Сюнгуаньлун
| Сюнгуаньлун | |
|---|---|

| |
| Скелет-монтировка на выставке в Китае рядом с монтировкой Бэйшаньлун. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Суперсемейство: | † Тираннозавроидеа |
| Клэйд : | † Пантираннозаврия |
| Род: | †Сюнгуаньлун Ли и др. , 2009 г. |
| Типовой вид | |
| † Сюнгуаньлун баймоенсис Ли и др. , 2009 г.
| |
Xiongguanlong вымерший род тираннозавроидных — теропод из раннего мелового периода на территории современного Китая . Тип и единственный вид — X. baimoensis . Общее название происходит от города Цзяюгуань (ранее называвшегося « Сюн Гуань » или «большой перевал») и мандаринского слова « длинный », что означает дракон . Видовой эпитет « baimoensis » представляет собой латинизацию мандаринского слова, обозначающего «белый призрак», в отношении одной из геологических особенностей типовой местности (прозванной «Замком Белого Призрака»). [2]
Открытие
[ редактировать ]
Сюнгуаньлун был обнаружен в верхней части формации Сягоу группы Синьминьбао в местности, получившей прозвище «Замок Белого Призрака», которая находится в бассейне Юйцзинцзы в Ганьсу , Китай . [3] Это то же место, где обнаружен голотип орнитомимозавра был Бэйшаньлуна . [3] [4]
Он был обнаружен и подготовлен в 2006 и 2007 годах Марком Нореллом , Питером Маковицки и командой учёных из Пекинского университета . [1] В своем первоначальном описании авторы предполагают, что он происходит либо из аптского , либо из альбского ярусов . [2] но последующие авторы ограничивали верхнюю пачку именно аптским ярусом. [1]
Голотипу и единственному экземпляру Сюнгуаньлуна было присвоено обозначение FRDC-GS JB16-2-1, и он был передан на хранение в Центр исследований и разработок ископаемых Бюро георазведки и разработки полезных ископаемых провинции Ганьсу в Ланьчжоу , Китай. [4] Он состоит из полного черепа (без нижних челюстей), полного ряда шейных и спинных позвонков , частичной правой подвздошной кости и правой бедренной кости . х годов, но не был описан до 2009 года была опубликована статья , когда в Трудах Королевского общества палеонтологов Первоначально он был обнаружен в начале 2000 - Дацин Ли, Марка Норелла , Кэ-Цинь Гао, Натана Д. Смита и Питера Дж. . Маковицкий. [2]
В его первоначальном описании и в многочисленных публикациях с тех пор он был отмечен как примечательный переходный вид между базальными тираннозавроидами юрского периода и производными тираннозавридами позднего мела . [2] [5] [6]
Описание
[ редактировать ]
Авторы первоначального описания подсчитали, что Сюнгуаньлун имел длину около 5 метров (16 футов). Они использовали регрессионный анализ, основанный на работах П. Кристиансена и Р. А. Фариньи, чтобы оценить его массу примерно в 270 килограммов (600 фунтов). [2] [7] Однако, по оценкам Грегори С. Пола , он весил всего около 200 килограммов (440 фунтов). [8] Считается, что голотип полностью или почти полностью вырос из-за наблюдаемого закрытия нейроцентральных швов. Его предполагаемый размер намного больше, чем у более ранних тираннозавроидов, таких как Дилонг и Гуанлун , что привело авторов к предположению, что увеличение размеров тела тираннозавроидов было непрерывным процессом на протяжении всего мелового периода . Хотя большая часть ног и хвоста не сохранилась, считается, что пропорции тела у него были схожи с Алиорамусом . [2]
К аутапоморфиям рода, приведенным авторами в его описании, относятся следующие: очень удлиненная предглазничная область, базикраниум , ширина которого превышает длину, отсутствие пневматических отверстий на латеральном крае носовой кости , гребень на середина предчелюстных зубов и латеральные отростки осевых нервных отростков. От более поздних тираннозавроидов он также отличается отсутствием складок на носовой кости и отсутствием пневматических элементов квадратной кости . [2]
Череп
