Циклинзависимый киназный комплекс
Комплекс циклин-зависимой киназы ( CDKC , циклин-CDK ) представляет собой белковый комплекс, образованный ассоциацией неактивной каталитической субъединицы протеинкиназы, циклин-зависимой киназы (CDK), с регуляторной субъединицей циклином . [ 1 ] Как только циклин-зависимые киназы связываются с циклином, образовавшийся комплекс находится в активированном состоянии. Субстратная специфичность активированного комплекса в основном определяется связанным с ним циклином внутри комплекса. Активность CDKC контролируется фосфорилированием белков-мишеней, а также связыванием ингибирующих белков. [ 2 ]
Структура и регулирование
[ редактировать ]
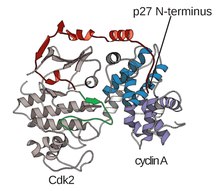
Структура CDK в комплексе с субъединицами циклина (CDKC) уже давно является целью структурных и клеточных биологов, начиная с 1990-х годов, когда Браун и др. определили структуру несвязанного циклина А. и в том же году Джеффри и др. расшифровал структуру комплекса циклин A-CDK2 человека с разрешением 2,3 ангстрема. [ 3 ] С тех пор многие структуры CDK были определены с более высоким разрешением, включая структуры CDK2 и CDK2, связанные с различными субстратами, как показано на рисунке 1. Всего в Банке данных белков существуют структуры высокого разрешения примерно для 25 комплексов CDK-циклин . [ 4 ] В зависимости от функции существует две основные популяции комплексных структур CDK-циклин: открытая и закрытая форма. Разница между формами заключается в связывании партнеров циклина, где комплексы закрытой формы связывают CDK-циклин как на C, так и на N-концах петли активации CDK, тогда как партнеры открытой формы связываются только на N-конце. Структуры открытой формы чаще всего соответствуют комплексам, участвующим в регуляции транскрипции (CDK 8, 9, 12 и 13), тогда как комплекс CDK-циклин закрытой формы чаще всего участвует в развитии и регуляции клеточного цикла (CDK 1, 2, 6). . Однако эти отдельные роли существенно не различаются в зависимости от гомологии последовательностей между компонентами CDK. В частности, среди этих известных структур, по-видимому, есть четыре основных консервативных участка: N-концевая петля, богатая глицином, шарнирная область, αC-спираль и сайт регуляции Т-петли. [ 4 ]
Цикл активации
[ редактировать ]Петля активации , также называемая Т-петлей, представляет собой область CDK (между мотивами DFG и APE во многих CDK). [ 4 ] который ферментативно активен, когда CDK связан со своим специфичным для функции партнером. В комплексах CDK-циклин эта область активации состоит из консервативной спирали αL-12 и содержит ключевой фосфорилируемый остаток (обычно треонин для партнеров CDK-циклина, но также включает серин и тирозин), который опосредует ферментативную активность CDK. Именно по этому важному остатку (T160 в комплексах CDK2, T177 в комплексах CDK6) происходит ферментативное АТФ-фосфорилирование комплексов CDK-циклин с помощью CAK (циклин-активирующей киназы, относящейся к комплексу CDK7-циклин H в клетках человека). После гидролиза АТФ до фосфорилирования в этом сайте эти комплексы способны выполнить свою намеченную функцию — фосфорилирование клеточных мишеней. Важно отметить, что в CDK 1, 2 и 6 Т-петля и отдельная С-концевая область являются основными сайтами связывания циклинов в CDK, и то, какие циклины связываются с каждым из этих CDK, опосредовано конкретная последовательность сайта активации Т-петли. Эти сайты связывания циклина являются областями наибольшей вариабельности в CDK, несмотря на относительно высокую гомологию последовательностей, окружающих мотив спирали αL-12 этого структурного компонента. [ 4 ]
Богатый глицином регион
[ редактировать ]Петля , богатая глицином (петля, богатая глицином), как видно из остатков 12-16 в CDK2, кодирует консервативный мотив GXGXXG как на дрожжевых, так и на животных моделях. Регуляторная область подвергается дифференциальному фосфорилированию по неглициновым остаткам в этом мотиве, что делает этот сайт подверженным ингибирующему фосфорилированию киназы Wee1 и/или Myt1 и Cdc25 дефосфорилированию у млекопитающих. Это обратимое фосфорилирование Gly-богатой петли в CDK2 происходит на Y15, активность которого дополнительно изучалась. Исследование этого остатка показало, что фосфорилирование способствует конформационному изменению, которое предотвращает связывание АТФ и субстрата за счет стерического вмешательства в эти необходимые сайты связывания в петле активации комплексов CDK-циклин. Этой активности способствует заметная гибкость, которую Gly-богатая петля имеет в структуре большинства CDK, что позволяет ее вращению в сторону активационной петли оказывать значительное влияние на снижение сродства к субстрату без серьезных изменений в общей структуре комплекса CDK-циклин. [ 3 ] [ 5 ]
Шарнирная область
