юрский период
| юрский период | |||||||||
|---|---|---|---|---|---|---|---|---|---|
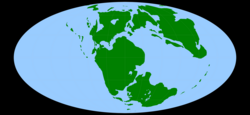 Карта мира, какой она была в средней юре, ок. 170 млн лет. [ нужна ссылка ] | |||||||||
| Хронология | |||||||||
| |||||||||
| Этимология | |||||||||
| Формальность имени | Формальный | ||||||||
| Информация об использовании | |||||||||
| Региональное использование | Глобальный ( ICS ) | ||||||||
| Используемая шкала времени | Временная шкала ICS | ||||||||
| Определение | |||||||||
| Хронологическая единица | Период | ||||||||
| Стратиграфическая единица | Система | ||||||||
| Формальность временного интервала | Формальный | ||||||||
| Определение нижней границы | Первое появление аммонита Psiloceras spelae tirolicum . | ||||||||
| Нижняя граница ГССП | Участок Кухйох, горы Карвендель , Северные Известняковые Альпы , Австрия 47 ° 29'02 "N 11 ° 31'50" E / 47,4839 ° N 11,5306 ° E | ||||||||
| Нижний GSSP ратифицирован | 2010 | ||||||||
| Определение верхней границы | Формально не определено | ||||||||
| Кандидаты на определение верхней границы |
| ||||||||
| Верхняя граница участков-кандидатов GSSP | Никто | ||||||||

период Юрский ( / dʒ ʊ ˈ r æ s ɪ k / juurr- ASS -ik [ 2 ] ) — это геологический период и стратиграфическая система , охватывающая период с конца триасового периода 201,4 миллиона лет назад (млн лет назад) до начала мелового периода, примерно 145 млн лет назад. Юрский период представляет собой средний период мезозойской эры и назван в честь гор Юра , где впервые были обнаружены пласты известняка этого периода.
Начало юрского периода ознаменовалось крупным триасово-юрским вымиранием , связанным с извержением Центрально -Атлантической магматической провинции (CAMP). Начало тоарского этапа началось около 183 миллионов лет назад и отмечено Тоарским океаническим бескислородным событием , глобальным эпизодом океанической аноксии , закисления океана и повышенных глобальных температур, связанных с вымиранием, вероятно, вызванным извержением Кару-Феррара. крупные магматические провинции . Конец юрского периода, однако, не имеет четкой, окончательной границы с меловым периодом и является единственной границей между геологическими периодами, которая остается формально неопределенной.
К началу юрского периода суперконтинент Пангея начал раскалываться на два массива суши: Лавразию на севере и Гондвану на юге. Климат юрского периода был теплее современного, ледяных шапок не было . Вблизи полюсов росли леса, а в нижних широтах располагались большие засушливые пространства.
На суше фауна перешла от триасовой фауны, в которой доминировали совместно динозавроморфы и псевдозуховые архозавры , к фауне, в которой доминировали одни динозавры . Первые птицы появились в юрском периоде, развившись из ветви тероподовых динозавров. Другие важные события включают появление первых крабов и современных ящериц , а также диверсификацию ранних млекопитающих . Крокодиломорфы совершили переход от наземного образа жизни к водному. Океаны были населены морскими рептилиями, такими как ихтиозавры и плезиозавры , тогда как птерозавры были доминирующими летающими позвоночными животными . В этот период впервые появились и разнообразили современные акулы и скаты. Во флоре преобладали папоротники и голосеменные растения , включая хвойные , из которых в этот период впервые появились многие современные группы, а также другие группы — вымершие Bennettitales .
Этимология и история
[ редактировать ]
Хроностратиграфический термин «юрский период » связан с горами Юра , лесистым горным хребтом , который в основном следует за границей Франции и Швейцарии . Название «Юра» происходит от кельтского корня * jor через галльское *iuris «лесистая гора», которое было заимствовано на латынь как название места и превратилось в Juria и, наконец, в Jura .
Во время поездки по региону в 1795 году немецкий натуралист Александр фон Гумбольдт признал карбонатные отложения в горах Юра геологически отличными от триасового возраста мушелькалька на юге Германии , но он ошибочно пришел к выводу, что они старше. Затем в 1799 году он назвал их Юра-Калькштейн («Юрский известняк»). [ 3 ]
В 1829 году французский натуралист Александр Броньяр опубликовал книгу « Описание территорий, составляющих земную кору, или Очерк строения известных земель Земли». В этой книге Броньяр использовал термин « юрские ландшафты », соотнося «Юра-Калькштейн» Гумбольдта с оолитовыми известняками такого же возраста в Великобритании, таким образом придумав и опубликовав термин «юрский период». [ 4 ] [ 3 ]
Немецкий геолог Леопольд фон Бух в 1839 году установил тройное деление юры, первоначально названное от древнейшей к самой молодой: черная юра , коричневая юра и белая юра . [ 5 ] Термин « Лиас » ранее использовался Уильямом Конибером и Уильямом Филлипсом для слоев, эквивалентных возрасту черной юры в Англии, в 1822 году. Уильям Филлипс, геолог, работал с Уильямом Конибером, чтобы узнать больше о черной юре в Англии.
Французский палеонтолог Альсид д'Орбиньи в статьях между 1842 и 1852 годами разделил юру на десять стадий на основе аммонитовых и других ископаемых комплексов в Англии и Франции, семь из которых используются до сих пор, но ни одна не сохранила свое первоначальное определение. Немецкий геолог и палеонтолог Фридрих Август фон Квенстедт в 1858 году разделил три серии фон Буха в Швабской Юре на шесть подразделений, определяемых аммонитами и другими окаменелостями.
Немецкий палеонтолог Альберт Оппель в своих исследованиях 1856–1858 годов изменил первоначальную схему д'Орбиньи и дополнительно подразделил этапы на биостратиграфические зоны, основанные преимущественно на аммонитах. Большинство современных этапов юрского периода были формализованы на Коллоке Юрского периода в Люксембурге в 1962 году. [ 3 ]
Геология
[ редактировать ]Юрский период делится на три эпохи : раннюю, среднюю и позднюю. Аналогичным образом в стратиграфии юра разделяется на нижнеюрскую , среднеюрскую и верхнеюрскую серии . Геологи делят породы юрского периода на стратиграфический набор подразделений, называемых этапами , каждый из которых сформировался в соответствующие временные интервалы, называемые возрастами.
Этапы могут быть определены глобально или регионально. Для глобальной стратиграфической корреляции Международная комиссия по стратиграфии (ICS) ратифицирует глобальные этапы на основе разреза и точки стратотипа глобальной границы (GSSP) из одной формации ( стратотипа ), определяющей нижнюю границу этапа. [ 3 ] Возраст юрского периода от самого молодого до самого старого следующий: [ 6 ]
| Серия/эпоха | Стадия/возраст | Нижняя граница |
| Ранний мел | Берриазия | ~145 млн лет назад |
| Верхняя/Поздняя юра | Титонский | 149,2 ± 0,7 млн лет назад |
| Кимеридж | 154,8 ± 0,8 млн лет назад | |
| оксфордский | 161,5 ± 1,0 млн лет назад | |
| Средняя юра | келловейский век | 165,3 ± 1,1 млн лет назад |
| батский | 168,2 ± 1,2 млн лет назад | |
| Байосский | 170,9 ± 0,8 млн лет назад | |
| Ааленский | 174,7 ± 0,8 млн лет назад | |
| Нижняя/ранняя юра | Тоарский | 184,2 ± 0,3 млн лет назад |
| Плинсбахский | 192,9 ± 0,3 млн лет назад | |
| синемурийский | 199,5 ± 0,3 млн лет назад | |
| геттангианский | 201,4 ± 0,2 млн лет назад |
Стратиграфия
[ редактировать ]


Стратиграфия юрского периода основана главным образом на использовании аммонитов в качестве индексных окаменелостей . Дата первого появления конкретных таксонов аммонитов используется для обозначения начала стадий, а также меньших промежутков времени внутри стадий, называемых «аммонитовыми зонами»; они, в свою очередь, также иногда подразделяются на подзоны. Глобальная стратиграфия основана на стандартных европейских аммонитовых зонах, а другие регионы калибруются по европейским последовательностям. [ 3 ]

Ранняя юра
[ редактировать ]Самая старая часть юрского периода исторически называлась Лиас или Лиас, что примерно эквивалентно по протяженности ранней юре, но также включает часть предшествующего рэтского периода . Эттанжский ярус был назван швейцарским палеонтологом Эженом Реневье в 1864 году в честь Эттанж-Гранде на северо-востоке Франции. [ 3 ] GSSP основания геттангиана расположен на перевале Кухйох, горы Карвендель , Северные Известняковые Альпы , Австрия; он был ратифицирован в 2010 г. Начало геттанга и, следовательно, юры в целом ознаменовано первым появлением аммонита Psiloceras spelae tirolicum в кендльбахской свите, обнаженной в Кухйохе. [ 7 ] Основание юрского периода ранее было определено Альбертом Оппелем как первое появление Psiloceras planorbis в 1856–1858 годах, но это было изменено, поскольку появление рассматривалось как слишком локализованное событие для международной границы. [ 3 ]
Синемюрийский этап был впервые определен и введен в научную литературу Альсидом д'Орбиньи в 1842 году. Свое название он получил от французского города Семюр-ан-Оксуа , недалеко от Дижона . Первоначальное определение синемурийского языка включало в себя то, что сейчас называется геттангианом. GSSP Синемюриана расположен на скале к северу от деревни Ист-Куантоксхед , в 6 км к востоку от Уотчетта , Сомерсет , Англия , в пределах Голубого Лиаса , и был ратифицирован в 2000 году. Начало Синемюриана определяется первым появление аммонита Vermiceras quantoxense . [ 3 ] [ 8 ]
Альберт Оппель в 1858 году назвал плинсбахскую сцену в честь деревни Плинсбах в общине Целль-унтер-Айхельберг в Швабском Альбе , недалеко от Штутгарта , Германия. GSSP для основания плинсбаха обнаружен в местности Вайн-Хейвен в заливе Робин Гуд , Йоркшир , Англия, в формации Редкар-аргиллит , и был ратифицирован в 2005 году. Начало плинсбаха определяется первым появлением аммонита. Bifericeras donovani . [ 9 ]
