Паразауролоф
| Паразауролоф | |
|---|---|

| |
| P. cyrtocristatus Скелет в Полевом музее естественной истории . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Орнитоподы |
| Семья: | † Гадрозавриды |
| Подсемейство: | † Ламбеозаврины |
| Племя: | † Паразауролофини |
| Род: | † Паразауролоф Парки , 1922 год. |
| Типовой вид | |
| † Паразауролоф ходери Парки, 1922 год.
| |
| Другие виды | |
| |
| Синонимы [ 1 ] | |
| |
Паразауролоф ( / ˌ p ær ə s ɔː ˈ r ɒ l ə f ə s , - ˌ s ɔːr ə ˈ l oʊ f ə s / ; что означает «рядом с хохлатой ящерицей» в отношении зауролофа ) [ 2 ] — род гадрозавридных Азии «утконосых» динозавров , обитавших на территории нынешней западной части Северной Америки и, возможно, в позднемеловой период , около 76,9–73,5 миллионов лет назад. [ 3 ] Это было крупное травоядное животное , которое могло достигать более 9 метров (30 футов) в длину и весить более 5 метрических тонн (5,5 коротких тонн) и могло передвигаться как двуногое, так и четвероногое . три вида Общепризнанными являются : P. walkeri ( типовой вид ), P. Tubeicen и короткохохлый P. cyrtocristatus . Кроме того, был предложен четвертый вид, P. jiayinensis , хотя его чаще относят к отдельному роду Charonosaurus . Останки известны из Альберты , Нью-Мексико и Юты , а также, возможно, из Хэйлунцзяна, если Харонозавр действительно является частью этого рода. Род был впервые описан в 1922 году Уильямом Парксом по черепу и частичному скелету, найденным в Альберте.
Паразауролоф был гадрозавридом , частью разнообразного семейства крупных орнитопод позднего мела, известных своими причудливыми украшениями на голове, которые, вероятно, использовались для общения и улучшения слуха. Этот род известен своим большим и сложным черепным гребнем, который образует длинную изогнутую трубку, выступающую вверх и назад от черепа в самой большой форме . Харонозавр из Китая, который, возможно, был его ближайшим родственником, имел похожий череп и потенциально похожий гребень. визуальное распознавание вида и пола, акустический резонанс и терморегуляция В качестве функционального объяснения гребня были предложены . Это один из самых редких гадрозавридов, известный лишь по нескольким хорошим экземплярам.
Открытие и присвоение имени
[ редактировать ]
Имя Паразауролоф , означающее «около хохлатой ящерицы», происходит от греческих слов para /παρα («рядом» или «рядом»), saurus / σαυρος («ящерица») и lophos /λοφος («гребень»). [ 4 ] Он основан на ROM 768, черепе и частичном скелете, у которых отсутствует большая часть хвоста и задних ног ниже колен, который был найден полевой группой из Университета Торонто в 1920 году недалеко от Сэнд-Крик вдоль реки Ред-Дир в Альберте. [ 5 ] Эти породы теперь известны как кампанского периода позднемелового формация Парка динозавров периода . Уильям Паркс назвал образец P. walkeri в честь сэра Байрона Эдмунда Уокера , председателя Попечительского совета Королевского музея Онтарио . [ 5 ] Останки паразауролофа в Альберте редки. [ 6 ] только с еще одним частичным черепом, который, возможно, принадлежит формации Парка Динозавров. [ 7 ] и три экземпляра из Парка динозавров без черепов, которые, возможно, принадлежат к этому роду. [ 6 ] В некоторых списках фауны есть упоминание о возможном P. walkeri материале в формации Хелл-Крик в Монтане , горной толще позднего маастрихта . [ 8 ] Это явление не отмечено Салливаном и Уильямсоном в их обзоре рода 1999 г. [ 9 ] и не получил дальнейшего развития где-либо еще.
В 1921 году Чарльз Х. Штернберг обнаружил часть черепа ( PMU .R1250) из того, что сейчас известно как немного более молодая формация Киртланд в округе Сан-Хуан , штат Нью-Мексико. Этот экземпляр был отправлен в Уппсалу , где Карл Виман описал его как второй вид, P. Tubeicen , в 1931 году. [ 10 ] Видовой эпитет происходит от латинского слова tƔbǐcěn , что означает «трубач». [ 11 ] Второй, почти полный череп P. Tubeicen ( NMMNH P-25100) был найден в Нью-Мексико в 1995 году. Используя компьютерную томографию этого черепа, Роберт Салливан и Томас Уильямсон в 1999 году дали этому роду монографическое описание, в котором были рассмотрены аспекты его анатомии и систематика, а также функции ее гребня. [ 9 ] Позже Уильямсон опубликовал независимый обзор останков, который не соответствовал таксономическим выводам. [ 12 ]
Джон Остром описал еще один хороший экземпляр ( FMNH P27393) из Нью-Мексико как P. cyrtocristatus в 1961 году. Он включает частичный череп с коротким закругленным гребнем и большую часть посткраниального скелета, за исключением ступней, шеи и частей хвоста. [ 13 ] Его конкретное название происходит от латинских слов curtus , что означает «укороченный», и cristatus , что означает «хохлатый». [ 11 ] Образец был найден либо в кровле формации Фрутленд , либо, что более вероятно, в основании перекрывающей формации Киртланд. [ 9 ] Ареал этого вида был описан в 1979 году, когда Дэвид Б. Вейшампель и Джеймс А. Дженсен описали частичный череп с похожим гребнем ( BYU кампанского возраста 2467) из формации Кайпаровиц в округе Гарфилд , штат Юта . [ 14 ] С тех пор в Юте был найден еще один череп с коротким и округлым P. cyrtocristatus . гребнем [ 9 ]
Разновидность
[ редактировать ]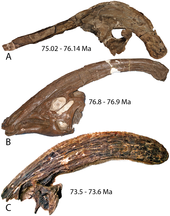
Паразауролоф известен из трех определенных видов: P. walkeri , P. Tubeicen и P. cyrtocristatus . [ 3 ] Все они четко отличаются друг от друга и имеют множество отличий. [ 15 ] [ 16 ] Первый названный вид, а значит и тип , — P. walkeri . К нему относят один конкретный экземпляр из формации Парка Динозавров. [ 17 ] но многие другие почти наверняка заслуживают внимания. [ 3 ] Как указано выше, он отличается от двух других видов тем, что имеет более простое внутреннее строение, чем P. Tubeicen . [ 9 ] наряду с более прямым гребнем и другой внутренней структурой, чем у P. cyrtocristatus . [ 15 ]
Следующий названный вид — P. Tubeicen , самый крупный из видов Parasaurolophus . [ 9 ] Он обитал в Нью-Мексико, где известны три экземпляра. [ 17 ] и его можно отличить от других его видов. [ 15 ] У него длинный и прямой гребень с очень сложной внутренней частью по сравнению с другими видами. [ 9 ] Все известные экземпляры P. Tubeicen происходят из пачки Де-На-Зин кертландской свиты . [ 18 ]
