Эоламбия
| Эоламбия Временной диапазон: сеноман ,
| |
|---|---|
| |
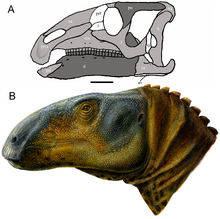
| Реконструкция черепа (А) и восстановление жизни (Б) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Орнитоподы |
| Суперсемейство: | † Хадрозавроида |
| Клэйд : | † Гадрозауроморфа |
| Род: | † Эоламбия Киркланд , 1998 г. |
| Типовой вид | |
| † Эоламбия карольхонесская Киркланд, 1998 г.
| |
Эоламбия рассветный ламбеозавр ») — род травоядных гадрозавроидных динозавров из раннего позднего мела США (что означает « . Он содержит единственный вид, E. caroljonesa , названный палеонтологом Джеймсом Киркландом в 1998 году. экземпляр Eolambia Типовой был обнаружен Кэрол и Рамалем Джонс в 1993 году; название вида дано в честь Кэрол. С тех пор были обнаружены сотни костей как взрослых, так и молодых людей, представляющих почти каждый элемент скелета. Все образцы на данный момент были найдены в округе Эмери , штат Юта , в слое породы, известном как член Муссентучит формации Кедровая гора .
Имея длину до 6 метров (20 футов), Эоламбия является крупным представителем своей группы. и конечностей он близко приближается к азиатским гадрозавроидам Equijubus , Probactrosaurus и Choyrodon Хотя по чертам черепа, позвонков , на самом деле он может быть более тесно связан с североамериканским протохадросом . Эта группа, основанная на прямолинейности квадратной кости и лопатки , представляет собой изолированную эндемическую группу гадрозавроидов. Несмотря на сходство с гадрозавридами - в частности с гадрозавридами ламбеозаврид - по некоторым признакам, что привело к его первоначальной идентификации как одного из них, эти сходства были отвергнуты как полностью конвергентные или неверно истолкованы.
Эоламбия жила в лесной среде на краю озер во влажной пойменной среде, питаясь голосеменными растениями , папоротниками и цветковыми растениями . Уровни воды в озерах со временем менялись с циклическими периодами дождей и засух, вызванными прецессией Земли , что отражается чередующимися полосами в отложениях пачки Муссентучит. В молодости Эоламбия стала жертвой крупных крокодиломорфов, обитающих в водах озера. Однако с возрастом они стали невосприимчивы к крокодиломорфам, а взрослые особи (в возрасте от восьми до девяти лет) стали жертвами крупных теропод, таких как неовенаториды Сиатс .
История открытия
[ редактировать ]Первоначальные открытия и описание
[ редактировать ]
В 1979 году Питер Гальтон и Джеймс А. Дженсен описали фрагментарную правую бедренную кость, BYU 2000, принадлежащую гадрозавровому динозавру, обнаруженную в отложениях формации Сидар-Маунтин в национальном парке Арчес , штат Юта . Несмотря на плохой материал, он был важен, поскольку (наряду со второй североамериканской бедренной костью, описанной в статье) был первым экземпляром гадрозавра из нижнего мела в любой точке мира. Гальтон и Дженсен предположили, что в будущем в этом пласте можно будет найти более полные останки гадрозавра. [ 1 ] Различные зубы гадрозавроидов также были обнаружены в карьерах мелких позвоночных в западном регионе Сан-Рафаэль-Суэлл , недалеко от Касл-Дейл в округе Эмери , штат Юта; они были описаны в 1991 году Дж. Майклом Пэрришем. Впоследствии, в 1993 году, Кэрол Джонс и ее муж Рамал Джонс обнаружили фрагментарные кости на окаменелом участке, расположенном в северо-западном районе Свелла. Они привлекли к этому месту внимание Дональда Берджа, директора учреждения, которое тогда называлось Колледжем доисторического музея Восточной Юты (CEUM). Это место, официально известное как CEUM Locality 42em366v, впоследствии будет названо «Участком Кэрол» ( так в оригинале ) в ее честь. Окаменелости, хранящиеся под номером экземпляра CEUM 9758, представляют собой частичные останки взрослого гадрозавроида, включая части черепа, позвонков, седалищной кости и ноги. CEUM 5212, частичный череп и передняя конечность взрослого человека, были найдены неподалеку в местонахождении CEUM 42em369v. [ 2 ] CEUM 8786, левая бедренная кость взрослого человека, была обнаружена позже на Участке Кэрол и не описывалась до 2012 года. [ 3 ]
В то же время Музей естественной истории Оклахомы (OMNH) обнаружил останки, принадлежащие тому же гадрозавроиду, из раскопок в юго-западном районе Свелла. [ 2 ] Образцы, обнаруженные OMNH, первоначально состояли из шести особей из пяти местонахождений: два молодых скелета, [ 4 ] включая позвонки, лопатки, локтевую кость, подвздошную и седалищную кости, а также элементы задних конечностей из местонахождения OMNH v237; частичный ювенильный скелет, включающий части черепа, передних и задних конечностей, а также спинной позвонок из OMNH v824; OMNH 27749, крестец и седалищная кость из OMNH v696; OMNH 24389, седалищная кость из OMNH v214; и OMNH 32812, частичный скелет, включающий лопатку, два хвостовых позвонка и другие нераскопанные элементы из OMNH v866. OMNH также совершила дальнейшие открытия зубов гадрозавроидов в различных карьерах. Ричард Сифелли из OMNH позволил Джеймсу Киркланду изучить собранные образцы. [ 2 ] Киркланд продолжил предварительное описание образцов в 1994 году в тезисах конференции на ежегодном собрании Общества палеонтологии позвоночных . [ 5 ]

В статье 1998 года, опубликованной в бюллетене Музея естественной истории и науки Нью-Мексико , Киркланд официально назвал эти экземпляры новым родом , Eolambia типом Eolambia и единственным видом которого является caroljonesa . Родовое название сочетает в себе греческий префикс eos /ἠώς («рассвет», «утро», подразумевающий «рано») с суффиксом лямбия (происходящий от Lambeosaurus , который, в свою очередь, назван в честь канадского палеонтолога Лоуренса Ламбе ). В целом название рода означает «рассветный (или ранний) ламбеозаврин », имея в виду его предполагаемое положение как самого базального ламбеозаврина. Между тем, видовое имя дано в честь Кэрол Джонс. Название Эоламбия было предложено палеохудожником Майклом Скрепником ; оно заменило неофициальное название «Eohadrosaurus caroljonesi», которое использовалось Киркландом до публикации статьи 1998 года. [ 2 ] Однако эпитет caroljonesa , следующий за ICZN , неверен, и в этом случае правильным написанием будет caroljonesae . [ 6 ]
Последующие открытия
[ редактировать ]Дополнительные раскопки, проведенные с 1998 года, выявили дополнительные элементы скелета, принадлежащие Эоламбии . Сейчас это наиболее известный игуанодонт между сеноманским и сантонским ярусами. Почти каждый элемент скелета, принадлежащий таксону, представлен множеством окаменелостей в существующих коллекциях. Не менее двенадцати человек, в том числе не менее восьми несовершеннолетних, [ 7 ] были обнаружены в карьере к югу от города Эмери , включая части черепа и остальной части тела. Карьер известен как карьер Cifelli № 2 «Eolambia caroljonesa», или, более формально, местонахождение CEUM 42em432v. В другом карьере к югу от города Эмери – карьер Уиллоу-Спрингс 8, или местонахождение CEUM 42Em576v – сохранилось как минимум четыре молодых особи. Материал из этих карьеров был описан в 2012 году Киркландом вместе с Эндрю Макдональдом, Джоном Бердом и Питером Додсоном, которые также предоставили обновленный диагноз Эоламбии . на основе этого материала [ 3 ]
