Дейнохейрус
| Дейнохейрус Временной диапазон: поздний мел , | |
|---|---|
 | |
| Реконструированный скелет в Японии. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | † Орнитомимозаврия |
| Семья: | † Дейнохейриды |
| Род: | † Дейнохейрус Осмольска и Роневич, 1970 г. |
| Разновидность: | † Д. замечательный |
| Биномиальное имя | |
| † Чудесный Дейнохейр Осмольска и Роневич, 1970 г. | |
Дейнохейрус ( / ˌ d aɪ n oʊ ˈ k aɪ r ə s / DY -no- KY -rəs ) — род крупных орнитомимозавров , живших в позднем меловом периоде около 70 миллионов лет назад. пара крупных рук, плечевые пояса и несколько других костей динозавра впервые были обнаружены В 1965 году в нэмегтской свите Монголии нового . В 1970 г. этот экземпляр стал голотипом единственного вида рода Deinocheirus mirificus ; Название рода по-гречески означает «ужасная рука». Никаких дальнейших останков не было обнаружено в течение почти пятидесяти лет, и его природа оставалась загадкой. В 2014 году были описаны еще два полных экземпляра, которые пролили свет на многие аспекты животного. Части этих новых экземпляров были украдены из Монголии несколько лет назад, но были репатриированы в 2014 году.
Дейнохейрус был необычным орнитомимозавром, самым крупным представителем клады , длиной 11 м (36 футов) и весом 6,5 т (7,2 коротких тонны). Хотя это было крупное животное, у него было много полых костей, что позволяло экономить вес. Руки были одними из самых больших среди всех двуногих динозавров, их длина составляла 2,4 м (7,9 футов), с большими тупыми когтями на трехпалых кистях. Ноги были относительно короткими и имели тупые когти. Его позвонки имели высокие нервные отростки, образующие « парус » вдоль спины. Большинство позвонков и некоторые другие кости были сильно пневматизированы из-за проникновения воздушных мешков . Хвост заканчивался пигостилевидными позвонками, что указывает на наличие веера из перьев. Череп имел длину 1,024 м (3,36 фута), с широким клювом и глубокой нижней челюстью, как у гадрозавров .
Классификация Deinocheirus долгое время была неопределенной, и первоначально его отнесли к теропод группе Carnosauria , но вскоре было отмечено сходство с орнитомимозаврами. После того, как были найдены более полные останки, было показано, что Deinocheirus является примитивным орнитомимозавром, наиболее тесно связанным с меньшими родами Garudimimus и Beishanlong , вместе образующими семейство Deinocheiridae . Представители этой группы не были приспособлены к скорости, в отличие от других орнитомимозавров. Считается, что дейнохейрус был всеяден ; форма его черепа указывает на растительную диету, у одного экземпляра была обнаружена рыбья чешуя, а гастролиты в области желудка экземпляра также присутствовали . Большие когти, возможно, использовались для копания и сбора растений. Следы укусов на костях дейнохейруса приписывают тираннозавриду тарбозавру .
Открытие
[ редактировать ]
Первые известные ископаемые останки дейнохейруса были обнаружены польским палеонтологом Зофией Килан-Яворовской 9 июля 1965 года на стоянке Алтан Ула III ( координаты : 43 ° 33,987' с.ш. , 100 ° 28,959' в.д. / 43,566450 ° с.ш., 100,482650 ° в.д. ) в котловине Немегта пустыни Гоби . Она входила в состав польской группы в сопровождении монгольского палеонтолога Ринчена Барсболда во время польско-монгольских палеонтологических экспедиций 1963–1965 годов, которые были организованы Польской академией наук и Монгольской академией наук . Экипаж провел 9–11 июля, выкапывая образец и загружая его на автомобиль. В отчете Килан-Яворовской и Найдин Довчин за 1968 год, в котором суммировались достижения экспедиций, было объявлено, что останки представляют собой новое семейство динозавров -теропод . [1] [2]

Экземпляр был обнаружен на небольшом холме в песчанике и представляет собой частичный разрозненный скелет, большая часть которого, вероятно, разрушилась на момент открытия. Экземпляр состоял из обеих передних конечностей, за исключением когтей правой руки, всего плечевого пояса, центральных частей трех спинных позвонков, пяти ребер, гастралий (брюшных ребер) и двух цератобранхиалей. Образец стал голотипом Deinocheirus mirificus , названного Хальшкой Осмолской и Евой Роневич в 1970 году. Родовое название происходит от греческого deinos (δεινός), что означает «ужасный», и cheir (χείρ), что означает «рука», из-за размер и сильные когти передних конечностей. Видовое название происходит от латинского языка и означает «необычный» или «своеобразный», выбранное из-за необычного строения передних конечностей. [3] Польско-монгольские экспедиции примечательны тем, что их возглавляли женщины, которые были одними из первых, кто назвал новые динозавры. [4] Первоначальный номер экземпляра голотипа был ZPal MgD-I/6, но с тех пор он был перекаталогизирован как MPC-D 100/18. [1]

Нехватка известных останков дейнохейруса препятствовала полному пониманию этого животного на протяжении почти полувека, и в научной литературе его часто описывали как одного из самых «загадочных», «таинственных» и «причудливых» динозавров. [1] [5] [6] Руки-голотипы стали частью передвижной выставки окаменелостей монгольских динозавров, путешествующей по разным странам. [7] В 2012 году Фил Р. Белл, Филип Дж. Карри и Юонг-Нам Ли объявили об открытии дополнительных элементов образца голотипа, включая фрагменты гастралии, найденные корейско-монгольской командой, которая переместила первоначальный карьер в 2008 году. Следы укусов на двух гастралиях были идентифицированы как принадлежащие тарбозавру , и было высказано предположение, что это объясняет рассеянное, разобщенное состояние экземпляра голотипа. [6]
Дополнительные образцы