[ редактировать ]Череп голотипа сохранился полностью, включая небо и черепную коробку , но лишен нижней челюсти . Он очень удлинен по сравнению с большинством других тираннозавроидов , со значительным удлинением верхнечелюстных , слезных и заглазничных костей . Несмотря на относительно полную целостность, череп не очень хорошо сохранился. Он окаменел в минерализованном узле, что привело к трудностям в дифференциации кости от матрицы, в которой она сохранилась. Это привело к затруднениям в определении общей топологии костей черепа и сокрытию существования верхнечелюстных окон . Однако, несмотря на качество сохранности, очевидны многочисленные отличительные особенности. [2]
Одной из отличительных особенностей черепов большинства тираннозавроидов являются носовые кости сросшиеся . Xiongguanlong также демонстрирует эту особенность, но ему не хватает сагиттальных складок, присутствующих у производных тираннозаврид . Носовая часть также демонстрирует развитую особенность отсутствия пневматических элементов на боковых сторонах носовой части, что наблюдается у менее производных родов, таких как Dilong и Eotyrannus . [2]
Остальная часть черепа также характерно компактна. Предлобная кость небольшая, треугольной формы, с глазницей не соприкасается . Теменные кости срослись и имеют короткий вогнутый сагиттальный гребень, служивший местом прикрепления кусающих мышц. Квадратная кость также большая и имеет расширенные латеральные мыщелки, которые, как предполагается, также связаны с мускулатурой челюсти. Заглазничная кость сильно удлинена в передне-заднем направлении и вертикальна по отношению к подвисочному оконному проему , что напоминает продвинутых тираннозаврид. Небо также очень прочное из -за глубокого, похожего на лезвие сошника и больших, глыбистых эктоптеригоидных костей. [2]
Некоторые зубы сохранились не очень хорошо, но зубные альвеолы видны . Это побудило авторов его описания определить, что у Сюнгуаньлуна было четыре предчелюстных зуба и пятнадцать верхнечелюстных зубов. Предчелюстные зубы имеют характерную d-образную форму в поперечном сечении и зазубрены, как и у большинства других тираннозавроидов. Однако верхнечелюстные зубы отличаются от более продвинутых тираннозавроидов тем, что они узкие в латеро-медиальном направлении и более похожи на лезвия. [2]
Посткраниальный скелет
[ редактировать ]
Сохранившиеся посткраниальные кости Сюнгуаньлуна включают весь ряд шейных и спинных позвонков , часть подвздошной и бедренной кости . Некоторые элементы скелета имеют сходство с другими тираннозавроидами , а также с некоторыми нетираннозавроидными тероподами . [2]
шейного отдела Центры сильно сращены с нервными дугами, центры вогнуты в переднем направлении и выпуклы в заднем направлении. У них также есть пара пневматических отверстий на позвонках, что не похоже на состояние, наблюдаемое у альбертозавра , дасплетозавра и тираннозавра . Спинные позвонки имеют широко расширенные нервные дуги, которые полностью перекрывают центры, если смотреть сверху, хотя это не относится к шейным позвонкам, которые больше напоминают нервные дуги, наблюдаемые у Дилонга . Высота нервных отростков увеличивается кзади, хотя точная высота задних спинных позвонков неизвестна, поскольку позвонки были сильнее повреждены во время окаменения . [2]
Сохранившиеся тазобедренные элементы Xiongguanlong напоминают таковые у производных тираннозавроидов, а также орнитомимозавров . Бедренная кость также очень похожа на производных тираннозавроидов относительно расположения и протяженности четвертого вертела . Вместе с остальной частью скелета также была обнаружена большеберцовая кость , хотя ее сравнительно небольшой размер привел Ли и его коллег к выводу, что она, вероятно, принадлежала не тому же животному, что и остальные кости. [2]
Классификация
[ редактировать ]Филогения
[ редактировать ]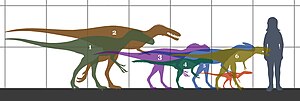