Деревня Туар (лат. Toarcium ), расположенная к югу от Сомюра в долине Луары во Франции , дала свое название тоарскому ярусу. Тоаркий был назван Альсидом д'Орбиньи в 1842 году, причем первоначальным местом обитания был карьер Врин, примерно в 2 км к северо-западу от Туара. GSSP для основания тоара расположен в Пенише, Португалия , и был ратифицирован в 2014 году. Граница определяется первым появлением аммонитов, принадлежащих к подроду Dactylioceras ( Eodactylites ) . [ 10 ]
Средняя юра
[ редактировать ]Ааленский назван в честь города Аален в Германии. Ааленский ярус был определен швейцарским геологом Карлом Майером-Эймаром в 1864 году. Нижняя граница первоначально проходила между темными глинами черной юры и перекрывающими глинистым песчаником и железистым оолитом коричневых юрских отложений юго-западной Германии. [ 3 ] GSSP для основания аалена расположен в Фуэнтельсасе в Иберийском хребте недалеко от Гвадалахары, Испания , и был ратифицирован в 2000 году. Основание аалена определяется первым появлением аммонита Leioceras opalinum . [ 11 ]
Альсид д'Орбиньи в 1842 году назвал Баджосскую сцену в честь города Байе (лат. Bajoce ) в Нормандии, Франция. GSSP для базы Байоса расположен на участке Муртинейра в Кабу-Мондего , Португалия; он был ратифицирован в 1997 году. Основание байоса определяется первым появлением аммонита Hyperlioceras mundum . [ 12 ]
Батский период назван в честь города Бат в Англии, введенного бельгийским геологом д'Омалиусом д'Аллоем в 1843 году, в честь неполного разреза оолитовых известняков в нескольких карьерах в этом регионе. GSSP для основания Бата — Равен-дю-Бес, район Нижний Оран, Альпы Верхнего Прованса , Франция; он был ратифицирован в 2009 году. Основание бата определяется первым появлением аммонитовой Gonolkites конвергентной зоны в основании зигзагообразной аммонитовой зоны Zigzagiceras . [ 13 ]
Келловейский век происходит от латинизированного названия деревни Келлауэйс в Уилтшире , Англия, и был назван Альсидом д'Орбиньи в 1852 году, первоначально на месте контакта между формациями Форест Мрамор и Формация Корнбраш . Однако позже выяснилось, что эта граница находится в пределах верхней части бата. [ 3 ] База келловейцев пока не имеет сертифицированного ГССП. Рабочее определение основания келловея — первое появление аммонитов, принадлежащих к роду Kepplerites . [ 14 ]
Поздняя юра
[ редактировать ]Оксфордиан назван в честь города Оксфорд в Англии и был назван Альсидом д'Орбиньи в 1844 году в отношении Оксфордской глины . В базе оксфордианца отсутствует четко выраженный GSSP. У. Дж. Аркелл в исследованиях 1939 и 1946 годов отнес нижнюю границу оксфорда к первому появлению аммонита Quenstedtoceras mariae (тогда отнесенного к роду Vertumniceras ). Последующие предложения предполагали первое появление Cardioceras redcliffense в качестве нижней границы. [ 3 ] [ 14 ]
Деревня Киммеридж на побережье Дорсета , Англия, является источником названия киммериджа. Сцена была названа Альсидом д'Орбиньи в 1842 году в честь Киммериджской глины . GSSP для основания кимериджа — разрез Флодигарри в заливе Стаффин на острове Скай , Шотландия . [ 15 ] который был ратифицирован в 2021 году. Граница определяется первым появлением аммонитов, обозначающим бореальную зону Баухини и суббореальную зону Байлей. [ 14 ]
Титониан был введен в научную литературу Альбертом Оппелем в 1865 году. Название Титониан необычно для геологических сценических названий, поскольку оно происходит из греческой мифологии, а не из географического названия. Тифон был сыном Лаомедонта Трои , и влюбился в Эос греческую богиню зари . Его имя было выбрано Альбертом Оппелем для этого стратиграфического этапа, потому что титон находится рука об руку с началом мелового периода. На базе Титона в настоящее время отсутствует GSSP. [ 3 ] Рабочее определение основания титона — первое появление аммонитов рода Gravesia . [ 14 ]
Верхняя граница юрского периода в настоящее время не определена, а граница юрского периода и мела в настоящее время является единственной границей системы, для которой отсутствует определенный GSSP. Размещение GSSP для этой границы было затруднено из-за сильной регионализации большинства биостратиграфических маркеров и отсутствия каких-либо хемостратиграфических событий, таких как изотопные отклонения (большие внезапные изменения в соотношениях изотопов ), которые можно было бы использовать для определения или корреляции границы. . Кальпионеллиды , загадочная группа планктонных протистов в форме урны, с кальцитовыми раковинами кратковременно встречавшаяся в период от поздней юры до раннего мела, были предложены как наиболее многообещающие кандидаты для установления границы юры и мела. [ 16 ] первое появление Calpionella alpina , совпадающее с основанием одноименной подзоны Alpina. В частности, в качестве определения основания мела предложено [ 17 ] Рабочее определение границы часто рассматривалось как первое появление аммонита Strambergella jacobi , ранее отнесенного к роду Berriasella , но его использование в качестве стратиграфического индикатора подвергалось сомнению, поскольку его первое появление не коррелирует с появлением C. альпина . [ 18 ]
Месторождения полезных ископаемых и углеводородов
[ редактировать ]Киммериджская глина и ее эквиваленты являются основной нефтематеринской породой нефти Северного моря . [ 19 ] Аравийский внутришельфовый бассейн, образовавшийся в средней и поздней юре, является местом расположения крупнейших в мире запасов нефти, включая месторождение Гавар , крупнейшее в мире нефтяное месторождение. [ 20 ] Саргелу юрского периода [ 21 ] и формации Наокелекан [ 22 ] являются основными нефтематеринскими породами в Ираке . Запасы юрского угля составляют более 1500 гигатонн на северо-западе Китая, главным образом в бассейнах Турфан-Хами и в бассейне Ордос . [ 23 ]
Ударные конструкции
[ редактировать ]Основные ударные структуры включают ударную структуру Мороквенг , ударную структуру диаметром 70 км, погребенную под пустыней Калахари на севере Южной Африки. Удар датируется титоном, примерно 146,06 ± 0,16 млн лет назад. [ 24 ] Еще одно крупное сооружение — кратер Пучеж-Катунки диаметром 40 километров, погребенный под Нижегородской областью на западе России. Воздействие датировано синемуром - 195,9 ± 1,0 млн лет назад. [ 25 ]
Палеогеография и тектоника
[ редактировать ]
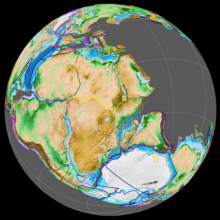
В начале юрского периода все основные массивы суши в мире были объединены в суперконтинент Пангея , который в раннем юрском периоде начал распадаться на северный суперконтинент Лавразию и южный суперконтинент Гондвану . [ 26 ] Рифтинг между Северной Америкой и Африкой был первым, начавшимся в ранней юре и связанным с возникновением Центрально -Атлантической магматической провинции . [ 27 ]
В юрский период Северная Атлантика оставалась относительно узкой, а Южная Атлантика открывалась только в меловой период. [ 28 ] [ 27 ] Континенты были окружены Панталассой , а океан Тетис между Гондваной и Азией находился . В конце триаса в Европе произошла морская трансгрессия , затопившая большую часть центральной и западной Европы, превратившая ее в архипелаг островов, окруженный мелководными морями. [ 29 ] В юрский период и Северный, и Южный полюс были покрыты океанами. [ 26 ] Начиная с ранней юры, Бореальный океан был связан с протоатлантикой «коридором викингов» или Трансконтинентальным Лавразийским морским путем — проходом между Балтийским щитом и Гренландией шириной в несколько сотен километров. [ 30 ] [ 31 ] [ 32 ] В келловейском периоде образовалось Тургайское эпиконтинентальное море , создавшее морской барьер между Европой и Азией. [ 33 ]
Мадагаскар и Антарктида начали отделяться от Африки в конце ранней юры в связи с извержением крупных магматических провинций Кару-Феррар , открыв западную часть Индийского океана и положив начало фрагментации Гондваны. [ 34 ] [ 35 ] В начале юрского периода Северная и Южная Америка оставались соединенными, но к началу поздней юры они разделились, образовав Карибский морской путь, также известный как Испаноязычный коридор, который соединял северную часть Атлантического океана с восточной частью Панталассы. Палеонтологические данные позволяют предположить, что морской путь был открыт еще в ранней юре. [ 36 ]
В рамках неваданской складчатости , которая началась в триасовый период, океан Кэш-Крик закрылся, и различные террейны , включая большой террейн Врангелия, аккрецировались на западной окраине Северной Америки. [ 37 ] [ 38 ] К средней юре произошло столкновение Сибирской плиты и Северо-Китайско-Амурского блока, что привело к закрытию Монголо -Охотского океана . [ 39 ]
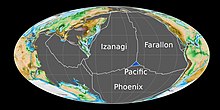
, около 190 миллионов лет назад, Тихоокеанская плита возникла на тройном стыке Фараллон В раннем юрском периоде , Феникс и Идзанаги тектонических плит , трёх основных океанических плит Панталасса. Ранее стабильное тройное соединение превратилось в нестабильное устройство, окруженное со всех сторон трансформными разломами из-за излома одной из границ плит, что привело к образованию Тихоокеанской плиты в центре соединения. [ 40 ] В период от среднего до начала позднего юрского периода Сандэнсский морской путь , мелкое эпиконтинентальное море , покрывал большую часть северо-запада Северной Америки. [ 41 ]

По оценкам, эвстатический уровень моря был близок к современному уровню в геттанге и синемуре, поднимаясь на несколько десятков метров в конце синемура – плинсбаха, а затем снизился до уровня, близкого к современному, к позднему плинсбаху. Судя по всему, в тоаре произошел постепенный подъем до пика примерно 75 м над нынешним уровнем моря. В поздней части тоара уровень моря снова упал на несколько десятков метров. Он постепенно повышался, начиная с аалена, за исключением провалов в несколько десятков метров в байосе и вокруг границы келловея и оксфорда, достигая, возможно, высоты 140 метров над современным уровнем моря на границе кимериджа и титона. Уровень моря падает в позднем титоне, возможно, примерно до 100 метров, а затем снова поднимается примерно до 110 метров на границе титона и берриаса.
