Гидрогеносома

Гидрогеносома обнаруженная — мембраной закрытая органелла, у некоторых анаэробных инфузорий , жгутиконосцев и грибов . Гидрогеносомы — это весьма изменчивые органеллы, которые, предположительно, развились из протомитохондрий для производства молекулярного водорода и АТФ в анаэробных условиях. [1]
Гидрогеносомы были открыты в 1973 г. Д.Г. Линдмарком и М. Мюллером. Поскольку гидрогеносомы имеют эволюционное значение для организмов, живущих в анаэробных или кислородно-дефицитных средах, многие исследовательские институты с тех пор документально задокументировали свои выводы о том, как органеллы различаются в различных источниках. [2]
История [ править ]
Гидрогеносомы были выделены, очищены, биохимически охарактеризованы и названы в начале 1970-х годов Линдмарком и Мюллером из Университета Рокфеллера. В дополнение к этому плодотворному исследованию гидрогеносом они также впервые продемонстрировали присутствие пируват:ферредоксиноксидоредуктазы и гидрогеназы у эукариот . [2] Впоследствии были проведены дальнейшие исследования биохимической цитологии и субклеточной организации нескольких анаэробных простейших паразитов (например, Trichomonas vaginalis , Tritrichomonas fetus , Giardia Lamblia и Entamoeba sp.) . [1]
Используя информацию, полученную в результате гидрогеносомных и биохимических цитологических исследований, эти исследователи определили механизм действия метронидазола (флагила). Сегодня метронидазол признан стандартным химиотерапевтическим средством для лечения анаэробных инфекций. [3] [4]
С момента открытия гидрогеносомы были обнаружены у множества анаэробных одноклеточных инфузорий, жгутиконосцев и грибов. Наиболее примечательным из них является паразитическая Trichomonas vaginalis . [5]
Описание [ править ]
Гидрогеносомы — это органеллы, которые, как предполагается, произошли от митохондрий и обеспечили другой механизм анаэробного синтеза АТФ с использованием пирувата. В результате реакции образуется молекулярный водород, от которого органелла получила свое название. [2]
Гидрогеносомы имеют размер 0,5-2 микрометра и связаны двойной мембраной. Чаще всего они имеют форму гантели и встречаются в больших комплексах сложенных друг на друга гидрогеносом. Эти стопки варьируются от 4 или 5 (так называемые ювенильные комплексы) до 20 и более гидрогеносом. [1]
В большинстве случаев гидрогеносомы являются безгеномными , поскольку большая часть митохондриального генома была перенесена в ядро; из-за этого все гидрогеносомные белки импортируются в органеллу. [6] [7] Однако гидрогеносомный геном был обнаружен у инфузории таракана Nyctotherus ovalis и страменопиля Blastocystis . [8]
В связи с тем, что многие организмы эволюционировали, чтобы приспособиться к своей анаэробной среде, у множества организмов независимо развились гидрогеносомы или структуры со схожими функциями. Сходство между Nyctotherus и Blastocystis , которые имеют лишь отдаленное родство, считается результатом конвергентной эволюции и ставит под сомнение существование четкого различия между митохондриями, гидрогеносомами и митосомами (еще один вид вырожденных митохондрий). [1] [8]
Организмы-источники [ править ]
Неисчерпывающий список организмов, содержащих гидрогеносомы, включает:
- парабазальные жгутиконосцы (например, Trichomonas vaginalis , Tritrichomonas fetus , Histomonas meleagridis )
- преакостилидные жгутиконосцы (например, Trimastixpyriformis )
- гетеролобозные амебофлагелляты (например, фонарь Psalteriomonas )
- анаэробные инфузории (например, Nyctotherus ovalis , Metopus palaeformis , Trimyema compressum , Caenomorpha uniserialis , Dasytricha ruminantium )
- анаэробные хитридиомицеты (например, Neocallimastix виды , виды Piromyces )
Огромное разнообразие исходных организмов можно объяснить теоретической конвергентной эволюцией гидрогеносом из митохондрий для приспособления к анаэробной среде. [1] [6] [8]
В 2010 году ученые также сообщили об открытии первых известных анаэробных многоклеточных животных с органеллами, подобными гидрогеносомам. Три многоклеточных вида Loricifera — Spinoloricus nov. сп. , Ругилорикус ноябрь. сп. и Pliciloricus nov. сп. — были обнаружены глубоко в средиземноморских отложениях и используют гидрогеносомы в своем анаэробном цикле метаболизма. [9]
Синтез АТФ [ править ]
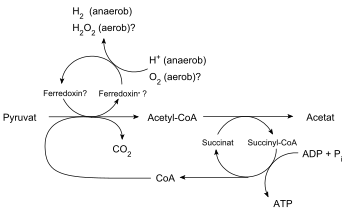
Гидрогеносомы трихомонад (наиболее изученных из гидрогеносомосодержащих микроорганизмов) продуцируют молекулярный водород , ацетат , углекислый газ и АТФ за счет совместного действия пируват:ферредоксиноксидоредуктазы , гидрогеназы , ацетат:сукцинат-КоА-трансферазы и сукцинаттиокиназы . супероксиддисмутаза , малатдегидрогеназа (декарбоксилирующая), ферредоксин , аденилаткиназа и НАДН:ферредоксиноксидоредуктаза. В гидрогеносоме также локализуются [10]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и де Грааф Р.М., Дуарте И., ван Ален Т.А., Койпер Дж.В., Шотанус К., Розенберг Дж. и др. (декабрь 2009 г.). «Гидрогеносомы Psalteriomonas lanterna» . Эволюционная биология BMC . 9 (1): 287. дои : 10.1186/1471-2148-9-287 . ПМЦ 2796672 . ПМИД 20003182 .