В своем описании Xiongguanlong в 2009 году Ли и его коллеги опубликовали филогенетический анализ tyrannosauroidea . Они обнаружили Сюнгуаньлуна как члена этой группы на основании следующих синапоморфий : вертикальный гребень на средней линии подвздошной кости , выемка на дорсальном крае подвздошной кости, сросшиеся носовые кости и предчелюстные зубы, намного меньшие, чем верхнечелюстные зубы. [2]
Они также обнаружили несколько клад внутри tyrannosauroidea, которым на момент описания еще не было присвоено официальное название. Первая из них («клада 1» на схеме) представляла собой кладу, включавшую всех тираннозавроидов, кроме процератозавридов . Эта клада была диагностирована по следующим синапоморфиям: носовые кости в форме песочных часов, увеличенное отверстие на квадратной кости и широкая заглазничная перемычка. [2] Эта клада также была обнаружена Рафаэлем Делькуром и Орландо Нельсоном Грилло в их филогенетическом исследовании тираннозавроидов в 2018 году . [9] Однако в их анализе синапоморфии, обнаруженные для этой клады, в первую очередь связаны с особенностями бедренных костей (в частности, подвздошной и седалищной костей ), без упоминания каких-либо объединяющих особенностей черепа. Они назвали эту кладу « пантираннозаврия » в связи со своей гипотезой о том, что эта клада имела глобальное распространение . [9]
Ли и его коллеги обнаружили, что Xiongguanlong принадлежит к немного более развитой кладе («клад 2» на диаграмме), которая включает таксоны, более тесно связанные с тираннозавридами , чем с Dilong или Eotyrannus . Они диагностируют эту кладу, используя следующие синапоморфии: медиальный гребень на предчелюстных зубах, квадратно-скуловую кость , которая расширяется наружу, базисфеноидную кость, ширина которой превышает длину, и латеральные отростки на осевых нервных отростках. [2] Эта клада конкретно не упоминается и не упоминается авторами каких-либо последующих анализов. [5] [6] [9] [10]
Они также обнаруживают третью, более эксклюзивную кладу («клада 3» на диаграмме), которая исключает Xiongguanlong и включает все таксоны, более тесно связанные с тираннозавридами. Эта клада была диагностирована, и Сюнгуаньлун был исключен из нее, по следующим синапоморфиям: морщинистости на вершине носовой кости и подмыщелковым углублениям в черепной коробке . Они обнаруживают еще несколько синапоморфий для каждой из этих клад, но они менее подтверждены, чем однозначные синапоморфии, перечисленные выше. [2] Эта клада была впоследствии обнаружена Делькуром и Грилло и получила название « эутираннозаврия »; однако обнаруженные ими однозначные синапоморфии этой клады связаны исключительно с посткраниальным скелетом. [9] Краткое изложение филогенетического анализа Ли и его коллег, включая обнаруженные ими клады, можно увидеть ниже. [2]
Подобное филогенетическое расположение Xiongguanlong было впоследствии обнаружено в анализе Сюй Сина и его коллег (2012). [11] Брусатте и коллеги (2016), [5] Брусатте и Карр (2016), [12] и Делькур и Грилло (2018), [9] Занно и коллеги (2019), [13] и Вулф и коллеги (2019). [14]
Филогенетический анализ (Ли и др., 2010) [2]
| |||||||||||||||||||||||||||
В 2013 году группа учёных под руководством Марка Лоуэна, в которую входили Филип Дж. Карри и Скотт Д. Сэмпсон , опубликовала описание тираннозаврид рода Lythronax , которое также содержало переоценку родственного таксона Teratophoneus . Они провели филогенетический анализ, используя набор данных из 54 таксонов (включая 26 таксонов тираннозавроидов) и более 300 оцененных признаков, чтобы проанализировать филогенетические отношения Lythronax и Teratophoneus . [10]

Их результаты выявили относительно уникальную топологию, которая противоречит анализам Брусатте и его коллег (2010) и Карру и коллег (2010), которые были наиболее современными анализами до их собственной публикации. Левен и его коллеги обнаружили, что Bistahieversor относится к тираннозавридам, а Alioramus - как к находящемуся за пределами этого семейства. Они также предполагают, что спорный род Raptorex является отдельным таксоном, независимо от каких-либо гипотез его онтогенеза . [10] Они также обнаружили Xiongguanlong и Alectrosaurus как сестринские таксоны и являются более производными, чем Dryptosaurus , что поместило бы их в эвтираннозаврию согласно определению, данному Делькуром и Грилло (наименее инклюзивная клада, содержащая как тираннозавра , так и дриптозавра ). [9] [10] Сокращенную версию филогении, которую они обнаружили, можно увидеть ниже. [10]