третий вид, P. cyrtocristatus назвал В 1961 году Джон Остром . [ 13 ] Три его известных экземпляра были найдены в формациях Фрутленд и Кайпаровитс в штатах Юта и Нью-Мексико. [ 3 ] Второй экземпляр, первый известный из свиты Кайпаровиц, изначально не был отнесен к определенному таксону. [ 14 ] Из Parasaurolophus видов P. cyrtocristatus самый маленький и имеет наиболее изогнутый гребень. [ 9 ] Из-за того, что он обладал двумя вышеуказанными признаками, часто предполагалось, что это была самка P. walkeri или P. Tubeicen , которые все считались самцами. [ 15 ] [ 19 ] хотя P. Tubecen жил примерно на миллион лет позже. [ 3 ] Как отметил Томас Уильямсон, типовой материал P. cyrtocristatus составляет около 72% размера P. Tubeicen , что близко к размеру, при котором другие ламбеозаврины интерпретируются как начинающие проявлять окончательный половой диморфизм в своих гребнях (~70% размера взрослых особей). ). [ 12 ] Несмотря на то, что многие ученые поддержали возможный факт того, что P. cyrtocristatus является самкой, [ 19 ] [ 20 ] многие другие исследования показали, что это не так. [ 17 ] [ 7 ] из-за различий в возрасте, распространении, а также больших различий в гребне и его внутреннем строении. [ 15 ]
Исследование, опубликованное в PLoS ONE можно отнести еще один вид в 2014 году, показало, что к Parasaurolophus . Это исследование, проведенное Сином, показало, что Charonosaurus jiayensis на самом деле находился глубоко внутри Parasaurolophus , что привело к созданию нового вида P. jiayensis . Если этот вид действительно находится внутри Parasaurolophus , то, следовательно, этот род просуществовал до вымирания K-Pg и известен на двух континентах. [ 21 ]
Описание
[ редактировать ]
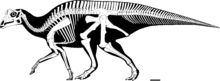
Как и у большинства динозавров, скелет паразауролофа известен не полностью. Длина типового экземпляра P. walkeri оценивается в 9,45 метра (31,0 фута). [ 22 ] а оценки массы тела, основанные на аллометрии, показывают, что человек длиной 9 метров (30 футов) весил бы более 5 метрических тонн (5,5 коротких тонн). [ 23 ] Грегори С. Пол подсчитал, что средняя взрослая особь этого типового вида будет иметь длину 7,5 метров (25 футов) и весить 2,6 метрических тонны (2,9 коротких тонны). [ 24 ] Его череп имеет длину около 1,6 м (5 футов 3 дюйма), включая гребень, тогда как типовой череп P. Tubeicen имеет длину более 2 м (6 футов 7 дюймов), что указывает на то, что это было более крупное животное. [ 25 ] Его единственная известная рука была относительно короткой для гадрозаврид, с короткой, но широкой лопаткой . у Длина бедренной кости составляет 103 см (41 дюйм) P. walkeri , и она прочнее по сравнению с другими гадрозавридами. [ 25 ] Кости плеча и таза также были массивными. [ 26 ]
Как и другие гадрозавриды, он мог ходить как на двух, так и на четырех ногах. Вероятно, он предпочитал добывать пищу на четырех ногах, но бегал на двух. [ 8 ] Нервные отростки позвонков . были высокими, как это часто бывает у ламбеозавринов [ 25 ] В самом высоком положении над бедрами они увеличили высоту спины. кожи известны отпечатки У P. walkeri , на которых видны однородные бугоркообразные чешуйки, но без более крупных структур. [ 5 ]
Череп
[ редактировать ]
Наиболее заметной особенностью был черепной гребень, выступавший из задней части головы и состоявший из предчелюстной и носовой костей . [ 25 ] Гребень был полым, с отдельными трубками, ведущими от каждой ноздри к концу гребня, а затем меняя направление и направляясь обратно вниз по гребню в череп. Трубочки были самыми простыми у P. walkeri и более сложными у P. Tubeicen , где некоторые трубки были слепыми, а другие встречались и разделялись. [ 9 ] В то время как P. walkeri и P. Tubeicen имели длинные гребни с небольшой кривизной, P. cyrtocristatus имел короткий гребень более круглого профиля. [ 13 ]
Классификация
[ редактировать ]Как следует из названия, паразауролоф первоначально считалось, что тесно связан с зауролофом из-за его внешне похожего гребня. [ 5 ] Однако вскоре он был признан членом подсемейства ламбеозавринов гадрозаврид — зауролоф является гадрозаврином . [ 27 ] Его обычно интерпретируют как отдельное ответвление ламбеозавринов, отличное от шлемообразного коритозавра , гипакрозавра и ламбеозавра . [ 8 ] [ 7 ] Его ближайшим известным родственником, по-видимому, является Харонозавр , ламбеозавр с похожим черепом (но пока без полного гребня) из Амурской области на северо-востоке Китая. [ 28 ] Эти двое могут образовывать кладу Parasaurolophini . P. cyrtocristatus с коротким закругленным гребнем может быть самым базальным из трех известных Parasaurolophus. видов [ 7 ] или это могут быть полувзрослые или женские особи P. Tubeicen . [ 12 ]

Следующая кладограмма взята после переописания Lambeosaurus magnicristatus в 2007 году (Evans and Reisz, 2007): [ 7 ]
| Гадрозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]
Диета и кормление
[ редактировать ]Будучи гадрозавридом, паразауролоф был крупным двуногим и четвероногим травоядным, питавшимся растениями , со сложным черепом, позволяющим совершать измельчающие движения, аналогичные жеванию . Его зубы постоянно заменялись и были упакованы в стоматологические батареи, содержащие сотни зубов, но лишь небольшая часть из них в любой момент времени использовалась. Он использовал свой клюв для сбора растительного материала, который удерживался в челюстях органом, похожим на щеку . Растительность могла быть поднята с земли на высоту около 4 м (13 футов). [ 17 ] Как отметил Роберт Баккер , ламбеозаврины имеют более узкие клювы, чем гадрозаврины, а это означает, что паразауролоф и его родственники могли питаться более избирательно, чем их ширококлювые собратья без гребня. паразауролофа состояла из листьев, веток и сосновых иголок, что означало, что это был браузер. Диета [ 29 ]
Рост
[ редактировать ]
Паразауролоф известен по многим взрослым особям, а также молодь, описанная в 2013 году, под номером RAM 140000 и по прозвищу Джо, [ 30 ] в честь волонтера Палеонтологического музея Раймонда М. Альфа (RAM). Молодь была обнаружена в формации Кайпаровиц в 2009 году. Выкопанная совместной экспедицией музея и Школы Уэбба , молодому человеку было около одного года, когда он умер. Относящаяся к Parasaurolophus sp., молодь является наиболее полной, а также самой молодой из когда-либо обнаруженных Parasaurolophus , ее размеры составляют 2,5 м (8,2 фута). Этот человек четко вписывается в известные в настоящее время стадии роста паразауролофа и жил примерно 75 миллионов лет назад. Несмотря на то, что полного черепа промежуточного возраста между RAM 14000 и взрослым паразауролофом еще не обнаружено, известна частичная черепная коробка примерно нужного размера. Молодые особи, составляющие 25% от общего размера взрослой особи, показывают, что рост гребня у Parasaurolophus начался раньше, чем у родственных родов, таких как Corythosaurus . Было высказано предположение, что взрослые особи паразауролофа имели такие большие гребни, особенно по сравнению с родственным коритозавром , из-за разницы в возрасте между моментами, когда их гребни начали развиваться. Гребень молоди не длинный и трубчатый, как у взрослых, а низкий и полусферический. [ 22 ]