Уровень моря в рамках многолетних трендов в юрском периоде имел циклический характер и имел 64 колебания, 15 из которых превышали 75 метров. Наиболее заметна цикличность в юрских породах четвертого порядка с периодичностью около 410 000 лет. [ 42 ]
В раннем юрском периоде химический состав мирового океана перешел от арагонитового моря к кальцитовому , что способствовало растворению арагонита и осаждению кальцита . [ 43 ] Рост количества известкового планктона в средней юре глубоко изменил химию океана, при этом отложение биоминерализованного планктона на дне океана служило буфером против крупных выбросов CO 2 . [ 44 ]
Климат
[ редактировать ]Климат юрского периода в целом был теплее нынешнего, примерно на 5–10 °C, а содержание углекислого газа в атмосфере, вероятно, было примерно в четыре раза выше. Известно, что в этот период времени происходили периодические периоды «похолоданий», однако они нарушали теплый парниковый климат. [ 45 ] Леса, вероятно, росли вблизи полюсов, где было теплое лето и холодная, иногда снежная зима; ледниковые щиты вряд ли существовали, учитывая высокие летние температуры, которые препятствовали накоплению снега, хотя горные ледники могли существовать. [ 46 ] Дропстоуны и глендониты на северо-востоке Сибири в период от ранней до средней юры указывают на холодные зимы. [ 47 ] Глубины океана, вероятно, были на 8 °C теплее, чем сейчас, а коралловые рифы выросли на 10 ° широты дальше к северу и югу. Зона внутритропической конвергенции, вероятно, существовала над океанами, в результате чего возникли большие площади пустынь и кустарников в нижних широтах между 40 ° северной и южной широты от экватора. Биомы тропических лесов и тундры , вероятно, были редки или отсутствовали. [ 46 ] В юрском периоде также наблюдался упадок пангейского мегамуссона, который характеризовал предшествующие пермский и триасовый периоды. [ 48 ] Изменение частоты лесных пожаров в юре определялось 405-тысячелетним циклом эксцентриситета . [ 49 ] Благодаря распаду Пангеи гидрологический цикл в юрском периоде значительно усилился. [ 50 ]
Начало юры, вероятно, ознаменовалось термальным скачком, соответствующим триас-юрскому вымиранию и извержению Центрально-Атлантической магматической провинции. Первая часть юрского периода ознаменовалась раннеюрским похолоданием между 199 и 183 миллионами лет назад. [ 47 ] Было высказано предположение, что оледенение присутствовало в Северном полушарии как в раннем плинсбахском периоде, так и в раннем плинсбахском периоде. [ 51 ] и последний плинсбахский. [ 52 ] [ 53 ] В начале тоара произошел скачок глобальной температуры примерно на 4–8 °C, соответствующий тоарскому океаническому бескислородному событию и извержению крупных магматических провинций Кару-Феррар на юге Гондваны, с теплым интервалом, простирающимся до Конец тоара около 174 миллионов лет назад. [ 47 ] Во время тоарского теплого периода температура поверхности океана, вероятно, превышала 30 °C, а экваториальные и субтропические (30 ° с.ш.–30 ° ю.ш.) регионы, вероятно, были чрезвычайно засушливыми, а температура внутри Пангеи, вероятно, превышала 40 °. С. За тоаровым теплым интервалом следует среднеюрский прохладный интервал (MJCI) между 174 и 164 миллионами лет назад. [ 47 ] которые, возможно, прерывались короткими, эфемерными интервалами в леднике. [ 54 ] [ 55 ] В ааленском периоде климатические изменения, вызванные прецессией, определяли масштабы и частоту лесных пожаров на торфяниках. [ 56 ] Европейский климат, по-видимому, стал заметно более влажным на границе аалена и байоса, но затем стал более засушливым в среднем байосе. [ 57 ] Переходный ледниковый период, возможно, произошел в конце байоса. [ 58 ] На границе келловея и оксфорда в конце MJCI наблюдалось особенно заметное глобальное похолодание. [ 59 ] [ 60 ] потенциально даже ледниковый период. [ 61 ] За этим следует Кимериджский теплый интервал (KWI) между 164 и 150 миллионами лет назад. [ 47 ] Судя по распространению ископаемой древесины , это был один из самых влажных интервалов юры. [ 62 ] Во внутренних районах Пангеи сезонные колебания были менее резкими, чем в предыдущие теплые периоды, поскольку расширение Центральной Атлантики и западной части Индийского океана предоставило новые источники влаги. [ 47 ] Заметное падение температуры произошло во время титона, известного как событие раннего титона похолодания (ETCE). [ 60 ] Конец юрского периода ознаменовался титонско-раннебарремским интервалом похолодания (TBCI), начавшимся 150 миллионов лет назад и продолжавшимся до раннего мела . [ 47 ]
Климатические события
[ редактировать ]Тоарское океаническое бескислородное событие
[ редактировать ]Тоарское океаническое аноксическое событие (TOAE), также известное как событие Дженкинса, было эпизодом широко распространенной океанической аноксии в начале тоарской эпохи, ок. 183 Мя. Он отмечен во всем мире зарегистрированным высокоамплитудным отрицательным выбросом изотопов углерода . [ 63 ] [ 64 ] а также отложения черных сланцев [ 65 ] а также вымирание и разрушение морских организмов, производящих карбонаты, что связано со значительным повышением глобальной температуры. [ 66 ]
TOAE часто связывают с извержением крупных магматических провинций Кару-Феррар и связанным с этим увеличением концентрации углекислого газа в атмосфере, а также с возможным связанным с этим выбросом клатратов метана . [ 66 ] Это, вероятно, ускорило гидрологический цикл и усилило выветривание силикатов , о чем свидетельствует увеличение количества органического вещества земного происхождения, обнаруженного в морских отложениях во время ТОАЭ. [ 67 ] Затронутые группы включают аммониты, [ 68 ] остракоды , [ 65 ] [ 69 ] фораминиферы , [ 70 ] [ 71 ] двустворчатые моллюски , [ 65 ] книдарии , и особенно брахиоподы , [ 72 ] [ 73 ] [ 74 ] для которых TOAE представляло собой одно из самых серьезных вымираний в их эволюционной истории. [ 75 ] Хотя это событие оказало значительное воздействие на морских беспозвоночных, оно оказало незначительное влияние на морских рептилий. [ 76 ] Во время TOAE бассейн Сычуань был преобразован в гигантское озеро , вероятно, в три раза превышающее по размеру современное озеро Верхнее , представленное пачкой Дааньчжай формации Цзылюцзин . Вероятно, во время этого события озеро изолировало около 460 гигатонн (Гт) органического углерода и около 1200 Гт неорганического углерода. [ 77 ] морской воды pH , который уже существенно снизился до события, немного увеличился на ранних стадиях TOAE, а затем упал до самой низкой точки примерно в середине события. [ 78 ] Это закисление океана является вероятной причиной прекращения производства карбонатов. [ 79 ] [ 80 ] Кроме того, бескислородные условия усугублялись усилением рециркуляции фосфора обратно в океанскую воду в результате высокой кислотности океана и температуры, препятствующих его минерализации в апатит; обилие фосфора в морской среде вызвало дальнейшую эвтрофикацию и, как следствие, аноксию в петле положительной обратной связи. [ 81 ]
Конец-юрский переход
[ редактировать ]Переход в конец юрского периода первоначально считался одним из восьми массовых вымираний, но теперь считается сложным интервалом круговорота фауны с увеличением разнообразия одних групп и сокращением других, хотя доказательства этого в основном европейские, вероятно, контролируется изменениями эвстатического уровня моря. [ 82 ]
Флора
[ редактировать ]Конец триасового вымирания
[ редактировать ]Данных о массовом вымирании растений на границе триаса и юры нет. [ 83 ] На границе триаса и юры в Гренландии запись спороморфов (пыльцы и спор) предполагает полный круговорот флоры. [ 84 ] Анализ макроископаемых цветочных сообществ в Европе позволяет предположить, что изменения произошли главным образом из-за местной экологической сукцессии . [ 85 ] В конце триаса Peltaspermaceae вымерли в большинстве частей мира, а Lepidopteris сохранились до ранней юры в Патагонии. [ 86 ] Дикроидий , користоспермовый семенной папоротник, который был доминирующей частью цветочных сообществ Гондваны в триасовый период, также пришел в упадок на границе триаса и юры, сохранившись как реликт в Антарктиде до ранней юры. [ 87 ]
Цветочная композиция
[ редактировать ]Хвойные деревья
[ редактировать ]
Хвойные породы составляли доминирующий компонент юрских флор. Поздний триас и юрский период были основным периодом диверсификации хвойных пород, при этом большинство современных групп хвойных деревьев появились в летописи окаменелостей к концу юрского периода и произошли от вольтзиалейных предков. [ 88 ] [ 89 ]
Первые однозначные находки араукарианских хвойных появились в ранней юре, а представители современного рода Araucaria были широко распространены в обоих полушариях к средней юре. [ 89 ] [ 90 ] [ 91 ]
В юрском периоде также в изобилии обитало вымершее семейство Cheirolepidiaceae , часто узнаваемое по весьма характерной пыльце Classopolis . К юрским представителям относятся пыльцевая шишка Classostrobus и семенная шишка Pararaucaria . Хвойные араукарианские и хейролепидиевые часто встречаются вместе. [ 92 ]
Самая старая окончательная находка семейства кипарисовых ( Cupressaceae ) - это Austrohamia minuta из ранней юры (плинсбах) Патагонии, известная по многим частям растения. [ 93 ] Репродуктивные структуры Austrohamia имеют большое сходство с таковыми у примитивных живых кипарисов родов Taiwania и Cunninghamia . К средней и поздней юре Cupressaceae были многочисленны в теплых умеренно-тропических регионах Северного полушария, наиболее обильно представлены родом Elatides . [ 94 ] В юрском периоде также появились первые виды некоторых современных родов кипарисов, таких как Секвойя . [ 95 ]
Представители вымершего рода Schizolepidopsis, которые, вероятно, представляют собой стволовую группу семейства сосновых ( Pinaceae ), были широко распространены по Евразии в юрском периоде. [ 96 ] [ 97 ] Самая старая однозначная находка Pinaceae - это сосновая шишка Eathiestrobus , известная из поздней юры (кимериджа) Шотландии, которая остается единственной известной однозначной окаменелостью группы до мелового периода. [ 98 ] является самым ранним известным представителем семейства Pinaceae, Несмотря на то, что Eathiestrobus он, по-видимому, является членом пиноидной клады этого семейства, что позволяет предположить, что первоначальная диверсификация Pinaceae произошла раньше, чем было обнаружено в летописи окаменелостей. [ 99 ] [ 89 ]
Самая ранняя находка семейства тисовых ( Taxaceae ) - это Palaeotaxus rediviva из геттанга Швеции, который, как предполагается, тесно связан с ныне живущим Austrotaxus , а Marskea jurassica из средней юры Йоркшира, Англия, и материал из келловейско-оксфордского слоя Даохугоу. в Китае считаются тесно связанными с Amentotaxus , причем последний материал отнесен к современному роду, что указывает на то, что Taxaceae существенно диверсифицировались к концу юрского периода. [ 100 ]
Самые старые однозначные представители Podocarpaceae известны из юрского периода, встречаются в обоих полушариях, включая Scarburgia и Harrisiocarpus из средней юры Англии, а также безымянные виды из средней-поздней юры Патагонии. [ 101 ]
В ранней юре во флоре средних широт Восточной Азии доминировал вымерший листопадный широколиственный хвойный Podozamites , который, по-видимому, не имеет близкого родства ни с одним из ныне живущих семейств хвойных. Его ареал простирался на север до полярных широт Сибири, а затем в средней и поздней юре сузился к северу, что соответствует увеличению засушливости региона. [ 102 ]
Гинкгоалы
[ редактировать ]
Ginkgoales , единственным живым видом которых является Ginkgo biloba , были более разнообразны в юрский период: они были одними из наиболее важных компонентов евразийских юрских флор и были адаптированы к самым разнообразным климатическим условиям. [ 103 ] Древнейшие представители рода Ginkgo , представленные семязачаточными и пыльцевыми органами, сходными с таковыми у современных видов, известны из средней юры в Северном полушарии. [ 103 ] Несколько других линий гинкговых известны из юрских пород, включая Yimaia , Grenana , Nagrenia и Karkenia . Эти линии связаны с гинкгоподобными листьями, но отличаются от живых и ископаемых представителей гинкго наличием по-разному устроенных репродуктивных структур. [ 103 ] [ 104 ] Умальтолепис из юрского периода Азии имеет ремневидные листья, похожие на гинкго, с весьма отчетливыми репродуктивными структурами, сходными со структурами семенных папоротников пельтасперма и користосперма; предполагается, что он принадлежит к Ginkgoales sensu lato. [ 105 ]
Беннеттиталес
[ редактировать ]
Bennettitales , впервые получившие широкое распространение в предшествующий триасовый период, были разнообразными и многочисленными представителями юрских флор обоих полушарий. [ 106 ] Листва Bennettitales имеет сильное сходство с листвой саговников до такой степени, что их невозможно надежно отличить только на основе морфологии. Листья Bennettitales можно отличить от листьев саговников по различному расположению устьиц , и эти две группы не считаются тесно связанными. [ 107 ] Юрские Bennettitales преимущественно относятся к группе Williamsoniaceae . [ 106 ] которые росли в виде кустарников и небольших деревьев. Считается, что Williamsoniaceae имели разветвленную структуру ветвления, аналогичную таковой у современных Banksia , и адаптировались к росту в открытых средах обитания с плохими питательными условиями почвы. [ 108 ] Bennettitales обладают сложными репродуктивными структурами, напоминающими цветы , некоторые из которых, как полагают, были опылены насекомыми. Несколько групп насекомых с длинными хоботками, включая вымершие семейства, такие как каллиграмматидные златоглазки. [ 109 ] и современные виды, такие как мухи -акроцериды , [ 110 ] Предполагается, что они были опылителями беннеттиталовых и питались нектаром , вырабатываемым шишками беннеттиталов.
