0-321-81380-0 .
- ^ Барфорд, Дэвид (12 декабря 2011 г.). «Структурное понимание сложных функций и механизмов, способствующих анафазе» . Философские труды Королевского общества B: Биологические науки . 366 (1584): 3605–3624. дои : 10.1098/rstb.2011.0069 . ПМК 3203452 . ПМИД 22084387 .
- ^ Деккер Дж. (25 ноября 2014 г.). «Два способа сворачивания генома во время клеточного цикла: данные, полученные с помощью захвата конформации хромосом» . Эпигенетика и хроматин . 7 (1): 25. дои : 10.1186/1756-8935-7-25 . ПМК 4247682 . ПМИД 25435919 .
- ^ Hetzer MW (март 2010 г.). «Ядерная оболочка» . Перспективы Колд-Спринг-Харбор в биологии . 2 (3): а000539. doi : 10.1101/cshperspect.a000539 . ПМК 2829960 . ПМИД 20300205 .
- ^ Гилберт С.Ф. (2000). «Оогенез» . Биология развития (6-е изд.). Синауэр Ассошиэйтс.
- ^ Гертин Д.А., Траутманн С., МакКоллум Д. (июнь 2002 г.). «Цитокинез у эукариот» . Обзоры микробиологии и молекулярной биологии . 66 (2): 155–78. дои : 10.1128/MMBR.66.2.155-178.2002 . ПМК 120788 . ПМИД 12040122 .
- ^ Смит, Лори Дж. (декабрь 1999 г.). «Разделяй и властвуй: цитокинез в растительных клетках» . Современное мнение в области биологии растений . 2 (6): 447–453. дои : 10.1016/S1369-5266(99)00022-9 .
- ^ Ячейка, GM Купер; изд 2 Книжная полка NCBI , Эукариотический клеточный цикл, Рисунок 14.7
- ^ «Фазовая голографическая визуализация деления клеток » . Интернет-архив . Архивировано из оригинала 29 июня 2013 года.
- ^ Чан К.Ю., Ян К.С., Роан Х.И., Сюй С.К., Ценг Т.Л., Сяо К.Д. и др. (апрель 2022 г.). «Клетки кожи подвергаются асинтетическому делению, что приводит к расширению поверхности тела рыбок данио». Природа . 605 (7908): 119–125. Бибкод : 2022Natur.605..119C . дои : 10.1038/s41586-022-04641-0 . ПМИД 35477758 . S2CID 248416916 .
- ^ Шентюрк, Эмир; Манфреди, Джеймс Дж. (2013). «Р53 и эффекты клеточного цикла после повреждения ДНК». Протоколы P53 . Методы молекулярной биологии (Клифтон, Нью-Джерси). Том. 962. стр. 49–61. дои : 10.1007/978-1-62703-236-0_4 . ISBN 978-1-62703-235-3 . ISSN 1064-3745 . ПМЦ 4712920 . ПМИД 23150436 .
- ^ Дин, Лей; Цао, Цзяци; Линь, Вэнь; Чен, Хунцзянь; Сюн, Сяньхуэй; Ао, Хуншунь; Ю, Мин; Линь, Цзе; Цуй, Цинхуа (13 марта 2020 г.). «Роль циклин-зависимых киназ в развитии клеточного цикла и терапевтические стратегии при раке молочной железы человека» . Международный журнал молекулярных наук . 21 (6): 1960. doi : 10.3390/ijms21061960 . ISSN 1422-0067 . ПМК 7139603 . ПМИД 32183020 .
- ^ Уильямс, Эшли Б.; Шумахер, Бьорн (2016). «Р53 в процессе восстановления повреждений ДНК» . Перспективы Колд-Спринг-Харбора в медицине . 6 (5): а026070. doi : 10.1101/cshperspect.a026070 . ISSN 2157-1422 . ПМЦ 4852800 . ПМИД 27048304 .
- ^ Энгеланд, Курт (2022). «Регуляция клеточного цикла: передача сигналов p53-p21-RB» . Смерть клеток и дифференциация . 29 (5): 946–960. дои : 10.1038/s41418-022-00988-z . ISSN 1476-5403 . ПМК 9090780 . ПМИД 35361964 .