Череп RAM 14000 почти цел, на левой стороне отсутствует только часть верхней челюсти . Однако череп был расколот посередине в результате эрозии , возможно, когда он лежал на дне русла реки . Обе стороны слегка смещены, при этом некоторые кости правой руки оторвались от основного блока, также в результате эрозии. После реконструкции череп, вид сбоку, напоминает других найденных молодых ламбеозавринов и имеет примерно трапеции . форму [ 22 ]
Частичный краниальный эндокаст для RAM 14000 был реконструирован по данным компьютерной томографии, впервые для паразауролофа на любой стадии онтогенеза. Эндокаст был реконструирован из двух частей: одна на части черепной коробки, сочлененной с левой половиной черепа, а другая на разъединенной части черепной коробки. Их относительное положение затем было аппроксимировано на основе краниальных ориентиров и сравнения с другими гадрозавридами. Из-за выветривания многие более мелкие нервные каналы и отверстия не удалось точно идентифицировать. [ 22 ]
Краниальный гребень
[ редактировать ]
Было выдвинуто множество гипотез относительно того, какие функции выполнял краниальный гребень паразауролофа , но большинство из них были дискредитированы. [ 31 ] [ 32 ] Сейчас считается, что он мог иметь несколько функций: визуальное отображение для определения вида и пола, усиление звука для общения и терморегуляция. Неясно, что и в какой момент имело наибольшее значение в эволюции гребня и его внутренних носовых ходов. [ 33 ]
Различия в гербах
[ редактировать ]Что касается других ламбеозавринов, то считается, что краниальный гребень Parasaurolophus менялся с возрастом и у взрослых имел половой диморфизм . Джеймс Хопсон , один из первых исследователей, описавших гребни ламбеозаврин с точки зрения таких различий, предположил, что P. cyrtocristatus с его маленьким гребнем был женской формой P. Tubeicen . [ 20 ] Томас Уильямсон предположил, что это ювенильная форма. Ни одна из гипотез не получила широкого признания. Как всего шесть хороших черепов, одна юная черепная коробка, [ 12 ] Известен один недавно обнаруженный череп молодого человека, [ 22 ] дополнительный материал поможет прояснить эти потенциальные связи. Уильямсон отметил, что в любом случае молодые особи Parasaurolophus, вероятно, имели маленькие округлые гребни, как у P. cyrtocristatus , которые, вероятно, росли быстрее по мере приближения особей к половой зрелости . [ 12 ] Недавнее повторное исследование черепной коробки молодых особей, ранее отнесенной к Lambeosaurus , а теперь отнесенной к Parasaurolophus , предоставило доказательства того, что у молодых особей присутствовал небольшой трубчатый гребень. У этого экземпляра сохранилось небольшое расширение лобных костей вверх , похожее на то, что наблюдается у взрослых особей, но меньшее; у взрослых лобные кости образовывали платформу, поддерживающую основание гребня. Этот образец также указывает на то, что рост гребня у Parasaurolophus и профиль лица молодых особей отличались от модели Corythosaurus – Hypacrosaurus – Lambeosaurus , отчасти потому, что на гребне Parasaurolophus отсутствует тонкий костный «гребешок», составляющий верхнюю часть гребень трех других ламбеозавринов. [ 7 ]
Гипотезы отклоненных функций
[ редактировать ]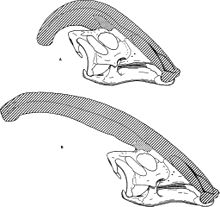
Многие ранние предложения были сосредоточены на адаптации к водному образу жизни, следуя гипотезе о том, что гадрозавриды были земноводными, что было распространено до 1960-х годов. Так, Альфред Шервуд Ромер предложил использовать его в качестве трубки . [ 34 ] Мартин Уилфарт, что это была насадка для подвижного хоботка, используемого в качестве дыхательной трубки или для сбора пищи, [ 35 ] Чарльз М. Штернберг , что он служил воздушной ловушкой, не допускающей попадания воды в легкие . [ 36 ] и Неда Колберта , что он служил резервуаром для воздуха при длительном пребывании под водой. [ 37 ]
Другие предложения носили более механический характер. Уильям Паркс в 1922 году предположил, что гребень соединен с позвонками над плечами связками или мышцами и помогает двигать и поддерживать голову. [ 5 ] Это маловероятно, поскольку у всех современных архозавров выйная связка прикрепляется к шее или основанию черепа. [ 38 ] Отенио Абель предположил, что его использовали в качестве оружия в бою между представителями одного и того же вида. [ 39 ] и Эндрю Милнер предположил, что его можно использовать в качестве дефлектора листвы, как гребень шлема (так называемый «каска») казуара . [ 32 ] Тем не менее, другие предложения сделали размещение специализированных органов основной функцией. Хальшка Осмольска предположила, что в нем находятся солевые железы . [ 40 ] были расширены области обонятельной ткани и значительно улучшено обоняние . и Джон Остром предположил, что у ламбеозавринов, которые не имели очевидных защитных способностей, [ 41 ]
Большинство этих гипотез были дискредитированы или отвергнуты. [ 31 ] Например, на конце гребня нет отверстия для подводного плавания. На хоботке нет мышечных рубцов, и сомнительно, что он понадобится животному с клювом. Будучи предлагаемым шлюзом, он не защищал бы воду. Предложенного резервуара с воздухом было бы недостаточно для животного размером с паразауролофа . У других гадрозаврид были большие головы, и им не требовались большие полые гребни, служащие точками крепления поддерживающих связок. [ 41 ] Кроме того, ни одно из предложений не объясняет, почему гребень имеет такую форму, почему у других ламбеозавридов должны быть гребни, которые выглядят совсем по-другому, но выполняют аналогичную функцию, как гадрозавриды без гребня или со сплошным гребнем обходились без таких способностей или почему некоторые гадрозавриды имели твердые гребни. гребни. Эти соображения особенно влияют на гипотезы, основанные на увеличении возможностей систем, уже присутствующих у животного, таких как гипотезы солевых желез и обоняния. [ 32 ] и указать, что это не были основные функции гребня. Кроме того, работа с носовой полостью ламбеозавринов показывает, что обонятельные нервы и соответствующая сенсорная ткань находились в основном за пределами носовых ходов в гребне, поэтому расширение гребня имело мало общего с обонянием. [ 33 ]
Гипотеза регулирования температуры