[ редактировать ]об открытии двух новых экземпляров дейнохейруса (SVP) объявили Общества палеонтологии позвоночных В 2013 году Ли, Барсболд, Карри и их коллеги перед ежегодной конференцией . Этим двум обезглавленным особям, размещенным в Академии наук Монголии, были присвоены номера экземпляров MPC-D 100/127 и MPC-D 100/128. MPC-D 100/128, полувзрослая особь, обнаружена учеными в местонахождении Алтан Ула IV (координаты: 43 ° 36,091' с.ш. 100 ° 27,066' в.д. / 43,601517 ° с.ш. 100,451100 ° в.д. ) формации Нэмегт во время Корейско-Монгольской международной экспедиции по динозаврам в 2006 году, но уже был поврежден ископаемыми браконьерами . Второй экземпляр, MPC-D 100/127, был найден учеными в местонахождении Бугийн Цав (координаты: 43 ° 54,025' с.ш. 99 ° 58,359' в.д. / 43,900417 ° с.ш. 99,972650 ° в.д. ) в 2009 году. Он немного крупнее голотипа, и его можно было четко идентифицировать как Deinocheirus по левой передней конечности, что помогло идентифицировать ранее собранный экземпляр как Deinocheirus . Образец также был выкопан браконьерами, которые удалили череп, руки и ноги, но оставили единственную кость пальца ноги. Судя по деньгам, оставшимся в карьере, его, вероятно, разграбили после 2002 года. [1] [8] Черепа, кости когтей и зубы часто становятся выборочной целью браконьеров в ущерб остальным скелетам (которые часто подвергаются вандализму) из-за их пригодности для продажи. [9] Карри заявил в интервью, что политика их команды заключалась в исследовании карьеров после того, как они были разграблены, и восстановлении чего-либо значимого, и что обнаружение любых новых окаменелостей дейнохейруса было поводом для празднования, даже без браконьерских частей. Виртуальная модель дейнохейруса , представленная на презентации СВП, вызвала аплодисменты толпы присутствовавших палеонтологов, а американский палеонтолог Стивен Л. Брусатт заявил, что он никогда не был так удивлен выступлением СВП, хотя на конференции регулярно представляются новые окаменелости. [10]
После того, как было объявлено о новых образцах, поползли слухи, что награбленный череп попал в европейский музей через черный рынок . [10] Добытые браконьерами элементы были обнаружены в частной европейской коллекции французским торговцем окаменелостями Франсуа Эскюйе, который уведомил о них бельгийского палеонтолога Паскаля Годфруа в 2011 году. Они заподозрили, что останки принадлежали дейнохейрусу , и связались с корейско-монгольской командой. Впоследствии Эскуйе приобрел окаменелости и передал их Королевскому бельгийскому институту естественных наук . [11] Найденный материал состоял из черепа, левой руки и ног, которые были собраны в Монголии, проданы японскому покупателю и перепроданы немецкой стороне (окаменелости также прошли через Китай и Францию). Команда пришла к выводу, что эти элементы принадлежали образцу MPC-D 100/127, поскольку единственная оставшаяся кость пальца ноги идеально вписывалась в неподготовленный матрикс отварной стопы, кость и матрикс совпадали по цвету, а также поскольку элементы принадлежали человеку из одинакового размера, без перекрытия элементов скелета. [1] [12] 1 мая 2014 года окаменелости были репатриированы в Монголию делегацией Бельгийского музея во время церемонии, состоявшейся в Монгольской академии наук. [13] Воссоединенный скелет был передан на хранение в Центральный музей монгольских динозавров в Улан-Баторе вместе со скелетом тарбозавра , который также был возвращен после кражи . Американский палеонтолог Томас Р. Хольц заявил в интервью, что новые останки дейнохейруса выглядели как «продукт тайной любви между гадрозавром и галлимимом ». [11]

В сочетании с браконьерскими элементами оба новых экземпляра представляют почти весь скелет дейнохейруса , поскольку MPC-D 100/127 включает весь материал, за исключением средних спинных позвонков, большинства хвостовых позвонков и правой передней конечности; MPC-D 100/128 заполняет большинство промежутков другого скелета, почти все спинные и хвостовые позвонки, подвздошную кость, частичную седалищную кость и большую часть левой задней конечности. В 2014 году экземпляры описали Ли, Барсболд, Карри, Ёсицугу Кобаяши, Ханг-Дже Ли Ли, Годфруа, Эскуилье и Цогтбаатар Чинзориг. [1] Похожая серия событий была зарегистрирована ранее в 2014 году со , еще одним парусным тероподом, который был известен лишь по небольшому количеству останков с 1912 года спинозавром . спинозавриды . Оба случая показали, что об образе жизни и внешнем виде малоизвестных вымерших животных не всегда можно с уверенностью судить по близким родственникам. [14] К 2017 году правительство Монголии активизировало усилия по изъятию у коллекционеров украденных браконьерами окаменелостей и их репатриации, но доказательство их происхождения стало научной и политической проблемой. Поэтому в исследовании была проверена возможность идентификации добытых браконьерами окаменелостей геохимическими методами на примере дейнохейруса и других динозавров немегта. [9] В 2018 году в местонахождении Немегт были зарегистрированы многочисленные крупные трехпалые (трехпалые) следы (обнаруженные в 2007 году рядом со следами зауроподов ). Хотя следы были похожи на следы гадрозавров, следов рук гадрозавров обнаружено не было, а поскольку теперь известно, что ступни дейнохейруса были похожи на ступни гадрозавров, нельзя исключать, что следы были оставлены представителями этого рода. [15]
Описание
[ редактировать ]