- ^ Руволо, ПП; Дэн, X.; Мэй, WS (2001). «Фосфорилирование Bcl2 и регуляция апоптоза» . Лейкемия . 15 (4): 515–522. дои : 10.1038/sj.leu.2402090 . ISSN 1476-5551 . ПМИД 11368354 . S2CID 2079715 .
- ^ Джаббур, AM; Эро, JE; Даунт, CP; Кауфманн, Т.; Сандов, Дж.; О'Рейли, Луизиана; Каллус, бакалавр; Лопес, А.; Штрассер, А.; Во, ДЛ; Экерт, П.Г. (2009). «Puma косвенно активирует Bax, вызывая апоптоз в отсутствие Bid или Bim» . Смерть клеток и дифференциация . 16 (4): 555–563. дои : 10.1038/cdd.2008.179 . ISSN 1476-5403 . ПМИД 19079139 .
- ^ Цзян Х., Шиффер Э., Сонг З., Ван Дж., Цюрбиг П., Тедик К. и др. (август 2008 г.). «Белки, вызванные дисфункцией теломер и повреждением ДНК, представляют собой биомаркеры старения и болезней человека» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (32): 11299–304. Бибкод : 2008PNAS..10511299J . дои : 10.1073/pnas.0801457105 . ПМК 2516278 . ПМИД 18695223 .
- ^ Коутон Р.М., Смит К.Р., О'Брайен Э., Сиваченко А., Кербер Р.А. (февраль 2003 г.). «Связь между длиной теломер в крови и смертностью у людей в возрасте 60 лет и старше». Ланцет . 361 (9355): 393–5. дои : 10.1016/S0140-6736(03)12384-7 . ПМИД 12573379 . S2CID 38437955 .
- ^ Джафри М.А., Ансари С.А., Алькахтани М.Х., Шей Дж.В. (июнь 2016 г.). «Роль теломер и теломеразы при раке, а также достижения в терапии, нацеленной на теломеразу» . Геномная медицина . 8 (1): 69. дои : 10.1186/s13073-016-0324-x . ПМЦ 4915101 . ПМИД 27323951 .
- ^ Биография, нем. «Моль, Гуго фон – немецкая биография» . www.deutsche-biography.de (на немецком языке) . Проверено 15 апреля 2019 г.
- ^ Магистр БР (15 декабря 2008 г.). «История оптического микроскопа в клеточной биологии и медицине». Энциклопедия наук о жизни . John Wiley & Sons, Ltd. doi : 10.1002/9780470015902.a0003082 . ISBN 978-0470016176 .
- ^ ZEISS Microscope (1 июня 2013 г.), исторический замедленный фильм доктора Курта Мишеля, Carl Zeiss Jena (ок. 1943 г.) , заархивировано из оригинала 07 ноября 2021 г. , получено 15 апреля 2019 г.
Дальнейшее чтение
[ редактировать ]- Морган Х.И. (2007). «Клеточный цикл: принципы контроля» Лондон: New Science Press.
- Дж. М. Превратите плод в человека (1978, 1989). Издательство Гарвардского университета. ISBN 0-674-30692-9
- Деление клетки: бинарное деление и митоз.
- Макдугал, В. Скотт и др. Обзор одиннадцатого издания Кэмпбелла-Уолша по урологии . Эльзевир, 2016.
- Раздел « Митоз и контроль клеточного цикла» из « Веховых статей по клеточной биологии» (Галл Дж.Г., Макинтош Дж.Р., ред.) содержит комментарии и ссылки на основополагающие исследовательские работы по митозу и делению клеток. Опубликовано онлайн в Библиотеке изображений и видео Американского общества клеточной биологии.
- Библиотека изображений и видео, заархивированная 10 июня 2011 г. в Wayback Machine Американского общества клеточной биологии, содержит множество видеороликов, показывающих деление клеток.
- Клеточный отдел библиотеки изображений клеток
- Изображения : Каланте обесцвечивает Линдл. – Тайный цветочный сад Флавона
- Модель деления клеток Тайсона и описание в базе данных BioModels
- WormWeb.org: Интерактивная визуализация клеточного C. elegans происхождения – визуализируйте весь набор клеточных делений нематоды C. elegans.
| Базы данных органов управления : Национальные |
|---|