[ редактировать ]Консервативная шарнирная область CDK в эукариотических клетках действует как важный мост между петлей, богатой Gly, и петлей активации. CDK характеризуются N-концевой долей, которая в основном представляет собой скрученный бета-лист, соединенный через эту шарнирную область с C-концевой долей, в которой преобладает альфа-спираль. При обсуждении Т-петли и Gly-rich петли важно отметить, что эти области, которые должны быть способны пространственно взаимодействовать, чтобы выполнять свои биохимические функции, лежат на противоположных долях самой CDK. Таким образом, эта шарнирная область, длина которой может незначительно различаться в зависимости от типа CDK и комплекса CDK-циклин, соединяет важные регуляторные области CDK, соединяя эти доли, и играет ключевую роль в получаемой структуре комплексов CDK-циклин, правильно ориентируя АТФ. для легкого катализа реакций фосфорилирования собранным комплексом. [ 3 ] [ 4 ]
αC-спираль
[ редактировать ]Область αC-спирали высококонсервативна во многих киномах млекопитающих (семейство киназ ). Его основная ответственность — поддержание аллостерического контроля активного центра киназы. Этот контроль проявляется в комплексах CDK-циклин путем специфического предотвращения активности CDK до тех пор, пока он не свяжется со своим партнерским регулятором (т.е. циклином или другим белком-партнером). Это связывание вызывает конформационное изменение в области αC-спирали CDK и позволяет ему переместиться из щели активного центра и завершает начальный процесс активации Т-петли. Учитывая, что эта область настолько консервативна во всем белковом суперсемействе киназ, было показано, что этот механизм, при котором αC-спираль сворачивается из N-концевой доли киназы, обеспечивает расширенный доступ к спирали αL-12, которая находится внутри Т-петля считается потенциальной мишенью для разработки лекарств. [ 6 ]
Клеточный цикл
[ редактировать ]Цикл дрожжевых клеток
[ редактировать ]Хотя эти комплексы выполняют разнообразные функции, CDKC наиболее известны своей ролью в клеточном цикле . Первоначально исследования проводились на Schizosaccharomyces pombe и Saccharomyces cerevisiae (дрожжи). S. pombe и S. cerevisiae наиболее известны своей ассоциацией с одним Cdk, Cdc2 и Cdc28 соответственно, которые образуют комплексы с несколькими различными циклинами. [ 7 ] В зависимости от циклина затрагиваются различные части клеточного цикла. Например, у S. pombe Cdc2 связывается с Cdk13, образуя комплекс Cdk13-Cdc2. У S. cerevisiae ассоциация Cdc28 с циклинами Cln1, Cln2 или Cln3 приводит к переходу от фазы G1 к фазе S. В S-фазе Cln1 и Cln2 диссоциируют с Cdc28, и образуются комплексы между Cdc28 и Clb5 или Clb6. В фазе G2 комплексы, образующиеся в результате ассоциации между Cdc28 и Clb1, Clb2, Clb3 или Clb4, приводят к переходу от G2 к фазе M ( фазы митотической). Эти комплексы присутствуют и в ранней М-фазе. [ 2 ] См. таблицу 1 для получения сводной информации о дрожжевых CDKC.
- Таблица 1. CDKC, связанные с фазами клеточного цикла у дрожжей
| СДК | Циклин | Фаза клеточного цикла |
|---|---|---|
| Cdc2 ( С. помб ) | Cdc13 | фазовый переход G2 в M; ранняя М-фаза |
| Cdc28 ( S. cerevisiae ) | Клн1, Клн2 | Фазовый переход G1 в S |
| Cdc28 | Клб5, Клб6 | S-фаза |
| Cdc28 | Клб1, Клб2, Клб3, Клб4 | фазовый переход G2 в M; ранняя М-фаза |
Из того, что известно о комплексах, образующихся на каждой фазе клеточного цикла у дрожжей, возникли предложенные модели, основанные на важных сайтах фосфорилирования и задействованных факторах транскрипции. [ 7 ] [ 8 ]
Клеточный цикл млекопитающих
[ редактировать ]Используя информацию, полученную в ходе исследований клеточного цикла дрожжей, был достигнут значительный прогресс в отношении клеточного цикла млекопитающих. Было установлено, что клеточные циклы схожи и CDKC прямо или косвенно влияют на течение клеточного цикла. Как упоминалось ранее, у дрожжей только одна циклинзависимая киназа (CDK) связана с несколькими различными циклинами. Однако в клетках млекопитающих несколько разных CDK связываются с различными циклинами, образуя CDKC. также известный как Cdc2 человека), первый идентифицированный человеческий CDK, ассоциируется с циклинами A или B. Например, Cdk1 ( Комплексы CyclinA/B-Cdk1 управляют переходом между фазой G2 и фазой M, а также ранней фазой M. Другой CDK млекопитающих, Cdk2, может образовывать комплексы с циклинами D1, D2, D3, E или A. Cdk4 и Cdk6 взаимодействуют с циклинами D1, D2 и D3. [ 9 ] Исследования показали, что нет никакой разницы между циклином D1-Cdk4/6 CDKC, поэтому любые уникальные свойства могут быть связаны со специфичностью или активацией субстрата. [ 1 ] Хотя уровни CDK остаются довольно постоянными на протяжении клеточного цикла, уровни циклина колеблются. Колебания контролируют активацию комплексов циклин-CDK и, в конечном итоге, прогрессирование на протяжении всего цикла. [ 10 ] В таблице 2 приведена сводная информация о CDKC клеток млекопитающих, участвующих в клеточном цикле.