Дейнохейрус - крупнейший из обнаруженных орнитомимозавров (страусовых динозавров); Согласно описанию 2014 года, самый крупный из известных экземпляров имел длину около 11 м (36 футов) и расчетный вес 6,36 т (7,01 коротких тонны). [1] Два других известных экземпляра меньше по размеру: размер голотипа составляет 94%, а размер самого маленького, полувзрослого экземпляра, составляет всего 74%. [1] В 2016 году Грегори С. Пол представил более высокую оценку длины - 11,5 м (38 футов), но меньшую оценку массы - 5 т (5,5 коротких тонн). [16] Также в 2016 году Асьер Ларраменди и Молина-Перес представили более высокую оценку длины - 12 м (39 футов) и более высокую оценку массы - 7 т (7,7 коротких тонн), а также расчетную высоту бедра - 4,4 м (14 футов). [17] В 2020 году Кампионе и Эванс оценили массу тела примерно в 6,5 т (7,2 коротких тонны). [18] Когда были известны только неполные рукава голотипов, на их основе разными методами экстраполировались различные размеры. По данным исследования 2010 года, высота бедер дейнохейруса составляет 3,3–3,6 м (11–12 футов). [19] Ранее вес оценивался от 2 тонн (2,2 коротких тонны) до 12 тонн (13 коротких тонн). Огромные размеры были также предположены путем сравнения рук с руками тираннозавров , хотя у членов этой группы не было больших рук, пропорциональных размеру их тела. [20] [21] [22]

Единственный известный череп, принадлежащий самому крупному экземпляру, имеет размеры 1,024 м (3,36 фута) от предчелюстной кости спереди до задней части затылочного мыщелка . Для сравнения, самая широкая часть черепа за глазами составляет всего 23 см (9,1 дюйма). Череп был похож на череп других орнитомимозавров тем, что был низким и узким, но отличался более удлиненной мордой. Стенки костей черепа были довольно тонкими, около 6 мм (0,24 дюйма). был бы покрыт кератином У него был округлый, приплюснутый клюв, который при жизни . Ноздри были обращены вверх, а носовая кость представляла собой узкую полоску, выступавшую вверх над глазницами. Внешний диаметр склеротических колец в глазах был небольшим - 8,4 см (3,3 дюйма) по сравнению с размером черепа. Нижние височные окна , отверстия за глазами, были частично закрыты скуловыми костями , подобно Галлимиму . Челюсти были беззубыми и повернуты вниз, а нижняя челюсть была очень массивной и глубокой по сравнению со тонкой и низкой верхней челюстью. Относительный размер нижней челюсти был ближе к тираннозавридам, чем к другим орнитомимозаврам. Морда была лопатообразной (расширенной в стороны) и шириной 25 см (9,8 дюйма), что шире, чем крыша черепа. [1] Эта форма была похожа на морду утконосых гадрозаврид. [14]
Посткраниальный скелет
[ редактировать ]
Дейнохейрус и Теризинозавр обладали самыми длинными передними конечностями среди всех двуногих динозавров. [19] Длина передних конечностей голотипа составляет 2,4 м (7,9 футов), плечевая кость (верхняя кость руки) — 93,8 см (36,9 дюйма), локтевая кость — 68,8 см (27,1 дюйма), а кисть — 77 см (30 дюймов), включая 19,6 дюйма. Загнутые когти длиной сантиметра (7,7 дюйма). Каждая лопатка плечевого пояса имеет длину 1,53 м (5 футов). Размер каждой половины парных цератобранхиалий составляет 42 см (17 дюймов). Лопатка длинная и узкая, гребень дельтопекторальной мышцы выраженный, треугольной формы. Плечевая кость была относительно тонкой и лишь немного длиннее кисти. Локтевая и лучевая кости (кости предплечья) были удлиненными и не были прочно соединены друг с другом в синдесмозе . Пясть . была длиннее пальцев Три пальца были примерно одинаковой длины: первый был самым толстым, а второй самым длинным. Различные шероховатые участки и вдавления на передних конечностях указывают на наличие мощной мускулатуры. Большинство суставных поверхностей костей рук были глубоко бороздчатыми, что указывает на наличие у животного толстых хрящевых прокладок между суставами. Хотя оружие Дейнохейрусы были крупными, соотношение между ними и плечевым поясом было меньше, чем у более мелкого орнитомимозавра Ornithomimus . [3] Кости рук Deinocheirus по пропорциям были аналогичны костям мелкого теропода Compsognathus . [23]
Хотя дейнохейрус был громоздким животным, его спинные ребра были высокими и относительно прямыми, что указывало на узкое тело. [8] Десять шейных позвонков были низкими и длинными и постепенно укорачивались назад от черепа. В результате шея стала более S-образной, чем у других орнитомимозавров, из-за большего черепа. Нервные отростки двенадцати задних позвонков становились все длиннее спереди назад, последний из них в 8,5 раз превышал высоту центральной части . Это почти то же самое, что и самое высокое соотношение в нервных отростках теропода Spinosaurus . Нервные отростки имели систему соединяющихся связок , которые укрепляли позвоночный столб, позволяя ему поддерживать живот, передавая нагрузку на бедра и задние конечности. [1] Вместе нервные отростки образовывали высокий « парус » вдоль поясницы, бедер и основания хвоста, чем-то похожий на таковой у спинозавра . [14]
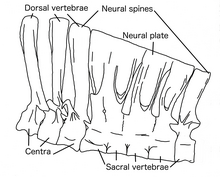
Все позвонки были сильно пневматизированы вторгшимися воздушными мешками , за исключением атланта и самых задних хвостовых позвонков, и, таким образом, были связаны с дыхательной системой . Задние позвонки были столь же пневматизированы, как и у динозавров- зауроподов , и имели обширную систему впадин. Эти адаптации могут быть связаны с гигантизмом , поскольку они уменьшают вес. Шесть позвонков крестца также были высокими и пневматизированными, и все, кроме первого, были слиты вместе наверху, а их нервные отростки образовывали нервную пластинку . , Подвздошная кость верхняя часть бедренной кости, также была частично пневматизирована вблизи крестцовых позвонков. Часть таза была гипертрофирована (увеличена) по сравнению с другими орнитомимозаврами, чтобы поддерживать вес животного с помощью сильных мышечных прикреплений. Передние бедренные кости при жизни наклонены вверх. Хвост дейнохейруса заканчивался как минимум двумя сросшимися позвонками, которые были описаны как похожие на пигостиль овирапторозавров теропод и теризинозавроидов . Известно, что орнитомимозавры имели перистые перья , поэтому эта особенность предполагает, что у них мог быть веер из перьев на конце хвоста. [1] [24]