Один экземпляр, обнаруженный в карьере Чифелли № 2, правая зубная кость, каталогизированная как CEUM 34447, необычен по сравнению с другими экземплярами Eolambia , включая как взрослых, так и молодых особей. В то время как соотношение между глубиной средней точки и общей длиной зубных рядов у других людей обычно колеблется от 0,19 до 0,24, то же значение у CEUM 34447 составляет 0,31, что делает его необычайно глубоким. Зубная кость также существенно не углубляется на переднем конце и, таким образом, лишена одной из отличительных черт зубных костей Eolambia . Однако у экземпляра присутствует другой отличительный признак — расширение переднего конца венечного отростка. Хотя вполне возможно, что этот человек представляет собой необычный экземпляр Eolambia , Макдональд и его коллеги предупредили, что он может представлять собой отдельный гадрозавроид, который отличается морфологией зубной кости от Eolambia (хотя и не предполагается ни одним из других материалов в карьере). ). Таким образом, они считали его неопределенным базальным гадрозавроидом. [ 3 ]

Полевой музей естественной истории (FMNH) также проводил раскопки к югу от города Эмери с 2009 по 2010 год с разрешения Геологической службы штата Юта . Эти раскопки проводились на участке, обнаруженном Акико Шинья в 2008 году, в местности FMNH UT080821-1, которая была названа «Участком Акико» в ее честь. В ходе раскопок в двух гипсовых оболочках было собрано 167 разрозненных, но тесно связанных друг с другом элементов. Одной из этих гипсовых курток, содержавшей элементы бедра и позвонков, был присвоен номер экземпляра FMNH PR 3847; впоследствии образец был описан Макдональдом вместе с Терри Гейтсом, Линдси Занно и Питером Маковицки в 2017 году. Еще одно место, обнаруженное FMNH, - это карьер Трипл-Пик, местоположение FMNH UT130904-2, который содержит костное ложе из более чем 400 молодых костей, принадлежащих как минимум четырем лицам. [ 8 ]
Геология и знакомства
[ редактировать ]
Все местонахождения, в которых были обнаружены Эоламбии , относятся к муссентучитской пачке кедровой горной свиты . Преобладающие породы Муссентучита состоят из переслаивающихся слоев от светло-серого до серого мутного песчаника и аргиллита . отложения, представляющие собой две отдельные системы русел рек В Муссентучите также присутствуют . Нижняя из этих систем состоит из четырех слоев соответственно илистого песчаника, мелкозернистого песчаника, алевритового аргиллита и мелкозернистого песчаника, причем два последних слоя содержат растительные остатки. С этой системой связан карьер Чифелли №2. Верхняя система сложена зеленоватым мелкозернистым песчаником. три слоя слоев бентонитовой глины , представляющие собой отложения вулканического пепла В Муссентучите также присутствуют , имеющие соответственно беловато-серый, желтый и коричневый цвет. Середина этих слоев пересекает карьер Чифелли №2. [ 7 ] Бентонит также присутствует в самом песчанике по всему Муссентучиту. [ 8 ] В карьере Чифелли №2 сами кости почернели, заместившись кальцитом , кварцем , пиритом и карбонатом - гидроксиапатитом . [ 7 ]
Палинология (изучение пыльцы), [ 9 ] [ 10 ] [ 11 ] а также исследования ископаемых мегафлоры и беспозвоночных, [ 12 ] первоначально предположил, что самые верхние кедровые горные формации, включая Муссентучит, относятся к альбскому ярусу. [ 2 ] , извлеченных из слоя вулканического пепла в верхней части формации Кедровой горы, проведенное Чифелли и его коллегами в 1997 году Аргон-аргоновое датирование кристаллов санидина , показало возраст 97 ± 0,1 миллиона лет, который тогда считался сразу после границы между альбом и сеноманом. этапы. Это похоже на ранний сеноманский возраст, указанный для вышележащей формации Дакота . [ 13 ] Впоследствии, в 2007 году, Джеймс Гаррисон и его коллеги датировали слой вулканического пепла, проходящий через карьер Чифелли № 2 (который, вероятно, является тем же слоем, датированным ранее Чифелли и его коллегами), 96,7 ± 0,5 миллиона лет назад. На приблизительном уровне оценки Чифелли и Гаррисона согласуются друг с другом, что указывает на сеноманский возраст Эоламбии . Гаррисон и его коллеги также повторно проанализировали комплекс пыльцы из местностей по всему Муссентучиту и пришли к выводу, что этот комплекс охватывает границу альба и сеномана. [ 7 ]
Описание
[ редактировать ]
Эоламбия — крупный гадрозавроид . По первоначальным оценкам длина его черепа составляла 1 метр (3 фута 3 дюйма). [ 2 ] хотя это произошло из-за непропорционально длинной морды, которая позже была исправлена открытием дополнительного материала. [ 3 ] В 2016 году Грегори С. Пол в 6 метров (20 футов) и вес в 1 метрическую тонну (0,98 длинных тонны; 1,1 коротких тонны) оценил длину тела Эоламбии . [ 14 ] что согласуется с предыдущей оценкой длины тела Томаса Р. Хольца-младшего в 2012 году в 6,1 метра (20 футов). [ 15 ] Ранее, в 2008 году, взрослая особь имела длину 5,2 метра (17 футов) и высоту в бедрах 2 метра (6 футов 7 дюймов). [ 7 ]
Череп
[ редактировать ]
Череп без гребня Eolambia имеет общую форму, аналогичную черепу Equijubus и Probactrosaurus . Передняя часть морды сильно шероховатая, с множеством отверстий . На кончике каждой предчелюстной кости есть две зубоподобные структуры, известные как зубчики, которые также наблюдаются у его ближайшего родственника Protohadros . Дальше назад задняя часть нижней ветви предчелюстной кости резко выступает вверх, закрывая ноздрю сзади, как у пробактрозавра , протохадроса и других гадрозавроидов. Эта часть соединяется с двумя пальцевидными отростками верхней челюсти , сходной с Протохадросом . На теле самой верхней челюсти нет выемки или каких-либо признаков анторбитального отверстия , как у Equijubus , Protohadros и других гадрозавроидов. Одной из характеристик, используемых для различения эоламбий, является вогнутый профиль зубного ряда верхней челюсти при взгляде сбоку, который подобен Equijubus , Probactrosaurus и некоторым другим гадрозавриформам , но в отличие от Протоадрос . [ 3 ]
Как и у пробактрозавра и других гадрозавроидов, задняя часть верхней челюсти соединяется со скуловой костью , которая граничит с нижней частью глазницы и подвисочным окном , через пальцеобразный выступ, который вписывается в углубление. Нижний край скуловой кости имеет сильный выступ ниже уровня подвисочного окна; это также наблюдается у Equijubus , Probactrosaurus , Protohadros и некоторых других гадрозавроидов. Сверху к скуловой кости соединяется заглазничная кость , имеющая шероховатую поверхность там, где она граничит с глазницами (как у Протохадроса ), но в остальном сторона кости гладкая. В задней части черепа квадратная кость соединяется с чешуйчатой костью с помощью сустава, который имеет D-образную форму, если смотреть сверху. Левая и правая чешуйчатые кости должны были широко контактировать друг с другом, будучи разделены лишь сзади небольшим отростком теменной кости . Надзатылочная кость , образующая верхнюю часть задней части черепа, плоская и почти вертикальная, как в случае с Пробактрозавр и другие гадрозавроиды. [ 3 ] \