Саговники
[ редактировать ]Саговники достигли своего апогея разнообразия в юрский и меловой периоды. [ 111 ] Несмотря на то, что мезозойскую эру иногда называют «эпохой саговников», считается, что саговники были относительно второстепенным компонентом среднемезозойских флор, при этом преобладали Bennettitales и Nilssoniales , имеющие саговниковую листву. [ 112 ] Nilssoniales часто считали саговниками или родственниками саговников, но было обнаружено, что они отличаются по химическому составу и, возможно, более тесно связаны с Bennettitales. [ 113 ] Отношения большинства мезозойских саговников с живыми группами неоднозначны. [ 112 ] при этом юрские саговники не принадлежат ни к одной из двух современных групп саговников, хотя некоторые юрские саговники, возможно, представляют собой родственников стеблевой группы современных Cycadaceae , таких как листовой род Paracycas, известный в Европе, и Zamiaceae , как некоторые европейские виды листового рода Pseudoctenis . В юрском периоде также была широко распространена вымершая линия Ctenis , которая, по-видимому, была отдаленно связана с современными саговниками. [ 114 ] Современные саговники опыляются жуками, и считается, что такая ассоциация образовалась в ранней юре. [ 111 ]
Другие семенные растения
[ редактировать ]Хотя было несколько заявленных записей, не существует широко признанных летописей окаменелостей юрского периода цветковых растений , которые составляют 90% ныне живущих видов растений, и ископаемые данные свидетельствуют о том, что группа диверсифицировалась в течение следующего мелового периода. [ 115 ]
Самые ранние известные гнетофиты , одна из четырех основных живых групп голосеменных растений , появились к концу юрского периода, при этом самым старым однозначно гнетофитом является семя Dayvaultia из поздней юры Северной Америки. [ 116 ]

«Семенные папоротники» ( Pteridospermatophyta ) — собирательный термин, обозначающий разрозненные линии папоротниковоподобных растений, которые производят семена, но имеют неопределенное сходство с живыми группами семенных растений . Выдающейся группой юрских семенных папоротников являются Caytoniales , которые достигли своего апогея в юрском периоде, с широко распространенными встречами в Северном полушарии, хотя записи в Южном полушарии остаются редкими. Из-за их ягодообразных семенных коробочек часто предполагалось, что они были близкими родственниками или, возможно, предками цветковых растений, но доказательства этого неубедительны. [ 117 ] Семенные папоротники, относящиеся к користосперму , такие как Pachypteris и Komlopteris, были широко распространены в обоих полушариях в юрском периоде. [ 118 ]
Czekanowskiales , также известные как Leptostrobales, представляют собой группу семенных растений неопределенного родства с устойчивыми сильно рассеченными листьями, растущими на лиственных коротких побегах, покрытых чешуйчатыми листьями, известных из позднего триаса (возможно, поздней перми). [ 119 ] ) до мелового периода. [ 120 ] Считается, что они имели древесную или кустарниковую форму и составляли заметный компонент мезозойской умеренной и теплоумеренной флоры Северного полушария. [ 119 ] Род Phoenicopsis был широко распространён в раннесреднеюрских флорах Восточной Азии и Сибири. [ 121 ]
Pentoxylales семенных , небольшая, но явно обособленная группа лианоподобных растений неясного родства, впервые появилась в юрском периоде. Их распространение, по-видимому, ограничивалось Восточной Гондваной. [ 122 ]
Папоротники и союзники
[ редактировать ]К современным семействам папоротников, широко распространенным в юрском периоде, относятся Dipteridaceae , Matoniaceae , Gleicheniaceae , Osmundaceae и Marattiaceae . [ 123 ] [ 124 ] Polypodiales , которые составляют 80% разнообразия современных папоротников, не имеют упоминаний о юрском периоде и, как полагают, разнообразны в меловом периоде. [ 125 ] хотя широко распространенный юрский травянистых род папоротников Coniopteris , исторически интерпретируемый как близкий родственник древовидных папоротников семейства Dicksoniaceae , недавно был интерпретирован как ранний родственник этой группы. [ 126 ]
Cytheales , группа , содержащая большинство современных древовидных папоротников, появилась в поздней юре и представлена представителями рода Cyathocaulis предположительно являются ранними представителями Cyatheaceae . , которые на основании кладистического анализа [ 127 ] Существует лишь несколько возможных находок Hymenophyllaceae из юрского периода, включая Hymenophyllites macrosporangiatus из российской юры. [ 128 ]
Древнейшие остатки современных хвощей рода Equisetum впервые появляются в ранней юре и представлены Equisetum dimorphum из ранней юры Патагонии. [ 129 ] и Equisetum Laterale из ранней и средней юры Австралии. [ 130 ] [ 131 ] Окремненные остатки Equisetum Thermale из поздней юры Аргентины обладают всеми морфологическими признаками современных представителей рода. [ 132 ] По оценкам, раскол между Equisetum bogotense и всеми остальными ныне живущими Equisetum произошел не позднее ранней юры. [ 131 ]
Низшие растения
[ редактировать ]Иглы, практически идентичные современным видам, известны начиная с юрского периода. Isoetites rolandii из средней юры штата Орегон — самый ранний известный вид, представляющий все основные морфологические особенности современных Isoetes . Более примитивные формы, такие как Nathorstiana , сохраняющие удлиненный стебель, сохранились до раннего мела. [ 133 ]
Считается, что мох Кулиндобрюм из средней юры России, который был обнаружен в костях динозавров, связан с Splachnaceae , которые растут на тушах животных. [ 134 ] Считается, что Bryokhutuliinia из того же региона связана с Dicranales . [ 134 ] Heinrichsiella Считается, что из юрского периода Патагонии принадлежит либо к Polytrichaceae , либо к Timmiellaceae . [ 135 ]
Печеночник Pellites hamiensis из среднеюрской формации Сишаньяо в Китае является старейшей находкой семейства Pelliaceae . [ 136 ] Pallaviciniites sandaolingensis Считается, что из того же месторождения принадлежит к подклассу Pallaviciniineae в составе Pallaviciniales . [ 137 ] Ricciopsis sandaolingensis , также из того же месторождения, является единственной юрской находкой Ricciaceae . [ 138 ]
фауна
[ редактировать ]Рептилии
[ редактировать ]Крокодиломорфы
[ редактировать ]
Триасово-юрское вымирание уничтожило разнообразие псевдозухий , при этом крокодиломорфы , возникшие в начале позднего триаса, оказались единственной группой псевдозухий, которая выжила. Все остальные псевдозухии, включая травоядных этозавров и плотоядных « раузухий », вымерли. [ 139 ] Морфологическое разнообразие крокодиломорфов в раннем юрском периоде было примерно таким же, как и у позднетриасовых псевдозухий, но они занимали разные области морфопространства, что позволяет предположить, что они занимали разные экологические ниши по сравнению со своими триасовыми собратьями и что имела место обширная и быстрая радиация крокодиломорфов. в течение этого интервала. [ 140 ] В то время как современные крокодилы в основном ведут образ жизни хищников, устраивающих водные засады, юрские крокодиломорфы демонстрировали широкое разнообразие жизненных привычек. Безымянный протозухид, известный по зубам ранней юры Аризоны, представляет собой самого раннего известного травоядного крокодиломорфа, адаптацию, которая появлялась несколько раз в мезозое. [ 141 ]
Thalattosuria . , клада преимущественно морских крокодиломорфов, впервые появилась в ранней юре и стала заметной частью морских экосистем [ 142 ] Внутри Thalattosuria Metriorhynchidae стали высоко адаптированными к жизни в открытом океане, включая преобразование конечностей в ласты, развитие хвостового плавника и гладкую кожу без чешуи. [ 143 ] Морфологическое разнообразие крокодиломорфов в ранней и средней юре было относительно низким по сравнению с таковым в более поздние периоды и доминировали наземные мелкотелые, длинноногие сфенозухи , ранние крокодилиформы и талаттозухи. [ 144 ] [ 142 ] Неозуха . , основная группа крокодиломорфов, впервые появилась в период от ранней до средней юры Неозуха представляет собой переход от наземного образа жизни предков к пресноводной водной экологии, подобной той, которую занимают современные крокодилы. [ 145 ] Время возникновения неозуха является спорным. Предполагается, что самой старой находкой неозухов является из Calsoyasuchus ранней юры Аризоны, который во многих анализах был обнаружен как самый ранний ветвящийся член неозухийского семейства Goniopholididae , что радикально меняет времена диверсификации крокодиломорфов. Однако это размещение оспаривается: некоторые анализы обнаружили его за пределами Неозухии, что указывает на то, что самые старые записи Неозухии относятся к средней юре. [ 145 ] Было высказано предположение, что разанандргобе из средней юры Мадагаскара представляет собой старейшую находку Notosurcia , преимущественно гондванской клады, состоящей в основном из наземных крокодиломорфов, также известных из мелового и кайнозойского периода. [ 146 ]
Черепахи
[ редактировать ]
стволовой группы Черепахи ( Testudinata ) разнообразовались в юрском периоде. Юрские стволовые черепахи принадлежат к двум все более продвинутым кладам: Mesochelydia и Perichelydia . [ 147 ] Считается, что предковым состоянием мезохелидий является водный, в отличие от наземного состояния семенников. [ 148 ] Две современные группы черепах ( Testudines ), Pleurodira и Cryptodira , разошлись к началу поздней юры. [ 147 ] Древнейшие известные плевродиры, Platychelyidae , известны из поздней юры Европы и Америки. [ 149 ] в то время как самый старый однозначный криптодир, Sinaspideretes , ранний родственник мягкопанцирных черепах , известен из поздней юры Китая. [ 150 ] Thalassochelydia морским , разнообразная линия морских черепах, не родственная современным черепахам , известна из поздней юры Европы и Южной Америки. [ 151 ]
Лепидозавры
[ редактировать ]Ринхоцефалы (единственный живой представитель — туатара ) достигли глобального распространения к началу юрского периода. [ 152 ] и представлял собой доминирующую группу мелких рептилий в юрском периоде во всем мире. [ 153 ] Ринхоцефалы достигли своего наивысшего морфологического разнообразия в своей эволюционной истории в юрском периоде, занимая широкий диапазон образов жизни, включая водных плеврозавров с длинными змееподобными телами и редуцированными конечностями, специализированных травоядных эйленодонтинов , а также сафеозавров, имевших широкие зубные пластины. указывает на дурофагию . [ 154 ] Ринхоцефалы исчезли из Азии после ранней юры. [ 152 ] По оценкам, последний общий предок ныне живущих чешуйчатых (в том числе ящериц и змей ) жил около 190 миллионов лет назад в ранней юре, при этом основные расхождения между современными линиями чешуйчатых, по оценкам, произошли в период от ранней до средней юры. [ 155 ] Чешуйчатые впервые появляются в летописи окаменелостей в средней юре. [ 156 ] включая представителей современных клад, таких как Scincomorpha , [ 157 ] хотя многие чешуйки юрского периода имеют неясные отношения с живыми группами. [ 158 ] эйхштеттизавр Предполагается, что из поздней юры Германии является ранним родственником гекконов и демонстрирует приспособления к лазанию. [ 159 ] Дорсетизавр из поздней юры Северной Америки и Европы представляет собой старейшую широко признанную находку Anguimorpha . [ 160 ] Было высказано предположение, что Marmoretta из средней юры Британии представляет собой поздно выживший лепидозавроморф, не являющийся как Rhynchocephalia, так и Squamata, хотя некоторые исследования обнаружили его как стеблево-чешуйчатый. [ 161 ]
-
 Vadasaurus herzogi , ринхоцефал из верхнеюрского известняка Зольнхофен в Германии.