[ редактировать ]Большая площадь поверхности и васкуляризация гребня также предполагают терморегуляторную функцию. [ 42 ] Первым, кто предположил, что краниальные гребни ламбеозавринов связаны с регуляцией температуры, был Wheeler (1978). существует нервная связь Он предположил, что между гребнем и мозгом , так что последний может охлаждаться первым. [ 43 ] [ 44 ] Следующими людьми, опубликовавшими схожую идею, были Тереза Марьянска и Осмольска, которые поняли, что, как и современные ящерицы, динозавры могли обладать солевыми железами и охлаждаться за счет осморегуляции. [ 44 ] [ 40 ] В 2006 году Эванс опубликовал аргументацию о функциях гребней ламбеозавров и объяснил, почему это может быть причиной эволюции гребня. [ 33 ]
Поведенческие гипотезы
[ редактировать ]
Часто предполагается, что паразауролоф использовал свой гребень в качестве резонирующей камеры для производства низкочастотных звуков, чтобы предупредить других членов группы или ее вида. [ 19 ] Эта функция была первоначально предложена Виманом в 1931 году, когда он описал P. Tubeicen . Он отметил, что внутренние структуры гребня похожи на структуру лебедя, и предположил, что животное может использовать свои удлиненные носовые ходы для создания шума. [ 19 ] [ 10 ] Однако носовые трубки гипакрозавра , коритозавра и ламбеозавра гораздо более разнообразны и сложны, чем дыхательные пути паразауролофа . Большое количество материала и данных подтверждает гипотезу о том, что большой трубчатый гребень паразауролофа представлял собой резонирующую камеру. Вейшампель в 1981 году предположил, что паразауролоф издает звуки в диапазоне частот от 55 до 720 Гц , хотя существовала некоторая разница в диапазоне отдельных видов из-за размера, формы и длины носового хода, что наиболее очевидно у P. cyrtocristatus (интерпретируется как возможная женщина). [ 19 ] Хопсон обнаружил, что существуют анатомические доказательства того, что гадрозавриды обладали сильным слухом. есть по крайней мере один пример У родственного коритозавра тонкого стремечка (рептильной ушной кости), что в сочетании с большим пространством для барабанной перепонки предполагает чувствительное среднее ухо. Кроме того, лагена гадрозаврид удлинена, как у крокодила, что указывает на хорошо развитую слуховую часть внутреннего уха. [ 20 ] Основываясь на сходстве внутренних ушей гадрозаврид с крокодилами , он также предположил, что взрослые гадрозавриды чувствительны к высоким частотам, которые могут воспроизводить их потомки. По мнению Вейшампеля, это соответствует общению родителей и детей. [ 19 ]
Компьютерное моделирование хорошо сохранившегося экземпляра P. Tubeicen с более сложными дыхательными путями, чем у P. walkeri , позволило реконструировать возможный звук, издаваемый его гребнем. [ 45 ] Основной путь резонирует на частоте около 30 Гц, но сложная анатомия пазухи вызывает пики и спады звука. [ 46 ] Другая основная поведенческая теория заключается в том, что гребень использовался для внутривидового распознавания. [ 44 ] Это означает, что герб мог использоваться для распознавания видов, в качестве предупреждающего сигнала и для других целей, не сексуальных. Это могло быть одной из причин появления гребней у паразауролофа и других гадрозаврид. [ 15 ] Вместо этого социальные и физиологические функции стали более поддерживаться как функции гребня с упором на визуальную и слуховую идентификацию и общение. Будучи крупным объектом, гребень имеет явное значение как визуальный сигнал и отличает это животное от его современников. Большой размер глазниц гадрозаврид и наличие склеротических колец в глазах предполагают острое зрение и дневной образ жизни, что свидетельствует о важности зрения для этих животных. Если бы, как обычно изображают, кожаный оборок простирался от гребня до шеи или спины, предполагаемое визуальное представление было бы еще более эффектным. [ 20 ] Как предполагают другие черепа ламбеозавров, гребень паразауролофа, вероятно, позволял как идентифицировать вид (например, отделить его от коритозавра или ламбеозавра ), так и половую идентификацию по форме и размеру. [ 33 ]
Оборка из мягких тканей
[ редактировать ]
Барнум Браун (1912) отметил наличие тонких полосок вблизи задней части гребня, которые, по его гипотезе, могли быть связаны с наличием оборки кожи, сравнимой с той, что встречается у современной ящерицы-василиска . Его гипотеза, по-видимому, была подтверждена кожей, сохранившейся над шеей и спиной коритозавра и эдмонтозавра . Впоследствии реконструкции паразауролофа со значительной оборкой кожи между гребнем и шеей появились во влиятельном палеоискусстве, включая фрески Чарльза Р. Найта и в Уолта Диснея анимационном фильме «Фантазия» . Это привело к тому, что оборка изображалась во многих других источниках, несмотря на появление ныне опровергнутой гипотезы о «трубке» и объединение гипотезы оборки с идеей о том, что гребень служит опорной точкой для связок шеи, наряду с отсутствием убедительные доказательства его присутствия, он потерял популярность в большинстве современных изображений. [ 38 ]
Палеопатология
[ редактировать ]
P. walkeri известен по одному экземпляру, который может содержать патологию . На скелете виден V-образный разрыв или выемка в позвонках у основания шеи. [ 16 ] Первоначально это считалось патологией, но Паркс опубликовал вторую интерпретацию этого явления: прикрепление связок, поддерживающих голову. Гребень прикреплялся к зазору с помощью мышц или связок и использовался для поддержки головы с оборкой, как, по прогнозам, существует у некоторых гадрозаврид. [ 5 ] Другая возможность заключается в том, что во время подготовки образец был поврежден, что привело к возможной патологии. [ 16 ] Однако насечка по-прежнему считается скорее патологией. [ 16 ] [ 31 ] хотя на некоторых изображениях Parasaurolophus восстанавливается кожный лоскут. [ 9 ]
Еще одну возможную патологию заметил Паркс, причем со стороны выреза. В четвертом, пятом и шестом позвонках, непосредственно перед вырезкой, были повреждены нервные отростки. У четвертого был очевидный перелом, а у двух других была опухоль у основания перелома. [ 5 ]
Анализ патологии, проведенный Бертоццо и др. и опубликованный в декабре 2020 года, предполагает, что патология плеча и грудных ребер у голотипа P. walkeri, вероятно, была результатом удара динозавра падающим деревом, возможно, во время серьезной шторм. Судя по возобновлению роста костей, предполагается, что гадрозавр выживал от одного до четырех месяцев, а возможно, и лет после травмы. Считается, что ни одна из патологий голотипа не вызвала и не способствовала его смерти. [ 47 ]
Палеоэкология
[ редактировать ]Альберта
[ редактировать ]