Как и предчелюстная кость, имел зубцы предентарий Эоламбии . Имеется заметный дорсомедиальный отросток, язычковая структура, также наблюдаемая у пробактрозавра и других гадрозавриформ. По обе стороны от дорсомедиального отростка присутствовало несколько дополнительных пластинчатых зубчиков, которые также присутствуют у Probactrosaurus . Преддентарий сзади соединяется с дентарием , составляющим большую часть нижней челюсти. Между сочленением предзубной кости с зубной и положением первого зуба на зубной кости имеется короткая выемка, или диастема , которая наблюдается у Equijubus , Probactrosaurus и других гадрозавроидов. Передняя часть зубной кости характерно углубляется, как у протохадроса , уранозавра и бактрозавра . На внешней поверхности зубной кости имеются две выпуклости, одна из которых представляет собой венечный отросток, как у пробактрозавра и других гадрозавроидов. Подобно протохадросу и некоторым другим гадрозавриформам, но в отличие от пробактрозавра , утолщен только передний конец этого отростка. Как в У Equijubus , Probactrosaurus и различных других игуанодонтов на стороне надугловой кости , расположенной позади зубной кости, имеется небольшое отверстие. [ 3 ]
Как и у других игуанодонтов, зубы эоламбии расположены плотно расположенными и переплетающимися рядами. В любой момент времени каждая из 32 лунок верхних зубов содержит три зуба, а каждая из 30 лунок зубных зубов — четыре зуба. Из них два зуба в каждой лунке являются замещающими, как у пробактрозавра ; остальные являются активными зубами. Каждый активный зуб имеет одну фасетку износа . Подобно Probactrosaurus , Gongpoquansaurus , Protohadros и другим гадрозавроидам, коронка каждого зуба верхней челюсти имеет только один гребень, слегка смещенный к средней линии рта. каждого зубного Между тем, коронка зуба обычно имеет первичный гребень и дополнительный гребень ближе к средней линии рта - состояние, которое также присутствует у протохадросов и других гадрозавроидов. Все коронки также имеют небольшие зубчатые зубцы на переднем и заднем краях, что также наблюдается у пробактрозавра . [ 3 ]
Позвоночник
[ редактировать ]
Подобно Equijubus , Probactrosaurus , Gongpoquansaurus и другим стиракостернам , Eolambia имеет шейные позвонки очень опистоцельные , что означает, что их передние концы сильно выпуклые, а задние — сильно вогнутые. Необычно то, что передний конец третьего шейного позвонка расположен немного выше заднего, чего не наблюдается ни в каких других шейных позвонках. Позвонки, расположенные в задней части шеи, имеют больший угол между удлиненными суставными отростками, известными как постзигапофизы, а также более удлиненные поперечные отростки . На поперечных отростках располагаются далее суставные отростки – округлые парапофизы и палочковидные диапофизы. В самых задних шейных отделах также имеется глубокая впадина, разделяющая постзигапофизы, а нервные отростки имеют круто наклоненные передние края, что приводит к созданию заметных шипообразных выступов. [ 3 ] [ 8 ]
Первые несколько спинных позвонков похожи на шейные, но имеют более высокие и зубчатые нервные отростки. Остальные амфиплатаны, то есть плоские с обоих концов. У них также есть постзигапофизы, которые менее удлинены и более похожи на пьедестал, а также более высокие и прямоугольные нервные отростки. Парапофизы, которые представляют собой углубления, а не выступы, переместились от поперечных отростков к нервной дуге , между поперечными отростками и суставными отростками, известными как презигапофизы, в передней части позвонков. Однако в последних нескольких дорсальных отделах они возвращаются к основанию поперечных отростков. [ 3 ] В третьем или четвертом дорсальном отделе парапофиз расположен очень близко к шву между нервным отростком и центром , что не похоже на другие дорсальные кости, но похоже на первые несколько дорсальных костей у эдмонтозавра . Также, как и у Эдмонтозавра , срединный гребень, разделяющий презигапофизы, становится более выраженным в задней части спины. [ 8 ]
Среди найденных на сегодняшний день экземпляров Эоламбии наиболее хорошо сохранившийся крестец включает семь позвонков. Учитывая, что этот человек незрелый, а у гадрозавров с возрастом увеличивается количество крестцовых позвонков, у взрослых особей могло быть больше крестцовых позвонков. [ 2 ] Хвостовые позвонки амфицельные , их передний и задний концы вогнуты. У них есть пьедесталообразные презигапофизы и пластинчатые постзигапофизы, последние из которых разделены углублением. Презигапофизы удлиняются, превращаясь в стебли, а постзигапофизы сжимаются в средней и задней каудальной части. При этом нервные отростки из прямоугольных переходят в сильно изогнутые (вогнутые спереди и выпуклые сзади), центры становятся более удлиненными, а поперечные отростки исчезают. [ 3 ] [ 8 ]
Конечности и пояса конечностей
[ редактировать ]
Как и у почти прямоугольное , его Probactrosaurus, лопаточное лезвие Eolambia стороны прямые и сходятся с концом лезвия под почти прямым углом. изогнута Плечевая кость к средней линии, а дельтопекторальный гребень на плечевой кости заметно утолщен. На внешней поверхности сустава с лучевой костью на плечевой кости имеется выступ. И локтевая , и лучевая кость представляют собой относительно прямые кости. Верхняя часть локтевой кости имеет три «зубца», а именно локтевой отросток и два дополнительных треугольных фланца. Между тем, лучевая кость имеет уплощенную внутреннюю поверхность на верхнем конце и отчетливую платформу на внутренней поверхности на нижнем конце, обе для сочленения с локтевой костью. На руке пять пальцев. Среди пястных костей кисти вторая изогнута, третья прямая и четвертая сильно изогнута. Как и у игуанодона , пробактрозавра и других базальных игуанодонтов, коготь на первом пальце имеет отчетливо коническую форму. Вторая и третья клешни похожи на копыта, причем третья коготь имеет шероховатые края по бокам – черта, присущая и другим игуанодонтам. [ 3 ]
Преацетабулярный (т. е. расположенный перед тазобедренным суставом ) отросток подвздошной кости несет горизонтальный «ботинок», образующий тупой угол с основанием отростка. Кроме того, верхний край кости выступает наружу, образуя ободок, расположенный над подвздошно-седалищным сочленением на нижнем крае кости. Эти черты также наблюдаются у пробактрозавра и других игуанодонтов. [ 3 ] Сама тазобедренная впадина у игуанодонта довольно велика по сравнению с подвздошной костью. [ 4 ] Вдоль верхней поверхности подвздошной кости присутствует характерный выступ, характерный для безымянного гадрозавроида из формации Вудбайн . Постацетабулярный (т. е. расположенный позади тазобедренного сустава) отросток подвздошной кости сужается сзади без разрыва ее верхнего края. На лобке выступающий вперед отросток расширяется кпереди, как у Probactrosaurus , Gongpoquansaurus и других игуанодонтов. Стержень седалищной кости у взрослых характерно прямой (у молодых особей изогнутый стержень, что чаще встречается у игуанодонтов), а нижняя часть стержня несет сжатый «сапог», расширенный вперед. [ 3 ] [ 8 ]
Как и плечевая кость, бедренная кость изогнута, но наружу. Однако характерно то, что нижняя половина бедра прямая. Четвертый вертел , расположенный на середине бедренной кости, вертикально высокий и треугольной формы, похож на пробактрозавра , гонгпоканзавра и других игуанодонтов, но отличается от более базальных орнитопод . Подобно локтевой и лучевой кости, голени и малоберцовые кости имеют прямые стержни и сочленяются друг с другом; шероховатый выступ на дне малоберцовой кости находится между двумя луковицеобразными мыщелками, расположенными сразу за кнемиальным гребнем на нижнем конце большеберцовой кости. Верх голени шероховатый, что указывает на наличие хрящевой крышки между бедренной и большеберцовой костью. Стопа имеет четыре пальца. Как и пястные кости, вторая плюсневая кость изогнута, третья — прямая, четвертая — сильно изогнута. Фаланги . стопы очень толстые, причем концевые фаланги на концах каждого пальца особенно толстые Как и вторые и третьи когти на руке, вторые, третьи и четвертые когти стопы одинаково широкие, плоские и напоминают копыта. [ 3 ]