Рычаг ( фуркула ), элемент, не известный ни у каких других орнитомимозавров, имел U-образную форму. Задние конечности были относительно короткими, а бедренная кость (бедренная кость) была длиннее голени (голень), как это обычно бывает у крупных животных. Плюсна аркто - была короткой и не метатарзальной , как у большинства других теропод. Когтевые кости стоп были тупыми и с широкими кончиками, а не заостренными, в отличие от других теропод, но напоминали когти крупных птицетазовых динозавров. Пропорции костей пальцев ног напоминали кости тираннозавров из-за большого веса, который им приходилось выдерживать. [1]
Классификация
[ редактировать ]Когда дейнохейрус был известен только по исходным передним конечностям, его таксономическое родство было трудно определить, и было предложено несколько гипотез. [14] Осмольска и Роневич первоначально пришли к выводу, что дейнохейрус не принадлежал ни к одному уже названному семейству теропод, поэтому они создали новое, монотипное семейство Deinocheiridae , помещенное в инфраотряд Carnosauria . Это объяснялось крупными размерами и толстостенными костями конечностей, но у них также обнаружены некоторые сходства с орнитомимами , и, в меньшей степени, с аллозаврами . [3] В 1971 году Джон Остром впервые предположил, что дейнохейрус принадлежал к отряду орнитомимозавров, отметив при этом, что он содержит как орнитомимозавровые, так и неорнитомимозавровые признаки . [1] [14] В 1976 году Ринчен Барсболд назвал отряд Deinocheirosauria , который должен был включать предположительно родственные роды Deinocheirus и Therizinosaurus . Связь между дейнохейрусом и длиннорукими теризинозаврами поддерживалась некоторыми более поздними авторами, но сегодня они не считаются близкими родственниками. [5]
В 2004 году Питер Маковицки, Кобаяши и Карри отметили, что дейнохейрус , вероятно, был примитивным орнитомимозавром, поскольку у него отсутствовали некоторые черты, типичные для семейства Ornithomimidae . [25] Примитивные черты включают загнутые когти, низкое соотношение плечевой кости к лопатке и отсутствие синдесмоза. Исследование, проведенное Кобаяши и Барсболдом в 2006 году, показало, что дейнохейрус , возможно, является самым примитивным орнитомимозавром, но не смогло определить его сходство из-за отсутствия элементов черепа и задних конечностей. [5] Кладистический анализ , сопровождавший описание двух гораздо более полных экземпляров в 2014 году, показал, что Deinocheirus образовал кладу с Garudimimus и Beishanlong , которые поэтому были включены в Deinocheiridae. Итоговая кладограмма представлена ниже: [1]

| Орнитомимозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Исследование 2014 года определило Deinocheiridae как кладу, включающую все таксоны с более поздним общим предком с Deinocheirus mirificus, чем с Ornithomimus velox . У всех троих общие анатомические особенности конечностей. Кладограмма 2014 года показала, что орнитомимозавры в раннем мелу разделились на две основные линии: Deinocheiridae и Ornithomimidae. В отличие от других орнитомимозавров, дейнохейриды не были созданы для бега. Анатомические особенности дейнохейруса по сравнению с другими, гораздо меньшими орнитомимозаврами, во многом можно объяснить его гораздо большими размерами и весом. [1] Дейнохейриды и более мелкие орнитомимиды не имели зубов, в отличие от более примитивных орнитомимозавров. [14] В 2020 году был назван дейнохейрид Paraxenisaurus из Мексики, что сделало его первым представителем группы, известным из Северной Америки. Его описатели предположили, что дейнохейриды произошли из Лавразии (северный суперконтинент того времени) или что они расселились по полярным регионам в Северном полушарии , и известно, что аналогичный обмен произошел и с другими группами динозавров с азиатским родством в кампан - маастрихтский период. . Это исследование также показало, что Harpymimus является базальным дейнохейридом, тогда как Бэйшаньлун находится вне группы, как базальный орнитомимозавр. [26]
Палеобиология
[ редактировать ]
Тупые и короткие когти дейнохейруса были похожи на когти теризинозавра Alxasaurus , что указывает на то, что длинные руки и когти использовались для рытья и сбора растений. Тупые когти ног могли помочь животному не проваливаться в грунт при переходе вброд. Крепкие задние конечности и область бедер указывают на то, что животное двигалось медленно. Большой размер животного, возможно, защитил его от хищников, таких как тарбозавр , но, в свою очередь, он потерял способность бегать, как другие орнитомимозавры. Длинные нервные отростки и, возможно, веер хвоста, возможно, использовались для демонстрации поведения . Дейнохейрус, вероятно, вел дневной образ жизни (активен в течение дня), поскольку склеротические кольца глаз были относительно небольшими по сравнению с длиной его черепа. [1] Кисть имела хорошую подвижность относительно предплечья, но была способна лишь на ограниченное сгибательное движение и не могла смыкаться при захвате. [3]
Мозг дейнохейруса был реконструирован с помощью компьютерной томографии и представлен на конференции Общества палеонтологии позвоночных в 2014 году. Мозг был шаровидным и по форме напоминал мозг птиц и троодонтидных теропод, головной мозг был расширен, как и у большинства теропод, а обонятельные тракты были относительно большими. Мозг был пропорционально маленьким и компактным, а его коэффициент энцефализации рептилий (отношение мозга к телу) оценивался в 0,69, что является низким показателем для теропод и аналогичен таковому у зауроподов. У других орнитомимозавров мозг пропорционально большой, а маленький мозг дейнохейруса может отражать его социальное поведение или диету. Его координация и баланс не были бы так важны, как для хищных теропод. [27]