Parasaurolophus walkeri из формации Парка Динозавров был представителем разнообразной и хорошо документированной фауны доисторических животных, включая таких известных динозавров, как рогатый центрозавр , хасмозавр и стиракозавр ; орнитомимиды Struthiomimus ; собратья-утконосы Грипозавр и Коритозавр ; тираннозавриды горгозавр и дасплетозавр ; и панцирные Эдмонтония , Эуоплоцефал и Диоплозавр . [ 8 ] Это был редкий представитель этой фауны. [ 6 ] Формация «Парк динозавров» интерпретируется как низкий рельеф рек и пойм , которые со временем стали более заболоченными и находились под влиянием морских условий по мере того, как Западный внутренний морской путь продвигался на запад. [ 6 ] Климат был теплее , чем в современной Альберте, без морозов , но с более влажными и сухими сезонами. Хвойные деревья, по-видимому, были доминирующими растениями кроны , с подлеском из папоротников , древовидных папоротников и покрытосеменных растений . [ 6 ]
Некоторые из менее распространенных гадрозавров в формации Парка динозавров Провинциального парка динозавров, такие как Паразауролоф , могут представлять собой останки особей, погибших во время миграции через регион. У них также могла быть более возвышенная среда обитания, где они могли гнездиться или питаться. Присутствие паразауролофа и критозавра в окаменелостях северных широт может свидетельствовать об обмене фауной между разными северными и южными биомами в позднемеловом периоде Северной Америки. Оба таксона редки за пределами южного биома, где, наряду с Pentaceratops , они являются преобладающими представителями фауны. [ 48 ]
Нью-Мексико
[ редактировать ]
В формации Фрутленд в Нью-Мексико P. cyrtocristatus делил свою среду обитания с другими птицетазовыми животными и тероподами. В частности, его современниками были цератопсы Pentaceratops sternbergii ; [ 8 ] пахицефалозавр Stegoceras nomexicanum ; [ 49 ] и некоторые неопознанные окаменелости, принадлежащие тираннозаврам , ? Орнитомим , ? Троодонтиды , ? Saurornitholestes langstoni , ? Струтиомим , Орнитопода , ? Хасмозавр , ? Corythosaurus , Hadrosaurinae , Hadrosauridae и Ceratopsidae . [ 8 ] Когда существовал Parasaurolophus , формация Fruitland была болотистой, располагалась в низменности и недалеко от берега Мелового внутреннего морского пути . Самая нижняя часть формации Fruitland возникла чуть моложе 75,56 ± 0,41 млн лет назад, а самая верхняя граница датируется 74,55 ± 0,22 млн лет назад. [ 50 ]
Существуя несколько позже видов из формации Фрутленд, P. Tubeicen встречается также в Нью-Мексико, в формации Киртланд . [ 8 ] Из этой формации происходят многочисленные группы позвоночных, в том числе рыбы , круротарсаны , [ 50 ] птицетазовые , ящеротазовые , [ 8 ] птерозавры , [ 51 ] и черепахи . Рыбы представлены двумя видами Melvius chauliodous и Myledalphus bipartitus . К круротарсанам относятся Brachychampsa montana и Denazinosuchus kirtlandicus . [ 50 ] Орнитисхии свиты представлены гадрозавридами Anasazisaurus Horneri , Naashoibitosaurus Ostromi , Kritosaurus navajovius , P. Tubeicen ; анкилозавриды Ahshislepelta major и Nodocephalosaurus kirtlandensis ; цератопсы Pentaceratops sternbergii [ 8 ] и Titanoceratops Ouranos ; [ 52 ] и пахицефалозавры Stegoceras nomexicanum. [ 49 ] и Sphaerotholus Goodwini [ 50 ] К ящерицам относятся тираннозавриды Bistahieversor sealeyi ; [ 53 ] орнитомимид sp Ornithomimus .; [ 8 ] и троодонтид Saurornitholestes Robustus . [ 54 ] Известен один птерозавр по имени Navajodactylus boerei . [ 51 ] Черепахи довольно многочисленны и известны из Denazinemys nodosa , Basilemys nobilis , Neurankylus baueri , Plastomenusrobustus и Thescelus hemispherica . Известны неопознанные таксоны, в том числе круротарсан? Лейдиозух , [ 50 ] а тероподы ? Struthiomimus , Troodontidae и Tyrannosauridae. [ 8 ] Начало формации Киртланд датируется 74,55 ± 0,22 млн лет назад, а окончание формирования - около 73,05 ± 0,25 млн лет назад. [ 50 ]
Юта
[ редактировать ]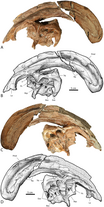

Аргон-аргоновое радиометрическое датирование показывает, что формация Кайпаровиц отложилась между 76,6 и 74,5 миллионами лет назад, в кампанский период позднего мелового периода. [ 55 ] [ 56 ] В позднемеловой период место формирования формации Кайпаровиц располагалось недалеко от западного берега Западного внутреннего морского пути , большого внутреннего моря, которое разделяло Северную Америку на два массива суши: Ларамидию на западе и Аппалачи на востоке. Плато, на котором жили динозавры, представляло собой древнюю пойму, в которой преобладали большие каналы и обильные водно-болотные торфяные болота, пруды и озера, и граничило с высокогорьем. Климат был влажным и влажным и поддерживал обильное и разнообразное разнообразие организмов. [ 57 ] Это образование содержит одни из лучших и наиболее непрерывных записей о земной жизни позднего мела в мире. [ 58 ]
Паразауролоф разделял свою палеосреду с другими динозаврами, такими как -дромеозавриды тероподы , троодонтид Talos sampsoni , орнитомимиды , такие как Ornithomimus velox , тираннозавриды, такие как Teratophoneus , панцирные анкилозавриды , утконосый гадрозавр Gryposaurus Monumentensis , цератопсы. Utahceratops gettyi , Nasutoceratops titusi и Kosmoceratops richardsoni и овирапторозавр Hagryphus giganteus . [ 59 ] Палеофауна, присутствующая в формации Кайпаровиц, включала хондрихтианов (акул и скатов), лягушек , саламандр , черепах , ящериц и крокодилов, таких как высший хищник Деинозух . множество ранних млекопитающих Присутствовало , включая многотуберкулезных , сумчатых и насекомоядных . [ 60 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]Сноски
[ редактировать ]- ^ Мартин 2014 .
- ^ Кольбер, Эдвин Х. (Эдвин Харрис); Найт, Чарльз Роберт (1951). Книга динозавров: правящие рептилии и их родственники . Нью-Йорк: МакГроу-Хилл. п. 152.
- ^ Перейти обратно: а б с д и Эванс и др. 2009 .
- ^ Лидделл и Скотт 1980 .
- ^ Перейти обратно: а б с д и ж г Парки 1922 года .
- ^ Перейти обратно: а б с д и Карри и Коппельхус 2005 .
- ^ Перейти обратно: а б с д и ж Эванс и Рейс 2007 .
- ^ Перейти обратно: а б с д и ж г час я дж к Вейшампель и др. 2004 .
- ^ Перейти обратно: а б с д и ж г час я дж Салливан и Уильямсон 1999 .
- ^ Перейти обратно: а б Виман 1931 г.
- ^ Перейти обратно: а б Симпсоны 1979 года .
- ^ Перейти обратно: а б с д и Уильямсон 2000 .
- ^ Перейти обратно: а б с Остром 1961 г.
- ^ Перейти обратно: а б Вейшампель и Дженсен, 1979 .
- ^ Перейти обратно: а б с д и ж Hone et al. 2011Хон и др. 2011
- ^ Перейти обратно: а б с д Бенсон и др. 2012 .