Классификация
[ редактировать ]Первоначальная идентификация гадрозаврид
[ редактировать ]Киркландом В первоначальном описании Эоламбии он считал ее представителем Hadrosauridae , как это было определено Дэвидом Б. Вейшампелем , Дэвидом Б. Норманом и Дэном Григореску в 1993 году. Вейшампель и его коллеги использовали семь объединяющих характеристик для определения Hadrosauridae: расширение восходящего отростка верхней челюсти вверх; отсутствие параквадратного отверстия, разделяющего квадратную и квадратноскуловую кости ; расположение углов на внутренней поверхности нижней челюсти; отсутствие надугольного отверстия на надугольной кости; узкие зубы верхней челюсти; наличие трех и более зубов в каждой зубной позиции; и уменьшение верхнего края лопатки. [ 16 ] Первая, пятая, шестая и седьмая из этих черт были обнаружены в Эоламбии , а остальные неизвестны из-за отсутствия материала. Киркланд далее отнес эоламбию к Euhadrosauria, которую Вейшампель и его коллеги определили как включающую общего предка Hadrosaurinae (ныне Saurolophinae) и Lambeosaurinae — двух основных ветвей гадрозаврид — и всех их потомков. [ 16 ] Это определение было основано на пяти характеристиках, которые присутствуют у Эоламбии : наличие зубчиков на предчелюстной кости; узкий сустав квадратной кости с нижней челюстью; узкие зубные зубы; наличие единственного края, или киля, на каждом зубном зубе; и наклонный дельтопекторальный гребень на плечевой кости. [ 2 ]
В пределах Hadrosauridae Киркланд далее считал Eolambia либо базальным представителем Lambeosaurinae, либо сестринской группой Lambeosaurinae. Он выделил пять характеристик, общих с Lambeosaurinae, определенных в различных исследованиях: [ 16 ] [ 17 ] [ 18 ] [ 19 ] отсутствие отверстия на предчелюстной кости; закрытие (по крайней мере частичное) ноздри предчелюстной костью; развитие полки на верхней челюсти; очень высокие нервные отростки хвостовых позвонков; прочность плечевой кости; и большой «сапог» седалищной кости у взрослых. Киркланд обнаружил, что две дополнительные черты отличают бесгребешков Eolambia от других хохлатых ламбеозавринов, которые считались связанными с развитием гребня: возвышение носовой полости над глазницей и укорочение теменной кости. Если бы Eolambia была признана ламбеозаврином, то Lambeosaurinae пришлось бы дать новое определение, чтобы исключить эти два признака. С другой стороны, если бы Eolambia была сестринской группой Lambeosaurinae, то она представляла бы морфологию гадрозаврид, близкую к расхождению между гадрозавринами и ламбеозавринами. Эта возможность подтверждается наличием бороздки на дне крестца Эоламбии , которая была признана Киркландом определяющим признаком гадрозавринов. [ 18 ] [ 19 ] - хотя один из них конвергентно присутствует у Ankylosauria и Ceratopsia . [ 2 ]
Чтобы подтвердить родство эоламбии с ламбеозаврами , Киркланд провел филогенетический анализ ; дерево , полученное в результате его анализа, который, соответственно, делает эоламбию самым базальным ламбеозаврином, воспроизведено ниже. Несмотря на доказательства, которые он привел в поддержку своей позиции, Киркланд также отметил, казалось бы, противоречивую черту. Как наблюдалось у ювенильных подвздошных костей, которые он первоначально описал, антитрохантер – выступ на подвздошной кости, ограничивающий большой вертел бедренной кости – развит хуже, чем можно было бы ожидать у базальных гадрозаврид. Майкл Бретт-Сурман отметил, что у взрослых особей, возможно, был более развит антивертел. С другой стороны, производный ламбеозаврин Hypacrosaurus имеет хорошо развитый антитрохантер даже на стадии вылупления. Киркланд отметил возможность того, что у базальных гадрозавридов свой антитрохантер развился позже в жизни, чем у производных гадрозавридов, таких как гипакрозавр . [ 20 ] Таким образом, он отметил, что взрослые экземпляры Eolambia решат эту проблему. [ 2 ]


| |||||||||||||||||||||||||||||||||||||||||||||||||
Повторная идентификация как негадрозаврид
[ редактировать ]
В 2001 году Джейсон Хед переоценил филогенетическое положение эоламбии на основе образцов, хранящихся в OMNH. Он признал идентификацию Киркландом предполагаемых отличительных черт ламбеозаврин в Эоламбии либо ошибочной, либо сомнительной. Как показал Хэд, на предчелюстной кости действительно есть отверстие; предчелюстная кость граничит с ноздрей только по нижнему краю; на передней части верхней челюсти помимо полки имеются два пальцевидных отростка; высокие нервные отростки хвостовых позвонков встречаются также у неламбеозавринов Ouranosaurus и Bactrosaurus ; а расширение кончика седалищной кости также встречается у камптозавра , игуанодона , уранозавра , гильмореозавра и бактрозавра , что указывает на то, что расширение является плезиоморфным для игуанодонтов и вторично утрачено у гадрозавринов. [ 19 ] В то время как М. Л. Казановас и его коллеги в 1999 году охарактеризовали ламбеозавринов как имеющие «расширенную стопу» на кончике седалищной кости, в отличие от «булавоподобного» состояния, [ 21 ] Хед утверждал, что коритозавр и ламбеозавр [ 22 ] седалищные кончики не более расширены, чем у Mantellisaurus или Ouranosaurus . [ 4 ] [ 23 ]
Хед также продемонстрировал, что зубы Эоламбии были шире и асимметричнее, чем предполагал Киркланд; он также отметил, что у молодых особей на каждую позицию зуба было только два зуба, в отличие от гадрозаврид. [ 4 ] хотя взрослые экземпляры на самом деле имеют три или более положений зубов, как первоначально продемонстрировал Киркланд. [ 3 ] Далее Хед показал, что у Эоламбии было семь крестцовых позвонков, в отличие от восьми или более у производных гадрозаврид, и что ее антитрохантер на самом деле был развит хуже, чем у Уранозавра и Алтирхинуса . [ 24 ] Исследование образцов OMNH также показало, что постацетабулярная область подвздошной кости не образовала отдельного отростка, а сочленения в нижней части бедренной кости не очень заметны, что отличает Eolambia от более производных игуанодонтов, таких как гадрозавриды. Таким образом, Хед пришел к выводу, что Eolambia была базальным представителем Hadrosauroidea, помимо Hadrosauridae, и этот вывод подтверждается филогенетическим анализом, показавшим, что это негадрозавроидный гадрозавроид, тесно связанный с Probactrosaurus . [ 4 ]
Последующие исследования
[ редактировать ]
включил Эоламбию В 2004 году Норман впервые в комплексный филогенетический анализ игуанодонтов как часть главы, посвященной игуанодонтам, во втором издании «Динозаврии» . Он обнаружил, что Eolambia была сестринской группой Altirhinus , причем обе они были более базальными, чем Protohadros , Probactrosaurus или Hadrosauridae. [ 25 ] В том же томе Хорнер, Вейшампель и Кэтрин Форстер провели отдельный филогенетический анализ гадрозаврид, включая Eolambia . Они снова обнаружили Эоламбию как негадрозавроидного гадрозавроида, хотя и как сестринскую группу Протохадроса . Хотя Хед специально переопределил Hadrosauridae, основываясь на общих характеристиках, включив в него Protohadros . [ 4 ] [ 26 ] Хорнер и его коллеги адаптировали определение, основанное на таксонах, которое исключило Протохадроса и, следовательно, Эоламбию . Они также выявили дополнительные признаки, отличающие эоламбий от гадрозаврид: на зубах зубной кости имеются грубые зубцы, а венечный отросток слабо расширен. [ 27 ]