что вместе с археорнитомимом и галлимимусом дейнохейрус В 2015 году Акинобу Ватанабэ и его коллеги обнаружили , имел наиболее пневматизированный скелет среди орнитомимозавров. Считается, что пневматизация полезна для полета современных птиц, но ее функция у нептичьих динозавров достоверно не известна. Было высказано предположение, что пневматизация использовалась для уменьшения массы крупных костей (связанная с гигантскими размерами в случае Deinocheirus ), что она была связана с высоким обменом веществ , балансом во время передвижения или использовалась для терморегуляции . [24]
костей Исследование микроструктуры , представленное Европейской ассоциацией палеонтологов позвоночных в 2015 году, показало, что дейнохейрус , вероятно, имел высокую скорость метаболизма и быстро рос до достижения половой зрелости . [28] Гистологическое сухожилий исследование фрагмента гастралии голотипа, представленного на конференции 2018 года, показало, что его внутреннее строение сходно со структурой окостеневших других теропод. Остеоны содержали возможные канальцы , что стало первым известным появлением таких структур у базального орнитомимозавра. Структура надкостницы и отсутствие линий остановки роста позволяют предположить, что голотип был взрослой особью. [29]
Диета
[ редактировать ]Отчетливая форма черепа показывает, что у дейнохейруса была более специализированная диета, чем у других орнитомимозавров. Клюв был похож на утиный , что указывает на то, что они, возможно, также добывали пищу в воде или паслись у земли, как некоторые зауроподы и гадрозавры. Места прикрепления мышц, открывающих и закрывающих челюсти, были очень маленькими по сравнению с размером черепа, что указывает дейнохейруса на слабую силу укуса . Череп, вероятно, был приспособлен для обрезки мягкого подлеска или водной растительности. Глубина нижней челюсти указывает на наличие крупного языка, который мог помогать животному всасывать пищевой материал, добытый широким клювом при кормлении на дне пресноводных водоемов. [1]

более 1400 гастролитов Среди ребер и гастралий особи MPC-D100/127 обнаружено (желудочные камни размером от 8 до 87 мм). Отношение массы гастролитов к общему весу, равное 0,0022, подтверждает теорию о том, что эти гастролиты помогали беззубым животным перемалывать пищу. Такие особенности, как наличие клюва и U-образной, загнутой вниз челюсти, являются индикаторами факультативного (факультативного) травоядности среди целурозавровых теропод. Несмотря на эти особенности, среди гастролитов были обнаружены также позвонки и чешуя рыбы, что позволяет предположить, что она была всеядной . [1] Считается, что орнитомимозавры питались как растениями, так и мелкими животными. [14]
Дэвид Дж. Баттон и Занно обнаружили в 2019 году, что травоядные динозавры в основном придерживались двух различных способов питания: либо переработка пищи в кишечнике, характеризующаяся изящными черепами и низкой силой укуса, либо рот, характеризующийся особенностями, связанными с интенсивной переработкой. Было обнаружено, что дейнохейрус , наряду с орнитомимидными орнитомимозаврами, диплодокоидными и титанозавровыми зауроподами, сегнозаврами и ценугнатидами , относится к первой категории. Эти исследователи предположили, что дейнохейриды и орнитомимиды-орнитомимозавры, такие как Gallimimus, вторглись в эти ниши по отдельности, конвергентно достигнув относительно больших размеров. Преимущества большой массы тела у травоядных включают повышенную скорость потребления пищи и устойчивость к голоданию, и, следовательно, эти тенденции могут указывать на то, что дейнохейриды и орнитомимиды были более травоядными, чем другие орнитомимозавры. Они предупредили, что корреляция между массой тела и массой тела непростая и что у этого клада не наблюдается направленной тенденции к увеличению массы. Кроме того, диета большинства орнитомимозавров плохо изучена. Дейнохейрус , по-видимому, был, по крайней мере, условно всеяден. [30]
В статье Вайсума Ма и его коллег, опубликованной в 2022 году, с помощью анализа методом конечных элементов было исследовано, как механика кормления различается между различными группами целурозавров, не относящихся к птицам , и было обнаружено, что у всех них произошло снижение стресса в челюстях, связанного с кормлением. Они обнаружили, что дейнохейрус демонстрировал другие модели распределения стресса и напряжения, чем другие орнитомимизавры, что указывает на то, что он был специализированным питателем. Они подозревали, что дейнохейрус , возможно, вернулся к всеядности/хищности. [31]
Различные варианты пищевого поведения были предложены до того, как были известны более полные останки дейнохейруса , и на раннем этапе его представляли как хищное, похожее на аллозавра животное с гигантскими руками. [14] В своем первоначальном описании Осмольска и Роневич обнаружили, что руки дейнохейруса не приспособлены для хватания, а вместо этого могут быть использованы для разрывания добычи. [3] В 1970 году российский палеонтолог Анатолий Константинович Рождественский сравнил передние конечности дейнохейруса с ленивцами , что привело его к гипотезе, что дейнохейрус был специализированным лазающим динозавром, который питался растениями и животными, обитающими на деревьях. [32] Вместо этого в 1988 году Пол предположил, что когти слишком тупы для хищнических целей, но могут быть хорошим защитным оружием. [22] Пытаясь определить экологические ниши для дейнохейруса и теризинозавра в 2010 году, Фил Сентер и Джеймс Х. Робинс предположили, что дейнохейрус имел самый большой диапазон вертикального питания из-за высоты бедра и специализировался на поедании высокой листвы. [19] В 2017 году было высказано предположение, что когти дейнохейруса приспособлены для вытягивания большого количества травянистых растений из воды и для уменьшения сопротивления воды. [33]