Самый последний комплексный филогенетический анализ tyrannosauroidea был проведен Дарреном Нэйшем и Андреа Кау в их монографии об остеологии и филогении Eotyrannus в 2022 году . Анализ, который они проводят в своей публикации, включал 83 таксона и более 1100 морфологических признаков, что привело к появлению нескольких новых гипотез. В их выводах больше всего обсуждается вопрос о том, что мегараптора является сестринским таксоном эутираннозаврия . Нэйш и Кау предполагают, что уникальность этого анализа является следствием отсутствия обширных выборок мегарапторов в предыдущих анализах. Они обнаружили Xiongguanlong как сестринский таксон «клады megaraptora + eutyrannosauria», хотя они не перечислили никаких синапоморфий этой клады. Они также обсуждают последствия, которые это имеет для палеобиогеографии тираннозавроидов, в частности, то, что они, вероятно, произошли из Евразии . Сокращенную версию дерева строгого консенсуса, полученную в результате их анализа, можно увидеть ниже. [6]
Эволюционные последствия
[ редактировать ]Информация об эволюции тираннозавроидов была относительно неясной с момента присвоения группе названия в начале 20 века. Есть несколько базальных таксонов, которые известны из средней юры (например, Guanlong , Kileskus , Proceratosaurus ), несколько более промежуточных форм из раннего мела (например, Yutyrannus , Dilong , Eotyrannus ), а также многочисленные мегафаунистические гиперплотоядные роды из кампана и маастрихта. (например, Тираннозавр , Альбертозавр , Дасплетозавр ). [9] Исследователи, описавшие таксоны «среднего мела», в том числе Xiongguanlong , [2] Тимурленгия , [5] мавры , [13] и Сускитираннус [14] отметил, что эти таксоны частично заполняют этот «пробел в тираннозаврах мелового периода». [6] [15]
Существует множество теорий относительно биогеографии тираннозавроидов среднего мелового периода. Суша Лавразии находилась в процессе распада в раннем меловом периоде , а это означало, что Атлантический океан был намного уже. Были также тираннозавроиды, известные из раннего мела Европы, такие как Eotyrannus , что заставило некоторых исследователей предположить, что тираннозавроиды мигрировали в Северную Америку из Европы , возможно, либо путем рафтинга , либо по сухопутному мосту . [15]
Было также высказано предположение, что к этому времени уже сформировалась суша Берингии , что позволило бы мигрировать животных из Азии в Ларамидию . Это подтверждается появлением в Северной Америке животных с очевидным недавним азиатским происхождением, включая Eolambia и Microvenator . [9] [14] [15] Зубы тираннозавроидов известны из апта - альбского возраста формации Клеверли в Вайоминге , а это означает, что некоторые тираннозавроиды, вероятно, мигрировали в Северную Америку до 108 миллионов лет назад. [15] Другие исследователи полагают, что тираннозавроиды уже обладали глобальным распространением к поздней юре . [9] Однако некоторые авторы полагают, что летопись окаменелостей этой части мелового периода слишком скудна, чтобы делать какие-либо конкретные выводы относительно природы эволюции или расселения тираннозавроидов. [12]
Палеоэкология
[ редактировать ]
Диета
[ редактировать ]Сюнгуаньлун — крупнейший хищник, обнаруженный на данный момент в формации Сягоу . Считается, что более крупные тероподы, обитающие в этом районе, Бэйшаньлун и Сучжоузавр , являются травоядными. [1] [2] [4]