Vadasaurus herzogi , ринхоцефал из верхнеюрского известняка Зольнхофен в Германии. -
 Homeosaurus maximiliani , ринхоцефал из известняка Зольнхофен.
Homeosaurus maximiliani , ринхоцефал из известняка Зольнхофен. -
 Плеврозавр , водный ринхоцефал из поздней юры Европы.
Плеврозавр , водный ринхоцефал из поздней юры Европы. -
 Eichstaettisaurus schroederi , вымершая ящерица из известняка Зольнхофен.
Eichstaettisaurus schroederi , вымершая ящерица из известняка Зольнхофен.




Хористодеры
[ редактировать ]
Самые ранние известные останки Choristodera , группы пресноводных водных рептилий с неопределенным родством с другими группами рептилий, обнаружены в средней юре. Из юры известны только два рода хористодер. Один из них — небольшой ящерицеобразный Cteniogenys , который считается самым базальным из известных хористодер; он известен от средней до поздней юры Европы и поздней юры Северной Америки, аналогичные остатки известны также из верхней средней юры Кыргызстана и Западной Сибири. [ 162 ] Другой - Coeruleodraco из поздней юры Китая, более развитый хористодер, хотя все еще небольшой и по морфологии напоминающий ящерицу. [ 163 ]
Ихтиозавры
[ редактировать ]
Ихтиозавры столкнулись с эволюционным затруднением во время вымирания в конце триаса, когда все ненеоихтиозавры вымерли . Ихтиозавры достигли своего пика видового разнообразия в раннем юрском периоде, имея множество морфологических форм, включая огромного хищника Temnodontosaurus и похожего на рыбу-меч Eurhinosaurus , хотя раннеюрские ихтиозавры были значительно менее морфологически разнообразны, чем их триасовые собратья. [ 164 ] [ 165 ] На границе ранней и средней юры, между концом тоара и началом байоса, большинство линий ихтиозавров, по-видимому, вымерло с первым появлением Ophthalmosauridae , клады, которая с тех пор будет охватывать почти всех ихтиозавров. , в раннем байосе. [ 166 ] Офтальмозавриды были разнообразны в поздней юре, но не смогли заполнить многие ниши, которые были заняты ихтиозаврами в ранней юре. [ 166 ] [ 164 ] [ 165 ]
Плезиозавры
[ редактировать ]
Плезиозавры возникли в конце триаса (ретийского периода). К концу триаса все остальные зауроптериги , включая плакодонтов и нотозавров , вымерли. По крайней мере шесть линий плезиозавров пересекли границу триаса и юры. [ 167 ] Плезиозавры уже были разнообразны в самой ранней юре, при этом большинство плезиозавров в голубых лиях геттангового возраста принадлежало к Rhomaleosauridae . Ранние плезиозавры, как правило, имели небольшое тело, а к тоарскому периоду размер тела увеличивался. [ 168 ] По-видимому, вокруг границы ранней и средней юры произошел сильный сдвиг: микроклеидиды и ромалеозавриды вымерли и почти вымерли соответственно после конца тоара с первым появлением доминирующей клады плезиозавров во второй половине юры. Cryptoclididae . в байосе [ 166 ] В средней юре произошла эволюция короткошеих и большеголовых талассофонических плиозавров от предков с маленькой головой и длинной шеей . [ 169 ] [ 166 ] Некоторые талассофоновые плиозавры, такие как некоторые виды плиозавров , имели черепа длиной до двух метров и длину тела около 10–12 метров, что делало их высшими хищниками океанов поздней юры. [ 170 ] [ 166 ] Плезиозавры вторглись в пресноводную среду в юрском периоде, при этом неопределенные останки плейозавров с мелким телом были известны из пресноводных отложений юрского периода Китая и Австралии. [ 171 ] [ 172 ]
Птерозавры
[ редактировать ]
Птерозавры впервые появились в позднем триасе. Основная группа юрских птерозавров — это Rhamphorhynchidae , которые впервые появились в конце ранней юры (тоар); [ 173 ] они считаются рыбоядными . [ 174 ] Анурогнатиды , впервые появившиеся в средней юре, обладали короткими головами и телом с густым мехом и, как полагают, были насекомоядными. [ 174 ] Производные монофенестратановые птерозавры, такие как вуконгоптериды, появились в конце средней юры. Передовые короткохвостые птеродактилоиды впервые появились на границе средней и поздней юры. К юрским птеродактилоидам относятся гребневики , подобные Ctenochasma , которые имеют близко расположенные игольчатые зубы, которые предположительно использовались для фильтрационного питания . [ 174 ] Причудливый позднеюрский ктенохазматоид Cycnorhamphus имел челюсть с зубами только на кончиках и изогнутыми челюстями, как у современных аистов с открытым клювом, которые, возможно, использовались для удержания и раздавливания твердых беспозвоночных. [ 174 ]
Динозавры
[ редактировать ]Динозавры , морфологически диверсифицированные в позднем триасе, испытали значительное увеличение разнообразия и численности в ранней юре после вымирания в конце триаса и вымирания других групп рептилий, став доминирующими позвоночными в наземных экосистемах. [ 175 ] [ 176 ] Чилезавр , морфологически аберрантный травоядный динозавр из поздней юры Южной Америки, имеет неопределенное отношение к трем основным группам динозавров, поскольку в различных анализах он был обнаружен как член всех трех. [ 177 ]
Тероподы
[ редактировать ]Передовые тероподы, относящиеся к Neotheropoda, впервые появились в позднем триасе. Базальные неотероподы, такие как целофизоиды и дилофозавры , сохранялись до ранней юры, но вымерли к средней юре. [ 178 ] Самые ранние авеространы появились в ранней юре, причем самым ранним известным представителем Ceratosauria был Saltriovenator из раннего синемура (199,3–197,5 миллионов лет назад) в Италии. [ 179 ] Необычный цератозавр Limusaurus из поздней юры Китая питался травоядной пищей: у взрослых особей были беззубые клювовидные челюсти. [ 180 ] что сделало его самым ранним известным тероподом, перешедшим от плотоядной диеты. [ 181 ] Самые ранние представители Tetanurae появились в конце ранней юры или начале средней юры. [ 182 ] Megalosauridae представляют собой старейшую ветвь Tetanurae, впервые появившуюся в Европе в байосском периоде. [ 183 ] самым старым представителем Allosauroidea Предполагается, что является Асфальтовенатор из средней юры Южной Америки. [ 182 ] Целурозавры впервые появились в средней юре, включая ранних тираннозавров, таких как процератозавр из бата Британии. [ 184 ] некоторые целурозавры из поздней юры Китая, включая Shishugounykus и Haplocheirus, Предполагается, что представляют собой ранних альвареззавров . [ 185 ] однако это было подвергнуто сомнению. [ 186 ] Скансориоптеригиды , группа небольших пернатых целурозавров с перепончатыми крыльями, напоминающими летучую мышь, предназначенными для планирования, известны от средней до поздней юры Китая. [ 187 ] Предполагается , что самой старой находкой троодонтид является Hesperornithoides из поздней юры Северной Америки. Останки зубов, предположительно представляющие останки дромеозавров, известны с юрского периода, но останки тел неизвестны до мелового периода. [ 188 ]
-
 Скелет цератозавра , цератозаврид из поздней юры Северной Америки.
Скелет цератозавра , цератозаврид из поздней юры Северной Америки. -
 Скелет монолофозавра , базального столбняка из средней юры Китая
Скелет монолофозавра , базального столбняка из средней юры Китая -
 Реставрация И Ци , скансориоптеригида из средней-поздней юры Китая.
Реставрация И Ци , скансориоптеригида из средней-поздней юры Китая.