Палеопатология
[ редактировать ]
Осмольска и Роневич сообщили о палеопатологиях в образце голотипа, таких как аномальные ямки, бороздки и бугорки на первой и второй фалангах левого второго пальца, которые могли быть результатом травм сустава между двумя костями. Повреждение могло вызвать изменения в расположении связок мышц. Два коракоида также развиты по-разному. [3] [34] На ребре образца MPC-D 100/127 видна зажившая травма, в результате которой кость ремоделировалась. [1] В 2012 г. сообщалось о следах укусов на двух гастралиях голотипа. Размер и форма следов укусов соответствуют зубам Тарбозавра , крупнейшего известного хищника из формации Немегт. Выявлены различные типы следов питания; проколы, сколы, борозды, фрагментированные зубы и комбинации вышеперечисленных признаков. Следы укусов, вероятно, отражают пищевое поведение, а не агрессию между видами, а тот факт, что следы укусов не были обнаружены на других участках тела, указывает на то, что хищник сосредоточился на внутренних органах. Следы укусов тарбозавра также были обнаружены на окаменелостях гадрозавров и зауроподов, но следы укусов теропод на костях других теропод очень редки в летописи окаменелостей. [6]
Палеосреда
[ редактировать ]
Три известных экземпляра Deinocheirus были обнаружены в формации Нэмегт в пустыне Гоби на юге Монголии. Это геологическое образование никогда не было датировано радиометрически , но фауна, присутствующая в летописи окаменелостей, указывает на то, что оно, вероятно, отложилось в ранний маастрихтский период, в конце позднего мела, около 70 миллионов лет назад. [35] [36] [37] пород Фации немегтинской свиты предполагают наличие русел ручьев и рек, илистых отмелей и мелких озер. Такие крупные речные русла и почвенные отложения свидетельствуют о гораздо более влажном климате, чем тот, который встречается в более древних формациях Барун-Гойот и Джадохта . Однако отложения калише указывают на то, что случались, по крайней мере, периодические засухи. Осадки отлагались в руслах и поймах крупных рек. [38]
Считается, что дейнохейрус был широко распространен в формации Немегт, поскольку единственные три найденных экземпляра находились на расстоянии 50 км (31 миль) друг от друга. Речные системы немегтинской свиты предоставили подходящую нишу для дейнохейруса с его всеядностью. [1] Окружающая среда была похожа на дельту Окаванго в современной Ботсване . В этой экосистеме дейнохейрус питался растениями и мелкими животными, включая рыбу. Возможно, он конкурировал за деревья с другими крупными травоядными динозаврами, такими как длинношеий теропод Теризинозавр , различные зауроподы -титанозавры и более мелкие гадрозавриды Saurolophus . Дейнохейрус, возможно, конкурировал с этими травоядными животными за более высокую листву, например деревья, но также мог питаться материалом, которого они не могли. Наряду с дейнохейрусами открытия теризинозавров и гиганторапторов показывают, что три группы травоядных теропод (орнитомимозавры, теризинозавры и овирапторозавры) независимо достигли максимальных размеров в позднем мелу Азии. [14]
Среда обитания в реках Немегт и вокруг них, где обитал дейнохейрус , служила домом для широкого спектра организмов. Иногда встречаются окаменелости моллюсков , а также множество других водных животных, таких как рыбы и черепахи, а также крокодиломорф- параллигатор . [35] [39] [40] Окаменелости млекопитающих редки в формации Немегт, но было обнаружено множество птиц, в том числе энантиорнитин Gurilynia , гесперорнитиформный Judinornis , а также Teviornis , возможный гусеобразный вид . Травоядные динозавры формации Немегт включают анкилозаврид , таких как Tarchia , пахицефалозавров Prenocephale , крупных гадрозавридов, таких как Saurolophus и Barsboldia , и зауроподов, таких как Nemegtosaurus и Opisthocoelicaudia . [35] [41] Хищные тероподы, жившие рядом с дейнохейрусами, включают тираннозавров, таких как Тарбозавр , Алиорам и Багараатан , и троодонтид, таких как Бороговия , Тохизавр и Занабазар . Группы теропод, включающие как всеядных, так и травоядных представителей, включают теризинозавров, таких как теризинозавр , овирапторозавров, таких как Elmisaurus , Nemegtomaia и Rinchenia , и других орнитомимозавров, таких как Anserimimus и Gallimimus . [42]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Ли, Ю.Н.; Барсболд, Р .; Карри, Пи Джей ; Кобаяши, Ю.; Ли, HJ; Годфруа, П .; Скуилли, ФО; Чинзориг, Т. (2014). Разгадка давней загадки гигантского орнитомимозавра Deinocheirus mirificus « Природа 515 (7526): 257–260. Бибкод : 2014Nature.515..257L . дои : 10.1038/nature13874 . ПМИД 25337880 . S2CID 2986017 .
- ^ Килан-Яворовска, З. ; Довчин, Н. (1968). «Рассказ о польско-монгольских палеонтологических экспедициях 1963–1965 годов» (PDF) . Палеонтологическая Полоника . 19:24 .
- ^ Jump up to: а б с д и ж Осмольска, Х. ; Роневич, Э. (1970). «Дейнохейриды, новое семейство тероподовых динозавров» (PDF) . Палеонтология Полоника (21): 5–19. Архивировано из оригинала (PDF) 3 марта 2016 года . Проверено 10 ноября 2009 г.
- ^ Додсон, П. (1998). Рогатые динозавры: естественная история . Издательство Принстонского университета. п. 9. ISBN 978-0-691-05900-6 .