Это почти наверняка было облигатным хищником, как и все другие известные тираннозавроиды. [16] Однако он не обладал приспособлениями для сильной силы укуса, характерными для производных тираннозаврид . [2] Считается, что подобные таксоны, такие как Suskityrannus, были хищниками среднего размера в своей экосистеме, но крупных хищных теропод в этой области пока не обнаружено. [16] происходил эволюционный переход то, что в позднем апте Китая что может указывать на . [12]
Палеосреда
[ редактировать ]Формация Сягоу является второй самой молодой формацией группы Синьминьбу и стратиграфически расположена между более старой формацией Чинджинпу и более молодой формацией Чжунгоу, все из которых расположены в бассейне Юйцзинцзы на западе Ганьсу . Сегодня этот регион является частью пустыни Гоби , но в раннемеловом периоде здесь была гораздо менее засушливая среда, и, вероятно, он находился на побережье моря Тетис или вблизи него . [17]
Отложения свиты представлены аргиллитами и алевролитами от серого до серого цвета с прослоями песчаников и конгломератов. [18]
Состав формации Сягоу указывает на то, что, вероятно, существовали низменности, окруженные узкими холмами, что позволяло фауне динозавров путешествовать по территориям без географических барьеров. У некоторых окаменевших водоплавающих птиц сохранились мягкие ткани их перепончатых ног и перья, что привело авторов к выводу, что озера с бескислородными озёрными днами. также существовали [17]
Современная фауна
[ редактировать ]

Предполагается, что фауна, присутствующая в группе Синьминьбао, является экологическим преемником гораздо более известной биоты Джехоль , которая существовала в Китае в барреме и раннем апте . Эта фауна, названная некоторыми авторами «фауной Мазонгшань», состоит в основном из динозавров , поскольку в породах формации Сягоу сохранились преимущественно крупные позвоночные животные. [1] Есть серьезное исключение из этой тенденции — птицы. В биоте Мазонгшань имеются многочисленные хорошо сохранившиеся окаменелости птиц, большинство из которых представляют собой энантиорнитины , с относительно небольшим количеством орнитуроморфов . В этом регионе также сохранилось несколько окаменелостей рыб и членистоногих. [19] [20]
Самые многочисленные останки, найденные в регионе, — это нептичьи динозавры. Из них мелкие цератопсы были наиболее многочисленными и на их долю приходится наибольшее количество окаменелостей. Встречаются также базальные гадрозавроиды , титанозаврообразные зауроподы , крупные травоядные орнитомимозавры и теризинозавры . Другие фрагментарные останки относят к овирапторозаврам , но некоторые авторы ставят их под сомнение. [1]
Известно, что Сюнгуаньлун сосуществовал с большим орнитомимозавром Бэйшаньлуном . [2] который был найден в том же месте, [3] а некоторые музейные экспонаты изображают отношения хищник-жертва между этими животными. современные животные включали Сюйвулонга и Цяованлуна Джинтазавра , эвгелоподида и Другие орнитоподов теризинозавра Сучжоузавра . небольшой неоцератопсовый археоцератопс . В этом районе также обитает [1] Энантиорнитиновые птицы были многочисленны и включали такие роды, как Avimaia , Feitianius и несколько безымянных форм, и их сопровождали ранние эворнитовые птицы, такие как утиноподобный Gansus и необычный зубастый Brevidentavis . [20] Также было найдено несколько скелетов черепах. [1]
См. также
[ редактировать ]- 2009 г. по палеонтологии архозавров.
- Провинциальный музей Ганьсу
- депозит
- Китайско-шведская экспедиция
- Хронология исследований тираннозавров
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час Ты, Х.; Моршхаузер, Э.М.; Ли, Д.; Додсон, П. (2018). «Знакомство с фауной динозавров Мазонгшаня» . Журнал палеонтологии позвоночных . 38 (доп. 1): 1−11. дои : 10.1080/02724634.2017.1396995 . S2CID 202867591 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х Ли, Дацин; Норелл, Марк А.; Гао, Кэ-Цинь; Смит, Натан Д.; Маковицкий, Питер Дж. (2009). «Лонгиростровый тираннозавроид из раннего мела Китая» . Труды Королевского общества B: Биологические науки . 277 (1679): 183–190. дои : 10.1098/rspb.2009.0249 . ПМЦ 2842666 . ПМИД 19386654 .