Различия в восстановленных филогенетических позициях Eolambia сохранялись и в последующие годы. В описании Левнесовии Протохадрос в 2009 году Ханс-Дитер Сьюс и Александр Аверьянов обнаружили, что занимал промежуточное положение относительно Алтирхина и Пробактрозавра , являясь сестринской группой Фукуизавра . [ 28 ] Между тем, в соответствии с результатами Хорнера и его коллег, Альберт Прието-Маркес и Марк Норелл в 2010 году обнаружили, что Эоламбия вместо этого была сестринской группой Протохадроса . Он также отметил черты Eolambia , сходные с гадрозавридами: наличие единственного зубного киля; почти квадратный внешний угол преддентария; а средняя точка квадратно-скуловой вырезки (которая сочленяется с квадратно-скуловой костью) расположена примерно на середине поверхности квадратной кости (находится менее чем на 60% высоты кости вниз от вершины). [ 29 ] В 2012 году Хольц классифицировал Эоламбию как примитивного представителя Стиракостерн наряду с Алтирхинусом . [ 15 ]
В 2012 году Макдональд провел филогенетический анализ игуанодонтов, включив данные из новых экземпляров Eolambia . В отличие от предыдущих авторов, вместо этого он обнаружил, что Эоламбия была сестринской группой пробактрозавра . Эта тесная связь была основана на двух общих характеристиках: квадрат был прямым, за исключением верхнего конца, который изогнут назад; а верхний и нижний края лопаточной пластинки почти параллельны. [ 3 ] Вэньцзе Чжэн и его коллеги добавили Джинтазавра в 2014 году. в группу [ 30 ] а Хосе Гаска и его коллеги восстановили первоначальную группу в 2015 году. [ 31 ] Однако Макдональд тщательно пересмотрел свой набор филогенетических данных при подготовке к описанию образцов FMNH в 2017 году. Как и Хорнер и его коллеги, а также Прието-Маркес и Норелл, Макдональд и его коллеги обнаружили, что Эоламбия была сестринской группой Протохадросов , причем оба они были отнесены к базальным гадрозавроморфам (результаты воспроизведены ниже). [ 8 ] Используя версию набора данных Макдональда 2012 года, Франсиско Верду и его коллеги независимо получили аналогичный результат: [ 32 ] [ 33 ] причем Эоламбия тесно связана с Протохадросом и Джеявати ; однако Макдональд и его коллеги обнаружили, что в 2017 году последний оказался более производным. [ 8 ]

В 2018 году Терри Гейтс и коллеги описали новый игуанодонтийский род Choyrodon , происходящий из того же местонахождения, что и Altirhinus , считавшийся в прошлом родственником Eolambia . Их филогенетический анализ, основанный на матрице Нормана (2015), показал, что Choyrodon является сестринским таксоном Eolambia . Протоадрос Эоламбии Обычно обнаруживаемый родственник не присутствовал в этой матрице. трапециевидной формы Три характеристики подтверждают это сестринское родство: затылок и две общие черты зубов. Расположение этих родов относительно других гадрозавриформных зависело от того, было ли предглазничное отверстие закодировано как присутствующее или отсутствующее у Choyrodon ; у экземпляра ювенильного типа он есть, но нельзя исключать возможность того, что у взрослых особей он будет закрытый. [ 34 ]
Палеобиология
[ редактировать ]Яичная скорлупа, возможно, принадлежащая Eolambia, была обнаружена в различных местах рядом с изолированными зубами. Они имеют сетчатый (сетчатый) рисунок на поверхности и толщину 2 миллиметра (0,079 дюйма). [ 2 ]
Рост
[ редактировать ]
известна по множеству молодых и взрослых экземпляров, изменения, которым Поскольку Eolambia она претерпела по мере роста, хорошо документированы. Количество зубов на верхней челюсти увеличивалось с возрастом: от 23 у подростка до 33 у взрослого. Аналогичное увеличение произошло и в зубных рядах : с 18–22 у молодых людей до 25–30 у взрослых. В то время как у молодых особей отсутствуют вторичные гребни на коронках зубных зубов, у взрослых присутствуют слабые вторичные гребни. [ 3 ] Размер «сапога» седалищной кости также увеличивался с возрастом, как документально подтверждено Киркландом. [ 2 ] Изменения также могли произойти в прямолинейности тела седалищной кости, о чем свидетельствуют образцы, обнаруженные FMNH. [ 8 ]
Макдональд и его коллеги сообщили о результатах гистологического анализа ребра из образца FMNH PR 3847 в 2017 году. обширное ремоделирование кости Во внутреннем матриксе ребра произошло . Медуллярная полость , в которой находится костный мозг , отмечена наличием множества полостей или лакун . На внешней поверхности кости остеоны почти полностью заместили внутреннюю часть кости; послойный рост кости более заметен внутри кости, где пять линий задержки роста видны (LAG). Шестой LAG, возможно, присутствует в медуллярной области, но его размеры относительно ограничены, что делает эту идентификацию сомнительной. [ 8 ]
Поскольку ЛАГ могут быть уничтожены в результате ремоделирования кости, это представляет собой недооценку истинного количества ЛАГ, которые действительно развились; Макдональд и его коллеги оценили три или четыре недостающих LAG, исследуя расстояние между видимыми LAG. Это указывает на то, что на момент смерти животному было восемь-девять лет. Расстояние между ЛАГами значительно уменьшается по направлению к внешней поверхности, что указывает на замедление роста, однако отсутствие внешней фундаментальной системы (серии плотно расположенных ЛАГов) указывает на то, что рост еще не прекратился. Это согласуется с несросшимися швами между центрами и нервными отростками шейных и спинных позвонков, а также с несросшимися швами между центрами крестца и соответствующими им ребрами. [ 35 ] В целом, темпы роста Eolambia , по-видимому, были сопоставимы с темпами роста Hypacrosaurus и Maiasaura , которые достигли зрелости через семь или восемь лет. [ 8 ] [ 36 ] [ 37 ]
Палеоэкология
[ редактировать ]Окружающая среда и тафономия
[ редактировать ]Седиментологические данные пачки Муссентучит указывают на то, что Эоламбия обитала в плохо дренированной пойме , вокруг системы крупных многолетних озер. Уровень воды в озерах колебался между высоким и низким уровнем воды в циклах Миланковича Земли продолжительностью примерно 21 000 лет, что было вызвано осевой прецессией . Эти циклы отражаются в чередующихся слоях илистого песчаника и аргиллита в современных отложениях горных пород. В засушливые периоды обнаженное дно озера образовывало широкий пляж, лишенный растений. Потрескавшаяся грязь на дне озера образовала куски грязи , а органический мусор и трупы были разбросаны по кромке воды по мере того, как озеро постепенно отступало. Между тем, в периоды повышения уровня моря небольшие реки вторгались в пойму, перерабатывая и вытесняя мусор, отложившийся в засушливый сезон. [ 7 ]