Птицы
[ редактировать ]
Самые ранние авиаланы , включающие птиц и их предков, появились в период от средней до поздней юры и окончательно представлены археоптериксами из поздней юры Германии. Авиаланы принадлежат к кладе Paraves внутри Coelurosauria, в которую также входят дромеозавры и троодонтиды. Anchiornithidae из средней - поздней юры Евразии часто предполагают как авиаланы, но также альтернативно обнаруживаются как отдельная линия паравианов. [ 189 ]

Орнитишии
[ редактировать ]Самые ранние окончательные птицетазовые животные появляются в ранней юре и представлены базальными птицетазовыми животными, такими как Lesothosaurus , гетеродонтозавридами и ранними представителями Thyreophora . Самые ранние представители Ankylosauria и Stegosauria появляются в средней юре. [ 190 ] Базальный неорнитиший Kulindadromeus из средней юры России указывает на то, что по крайней мере некоторые птицетазовые были покрыты протоперьями . [ 191 ] Самые ранние представители Ankylopollexia , которые стали заметными в меловом периоде, появились в поздней юре и были представлены двуногими формами, такими как камптозавр . [ 192 ] Цератопсы впервые появились в поздней юре Китая, представленные представителями Chaoyangsauridae . [ 193 ]
Зауроподоморфы
[ редактировать ]
Зауроподы стали доминирующими крупными травоядными животными в наземных экосистемах в юрском периоде. [ 194 ] Некоторые юрские зауроподы достигали гигантских размеров, став крупнейшими организмами, когда-либо жившими на суше. [ 195 ]
Базальные двуногие зауроподоморфы , такие как массоспондилиды , продолжали существовать до ранней юры, но вымерли к началу средней юры. [ 194 ] Четвероногие зауропоморфы появились в позднем триасе. Четвероногие ледумахади из самой ранней юры Южной Африки достигали предполагаемого веса 12 тонн, что намного превышает вес других известных базальных зауроподоморфов. [ 196 ] Гравизаврийские зауроподы впервые появились в раннем юрском периоде, причем самой старой окончательной находкой является вулканодон из Зимбабве, вероятно, синемурийского возраста. [ 197 ] Эузавроподы впервые появились в конце ранней юры (тоар) и разнообразились в средней юре; [ 194 ] в их число входили цетиозавриды , туриазавры , [ 198 ] и маменчизавры . [ 199 ] Неозавроподы, такие как макронарии и диплодокоиды, впервые появились в средней юре, а затем стали многочисленными и глобально распространились в поздней юре. [ 200 ]
Земноводные
[ редактировать ]
Разнообразие темноспондилов постепенно сокращалось в течение позднего триаса, и только брахиопоиды . в юрский период и далее дожили [ 201 ] Представители семейства Brachyopidae известны из юрских отложений Азии. [ 202 ] а чигутисавр Сидеропс известен из ранней юры Австралии. [ 203 ] Современные лисамфибии начали диверсифицироваться в юрском периоде. Считается, что раннеюрский прозалирус представлял собой первого родственника лягушки, морфология которого позволяла прыгать, как живые лягушки. [ 204 ] Морфологически узнаваемые стволовые лягушки, такие как южноамериканский Notobatrachus, известны из средней юры, [ 205 ] современные лягушки кроновой группы, такие как Enneabatrachus и Rhadinosteus, появились в поздней юре. [ 206 ] Хотя самые ранние земноводные линии саламандр известны из триаса, [ 207 ] Саламандры кроновой группы впервые появляются в период от средней до поздней юры в Евразии вместе с родственниками стволовой группы. Считается, что многие саламандры юрской стволовой группы, такие как Marmorerpeton и Kokartus , были неотеническими . [ 208 ] К ранним представителям саламандр кроновой группы относятся Chunerpeton , Pangerpeton и Linglongtriton из средне-позднеюрской биоты Янляо Китая. Они принадлежат к Cryptobranchoidea , в состав которого входят современные азиатские и гигантские саламандры . [ 209 ] Считается, что Beiyanerpeton и Qinglongtriton из одной и той же биоты являются ранними членами Salamandroidea , группы, в которую входят все остальные ныне живущие саламандры. [ 210 ] [ 211 ] Саламандры расселились по Северной Америке к концу юрского периода, о чем свидетельствует Иридотритон , обнаруженный в позднеюрской формации Моррисон . [ 212 ] Стебель -цецилия Eocaecilia известна из ранней юры Аризоны. [ 213 ] Четвертая группа лиссамфибий — вымершие альбанерпетонтиды — впервые появилась в средней юре, представленная Anoualerpeton priscus из бата Британии, а также неопределенными остатками из равновозрастных отложений Франции и формации Аноуал Марокко. [ 214 ]

Млекопитающие
[ редактировать ]Mammaliaformes , включая млекопитающих , произошедших от цинодонтов в конце триаса, широко диверсифицировались в юрском периоде. [ 215 ] Хотя большинство юрских млекопитающих известны исключительно по изолированным фрагментам зубов и челюстей, исключительно сохранившиеся останки свидетельствуют о разнообразии образа жизни. [ 215 ] Докодонтан Castorocauda и был приспособлен к водной жизни, как утконос и выдра . [ 216 ] Некоторые члены Харамииды [ 217 ] и эвтриконодонтное племя Volaticotherini [ 218 ] имели патагий, похожий на таковой у белок-летяг , позволяющий им планировать по воздуху. Муравьедоподобное неопределенной таксономии, вероятно млекопитающее Fruitafossor , было специалистом по колониальным насекомым, как и ныне живущие муравьеды . [ 219 ] Australosphenida — группа млекопитающих, возможно, родственная ныне живущим однопроходным , впервые появилась в средней юре Гондваны. [ 220 ] Самые ранние находки мультитуберкулятов , самых продолжительных и успешных отрядов млекопитающих, известны из средней юры. [ 221 ] Терианские млекопитающие, представленные сегодня живыми плацентами и сумчатыми , имеют свои самые ранние записи в начале поздней юры, представленные Джурамайей , млекопитающим , более близким к предкам плацентарных, чем сумчатые. [ 222 ] Джурамайя гораздо более развит, чем ожидалось для своего возраста, поскольку другие териевые млекопитающие не известны до раннего мела, и было высказано предположение, что Джурамайя также может произойти из раннего мела. [ 223 ] Две группы цинодонтов, не имеющих млекопитающих, сохранились и после конца триаса. Насекомоядные Tritheledontidae имеют несколько находок из ранней юры. Tritylodontidae , травоядная группа цинодонтов , впервые появившаяся в рэтском периоде, имеет многочисленные находки из юрского периода, преимущественно из Северного полушария. [ 224 ] [ 225 ]
Рыба
[ редактировать ]Бесчелюстная рыба
[ редактировать ]
Последний известный вид конодонтов , класса бесчелюстных рыб , чьи твердые, похожие на зубы элементы являются ключевыми окаменелостями, окончательно вымер в самом раннем юрском периоде после более чем 300 миллионов лет эволюционной истории, причем асинхронное вымирание произошло сначала в Тетисе и восточная Панталасса и выжившие, сохранившиеся до самого раннего геттанга Венгрии и центральной Панталассы. [ 226 ] Конодонты конца триаса были представлены лишь несколькими видами, и их численность постепенно сокращалась в среднем и позднем триасе. [ 227 ] Янляомизон из средней юры Китая представляет собой старейшую постпалеозойскую миногу и старейшую миногу, имеющую зубчатый пищевой аппарат и, вероятно, трехстадийный жизненный цикл, типичный для современных представителей группы. [ 228 ]
Саркоптеригии
[ редактировать ]
Двоякодышащие рыбы (Dipnoi) обитали в пресноводных средах обоих полушарий в юрском периоде. [ 229 ] Некоторые исследования предполагают, что последний общий предок всех ныне живущих двоякодышащих рыб жил в юрском периоде. [ 230 ] Мавсонииды , морская и пресноводная/солоноватая группа целакантов , впервые появившаяся в Северной Америке в триасовом периоде, к концу юрского периода распространилась на Европу и Южную Америку. [ 231 ] Морские Latimeriidae , которые содержат живых целакантов рода Latimeria , также присутствовали в юрском периоде, возникнув в триасе, с рядом записей из юрского периода Европы. [ 232 ]
Актиноптеригии
[ редактировать ]
Лучеперые рыбы ( Actinopterygii ) были основными компонентами юрских пресноводных и морских экосистем. Архаичные « палеонисковые » рыбы, которые были распространены как в морских, так и в пресноводных средах обитания в предшествующий триасовый период, сократились в юрском периоде, будучи в значительной степени заменены более производными актиноптеригическими линиями. [ 233 ] Самые старые из известных Acipenseriformes , группа, в которую входят современные осетры и веслоносы , происходят из ранней юры. [ 234 ] Амиеформная рыба (к которой сегодня относится только боуфин ) впервые появилась в ранней юре и представлена Caturus из плинсбахского яруса Британии; после появления в западной части Тетиса к концу юры они распространились на Африку, Северную Америку, Юго-Восточную и Восточную Азию, [ 235 ] с современным семейством Amiidae, появившимся в поздней юре. [ 236 ] Pycnodontiformes , впервые появившиеся в западной части Тетиса в позднем триасе, к концу юрского периода распространились на Южную Америку и Юго-Восточную Азию, имея высокое разнообразие в Европе в позднем юрском периоде. [ 235 ] В юрском периоде гинглимоди , единственные живые представители — гары (Lepisosteidae), были разнообразны как в пресноводной, так и в морской среде. Древнейшие известные представители анатомически современных гаров появились в поздней юре. [ 237 ] стволовой группы Костистые кости , составляющие более 99% современных Actinopterygii, впервые появились в триасе в западной части Тетиса; первые представители современных клад костистых рыб, такие как Elopomorpha и Osteoglossoidei . они претерпели серьезную диверсификацию, начавшуюся в поздней юре, когда в это время появились [ 238 ] [ 239 ] Pachycormiformes Leedsichthys , группа морских стволовых костистых рыб, впервые появилась в ранней юре и включала как тунцеподобные хищные, так и фильтрующие формы; последняя включала в себя крупнейшую костную рыбу, которая, как известно, существовала: , с предполагаемой максимальной длиной более 15 метров, известны от позднего среднего до поздней юры. [ 240 ]
Хондриктиес
[ редактировать ]
В ранней юре акулоподобные гибодонты , которые представляли доминирующую группу хондрихтиев в предшествующий триасовый период, были обычным явлением как в морских, так и в пресноводных условиях; однако к поздней юре гибодонты стали второстепенными компонентами большинства морских сообществ, будучи в значительной степени заменены современными неоселахиями , но оставались обычными в пресноводных и ограниченных морских средах. [ 241 ] [ 242 ] Neoselachii, в котором обитают все живые акулы и скаты, распространился начиная с ранней юры. [ 243 ] Самый старый известный скат ( Batoidea ) — Antiquaobatis из плиенсбаха Германии. [ 244 ] Юрские батоиды, известные по полным останкам, сохраняют консервативную морфологию, напоминающую рыбу- гитару . [ 245 ] Самые старые известные Hexanchiformes и ковровые акулы (Orectolobiformes) происходят из ранней юры (плинсбахского и тоарского периода соответственно) Европы. [ 246 ] [ 247 ] Древнейшие известные представители Heterodontiformes , единственными живыми представителями которых являются бычья акула ( Heterodontus ), впервые появились в ранней юре, а представители живого рода появились в позднеюрском периоде. [ 248 ] Самые старые из известных макрельевых акул (Lamniformes) происходят из средней юры и представлены родом Palaeocarcharias , который имеет тело, напоминающее оректолобиформное, но имеет ключевые сходства в гистологии зубов с ламнообразными, включая отсутствие ортодентина. [ 249 ] Самая старая находка акул-ангелов ( Squatiniformes ) — это Pseudorhina из поздней юры (оксфорд-титон) Европы, которая уже имеет форму тела, сходную с представителями единственного ныне живущего рода отряда Squatina . [ 250 ] Самые старые известные останки Carcharhiniformes , крупнейшего отряда ныне живущих акул, впервые появляются в поздней средней юре (бате) западного Тетиса (Англия и Марокко). Известные останки зубов и исключительно сохранившихся тел юрских Carchariniformes подобны останкам ныне живущих кошачьих акул . [ 251 ] Synechodontiformes , вымершая группа акул, тесно связанных с Neoselachii, также были широко распространены в юрском периоде. [ 252 ] Самые старые останки современных химер относятся к ранней юре Европы, а представители современного семейства Callorhinchidae появились в средней юре. В отличие от большинства современных химер, юрские химеры часто встречаются на мелководье. [ 253 ] Близкородственные Squaloraja и мириакантоиды также известны из юры Европы. [ 254 ]