- ^ Jump up to: а б с Кобаяши, Ю.; Барсболд, Р. (2006). «Орнитомимиды из немегтинской свиты Монголии» (PDF) . Журнал Палеонтологического общества Кореи . 22 (1): 195–207.
- ^ Jump up to: а б с Белл, PR; Карри, Пи Джей ; Ли, Ю.Н. (2012). «Следы питания тираннозавров на дейнохейрусах (Theropoda:?Ornithomimosauria) остаются из формации Немегт (поздний мел), Монголия» . Меловые исследования . 37 : 186–190. Бибкод : 2012CrRes..37..186B . дои : 10.1016/j.cretres.2012.03.018 .
- ^ «Меловые монгольские динозавры» . DinoCasts.com. Архивировано из оригинала 2 февраля 2015 года.
- ^ Jump up to: а б Ли, Ю.Н.; Барсболд, Р. ; Карри, Пи Джей ; Кобаяши, Ю.; Ли, HJ (2013). «Новые экземпляры Deinocheirus mirificus из позднего мела Монголии» (PDF) . Рефераты статей Общества палеонтологии позвоночных : 161. Архивировано из оригинала (PDF) 4 декабря 2014 г.
- ^ Jump up to: а б Фанти, Ф.; Белл, PR; Тайге, М.; Милан, Луизиана; Динелли, Э. (2017). «Геохимическая дактилоскопия как инструмент репатриации окаменелостей динозавров, добытых браконьерами в Монголии: тематическое исследование для местности Нэмэгт, пустыня Гоби». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 51–64. Бибкод : 2018PPP...494...51F . дои : 10.1016/j.palaeo.2017.10.032 .
- ^ Jump up to: а б Свитек, Б. (4 ноября 2013 г.). «Таинственный динозавр наконец получил тело» . Национальное географическое общество . Архивировано из оригинала 7 ноября 2013 года.
- ^ Jump up to: а б Хехт, Дж. (12 мая 2014 г.). «Украденная голова динозавра открывает странные гибридные виды» . Новый учёный .
- ^ Пикрелл, Дж. (2020). «Забавный случай с похищенным динозавром-лебедем». Новый учёный . 245 (3269): 40–43. Бибкод : 2020НовыйSc.245...40P . дои : 10.1016/S0262-4079(20)30314-6 . S2CID 212997151 .
- ^ «Окаменелости динозавра дейнохейруса «ужасной руки» репатриированы на родину» . InfoMongolia.com. Архивировано из оригинала 12 мая 2014 года.
- ^ Jump up to: а б с д и ж г час я Хольц, ТР (2014). «Палеонтология: Тайна ужасных рук раскрыта». Природа . 515 (7526): 203–205. Бибкод : 2014Natur.515..203H . дои : 10.1038/nature13930 . ПМИД 25337885 . S2CID 205241353 .
- ^ Стеттнер, Б.; Лица, WS; Карри, Пи Джей (2018). «След гигантского зауропода из нэмегтской свиты (верхний мел) Монголии». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 168–172. Бибкод : 2018PPP...494..168S . дои : 10.1016/j.palaeo.2017.10.027 .
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон: Издательство Принстонского университета. п. 129. ИСБН 9780691167664 .
- ^ Молина-Перес; Ларраменди (2016). Записи и курьезы о динозаврах-тероподах и других динозаврах . Барселона, Испания: Ларусс. п. 268.
- ^ Кампионе, Николас Э.; Эванс, Дэвид С. (2020). «Точность и точность оценки массы тела нептичьих динозавров» . Биологические обзоры . 95 (6): 1759–1797. дои : 10.1111/brv.12638 . ISSN 1469-185Х . ПМИД 32869488 . S2CID 221404013 .
- ^ Jump up to: а б с Сентер, П.; Робинс, Дж. Х. (2010). «Высота бедер гигантских динозавров-теропод Deinocheirus mirificus и Therizinosaurus cheloniformis и значение для музейного монтажа и палеоэкологии» (PDF) . Бюллетень Музея естественной истории Гуммы (14): 1–10. Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 9 мая 2010 г.
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 112 . ISBN 978-0-691-13720-9 .
- ^ Валкенбург, БВ; Мольнар, Р.Э. (2002). «Сравнение хищников-динозавров и млекопитающих» . Палеобиология . 28 (4): 530–540. doi : 10.1666/0094-8373(2002)028<0527:dampc>2.0.co;2 . JSTOR 3595499 . S2CID 85725299 .
- ^ Jump up to: а б Пол, GS (1988). Хищные динозавры мира . Саймон и Шустер. стр. 228, 382 . ISBN 978-0-671-61946-6 .
- ^ Миддлтон, КМ; Гейтси, С.М. (2000). «Дизайн и эволюция передних конечностей теропода» . Зоологический журнал Линнеевского общества . 128 (2): 160, 172. doi : 10.1111/j.1096-3642.2000.tb00160.x .
- ^ Jump up to: а б Ватанабэ, А.; Евгения Леоне Голд, М.; Брусатте, СЛ; Бенсон, РБЖ; Шуаньер, Ж.; Дэвидсон, А.; Норелл, Массачусетс; Классенс, Л. (2015). «Пневматичность позвонков у орнитомимозавра Archaeornithomimus (Dinosauria: Theropoda), выявленная с помощью компьютерной томографии, и переоценка осевой пневматичности у орнитомимозаврии» . ПЛОС ОДИН . 10 (12): e0145168. Бибкод : 2015PLoSO..1045168W . дои : 10.1371/journal.pone.0145168 . ПМЦ 4684312 . ПМИД 26682888 .
- ^ Маковицкий, П.Дж.; Кобаяши, Ю.; Карри, Пи Джей (2004). «Орнитомимозаврия». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. стр. 137–150. ISBN 978-0-520-24209-8 .