- ^ Перейти обратно: а б с д Хорнер и др. 2004 .
- ^ Салливан и др. 2011 .
- ^ Перейти обратно: а б с д и ж Вейшампель 1981 .
- ^ Перейти обратно: а б с д Хопсон 1975 .
- ^ Син и др. 2014 .
- ^ Перейти обратно: а б с д и Фарке и др. 2013 .
- ^ Сибахер, Ф. (2001). «Новый метод расчета аллометрических отношений длины и массы динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 . JSTOR 4524171 . S2CID 53446536 .
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 341. ИСБН 978-1-78684-190-2 . OCLC 985402380 .
- ^ Перейти обратно: а б с д Лулл и Райт, 1942 год .
- ^ Бретт-Сурман и Вагнер 2006 .
- ^ Гилмор 1924 .
- ^ Годфруа и др. 2000 .
- ^ Баккер 1986 .
- ^ «Динозавр Джо» . Музей Раймонда Альфа . Проверено 31 марта 2021 г.
- ^ Перейти обратно: а б с Перенасыщение 1997 года .
- ^ Перейти обратно: а б с Норман 1985 .
- ^ Перейти обратно: а б с д Эванс 2006 .
- ^ Ромер 1933 .
- ^ Уилфарт 1947 .
- ^ Штернберг 1935 .
- ^ Кольбер 1945 .
- ^ Перейти обратно: а б Мануччи Ф., Демпси М., Танке Д. Х. и др. Описание и этиология палеопатологических поражений типового экземпляра Parasaurolophus walkeri (Dinosauria: Hadrosauridae) с предлагаемыми реконструкциями выйной связки J. Anat. 2020; 00:1– 15. https://doi.org/10.1111/joa.13363
- ^ Абель 1924 .
- ^ Перейти обратно: а б Марьянская и Осмольская 1979 .
- ^ Перейти обратно: а б Остром 1962 г.
- ^ Салливан и Уильямсон 1996 .
- ^ Уиллер 1978 .
- ^ Перейти обратно: а б с Вейшампель 1997 .
- ^ Сандия 1997 .
- ^ Дигерт и Уильямсон 1998 .
- ^ Бертоццо, Филиппо; Мануччи, Фабио; Демпси, Мэтью; Танке, Даррен Х.; Эванс, Дэвид С.; Раффелл, Аластер; Мерфи, Эйлин (2020). «Описание и этиология палеопатологических поражений типового экземпляра Parasaurolophus walkeri (Dinosauria: Hadrosauridae) с предлагаемыми реконструкциями выйной связки» . Журнал анатомии . 238 (5): 1055–1069. дои : 10.1111/joa.13363 . ПМЦ 8053592 . ПМИД 33289113 .
- ^ Танке и Карпентер 2001 .
- ^ Перейти обратно: а б Ясински и Салливан 2011 .
- ^ Перейти обратно: а б с д и ж Салливан и Лукас 2006 .
- ^ Перейти обратно: а б Салливан и Фаулер 2011 .
- ^ Лонгрич 2011 .
- ^ Карр и Уильямсон 2010 .
- ^ Эванс и др. 2014 .
- ^ Робертс и др. 2005
- ^ Итон 2002 .
- ^ Титус и Лоуэн, 2013 .
- ^ Клинтон 1996 .
- ^ Занно и Сэмпсон 2005 .
- ^ Итон и др. 1999
Цитаты
[ редактировать ]- Абель, Отенио (1924). «Новые находки динозавров в верхнем мелу Канады». Ежегодник естественных наук (на немецком языке). 12 (36): 709–716. Бибкод : 1924NW.....12..709A . дои : 10.1007/BF01504818 . S2CID 1133858 .
- Баккер, RT (1986). Ересь динозавров: новые теории, раскрывающие тайны динозавров и их вымирания . Уильям Морроу. п. 194. ИСБН 978-0-8217-2859-8 .
- Бенсон, Р.Дж.; Брюссатт, SJ; Андерсон; Хон, Д.; Парсонс, К.; Сюй, Х.; Милнер, Д.; Нэйш, Д. (2012). Доисторическая жизнь . Дорлинг Киндерсли. п. 342. ИСБН 978-0-7566-9910-9 .
- Бретт-Сурман, Майкл К.; Вагнер, Джонатан Р. (2006). «Аппендикулярная анатомия кампанских и маастрихтских североамериканских гадрозаврид». В Карпентер, Кеннет (ред.). Рога и клювы: цератопсы и динозавры-орнитоподы . Блумингтон и Индианаполис: Издательство Университета Индианы. стр. 135–169. ISBN 978-0-253-34817-3 .
- Карр, Т.Д.; Уильямсон, Т.Э. (2010). « Bistahievers sealeyi , gen. et sp. nov., новый тираннозавроид из Нью-Мексико и происхождение глубоких морд у Tyrannosauroidea». Журнал палеонтологии позвоночных . 30 (1): 1–16. Бибкод : 2010JVPal..30....1C . дои : 10.1080/02724630903413032 . S2CID 54029279 .
- Кольбер, Эдвин Х. (1945). Книга о динозаврах: правящие рептилии и их родственники . Нью-Йорк: Американский музей естественной истории, Публикации «Человек и природа», 14 с. 156 . OCLC 691246 .
- Дигерт, CF; Уильямсон, Т.Е. (1998). «Цифровая акустическая модель гадрозавра ламбеозавра Parasaurolophus Tubeicen ». Журнал палеонтологии позвоночных . 18 (3):38А. дои : 10.1080/02724634.1998.10011116 .
- Карри, Филипп Дж.; Коппельхус, Ева, ред. (2005). Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема . Блумингтон: Издательство Университета Индианы. стр. 312–348 . ISBN 978-0-253-34595-0 .
- Клинтон, Уильям. «Президентская декларация: создание национального памятника Большой лестницы-Эскаланте» . 18 сентября 1996 года . Архивировано из оригинала 28 августа 2013 года . Проверено 9 ноября 2013 г.
- Итон, Дж. Г. (2002). «Многотуберкулезные млекопитающие из формаций Вахвеап (Кампан, Аквилан) и Кайпаровит (Кампан, Юдит) на территории национального памятника Большая Лестница-Эскаланте и рядом с ним, южная Юта». Разные публикации 02-4, Геологическая служба Юты : 1–66.
- Итон, Дж. Г.; Чифелли, РЛ; Хатчинсон, Дж. Х.; Киркланд, Дж.И.; Пэрриш, MJ (1999). «Меловая фауна позвоночных с плато Кайпаровиц, юго-центральная часть штата Юта». В Gillete, Дэвид Д. (ред.). Палеонтология позвоночных в Юте . Разная публикация 99-1. Солт-Лейк-Сити: Геологическая служба Юты. стр. 345–353. ISBN 978-1-55791-634-1 .
- Эванс, округ Колумбия (2006). «Гомологии полости носа и функция черепного гребня у динозавров-ламбеозавров». Палеобиология . 32 (1): 109–125. Бибкод : 2006Pbio...32..109E . дои : 10.1666/04027.1 . S2CID 198152630 .