Насекомые и паукообразные
[ редактировать ]
Судя по всему, на границе триаса и юры не произошло крупного вымирания насекомых. [ 83 ] В юрском периоде Евразии известно множество важных местонахождений окаменелостей насекомых, наиболее важными из которых являются формация Карабастау в Казахстане и различные отложения биоты Янляо во Внутренней Монголии, Китай, такие как пласт Даохугоу, датируемый келловей-оксфордским периодом. Разнообразие насекомых оставалось на прежнем уровне на протяжении всей ранней и средней юры, но во второй трети юры темпы возникновения существенно возросли, в то время как темпы вымирания остались на прежнем уровне. [ 255 ] Увеличение разнообразия насекомых в средней-поздней юре соответствует существенному увеличению разнообразия ротового аппарата насекомых . [ 256 ] Средняя и поздняя юра была временем значительной диверсификации жуков . [ 257 ] Долгоносики впервые появляются в летописи окаменелостей в период от средней до поздней юры, но предположительно они возникли в период от позднего триаса до ранней юры. [ 258 ] Прямокрылые процветали в ранней юре, особенно в Джунгарском бассейне . [ 259 ] Самые древние из известных чешуекрылых (группа бабочек и мотыльков) известны с границы триаса и юры, с чешуйками крыльев, принадлежащими подотряду Glossata и бабочкам класса Micropterigidae из отложений этого возраста в Германии. [ 260 ] Современные представители как стрекоз , так и стрекоз также впервые появились в юрском периоде. [ 261 ] Хотя современные представители не известны до кайнозоя, эктопаразитические насекомые, которые, как полагают, представляют собой примитивных блох , принадлежащих к семейству Pseudopulicidae , известны из средней юры Азии. Эти насекомые существенно отличаются от современных блох, поскольку не имеют специализированной морфологии последних и имеют более крупные размеры. [ 262 ] [ 263 ] Осы-паразитоиды ( Apocrita ) впервые появились в ранней юре и впоследствии получили широкое распространение, изменив наземные пищевые сети. [ 264 ] Юрская пила также стала свидетелем первых выступлений нескольких других групп насекомых, в том числе Phasmatodea (палочников), [ 265 ] Мантофазматиды , [ 266 ] Embioptera (прядильщики паутины), [ 267 ] и Raphidioptera (змеекрылые). [ 268 ]

Из юрского периода известно лишь несколько находок клещей, в том числе Jureremus , панцирный клещ, принадлежащий к семейству Cymbaeremaeidae, известный из поздней юры Великобритании и России. [ 269 ] и представитель ныне живущего рода орбатид Hydrozetes из ранней юры Швеции. [ 270 ] Пауки диверсифицировались в юрском периоде. [ 271 ] Раннеюрский Seppo koponeni может представлять собой стволовую группу Palpimanoidea . [ 272 ] Eoplectreurys из средней юры Китая считается стволовой линией Synspermiata . Самый древний представитель семейства Archaeidae — Patarchaea — известен из средней юры Китая. [ 271 ] Mongolarachne из средней юры Китая — один из крупнейших известных ископаемых пауков, длина ног которого превышает 5 сантиметров. [ 273 ] Единственный скорпион, известный из юрского периода, - это Liassoscorpionides из ранней юры Германии, местонахождение неопределенного происхождения. [ 274 ] Эупнои- сенокосцы ( Opiliones ) известны из средней юры Китая, включая представителей семейства Sclerosomatidae . [ 275 ] [ 276 ]
Морские беспозвоночные
[ редактировать ]Конец триасового вымирания
[ редактировать ]Во время вымирания в конце триаса вымерло 46–72% всех морских родов. Последствия вымирания в конце триаса были наибольшими в тропических широтах и были более серьезными в Панталассе, чем в Тетисе или северных океанах. Экосистемы тропических рифов рухнули во время этого события и полностью восстановились лишь намного позже, в юрском периоде. сидячие фильтраторы и фотосимбиотические организмы. Наиболее сильно пострадали [ 277 ]
Морские экосистемы
[ редактировать ]Уменьшившись на границе триаса и юры, рифы существенно расширились в течение поздней юры, включая как губковые рифы , так и склерактиновые коралловые рифы . Позднеюрские рифы по форме были похожи на современные рифы, но имели больше микробных карбонатов и гиперкальцинированных губок , а также имели слабую биогенную связь. В конце юрского периода численность рифов резко сократилась. [ 278 ] что вызвало связанное с этим снижение разнообразия десятиногих ракообразных. [ 279 ] Самые ранние планктонные фораминиферы, составляющие подотряд Globigerinina , известны из поздней ранней юры (середины тоара) западного Тетиса, распространились по всему Тетису к средней юре и стали глобально распространены в тропических широтах к поздней юре. [ 280 ] Кокколитофоры и динофлагелляты , впервые появившиеся в триасе, распространились в период от ранней до средней юры, став видными представителями фитопланктона . [ 281 ] Трубчатые черви Microconchid , последний оставшийся отряд Tentaculita , группы животных неопределенного родства, которые сходились с трубчатыми червями Spirorbis , были редки после триаса и сведены к единственному роду Punctaconchus , который вымер в позднем бате. [ 282 ] Самая старая известная диатомовая водоросль происходит из янтаря позднеюрского периода из Таиланда, отнесенного к современному роду Hemiaulus . [ 283 ]
Иглокожие
[ редактировать ]Криноидеи диверсифицировались на протяжении всего юрского периода, достигая пика мезозойского разнообразия в поздней юре, в первую очередь за счет радиации сидячих форм, принадлежащих к отрядам Cyrtocrinida и Millericrinida . [ 284 ] Ехиноиды (морские ежи) претерпели существенную диверсификацию, начиная с ранней юры, в первую очередь за счет распространения неправильных (асимметричных) форм, которые адаптировались к питанию отложениями. Темпы диверсификации резко упали в позднеюрском периоде. [ 285 ]
Ракообразные
[ редактировать ]
Юрский период был важным временем для эволюции десятиногих . [ 279 ] Первые настоящие крабы ( Brachyura ) известны из ранней юры, причем самым ранним из них был Eocarcinus praecursor из раннего плинсбахского периода в Англии, у которого отсутствовала крабоподобная морфология ( карцинизация ) современных крабов. [ 286 ] и Eoprosopon klugi из позднего плинсбаха Германии, которые могут принадлежать современному семейству Homolodromiidae . [ 287 ] Большинство юрских крабов известны только по кусочкам панциря , что затрудняет определение их родства. [ 288 ] Хотя крабы были редкостью в ранней и средней юре, они стали многочисленными в поздней юре, когда они распространились из своей древней среды обитания на илистом морском дне в места обитания с твердым субстратом, такие как рифы, с расщелинами в рифах, обеспечивающими убежище от хищников. [ 288 ] [ 279 ] Раки-отшельники также впервые появились в юрском периоде, причем самым ранним из них был Schobertella hoelderi из позднего геттанга Германии. [ 289 ] Ранние раки-отшельники связаны с раковинами аммонитов, а не с раковинами брюхоногих моллюсков. [ 290 ] Глифеиды , которые сегодня известны только из двух видов, достигли своего пика разнообразия в юрском периоде: около 150 видов из общей летописи окаменелостей, насчитывающей 250, известных за этот период. [ 291 ] Юрские ракушки отличались меньшим разнообразием по сравнению с современными. [ 292 ] но известно несколько важных эволюционных нововведений, в том числе первые появления кальцитовых панцирных форм и видов с эпипланктонным образом жизни. [ 293 ]
Брахиоподы
[ редактировать ]Разнообразие брахиопод сократилось во время триасово-юрского вымирания. Шпильоносные брахиоподы ( Spiriferinida и Athyridida ) не восстановили своего биоразнообразия, вымерев в ТОАЭ. [ 294 ] Число Rhynchonellida и Terebratulida также сократилось во время триасово-юрского вымирания, но восстановилось в раннем юрском периоде; ни одна из клад не претерпела особых морфологических изменений. [ 295 ] В поздней юре количество брахиопод существенно сократилось; причины плохо изучены. Предполагаемые причины включают усиление хищничества, конкуренцию с двустворчатыми моллюсками, усиленную биотурбацию или повышенную нагрузку на выпас . [ 296 ]
Мшанки
[ редактировать ]Как и в предшествующий триас, разнообразие мшанок было относительно низким по сравнению с палеозоем. Подавляющее большинство юрских мшанок относятся к представителям Cyclostomatida , испытавшим радиацию в средней юре, при этом все юрские представители принадлежали к подотрядам Tubuliporina и Cerioporina . Cheilostomata , доминирующая группа современных мшанок, впервые появилась в позднеюрском периоде. [ 297 ]
Моллюски
[ редактировать ]Двустворчатые моллюски
[ редактировать ]Вымирание в конце триаса оказало серьезное влияние на разнообразие двустворчатых моллюсков, хотя и мало повлияло на экологическое разнообразие двустворчатых моллюсков. Вымирание было избирательным и оказало меньшее влияние на глубоко норящих, но нет никаких свидетельств дифференцированного воздействия между поверхностно живущими (эпифаунальными) и роющими (инфаунальными) двустворчатыми моллюсками. [ 298 ] Разнообразие на уровне семейства двустворчатых моллюсков после ранней юры оставалось неизменным, хотя видовое разнообразие постепенно увеличивалось на протяжении всего периода. [ 299 ] Рудисты , доминирующие рифообразующие организмы мелового периода, впервые появились в поздней юре (середине оксфорда) на северной окраине западного Тетиса, а к концу юры распространились на восточный Тетис. [ 300 ]
Головоногие моллюски
[ редактировать ]
Аммониты были опустошены вымиранием в конце триаса, и лишь несколько родов, принадлежащих к семейству Psiloceratidae подотряда Phylloceratina, выжили и стали предками всех более поздних юрских и меловых аммонитов. отряды Psiloceratina , Ammonitina , Lytoceratina , Haploceratina , Perisphinctina и Ancyloceratina Аммониты резко разнообразились в раннем юрском периоде: в юрском периоде появились . Фауна аммонитов в юрском периоде была региональной и была разделена примерно на 20 различных провинций и субпровинций в двух областях: северной высокоширотной панбореальной области, состоящей из Арктики, северной Панталассы и северной Атлантики, и экваториально-южной Пан-Тетийской области. царство, включавшее Тетис и большую часть Панталассы. [ 301 ] Диверсификация аммонитов происходила одновременно с морскими трансгрессиями , тогда как надиры их разнообразия происходили во время морских регрессий . [ 302 ]
Самые старые окончательные находки кальмароподобных белемнитов относятся к самой ранней юре (геттан-синемюр) Европы и Японии; они распространились по всему миру во время юрского периода. [ 303 ] Белемниты были мелководными обитателями, населявшими верхние 200 метров толщи воды на континентальных шельфах и в прибрежной зоне . Они были ключевыми компонентами юрских экосистем как хищниками, так и жертвами, о чем свидетельствует обилие белемнитов-охранников в юрских породах. [ 304 ]
Самые ранние вампироморфы , единственным живым представителем которых является кальмар-вампир , впервые появились в раннем юрском периоде. [ 305 ] Самые ранние осьминоги появились в средней юре, отделившись от своих ближайших ныне живущих родственников, вампироморфов, в период от триаса до ранней юры. [ 306 ] Все юрские осьминоги известны исключительно по твердому гладиусу . [ 306 ] [ 307 ] Осьминоги, вероятно, произошли от донных ( бентических ) предков, обитавших на мелководье. [ 306 ] Протероктопус из позднесреднеюрского лагерштетта Ла Вульт-сюр-Рон , ранее интерпретируемый как ранний осьминог, теперь считается базальным таксоном вне клады, содержащей вампироморфов и осьминогов. [ 308 ]
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ «Международная хроностратиграфическая карта» (PDF) . Международная комиссия по стратиграфии.