- Таблица 2. CDKC, связанные с фазами клеточного цикла в клетках млекопитающих [ 4 ]
| СДК | Циклин | Фаза клеточного цикла | Нециклиновые партнерские белки |
|---|---|---|---|
| Кдк1 (Кдк2) | Циклины А и В | фазовый переход G2 в M; ранняя М-фаза | Cks1 и Cks2 |
| CDK2 | Циклины D1, D2, D3 | Фаза G1 | KAP, Cks1, p27KIP1 и Spy-1 |
| CDK2 | Циклин Е | Фазовый переход G1 в S | KAP, Cks1, p27KIP1 и Spy-1 |
| CDK2 | Циклин А | S-фаза | KAP, Cks1, p27KIP1 и Spy-1 |
| CDK4 | Циклины D1, D2, D3 | Фаза G1 | HSP90-Cdc37 |
| CDK6 | Циклины D1, D2, D3 | Фаза G1 | p16INK4A, p19INK4D и P18INK4C-циклин K |
| Cdk8 | Циклин С | --- | --- |
| CDK9 | Циклин Т | --- | Тат, AFF4 и TAR |
| Cdk12 | Циклин К | --- | --- |
| Cdk13 | Циклин К | --- | --- |

Переход фазы G 1 к S
[ редактировать ]Во время поздней фазы G1 CDKC связывают и фосфорилируют членов семейства белков ретинобластомы (Rb) . Члены семейства белков Rb являются супрессорами опухолей, которые предотвращают неконтролируемую пролиферацию клеток, которая может произойти во время образования опухоли. Однако считается, что pRbs также подавляют гены, необходимые для перехода от фазы G 1 к фазе S. Когда клетка готова перейти к следующей фазе, CDKC, циклин D1-Cdk4 и циклин D1-Cdk6 фосфорилируют pRB с последующим дополнительным фосфорилированием из циклина E-Cdk2 CDKC. [ 11 ] [ 12 ] Как только происходит фосфорилирование, транскрипционные факторы , которые необратимо инактивируют pRB, и наступает переход в S-фазу клеточного цикла. высвобождаются [ 13 ] Циклин E-Cdk2 CDKC, образующийся в фазе G1 , затем способствует инициации репликации ДНК во время S-фазы. [ 1 ]
G 2 к M Переход фазы
[ редактировать ]В конце фазы S циклин А связывается с Cdk1 и Cdk2. Во время фазы G2 циклин A разлагается, в то время как циклин B синтезируется и образуются комплексы циклин B-Cdk1. Комплексы циклин B-Cdk1 не только важны для перехода в фазу М, но и эти CDKC играют роль в следующих регуляторных и структурных процессах: [ 1 ]
- Хромосомная конденсация
- Фрагментация сети Гольджи
- Разрушение ядерной пластинки
Инактивация комплекса циклин B-Cdk1 посредством деградации циклина B необходима для выхода из М-фазы клеточного цикла. [ 1 ]
Другой
[ редактировать ]Несмотря на то, что большинство известных CDKC участвуют в клеточном цикле, не все киназные комплексы функционируют таким образом. Исследования показали, что другие CDKC, такие как циклин k-Cdk9 и циклин T1-Cdk9, участвуют в реакции репликационного стресса . [ 14 ] и влияют на транскрипцию . [ 15 ] [ 16 ] [ 17 ] Кроме того, комплексы циклин H-Cdk7 могут играть роль в мейозе мужских половых клеток. [ 18 ] и было показано, что он также участвует в транскрипционной деятельности. [ 1 ] [ 19 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Малумбрес М., Барбацид М. Циклинзависимые киназы млекопитающих. Тенденции биохимии. наук. 2005 ноябрь;30(11):630-41
- ^ Jump up to: а б Лодиш Х., Балтимор Д., Берк А., Зипурски С.Л., Мацудайра П., Дарнелл Дж. 1995. Молекулярная клеточная биология. 3-е изд. Нью-Йорк: Книги Scientific American
- ^ Jump up to: а б с Кристи Левин, Фредерик Р. Кросс, Структурирование биологии клеточного цикла, Структура, Том 3, Выпуск 11, 1995, Страницы 1131–1134, ISSN 0969–2126, два : 10.1016/S0969-2126(01)00248-9 .