Большинство окаменелостей, обнаруженных в Муссентучите, разбросаны и разрознены, а неповрежденные скелеты встречаются относительно редко. Хотя озера активно не разрушали кости, это не особенно способствовало их хорошей сохранности из-за длительности процесса захоронения. Перед захоронением мусорщики могли разрушить скелетный материал, о чем свидетельствуют отрубленные голени Eolambia со следами зубов, обнаруженные в карьере Чифелли №2. Регрессия озера могла время от времени создавать болота , но они не были бы достаточно постоянными, чтобы заманить в ловушку и сохранить более крупных позвоночных. Воздействие водных потоков, по-видимому, сыграло роль в отложении костей эоламбии в карьере Чифелли № 2, где кости в основном откладываются под азимутами 28°, 69°, 93°, 131° и 161° (т.е. относительно северной широты) . ). Эти направления должны были быть параллельны или субпараллельны краям озера, что указывает на ориентацию по озерным течениям, за исключением азимутов 69 ° и 93 °, которые, вероятно, представляют собой речные течения. [ 7 ]
Обилие растительных остатков в сочетании с отсутствием раковинных (плайских) отложений указывает на то, что местный климат во время отложения Муссентучита был относительно влажным. Это резко контрастирует с более ранними отложениями в этом районе, которые являются известковыми (меловыми), что указывает на периоды засушливости продолжительностью до 10 000 лет. Палеоклиматические реконструкции подтверждают эти интерпретации: вторжение моря Моури с севера (процесс, который в конечном итоге сформировал Западный внутренний морской путь ), в результате чего засушливый (внутренние континентальные широты) климат сменился более влажным (средние широты). Восточное побережье) климат. [ 38 ] При палеошироте 46 ° северной широты средняя температура поверхности Муссентучита должна была составлять 40–45 ° C (104–113 ° F), что примерно на 6–8 ° C (43–46 ° F) выше, чем сегодня. , по моделям общего обращения . [ 7 ] [ 39 ]
Экосистема
[ редактировать ]
Крокодиломорфы широко распространены в отложениях Муссентучит и представлены преимущественно зубами. Самые крупные из известных зубов были отнесены к Pholidosauridae ; [ а ] они широкие, с округлым основанием, некоторые из них имеют продольные гребни. [ 8 ] Эти крупные крокодилы были способны охотиться по крайней мере на молодых эоламбий у кромки воды. Обилие молодых экземпляров Eolambia в ископаемом материале на берегу озера подтверждает эту гипотезу; более крупная Эоламбия была бы неуязвима для крокодилов. [ 7 ] Также присутствуют зубы крокодила меньшего размера с двумя различными морфотипами: яйцевидной формы с гребнями, которая была отнесена к двум разным видам Bernissartia ; [ 7 ] [ а ] и сжатая треугольная форма с тяжелым орнаментом, приписываемая Atoposauridae . [ 13 ] [ 41 ] [ а ] Однако, учитывая одинаковый небольшой размер этих зубов и одновременное появление этих морфотипов у некоторых крокодилов, [ 40 ] они могут принадлежать к одному и тому же виду. [ 8 ]
Большая Эоламбия была бы уязвима. [ 7 ] к большому неовенаториду тероподу- Сиацу , [ 42 ] который был обнаружен экспедициями ФМНХ. [ 8 ] В Муссентучите также присутствует множество более мелких теропод -манирапторов , которые действовали как падальщики; их останки включают зубы, приписываемые Dromaeosauridae , Paronychodon и Richardoestesia . [ 41 ] Что касается травоядных динозавров, то нодозаврид Animantarx был обнаружен в Участке Кэрол, рядом с типовым экземпляром Эоламбии . [ 43 ] Также были обнаружены зубы, принадлежащие более базальным орнитоподам, причем аналогичные зубы Чифелли в 1999 году отнес к Pachycephalosauria или зубастым птицам. [ 7 ] [ 41 ] Более полные останки, относящиеся к двум разным орнитоподам – ородромину и базальному игуанодонту – остаются неопубликованными. [ 44 ] гигантский овирапторозавр . Также был обнаружен [ 45 ] рядом с зубами цератопсов и зауроподов . [ 8 ] [ 41 ]
Рыбы, на которых, возможно, охотились более мелкие крокодилы, представлены зубами и чешуей. К ним относятся многолопастный скат, относящийся к роду Baibisha ; [ 41 ] неопределенный член Neopterygii ; семионотид , относящийся к роду Lepidotes ; гар , родственный Атрактостею ; [ 8 ] пикнодонтид , относящийся к роду Stephanodus ; и неопределенный боуфин . индетерминированные хвостатые Также присутствуют (саламандры) и млекопитающие. Что касается растений, образцы пыльцы указывают на многоярусный лес, окружающий озера Муссентучит, содержащий полог древовидных папоротников , араукариев и хвойных деревьев , сопровождаемый подлеском папоротников , мелких голосеменных и цветковых растений . В водах озер также присутствовали водоросли. [ 10 ] [ 46 ] Эти растения могли стать обильным источником пищи для Эоламбии . [ 7 ]
Палеобиогеография
[ редактировать ]Киркланд, Чифелли и коллеги отметили, что фауна муссентучита – игуанодонты, пахицефалозавры и цератопсы – имеет большое сходство с современной азиатской фауной. Они предположили, что Эоламбия была частью притока азиатских динозавров в Северную Америку во время сеномана, который вытеснил более раннюю местную фауну с низким разнообразием. [ 2 ] [ 13 ] Эта гипотеза подтверждается тесным родством между Eolambia и Probactrosaurus или Fukuisaurus , которые были соответственно обнаружены филогениями Хэда и Сьюса и Аверианова. [ 3 ] [ 4 ] [ 28 ]
Однако результаты Хорнера и его коллег, Прието-Маркеса и Макдональда и его коллег, которые считают североамериканского протохадроса ближайшим родственником Эоламбии , противоречат этой гипотезе. [ 8 ] [ 27 ] [ 29 ] [ 47 ] Эта альтернативная интерпретация предполагает, что общий предок Эоламбии , Протохадроса и других производных гадрозавроидов не был исключительно азиатским, а вместо этого был распространен по всей Азии и Северной Америке. [ 3 ] Макдональд и его коллеги далее предположили, что Eolambia и Protohadros представляют собой локализованную группу гадрозавроидов, эндемичных для Северной Америки, во многом как и другие группы стиракостернов на протяжении мелового периода. К числу таких современных групп относились центральноазиатские группы Shuangmiosaurus , Zhanghenglong и Plesiohadros . [ 8 ]
Примечания
[ редактировать ]- ^ Jump up to: а б с Учитывая, что эти классификации основаны исключительно на зубах, они ненадежны. [ 40 ] Макдональд и его коллеги решили признать эти морфотипы неопределенными неозухиями в 2017 году. [ 8 ]
Ссылки
[ редактировать ]- ^ Гальтон, Питер; Дженсен, Джеймс А. (1979). «Гальтон П.М. и Дженсен Дж.А. (1979). Остатки динозавров-орнитопод из нижнего мела Северной Америки» (PDF) . Геологические исследования в Университете Бригама Янга . 25 (3): 1–10. ISSN 0068-1016 . Архивировано из оригинала (PDF) 26 января 2021 года . Проверено 26 июня 2019 г.
- ^ Jump up to: а б с д и ж г час я дж к л м Киркланд, Дж.И. (1998). «Новый гадрозаврид из верхней формации Кедровой горы (альб-сеноман: мел) восточной части штата Юта – самый старый из известных гадрозавридов (ламбеозавр?)» . В Лукасе, СГ; Киркланд, Дж.И.; Эстеп, JW (ред.). Нижне- и среднемеловые наземные экосистемы . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 14. С. 283–295.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Макдональд, AT; Берд, Дж.; Киркланд, Дж.И.; Додсон, П. (2012). «Остеология базального гадрозавроида Eolambia caroljonesa (Dinosauria: Ornithopoda) из формации Кедр-Маунтин в штате Юта» . ПЛОС ОДИН . 7 (10): е45712. Бибкод : 2012PLoSO...745712M . дои : 10.1371/journal.pone.0045712 . ПМК 3471925 . ПМИД 23077495 .