- ^ Перейти обратно: а б с Каррано, Мэтью (2009). «Замок Белого Призрака (мел Китая), также известный как бассейн Юцзинцзы» . База данных палеобиологии . Проверено 15 августа 2023 г.
Где: Ганьсу, Китай (40,5° с.ш., 98,1° в.д.: палеокоординаты 40,0° с.ш., 98,6° в.д.); Когда: верхняя пачка (формация Сягоу), апт (125,0–113,0 млн лет назад); Саурисхия — Xiongguanlong baimoensis n. ген. н. сп.
- ^ Перейти обратно: а б с Маковицкий, Питер Дж.; Ли, Дацин; Гао, Кэ-Цинь; Левин, Мэтью; Эриксон, Грегори М.; Норелл, Марк А. (2010). «Гигантский орнитомимозавр из раннего мела Китая» . Труды Королевского общества B: Биологические науки . 277 (1679): 191–198. дои : 10.1098/rspb.2009.0236 . ПМЦ 2842665 . ПМИД 19386658 .
- ^ Перейти обратно: а б с д Брусатте, Стивен Л.; Аверьянов, Александр; Сьюс, Ханс-Дитер; Мьюир, Эми; Батлер, Ян Б. (2016). «Новый тираннозавр из середины мелового периода Узбекистана проясняет эволюцию гигантских размеров тела и развитых чувств у динозавров-тиранов» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (13): 3447–3452. Бибкод : 2016PNAS..113.3447B . дои : 10.1073/pnas.1600140113 . ПМЦ 4822578 . ПМИД 26976562 .
- ^ Перейти обратно: а б с д Нэйш, Д.; Кау, А. (июль 2022 г.). «Остеология и сходство Eotyrannus lengi , теропода-тираннозавра из супергруппы Уилдена на юге Англии» . ПерДж . 10 : е12727. дои : 10.7717/peerj.12727 . ПМЦ 9271276 . ПМИД 35821895 .
- ^ Кристиансен, П.; Фаринья †, РА (2004). «Массовое предсказание динозавров-теропод» . Историческая биология . 16 (2–4): 85–92. дои : 10.1080/08912960412331284313 . S2CID 84322349 .
- ^ Пол, Грегори С. (2024). Принстонский полевой справочник по динозаврам (третье издание) . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0691231570 .
- ^ Перейти обратно: а б с д и ж г час я Делькур, Рафаэль; Грилло, Орландо Нельсон (2018). «Тираннозавроиды из южного полушария: значение для биогеографии, эволюции и таксономии» . Палеогеография, Палеоклиматология, Палеоэкология . 511 : 379–387. Бибкод : 2018PPP...511..379D . дои : 10.1016/j.palaeo.2018.09.003 . S2CID 133830150 .
- ^ Перейти обратно: а б с д и Лоуэн, Массачусетс; Ирмис, РБ; Сертич, JJW; Карри, Пи Джей; Сэмпсон, SD (2013). Эванс, Дэвид С. (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение позднемеловых океанов» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- ^ Сюй, Кебай; Чжан, Кэ; Ма, Цинъюй » . Природа . 484 (7392): 92–95. Бибкод : 2012Natur.484...92X doi : 10.1038 /nature10906 . PMID 22481363. . S2CID 29689629 .
- ^ Перейти обратно: а б с Брусатте, Стивен Л.; Карр, Томас Д. (2016). «Филогения и история эволюции тираннозавроидов» . Научные отчеты . 6 : 20252. Бибкод : 2016NatSR...620252B . дои : 10.1038/srep20252 . ПМЦ 4735739 . ПМИД 26830019 .