- ^ Jump up to: Перейти обратно: а б с Линдмарк, Дональд Г.; Мюллер, Миклош (25 ноября 1973 г.). «Гидрогеносома, цитоплазматическая органелла плода анаэробных жгутиковых Tritrichomonas, и ее роль в метаболизме пирувата» . Журнал биологической химии . 248 (22): 7724–7728. дои : 10.1016/S0021-9258(19)43249-3 . ISSN 0021-9258 . ПМИД 4750424 .
- ^ «Дозировка флагила, флагила ER (метронидазола), показания, взаимодействие, побочные эффекты и многое другое» . ссылка.medscape.com . Проверено 11 апреля 2021 г.
- ^ Хрды И., Каммак Р., Стопка П., Кулда Дж., Тачези Дж. (декабрь 2005 г.). «Альтернативный путь активации метронидазола в гидрогеносомах Trichomonas vaginalis» . Антимикробные средства и химиотерапия . 49 (12): 5033–6. doi : 10.1128/AAC.49.12.5033-5036.2005 . ПМЦ 1315937 . ПМИД 16304169 .
- ^ Шнайдер Р.Э., Браун М.Т., Шифлетт А.М., Дьялл С.Д., Хейс Р.Д., Се Ю и др. (ноябрь 2011 г.). «Протеом гидрогеносомы Trichomonas vaginalis сильно редуцирован по сравнению с митохондриями, но сложен по сравнению с митосомами» . Международный журнал паразитологии . 41 (13–14): 1421–34. дои : 10.1016/j.ijpara.2011.10.001 . ПМЦ 4437511 . ПМИД 22079833 .
- ^ Jump up to: Перейти обратно: а б Рада П., Долежал П., Едельски П.Л., Бурсак Д., Перри А.Дж., Шединова М. и др. (15 сентября 2011 г.). «Основные компоненты биогенеза органелл и мембранного транспорта в гидрогеносомах Trichomonas vaginalis» . ПЛОС ОДИН . 6 (9): e24428. Бибкод : 2011PLoSO...624428R . дои : 10.1371/journal.pone.0024428 . ПМК 3174187 . ПМИД 21935410 .
- ^ Май З., Гош С., Фрисарди М., Розенталь Б., Роджерс Р., Самуэльсон Дж. (март 1999 г.). «Hsp60 нацелен на загадочную органеллу митохондриального происхождения («криптон») микроаэрофильного простейшего паразита Entamoeba histolytica» . Молекулярная и клеточная биология . 19 (3): 2198–205. дои : 10.1128/MCB.19.3.2198 . ПМК 84012 . ПМИД 10022906 .
- ^ Jump up to: Перейти обратно: а б с Штехманн А., Хэмблин К., Перес-Брокаль В., Гастон Д., Ричмонд Г.С., ван дер Гизен М. и др. (апрель 2008 г.). «Органеллы Blastocystis, которые стирают различие между митохондриями и гидрогеносомами» . Современная биология . 18 (8): 580–5. дои : 10.1016/j.cub.2008.03.037 . ПМК 2428068 . ПМИД 18403202 .
- ^ Дановаро Р., Делл'Анно А., Пушедду А., Гамби С., Хайнер И., Кристенсен Р.М. (апрель 2010 г.). «Первые многоклеточные животные, живущие в постоянно бескислородных условиях» . БМК Биология . 8:30 . дои : 10.1186/1741-7007-8-30 . ПМЦ 2907586 . ПМИД 20370908 .
- ^ Хьорт К., Голдберг А.В., Цаусис А.Д., Хирт Р.П., Эмбли Т.М. (март 2010 г.). «Разнообразие и редуктивная эволюция митохондрий микробных эукариот» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 365 (1541): 713–27. дои : 10.1098/rstb.2009.0224 . ПМЦ 2817227 . ПМИД 20124340 .