- ^ Jump up to: а б с д и ж Вуд, DJ, и Эндикотт, JA (2018). Структурное понимание функционального разнообразия семейства CDK-циклинов. Открытая биология, 8(9), 180112.
- ^ Malumbres: Циклин-зависимые киназы. Геномная биология, 2014, 15:22, два : 10.1186/gb4184
- ^ Лоренцо Палмьери, Джулио Растелли, Смещение спирали αC как общий подход к аллостерической модуляции протеинкиназ, Drug Discovery Today, том 18, выпуски 7–8, 2013, страницы 407–414, ISSN 1359–6446, дои : 10.1016/j.drudis.2012.11.009 .
- ^ Jump up to: а б Саймон И, Барнетт Дж, Ханнетт Н, Харбисон Коннектикут, Ринальди Н.Дж., Волкерт Т.Л., Вайрик Дж.Дж., Зейтлингер Дж., Гиффорд Д.К., Яаккола Т.С., Янг Р.А. Серийная регуляция регуляторов транскрипции в клеточном цикле дрожжей. Клетка. 21 сентября 2001 г.; 106 (6): 697-708.
- ^ Барик Д., Бауманн В.Т., Пол М.Р., Новак Б., Тайсон Дж.Дж. Модель регуляции клеточного цикла дрожжей, основанная на многосайтовом фосфорилировании. Мол сист биол. 2010, 24 августа;6:405.
- ^ Малумбрес М., Барбацид М. Клеточный цикл, CDK и рак: меняющаяся парадигма. Нат Преподобный Рак. Март 2009 г.;9(3):153-66.
- ^ Вермюлен К., Ван Бокстеле Д.Р., Бернеман З.Н. Клеточный цикл: обзор регуляции, дерегуляции и терапевтических целей при раке. Сотовый Пролиф. Июнь 2003 г.;36(3):131-49.
- ^ Миттнахт С. Контроль фосфорилирования PRB. Curr Opin Genet Dev. 1998 февраль;8(1):21-7.
- ^ Кэлин В.Г. младший. Функции белка ретинобластомы. Биоэссе. 1999 ноября;21(11):950-8.
- ^ Лундберг А.С., Вайнберг Р.А. Функциональная инактивация белка ретинобластомы требует последовательной модификации по крайней мере двумя различными комплексами циклин-cdk. Мол Клеточная Биол. 1998 февраля;18(2):753-61.
- ^ Ю Д.С., Чжао Р., Сюй Э.Л., Кайер Дж., Йе Ф, Гуо Ю, Шир Ю, Кортес Д. Циклин-зависимая киназа 9-циклин K действует в ответной реакции на репликационный стресс. Представитель ЭМБО. 2010 ноября;11(11):876-82.
- ^ Фу Т.Дж., Пэн Дж., Ли Дж., Прайс Д.Х., Флорес О. Циклин K действует как регуляторная субъединица CDK9 и участвует в транскрипции РНК-полимеразы II. J Биол Хим. 3 декабря 1999 г.; 274 (49): 34527-30.
- ^ Ян З, Чжу К, Луо К, Чжоу К. Малая ядерная РНК 7SK ингибирует киназу CDK9/циклин Т1, контролируя транскрипцию. Природа. 15 ноября 2001 г.; 414 (6861): 317-22.
- ^ Ю Д.С., Кортес Д. Роль CDK9-циклина K в поддержании целостности генома. Клеточный цикл. 1 января 2011 г.; 10 (1): 28–32.
- ^ Ким Дж.М., Макгоги Дж.Т., Богл Р.К., Равник С.Е. Мейотическая экспрессия комплекса циклин H/Cdk7 в половых клетках самцов мыши. Биол Репрод. 2001 май;64(5):1400-8.
- ^ Патель С.А., Саймон MC. Функциональный анализ комплекса Cdk7.cyclin H.Mat1 в эмбриональных стволовых клетках и эмбрионах мыши. J Биол Хим. 14 мая 2010 г.; 285(20): 15587-98.