- ^ Серрано-Браньяс, CI; Эспиноса-Чавес, Б.; Маккракен, ЮАР; Гутьеррес-Бландо, К.; де Леон-Давила, К.; Вентура, Дж. Ф. (2020). « Paraxenisaurusnormalensis , крупный дейнохейрид-орнитомимозавр из формации Серро-дель-Пуэбло (верхний мел), Коауила, Мексика». Журнал южноамериканских наук о Земле . 101 : 102610. Бибкод : 2020JSAES.10102610S . дои : 10.1016/j.jsames.2020.102610 . S2CID 218968100 .
- ^ Лаутерс, П.; Ли, Ю.Н.; Барсболд, Р. ; Карри, Пи Джей ; Кобаяши, Ю.; Эскуийе, ФО; Годфруа, П. (2014). «Мозг Deinocheirus mirificus , гигантского динозавра-орнитомимозавра из мелового периода Монголии» (PDF) . Рефераты статей Общества палеонтологии позвоночных : 166. Архивировано из оригинала (PDF) 4 декабря 2014 г.
- ^ Кундрат, М.; Ли, Ю.Н. (2015). «Первые сведения о микроструктуре кости Deinocheirus mirificus » (PDF) . 13-е ежегодное собрание Европейской ассоциации палеонтологов позвоночных : 25. Архивировано из оригинала (PDF) 20 июля 2015 г .. Проверено 17 июля 2015 г.
- ^ Рой, Б.; Райан, MJ; Карри, Пи Джей; Коппельхус, Е.Б.; Цогтбаатар, К. (2018). «Гистологический анализ гастралий Deinocheirus mirificus из нэмегтинской свиты Монголии». 6-е ежегодное собрание Канадского общества палеонтологии позвоночных, 14–16 мая 2018 г., Оттава, Онтарио . Оттава. п. 46.
- ^ Баттон, диджей; Занно, LE (2019). «Повторяющаяся эволюция различных способов травоядности у нептичьих динозавров» . Современная биология . 30 (1): 158–168.e4. дои : 10.1016/j.cub.2019.10.050 . ПМИД 31813611 . S2CID 208652510 .
- ^ Ма, Вайсум; Питтман, Майкл; Батлер, Ричард Дж.; Лаутеншлагер, Стефан (2022). «Макроэволюционные тенденции в механике питания тероподных динозавров» . Современная биология . 32 (3): 677–686.e3. дои : 10.1016/j.cub.2021.11.060 . ПМИД 34919807 . S2CID 245257271 .
- ^ Rozhdestvensky, A.K. (1970). "Gigantskiye kogti zagadochnykh mezozoyskikh reptiliy" [Giant claws of enigmatic Mesozoic reptiles]. Paleontologicheskii Zhurnal (in Russian). 1970 (1): 117–125.
- ^ Чинзориг, Т.; Кобаяши, Ю.; Цогтбаатар, К.; Карри, Пи Джей; Такасаки, Р.; Танака, Т.; Иидзима, М.; Барсболд, Р. (2017). «Орнитомимозавры из немегтинской свиты Монголии: морфологическая изменчивость и разнообразие кисти». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 91–100. Бибкод : 2018PPP...494...91C . дои : 10.1016/j.palaeo.2017.10.031 .
- ^ Мольнар, Р.Э. (2001). «Палеопатология теропод: обзор литературы» . В Танке, Даррен Х.; Карпентер, Кеннет (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. п. 343 . ISBN 978-0-253-33907-2 .
- ^ Jump up to: а б с Ежикевич, Т.; Рассел, Д.А. (1991). «Позднемезозойская стратиграфия и позвоночные животные бассейна Гоби» . Меловые исследования . 12 (4): 345–377. Бибкод : 1991CrRes..12..345J . дои : 10.1016/0195-6671(91)90015-5 .
- ^ Салливан, РМ (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» (PDF) . В Лукасе, Спенсер Г.; Салливан, Роберт М. (ред.). Позднемеловые позвоночные Западного Внутреннего региона . Том. 35. Бюллетень Музея естественной истории и науки Нью-Мексико. стр. 347–366.
- ^ Градштейн, FM; Огг, Дж.Г.; Смит, АГ (2005). Геологическая шкала времени, 2004 г. Издательство Кембриджского университета. стр. 344–371. ISBN 978-0-521-78142-8 .
- ^ Новачек, М. (1996). Динозавры Пылающих скал . Якорь. п. 133 . ISBN 978-0-385-47775-8 .
- ^ Efimov, M.B. (1983). "Peresmotr iskopayemykh krokodilov Mongolii" [Revision of the fossil crocodiles of Mongolia]. Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya Trudy (in Russian). 24 : 76–96.
- ^ Тернер, А.Х. (2015). «Обзор шамозуха и параллигатора (Crocodyliformes, Neosuria) из мела Азии» . ПЛОС ОДИН . 10 (2): e0118116. Бибкод : 2015PLoSO..1018116T . дои : 10.1371/journal.pone.0118116 . ПМК 4340866 . ПМИД 25714338 .
- ^ Хурум, Дж. Х.; Сабат, К. (2003). «Гигантские динозавры-тероподы из Азии и Северной Америки: сравнение черепов Tarbosaurus bataar и Tyrannosaurus rex » (PDF) . Acta Palaeontologica Polonica . 48 (2): 188.
- ^ Хольц, Т.Р. (2004). «Тираннозавроидея». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. п. 124. ИСБН 978-0-520-24209-8 .
Внешние ссылки
[ редактировать ]- TEDx Talk о дейнохейрусе с участием Филипа Дж. Карри на YouTube
- Компьютерная анимация дейнохейруса , созданная Корейским институтом геонаук и минеральных ресурсов, на YouTube















| Базы данных органов управления : Национальные |
|---|