- ^ Jump up to: а б с д и ж г Хед, Джей-Джей (2001). «Реанализ филогенетического положения Eolambia caroljonesa (Dinosauria, Iguanodontia)». Журнал палеонтологии позвоночных . 21 (2): 392–396. doi : 10.1671/0272-4634(2001)021[0392:AROTPP]2.0.CO;2 . S2CID 85705882 .
- ^ Киркланд, Дж.И.; Бердж, Д. (1994). Крупный примитивный гадрозавр из нижнего мела штата Юта . Пятьдесят четвертое ежегодное собрание Общества палеонтологии позвоночных. Журнал палеонтологии позвоночных . Том. 14, нет. 3. п. 32А. JSTOR i406457 .
- ^ Коста, ТВВ (2019). «Замечания к названию гадрозавроидного динозавра Eolambia caroljonesa Kirkland, 1998 (Ornithischia)» (PDF) . Бразильский журнал палеонтологии . 22 (2): 165–166. дои : 10.4072/rbp.2019.2.07 . S2CID 213724827 .
- ^ Jump up to: а б с д и ж г час я дж к л м Гаррисон-младший; Бринкман, Д.; Николс, диджей; Слой, П.; Бердж, Д.; Тейн, Д. (2007). «Многодисциплинарное исследование формации кедровой горы нижнего мела, Муссентучит-Уош, штат Юта: определение палеосреды и палеоэкологии карьера динозавров Eolambia caroljonesa ». Меловые исследования . 28 (3): 461–494. Бибкод : 2007CrRes..28..461G . дои : 10.1016/j.cretres.2006.07.007 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Макдональд, AT; Гейтс, штат Калифорния; Занно, Ле; Маковицкий, П.Дж. (2017). «Анатомия, тафономия и филогенетическое значение нового экземпляра Eolambia caroljonesa (Dinosauria: Ornithopoda) из формации Сидар-Маунтин, штат Юта, США» . ПЛОС ОДИН . 12 (5): e0176896. Бибкод : 2017PLoSO..1276896M . дои : 10.1371/journal.pone.0176896 . ПМК 5425030 . ПМИД 28489871 .
- ^ Крейг, LC (1981). «Породы нижнего мела, юго-запад Колорадо и юго-восток Юты» . Полевая конференция Ассоциации геологов Скалистых гор, 1981 г .: 195–200.
- ^ Jump up to: а б Чуди, Р.Х.; Чуди, Б.Д.; Крейг, LC (1984). Палинологическая оценка формаций Кедровой горы и каньона Берро, плато Колорадо (Отчет). Геологическая служба США, Профессиональная статья. Том. 1821. Геологическая служба США. стр. 1–24.
- ^ Николс, диджей; Сладкий, АР (1993). «Биостратиграфия неморских палинофлор верхнего мела на разрезе с севера на юг Западного внутреннего бассейна». В Колдуэлле, WGE; Кауфман, Э.Г. (ред.). Эволюция Западного внутреннего бассейна . Геологическая ассоциация Канады, Специальный доклад. Том. 39. Геологическая ассоциация Канады. стр. 539–584.
- ^ Янг, Р.Г. (1960). «Группа Дакота на плато Колорадо» . Бюллетень Американской ассоциации геологов-нефтяников . 44 (2): 156–194. doi : 10.1306/0bda5fd2-16bd-11d7-8645000102c1865d .
- ^ Jump up to: а б с Чифелли, РЛ; Киркланд, Дж.И.; Вейль, А.; Дейно, Алабама; Коваллис, Би Джей (1997). «Высокоточная геохронология 40Ar/39Ar и появление наземной фауны позднего мела в Северной Америке» . Труды Национальной академии наук . 94 (21): 11163–11167. Бибкод : 1997PNAS...9411163C . дои : 10.1073/pnas.94.21.11163 . ПМК 23402 . ПМИД 9326579 .
- ^ Пол, GS (2010). «Орнитишиане» . Принстонский полевой справочник по динозаврам . Принстон: Издательство Принстонского университета. п. 327. ИСБН 9780691167664 .
- ^ Jump up to: а б Хольц, ТР (2012). Хольца (2007) «Список родов динозавров » (PDF) . Дополнительная информация к динозаврам: самая полная и современная энциклопедия для любителей динозавров всех возрастов Томаса Р. Хольца-младшего, иллюстрации Луиса Рея . Департамент геологии Мэрилендского университета . Проверено 30 августа 2017 г.
- ^ Jump up to: а б с Вейшампель, Д.Б.; Норман, Д.Б.; Григореску, Д. (1993). « Telmatosaurus transsylvanicus из позднего мела Румынии: самый базальный гадрозавридный динозавр» . Палеонтология . 36 (2): 361–385.
- ^ Серено, ПК (1986). «Филогения птицетазовых динозавров (отряд Ornithischia)». Национальное географическое исследование . 2 : 234–256.
- ^ Jump up to: а б Бретт-Сурман, депутат Кнессета (1989). Пересмотр Hadrosauridae (Reptilia: Ornithischia) и их эволюция в кампане и маастрихте (PDF) (доктор философии). Вашингтон, округ Колумбия: Университет Джорджа Вашингтона. стр. 1–373.
- ^ Jump up to: а б с Вейшампель, Д.Б.; Хорнер, младший (1990). «Гадрозавриды». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. стр. 534–561.
- ^ Хорнер, младший; Карри, Пи Джей (1994). «Эмбриональная и неонатальная морфология и онтогенез нового вида Hypacrosaurus (Ornithischia, Lambeosauridae) из Монтаны и Альберты» . В Карпентере, К.; Хирш, К.Ф.; Хорнер, младший (ред.). Яйца динозавров и детеныши . Кембридж: Издательство Кембриджского университета. стр. 312–336. ISBN 9780521567237 .
- ^ Казановас, ML; Переда Субербиола, X.; Сантафе, СП; Вейшампель, Д.Б. (1999). «Первый ламбеозавровый гадрозаврид из Европы: палеобиогеографические последствия». Геологический журнал . 136 (2): 205–211. Бибкод : 1999GeoM..136..205C . дои : 10.1017/s0016756899002319 . S2CID 128603347 .
- ^ Лулл, РС; Райт, штат Невада (1942). «Гадрозавровые динозавры Северной Америки». Специальные статьи Геологического общества Америки . 40 : 1–242. дои : 10.1130/SPE40-p1 .
- ^ Норман, Д.Б. (1986). «К анатомии Iguanodon atherfieldensis (Ornithischia: Ornithopoda)» . Бюллетень Королевского института естественных наук Бельгии . 56 : 281–372.
- ^ Норман, Д.Б. (1998). «Об азиатских орнитоподах (Dinosauria: Ornithischia). 3. Новый вид динозавров-игуанодонтид» (PDF) . Зоологический журнал Линнеевского общества . 122 (1): 291–348. дои : 10.1111/j.1096-3642.1998.tb02533.x .
- ^ Норман, Д.Б. (2004). «Базальная игуанодонтия» . В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 413–437.
- ^ Хед, Джей-Джей (1998). «Новый вид базальных гадрозаврид (Dinosauria, Ornithischia) из сеномана Техаса». Журнал палеонтологии позвоночных . 18 (4): 718–738. Бибкод : 1998JVPal..18..718H . дои : 10.1080/02724634.1998.10011101 .
- ^ Jump up to: а б Хорнер, младший; Вейшампель, Д.Б.; Форстер, Калифорния (2004). «Базальная игуанодонтия» . В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 438–463.