- ^ Перейти обратно: а б Занно, Линдси Э.; Такер, Райан Т.; Кановиль, Аврора; Авраами, Хавив М.; Гейтс, Терри А.; Маковицкий, Питер Дж. (2019). «Маленький быстроногий тираннозавроид сокращает разрыв в 70 миллионов лет в летописи окаменелостей Северной Америки» . Коммуникационная биология . 2 : 64. doi : 10.1038/s42003-019-0308-7 . ПМК 6385174 . ПМИД 30820466 .
- ^ Перейти обратно: а б с Вульф, генеральный директор; Макдональд, AT; Киркланд, Дж.И.; Тернер, А.Х.; Смит, Северная Дакота; Брусатте, СЛ; Лоуэн, Массачусетс; Дентон, РК; Несбитт, SJ (6 мая 2019 г.). «Тираннозавроид среднего мела и происхождение скоплений динозавров конца мела в Северной Америке» (PDF) . Экология и эволюция природы . 3 (6): 892–899. дои : 10.1038/s41559-019-0888-0 . hdl : 20.500.11820/a6709b34-e3ab-416e-a866-03ba1162b23d . ПМИД 31061476 . S2CID 256707614 .
- ^ Перейти обратно: а б с д Занно, Линдси Э.; Маковицкий, Питер Дж. (2011). «О самых ранних находках меловых тираннозавроидов на западе Северной Америки: последствия для события обмена в раннем меловом периоде в Лавразии» . Историческая биология . 23 (4): 317–325. дои : 10.1080/08912963.2010.543952 . S2CID 140721871 .
- ^ Перейти обратно: а б Хольц, Томас Р. (2021). «Структура гильдии теропод и гипотеза ассимиляции ниш тираннозаврид: значение для макроэкологии и онтогенеза хищных динозавров в позднем меловом периоде в Азии». 1 « . Канадский журнал наук о Земле . 58 (9): 778–795. doi : 10.1139/cjes-2020-0174 . hdl : 1903/28566 .
- ^ Перейти обратно: а б Ты, Хай-лу; Ламанна, Мэтью С.; Харрис, Джеральд Д.; Кьяппе, Луис М.; О'Коннор, Цзинмай; Цзи, Шу-ан; Лю, Цзюнь-чан; Юань, Чун-си; Ли; Чжан, Син; Лаковара, Кеннет Дж.; Додсон, Питер и Цзи, Цян, Да-цин (2006). «Почти современная птица-амфибия из раннего мела Северо-Западного Китая» . Наука . 312 (5780): 1640–1643. Бибкод : 2006Sci...312.1640Y . дои : 10.1126/science.1126377 . ПМИД 16778053 . S2CID 42723583 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Суарес, Марина Б.; Милдер, Тимоти; Пэн, Нан; Суарес, Селина А.; Ты, Хайлу; Ли, Дацин; Додсон, Питер (13 декабря 2018 г.). «Хемостратиграфия формаций Сягоу и Чжунгоу, содержащих динозавров нижнего мела, бассейн Юйцзинцзы, северо-запад Китая». Журнал палеонтологии позвоночных . 38 (суп1): 12–21. дои : 10.1080/02724634.2018.1510412 . ISSN 0272-4634 . S2CID 202865132 .
- ^ AM Мюррей, HL You и К. Пэн. 2010. Новая меловая рыба-остеоглоссоморф из провинции Ганьсу, Китай. Журнал палеонтологии позвоночных 30 (2): 322-332.
- ^ Перейти обратно: а б Ван, Ю.; О'Коннор, Дж. К.; Ли, Д.; Ты, Х. (2013). «Ранее непризнанное разнообразие птиц-орнитуроморфов в раннемеловом бассейне Чангма, провинция Ганьсу, Северо-Западный Китай» . ПЛОС ОДИН . 8 (10): e77693. Бибкод : 2013PLoSO...877693W . дои : 10.1371/journal.pone.0077693 . ПМЦ 3795672 . ПМИД 24147058 .
Внешние ссылки
[ редактировать ]- «Предок тираннозавра найден в Китае» BBC News
- «Ископаемые свидетельства существования тираннозавра Златовласки» ScienceNews.org