- Эванс, округ Колумбия; Рейс, Р.Р. (2007). «Анатомия и взаимоотношения Lambeosaurus magnicristatus , хохлатого гадрозавридного динозавра (Ornithischia) из формации Парк динозавров, Альберта». Журнал палеонтологии позвоночных . 27 (2): 373–393. doi : 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2 . S2CID 86070917 .
- Эванс, округ Колумбия; Бавингтон, Р.; Кампионе, Нью-Йорк (2009). «Необычная черепная коробка гадрозавридов из формации Парк динозавров и биостратиграфия паразауролофа (Ornithischia: Lambeosaurinae) из южной Альберты» . Канадский журнал наук о Земле . 46 (11): 791–800. Бибкод : 2009CaJES..46..791E . дои : 10.1139/E09-050 .
- Эванс, округ Колумбия; Ларсон, Д.В.; Каллен, ТМ; Салливан, РМ (2014). Сьюс, Ханс-Дитер (ред.). « Saurornitholestes»robustus — троодонтид (Dinosauria: Theropoda)». Канадский журнал наук о Земле . 51 (7): 730–734. Бибкод : 2014CaJES..51..730E . doi : 10.1139/cjes-2014-0073 .
- Фарке, А.А.; Чок, диджей; Кузнец, А.; Скольери, Б.; Вернинг, С. (2013). Хатчинсон, Джон (ред.). «Онтогенез трубчатохохлого динозавра Parasaurolophus (Hadrosauridae) и гетерохрония у гадрозаврид» . ПерДж . 1 : е1 дои : 10.7717/peerj.182 . ПМЦ 3807589 . ПМИД 24167777 .
- Гилмор, Чарльз В. (1924). «О роде Stephanosaurus , с описанием типового экземпляра Lambeosaurus lampei , Паркс». Бюллетень геологической службы Министерства горнодобывающей промышленности Канады (геологическая серия) . 38 (43): 29–48.
- Перенасыщение, Д.Ф. (1997). « Паразауролоф » . В «Перенасыщении», Дональд Ф. (ред.). Динозавры: Энциклопедия . МакФарланд и компания. стр. 678–940. ISBN 978-0-899-50917-4 .
- Годфруа, Паскаль; Шукин Зан; Лиён Джин (2000). « Charonosaurus jiayinensis ng, n. sp., динозавр-ламбеозавр из позднего маастрихта на северо-востоке Китая» (PDF) . Comptes Rendus de l'Académie des Sciences, Série IIA . 330 (12): 875–882. Бибкод : 2000CRASE.330..875G . дои : 10.1016/S1251-8050(00)00214-7 .
- Хон, DWE; Нэйш, Д.; Катхилл, IC (2011). «Объясняет ли взаимный половой отбор эволюцию гребней на голове у птерозавров и динозавров?» (PDF) . Летайя . 45 (2): 139–156. дои : 10.1111/j.1502-3931.2011.00300.x . Архивировано из оригинала (PDF) 6 октября 2014 г.
- Хопсон, Дж. А. (1975). «Эволюция структур черепного дисплея у динозавров-гадрозаврид». Палеобиология . 1 (1): 21–43. Бибкод : 1975Pbio....1...21H . дои : 10.1017/S0094837300002165 . JSTOR 2400327 . S2CID 88689241 .
- Хорнер, Дж.А.; Вейшампель, Д.Б.; Форстер, Калифорния (2004). «Гадрозавриды». В Вейшампеле, Дэвид Б.; Осмольска, Гальшка; Додсон, Питер (ред.). Динозаврия (Второе изд.). Издательство Калифорнийского университета. стр. 438–463. ISBN 978-0-520-24209-8 .
- Ясински, SE; Салливан, РМ (2011). «Повторная оценка пахицефалозаврид из перехода Фрутленд-Киртланд (Киртланд, поздний кампан), бассейн Сан-Хуан, Нью-Мексико, с описанием нового вида Stegoceras и повторной оценкой Texascephale langstoni ». Музей естественной истории и науки Нью-Мексико, Бюллетень . Ископаемая запись 3. 53 : 202–215.
- Лидделл, Генри Джордж и Роберт Скотт (1980). Греко-английский лексикон (сокращенное изд.). Соединенное Королевство: Издательство Оксфордского университета. ISBN 978-0-19-910207-5 .
- Лонгрич, Северная Каролина (2011). « Titanoceratops Ouranous , гигантский рогатый динозавр из позднего кампана Нью-Мексико» (PDF) . Меловые исследования . 32 (3): 264–276. Бибкод : 2011CrRes..32..264L . дои : 10.1016/j.cretres.2010.12.007 .
- Лулл, RS ; Райт, штат Невада (1942). Гадрозавровые динозавры Северной Америки . Специальный доклад Геологического общества Америки 40 . Геологическое общество Америки. п. 229.
- Мартин, Эй Джей (2014). Динозавры без костей: жизнь динозавров, обнаруженная по их окаменелостям . Книги Пегаса. п. 42. ИСБН 978-1-60598-499-5 .
- Марьянская, Т.; Осмольска, Х. (1979). «Аспекты черепной анатомии гадрозавров». Летайя . 12 (3): 265–273. Бибкод : 1979Лета..12..265М . дои : 10.1111/j.1502-3931.1979.tb01006.x .
- Норман, Дэвид Б. (1985). «Гадрозавриды II». Иллюстрированная энциклопедия динозавров: оригинальный и убедительный взгляд на жизнь в царстве динозавров . Нью-Йорк: Crescent Books. стр. 122–127. ISBN 978-0-517-46890-6 .
- Остром, Дж. Х. (1961). «Новый вид гадрозаврового динозавра из мелового периода Нью-Мексико». Журнал палеонтологии . 35 (3): 575–577. JSTOR 1301139 .
- Остром, Джон Х. (1962). «Черепные гребни гадрозавровых динозавров». Постилла . 62 : 1–29.
- Паркс, Вашингтон (1922). « Parasaurolophus walkeri , новый род и вид траходонтных динозавров» . Исследования Университета Торонто: Геологическая серия . 13 :5–32.
- Робертс, EM; Дейно, Алабама; Чан, Массачусетс (2005). «40Ar/39Ar возраст формации Кайпаровиц, южная Юта, и корреляция одновременных слоев кампана и фауны позвоночных вдоль окраин Западного внутреннего бассейна». Меловые исследования . 26 (2): 307–318. дои : 10.1016/j.cretres.2005.01.002 .
- Ромер, Альфред Шервуд (1933). Палеонтология позвоночных . Издательство Чикагского университета. п. 491. OCLC 1186563 .
- Национальные лаборатории Сандии (5 декабря 1997 г.). «Ученые используют цифровую палеонтологию для создания голоса динозавра -паразауролофа » . Сандианские национальные лаборатории. Архивировано из оригинала 17 августа 2014 года.
- Симпсон, ДП (1979). Латинский словарь Касселла (5-е изд.). Лондон: Cassell Ltd., с. 883. ИСБН 978-0-304-52257-6 .