- ^ Jump up to: а б Сьюс, Х.-Д.; Аверьянов, А. (2009). «Новый базальный гадрозавроидный динозавр из позднего мела Узбекистана и ранняя радиация утконосых динозавров» . Труды Королевского общества Б. 276 (1667): 2549–2555. дои : 10.1098/rspb.2009.0229 . ПМЦ 2686654 . ПМИД 19386651 .
- ^ Jump up to: а б Прието-Маркес, А.; Норелл, Массачусетс (2010). «Анатомия и взаимоотношения Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) из позднего мела Центральной Азии » Новитаты Американского музея (3694): 1–49. дои : 10.1206/3694.2 . S2CID 56372891 .
- ^ Чжэн, В.; Джин, X.; Сибата, М.; Азума, Ю. (2014). «Ранний молодой экземпляр Bolong yixianensis (Ornithopoda: Iguanodontia) из нижнего мела округа Нинчэн, Нэй-Монгол, Китай». Историческая биология . 26 (2): 236–251. Бибкод : 2014HBio...26..236Z . дои : 10.1080/08912963.2013.809347 . S2CID 129081459 .
- ^ Гаска, Дж. М.; Морено-Азанза, М.; Руис-Оменьака, Дж.И.; Канудо, Джи (2015). «Новый материал и филогенетическое положение базального игуанодонтного динозавра Delapparentia turolensis из баррема (раннего мела) Испании» . Журнал иберийской геологии . 41 (1): 57–70. doi : 10.5209/rev_jige.2015.v41.n1.48655 .
- ^ Верду, Ф.Дж.; Ройо-Торрес, Р.; Кобос, А.; Алькала, Л. (2015). «Перинаты нового вида игуанодона (Ornithischia: Ornithopoda) из нижнего баррема Гальве (Теруэль, Испания)». Меловые исследования . 56 : 250–264. Бибкод : 2015CrRes..56..250В . дои : 10.1016/j.cretres.2015.05.010 .
- ^ Верду, Ф.Дж.; Ройо-Торрес, Р.; Кобос, А.; Алькала, Л. (2017). «Новые систематические и филогенетические данные о раннем барремском Iguanodon galvensis (Ornithopoda: Iguanodontoidea) из Испании». Историческая биология . 30 (4): 437–474. Бибкод : 2018HBio...30..437V . дои : 10.1080/08912963.2017.1287179 . S2CID 89715643 .
- ^ Гейтс, штат Калифорния; Цогтбаатар, К.; Зано, Ле; Чинзориг, Т.; Ватабе, М. (2018). «Новый игуанодонт (Dinosauria: Ornithopoda) из раннего мела Монголии» . ПерДж . 6 : е5300. дои : 10.7717/peerj.5300 . ПМК 6078070 . ПМИД 30083450 .
- ^ Брочу, Калифорния (1994). «Закрытие нейроцентральных швов во время онтогенеза крокодилов: значение для оценки зрелости ископаемых архозавров». Журнал палеонтологии позвоночных . 16 (1): 49–62. Бибкод : 1996JVPal..16...49B . дои : 10.1080/02724634.1996.10011283 .
- ^ Хорнер, младший; де Риклес, А.; Падиан, К. (1999). «Вариации показателей скелетохронологии динозавров: значение для оценки возраста и физиологии». Палеобиология . 25 (3): 295–304. Бибкод : 1999Pbio...25..295H . дои : 10.1017/S0094837300021308 . S2CID 88428612 .
- ^ Вудворд, Х.Н.; Фридман Фаулер, Э.А.; Фарлоу, Джо; Хорнер, младший (2015). « Майазавр , модельный организм для биологии вымерших популяций позвоночных: большая выборочная статистическая оценка динамики роста и выживаемости». Палеобиология . 41 (4): 503–527. Бибкод : 2015Pbio...41..503W . дои : 10.1017/pab.2015.19 . S2CID 85902880 .
- ^ Старейшина, В.П.; Киркланд, Дж. И. «Меловая палеогеография южно-западного внутреннего региона» . В Капуто, МВ; Петерсон, Дж.А.; Францик, К.Дж. (ред.). Мезозойские системы региона Скалистых гор, США . Денвер: Раздел Скалистых гор SEPM. стр. 415–440.
- ^ Глэнси, Ти Джей младший; Артур, Массачусетс; Бэррон, Э.Дж.; Кауфман, Э.Г. (1993). «Палеоклиматическая модель североамериканского мелового (сеноман-туронского) эпиконтинентального моря». В Колдуэлле, WGE; Кауфман, Э.Г. (ред.). Эволюция Западного внутреннего бассейна . Геологическая ассоциация Канады, Специальные статьи. Том. 39. Геологическая ассоциация Канады. стр. 219–241.
- ^ Jump up to: а б Ирмис, РБ; Хатчисон, Дж. Х.; Сертич, JJW; Титус, Ал. (2013). «Крокодилформы из позднего мела национального памятника и окрестностей Гранд-Лестница-Эскаланте, Южная Юта, США» . В Титусе, Алабама; Лоуэн, Массачусетс (ред.). На вершине парадной лестницы: поздний мел Южной Юты . Блумингтон: Издательство Университета Индианы. стр. 424–444. ISBN 9780253008961 .
- ^ Jump up to: а б с д и Чифелли, РЛ; Нидам, РЛ; Гарднер, доктор медицинских наук; Вейль, А.; Итон, Дж. Г.; Киркланд, Дж.И.; Мэдсен, СК (1999). «Позвоночные животные среднего мелового периода из формации Сидар-Маунтин, округ Эмери, штат Юта: местная фауна Муссентучита» . В Gillette, Д. (ред.). Палеонтология позвоночных в Юте . Разные публикации Геологической службы Юты. Том. 99–1. стр. 219–242.
- ^ Занно, Ле; Маковицкий, П.Дж. (2013). «Тероподы-неовенаториды — высшие хищники позднего мела Северной Америки» . Природные коммуникации . 4 : 2827. Бибкод : 2013NatCo...4.2827Z . дои : 10.1038/ncomms3827 . ПМИД 24264527 .
- ^ Карпентер, К.; Киркланд, Дж.И.; Бердж, ДЛ; Берд, Дж. (1999). «Анкилозавры (Dinosauria: Ornithischia) формации Кедр-Маунтин, штат Юта, и их стратиграфическое распространение» . В Gillette, Д. (ред.). Палеонтология позвоночных в Юте . Разные публикации Геологической службы Юты. Том. 99–1. стр. 243–251.
- ^ Занно, Ле; Маковицкий, П.Дж. (2016). Новый вид раннедивергентных орнитопод увеличивает палеобиоразнообразие травоядных динозавров в позднемеловых экосистемах Северной Америки . 76-е ежегодное собрание Общества палеонтологии позвоночных. Солт-Лейк-Сити: Общество палеонтологии позвоночных. п. 256. Архивировано из оригинала 18 октября 2016 года . Проверено 17 сентября 2017 г.
- ^ Маковицкий, П.Дж.; Занно, Л.Е. (2014). Киркланд, Дж.И.; Хант-Фостер, Р.; Лиггетт, Джорджия; Трухильо, К. (ред.). Новые открытия увеличивают разнообразие фауны динозавров члена Муссентучит (формация Кедровой горы) . Средний мезозой: переходная эпоха динозавров (PDF) . Фрута: Геологическая служба Юты. стр. 70–71.
- ^ Равн, РЛ (1995). «Миоспоры из илистого песчаника (верхний альб), бассейн реки Уинд, Вайоминг, США» . Палеонтографика Абтейлунг Б. 234 (3): 41–91.
- ^ Прието-Маркес, А. (2010). «Глобальная историческая биогеография динозавров гадрозаврид» . Зоологический журнал Линнеевского общества . 159 (2): 503–525. дои : 10.1111/j.1096-3642.2010.00642.x .