- Штернберг, Чарльз М. (1935). «Хадрозавры серии Белли-Ривер верхнего мела». Бюллетень Министерства горнодобывающей промышленности Канады (геологическая серия) . 77 (52): 1–37.
- Салливан, РС; Уильямсон, Т.Э. (1996). «Новый череп паразауролофа (форма с длинным гребнем) из Нью-Мексико: внешние и внутренние (компьютерная томография) особенности и их функциональное значение». Журнал палеонтологии позвоночных . 16 (3): 1–68. дои : 10.1080/02724634.1996.10011371 .
- Салливан, РС; Уильямсон, Т.Е. (1999). «Новый череп Parasaurolophus (Dinosauria: Hadrosauridae) из формации Киртланд в Нью-Мексико и пересмотренная версия рода» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 15 :1–52. Архивировано из оригинала (PDF) 8 апреля 2023 г.
- Салливан, РМ; Лукас, СГ (2006). «Возрастной состав наземных и позвоночных животных Киртландии, временное положение и биостратиграфическая корреляция в неморском верхнем мелу западной части Северной Америки» . В Лукасе, СГ; Салливан, Р.М. (ред.). Позднемеловые позвоночные Западного Внутреннего региона . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 35. стр. 7–23.
- Салливан, RS; Ясински, SE; Гюнтер, М.; Лукас, СГ (2011). Салливан, Роберт С.; Лукас, Спенсер Г. (ред.). «Ископаемая запись 3: первый« ламбеозавр » (Dinosauria, Hadrosauridae, Lambeosaurinae) из верхнемеловой формации Охо Аламо (пачка Наашойбито), бассейн Сан-Хуан, Нью-Мексико» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 53 : 405–417. Архивировано из оригинала (PDF) 6 октября.
- Салливан, РМ; Фаулер, Д.В. (2011). « Navajodactylus boerei , n. gen., n. sp., (Pterosauria, ?Azhdarchidae) из верхнемеловой формации Киртланд (верхний кампан) в Нью-Мексико» (PDF) . Ископаемая запись 3. Музей естественной истории и науки Нью-Мексико, Бюллетень . 53 : 393–404.
- Танке, DH; Карпентер, К., ред. (2001). Жизнь мезозойских позвоночных . Издательство Университета Индианы. стр. 206–328 . ISBN 978-0-253-33907-2 .
- Титус, Алабама; Лоуэн, Массачусетс, ред. (2013). На вершине парадной лестницы: поздний мел Южной Юты . Издательство Университета Индианы. стр. 1–634.
- Вейшампель, Д.Б.; Дженсен, Дж. А. (1979). « Паразауролоф (Reptilia: Hadrosauridae) из Юты». Журнал палеонтологии . 53 (6): 1422–1427. JSTOR 1304144 .
- Вейшампель, Д.Б. (1981). «Акустический анализ вокализации динозавров-ламбеозавров (Reptilia: Ornithischia)» (PDF) . Палеобиология . 7 (2): 252–261. Бибкод : 1981Pbio....7..252W . дои : 10.1017/S0094837300004036 . JSTOR 2400478 . S2CID 89109302 . Архивировано из оригинала (PDF) 6 октября 2014 г.
- Вейшампель, Д.Б. (1997). «Какофония динозавров: определение функции вымерших организмов» . Бионаука . 47 (3): 150–155. дои : 10.2307/1313034 . JSTOR 1313034 .
- Вейшампель, Дэвид Б.; Барретт, Пол М.; Кориа, Родольфо А.; ЛеЛёфф, Жан; Сюй Син; Чжао Сицзинь; Сахни, Ашок; Гомани, Элизабет, член парламента; Ното, Кристофер Р. (2004). «Распространение динозавров». Динозаврия (2-е изд.). стр. 517–606.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Уиллер, ЧП (1978). «Развитая структура охлаждения ЦНС у крупных динозавров». Природа . 275 (5679): 441–443. Бибкод : 1978Natur.275..441W . дои : 10.1038/275441a0 . ПМИД 692723 . S2CID 4160470 .
- Уилфарт, Мартин (1947). «Тоботоносные динозавры». Орион (Мюнхен) (на немецком языке). 2 :525-532.
- Уильямсон, Т.Е. (2000). Лукас, Спенсер Г.; Хеккерт, Эндрю Б. (ред.). «Динозавры Нью-Мексико: обзор Hadrosauridae (Dinosauria: Ornithischia) из бассейна Сан-Хуан, Нью-Мексико». Бюллетень Музея естественной истории и науки Нью-Мексико . 17 : 191–213.
- Виман, К. (1931). « Parasaurolophus Tubeicen , n. sp. aus der Kreide в Нью-Мексико». Новые труды Королевского общества наук Уппсалы . Серия 4 (на немецком языке). 7 (5): 1–11.
- Син, Х.; Ван, Д.; Хан, Ф.; Салливан, К.; Ма, Кью; Привет.; Хон, DWE; Ян, Р.; Ду, Ф.; Сюй, X. (2014). Эванс, Дэвид С. (ред.). «Новый базальный гадрозавроидный динозавр (Dinosauria: Ornithopoda) с переходными особенностями из позднего мела провинции Хэнань, Китай» . ПЛОС ОДИН . 9 (6): e98821. Бибкод : 2014PLoSO...998821X . дои : 10.1371/journal.pone.0098821 . ПМК 4047018 . ПМИД 24901454 .
- Занно, Ле; Сэмпсон, С.Д. (2005). «Новый овирапторозавр (Theropoda; Maniraptora) из позднего мела (кампан) штата Юта». Журнал палеонтологии позвоночных . 25 (4): 897–904. doi : 10.1671/0272-4634(2005)025[0897:ANOTMF]2.0.CO;2 . S2CID 131302174 .
Внешние ссылки
[ редактировать ]- Штраус, Боб (2014). «Десять фактов о паразауролофах » . О.Com Динозавры. Архивировано из оригинала 7 апреля 2015 года . Проверено 5 октября 2014 г.
- «Звук паразауролофа» . Сандианские национальные лаборатории. 5 декабря 1997 г. Архивировано из оригинала 6 октября 2014 г.
- Хартман, Скотт (2004). «Порнитисхи: Parasaurolophus cyrtocristatus » . Рисунок скелета Скотта Хартмана.
- Хартман, Скотт (2013). «Порнитисхи: Parasaurolophus walkeri » . Рисунок скелета Скотта Хартмана.
 СМИ, связанные с паразауролофом, на Викискладе?
СМИ, связанные с паразауролофом, на Викискладе?  Данные, относящиеся к паразауролофу , в Wikispecies
Данные, относящиеся к паразауролофу , в Wikispecies

- Позднемеловые динозавры Северной Америки.
- Ламбеозаврины
- Ископаемые таксоны описаны в 1922 году.
- Таксоны, названные Уильямом Парксом
- Фауна Киртланда
- Фауна парка динозавров
- Палеонтология в Альберте
- Палеонтология в Нью-Мексико
- Палеонтология в Юте
- Кайпаровицкая свита
- Первые появления кампанского рода
- Орнитишские роды
- Маастрихтское вымирание родов
- Многовидовые нептичьи роды динозавров
- Орнитоподы Северной Америки