- ^ «Dictionary.com | Значения и определения английских слов» . Словарь.com . 17 апреля 2024 г. Проверено 18 апреля 2024 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Огг, Дж.Г.; Хиннов, Луизиана; Хуанг, К. (2012), «Юрский период» , Шкала геологического времени , Elsevier, стр. 731–791, doi : 10.1016/b978-0-444-59425-9.00026-3 , ISBN 978-0-444-59425-9 , получено 5 декабря 2020 г.
- ^ Броньяр, Александр (1829). Таблица ландшафтов, составляющих земную кору или Очерк строения известной части земли [ Описание ландшафтов, составляющих земную кору или Очерк строения известных земель Земли ] ( на французском языке). Страсбург, Франция: Ф.Г. Левро – через Галлику. Из стр. 221, сноска 2: «Часто также средний или основной оолитовый известняк (большой оолит); но название «юрские террейны» кажется нам предпочтительнее […] аналогично названию цепи Юра». (Часто также средний или главный оолитовый известняк (большой оолит); но название «юрские террейны» кажется нам предпочтительнее, потому что оно более общее, потому что указывает на ландшафт, сложенный разными породами, находящийся в геогностическом положении, аналогичном тому, сети Джура.)
- ^ фон Бух, Л., 1839. О Юре в Германии. Королевская прусская академия наук, Берлин, с. 87.
- ^ Коэн, К.М., Финни, СК, Гиббард, П.Л. и Фан, Дж.-Х. (2013; обновлено) Международная хроностратиграфическая карта ICS . Эпизоды 36: 199–204.
- ^ Хиллебрандт, Авеню; Кристин, Л.; Кюршнер, ВМ; Бонис, Северная Каролина; Руль, М.; Ричоз, С.; Шоббен, МАН; Урличс, М.; Баун, PR; Кмент, К.; МакРобертс, Калифорния (01 сентября 2013 г.). «Глобальные стратотипические разрезы и точки (GSSP) основания юрской системы в Куйохе (горы Карвендель, Северные известняковые Альпы, Тироль, Австрия)» . Эпизоды . 36 (3): 162–198. дои : 10.18814/epiiugs/2013/v36i3/001 . ISSN 0705-3797 .
- ^ Блоос, Герт; Пейдж, Кевин Н. (01 марта 2002 г.). «Глобальный стратотипический разрез и точка основания синемурского яруса (нижняя юра)» . Эпизоды . 25 (1): 22–28. дои : 10.18814/epiiugs/2002/v25i1/003 . ISSN 0705-3797 .
- ^ Мейстер, Кристиан; Аберхан, Мартин; Блау, Иоахим; Доммерг, Жан-Луи; Файст-Буркхардт, Сюзанна; Хейлвуд, Эрни А.; Харт, Малком; Хессельбо, Стивен П.; Хаунслоу, Марк В.; Хилтон, Марк; Мортон, Никол (1 июня 2006 г.). «Разрез и точка стратотипа глобальной границы (GSSP) для основания плинсбахского яруса (нижняя юра), Уайн-Хейвен, Йоркшир, Великобритания» . Эпизоды . 29 (2): 93–106. дои : 10.18814/epiiugs/2006/v29i2/003 . ISSN 0705-3797 .
- ^ РОША, Рожерио Бордало да; Маттиоли, Эмануэла; Дуарте, Луис Витор; Питтет, Бернард; Эльми, Серж; Мутерде, Рене; КАБРАЛЬ, Мария Кристина; Комас-Ренхифо, Мария Хосе; ГОМЕС, Хуан Хосе; ГОЙ, Антонио; Хессельбо, Стивен П. (01 сентября 2016 г.). «Основа тоарского яруса нижней юры, определяемая разрезом и точкой стратотипа глобальной границы (GSSP) в разрезе Пенише (Португалия)» . Эпизоды . 39 (3): 460–481. дои : 10.18814/epiiugs/2016/v39i3/99741 . hdl : 10261/140775 . ISSN 0705-3797 . S2CID 131791652 .
- ^ Бэррон, Эдвард; Уретра, Одиночество; Гой, Антонио; Лассалетта, Луис (август 2010 г.). «Палинология тоарско-ааленского стратотипического разреза и точки глобальной границы (GSSP) в Фуэнтельсасе (нижняя – средняя юра, Иберийский хребет, Испания)» . Обзор палеоботаники и палинологии . 162 (1): 11–28. Бибкод : 2010RPaPa.162...11B . дои : 10.1016/j.revpalbo.2010.04.003 .
- ^ Павия, Г.; Энай, Р. (1 марта 1997 г.). «Определение границы аален-байосского яруса» . Эпизоды . 20 (1): 16–22. дои : 10.18814/epiiugs/1997/v20i1/004 . ISSN 0705-3797 .
- ^ Лопес, Фернандес; Рафаэль, Сикст; Павия, Хулио; Эрба, Элизабет; Гиомар, Мьетт; Пайва Энрикес, Мария Хелена; Ланца, Роберто; Мангольд, Чарльз; Мортон, Никол; Оливеро, Давиде; Тирабоски, Даниэле (2009). «Разрез и точка стратотипа глобальной границы (GSSP) основания батского яруса (средняя юра), разрез Равен-дю-Бес, юго-восток Франции» (PDF) . Эпизоды . 32 (4): 222–248. дои : 10.18814/epiiugs/2009/v32i4/001 . S2CID 51754708 . Архивировано из оригинала (PDF) 4 марта 2016 года . Проверено 5 июня 2015 г.
- ^ Перейти обратно: а б с д «Международная комиссия по стратиграфии-Подкомиссия по стратиграфии юрского периода» . jurassic.stratigraphy.org . Проверено 9 апреля 2021 г.
- ^ БАРСКИЙ, Марцин (06 сентября 2018 г.). «Комплексы кист динофлагеллят на границе оксфорда и кимериджа (верхняя юра) в Флодигарри, залив Стаффин, остров Скай, Шотландия - предлагаемый GSSP для основания кимериджа» . Юрский том . XV (1): 51–6 doi : 10.5604/01.3001.0012.4594 (неактивен 20 мая 2024 г.). ISSN 1731-3708 . S2CID 133861564 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ УИМБЛДОН, Уильям А.П. (27 декабря 2017 г.). «Разработки по установлению границы титона и берриаса (J/K)» . Волюмина Юрская . 15 (1): 107–112. дои : 10.5604/01.3001.0010.7467 . ISSN 1731-3708 .
- ^ Уимблдон, Уильям А. П.; Рехакова, Даниэла; Свободова, Андреа; Шнабль, Питер; Прюнер, Питер; Эльбра, Тииу; Шифнерова, Кристина; Кдыр, Саймон; Фрау, Камилла; Шнайдер, Иоганн; Гальбрун, Бруно (11 февраля 2020 г.). «Установка границы J/K: сравнительный анализ ключевых профилей титона и берриаса в департаментах Дром и Верхние Альпы, Франция» . Геология Карпатика . 71 (1). дои : 10.31577/GeolCarp.71.1.3 . S2CID 213694912 .
- ^ Сестра Камилла; Булот, Люк Г.; Рехакова, Даниэла; Уимблдон, Уильям А. П.; Ифрим, Кристина (ноябрь 2016 г.). «Ревизия индексного вида аммонитов Berriasella jacobi Mazenot, 1939 и ее последствия для биостратиграфии берриасского яруса» . Меловые исследования . 66 : 94–114. Бибкод : 2016CrRes..66...94F . дои : 10.1016/j.cretres.2016.05.007 .
- ^ Готье Д.Л. (2005). «Общая нефтегазовая система киммериджских сланцев провинции Грабен Северного моря» (PDF) . Геологическая служба США . Проверено 2 ноября 2018 г.
- ^ Уилсон, АО (2020). «Глава 1. Знакомство с юрским Аравийским внутришельфовым бассейном» . Геологическое общество, Лондон, Мемуары . 53 (1): 1–19. дои : 10.1144/M53.1 . ISSN 0435-4052 . S2CID 226967035 .
- ^ Абдула, Рзгер А. (август 2015 г.). «Углеводородный потенциал формации Саргелу и корреляция с источниками нефти, Иракский Курдистан» . Арабский журнал геонаук . 8 (8): 5845–5868. Бибкод : 2015ArJG....8.5845A . дои : 10.1007/s12517-014-1651-0 . ISSN 1866-7511 . S2CID 129120960 .
- ^ Соранский университет; Абдула, Рзгер А. (16 октября 2016 г.). «Оценка исходных пород формации Наокелекан в Иракском Курдистане» . Журнал Занкоя Сулеймани – Часть А. 19 (1): 103–124. дои : 10.17656/jzs.10589 .
- ^ Ао, Вэйхуа; Хуан, Вэньхуэй; Венг, Чэнминь; Сяо, Сюлин; Лю, Дамэн; Тан, Сюи; Чен, Пин; Чжао, Чжиген; Ван, Хуан; Финкельман, Роберт Б. (январь 2012 г.). «Петрология угля и генезис юрского угля в бассейне Ордос, Китай» . Геонаучные границы . 3 (1): 85–95. Бибкод : 2012GeoFr...3...85A . дои : 10.1016/j.gsf.2011.09.004 .
- ^ Кенни, Гэвин Г.; Харриган, Клэр О.; Шмитц, Марк Д.; Кроули, Джеймс Л.; Уолл, Кори Дж.; Андреоли, Марко АГ; Гибсон, Роджер Л.; Майер, Вольфганг Д. (01 августа 2021 г.). «Временные масштабы кристаллизации пластинчатого расплава и точный возраст ударной структуры Мороквенг, Южная Африка» . Письма о Земле и планетологии . 567 : 117013. Бибкод : 2021E&PSL.56717013K . дои : 10.1016/j.epsl.2021.117013 . ISSN 0012-821X . S2CID 235666971 .
- ^ Хольм-Альвмарк, Санна; Журдан, Фред; Ферьер, Людовик; Алвмарк, Карл; Кеберл, Кристиан (15 мая 2021 г.). «Определение возраста ударной структуры Пучеж-Катунки (Россия) на предмет изменений и унаследованного 40Ar* – Нет связи с вымиранием» . Geochimica et Cosmochimica Acta . 301 : 116–140. Бибкод : 2021GeCoA.301..116H . дои : 10.1016/j.gca.2021.03.001 . S2CID 233620694 .
- ^ Перейти обратно: а б Скотезе, Кристофер Р. (30 мая 2021 г.). «Атлас палеогеографических карт фанерозоя: моря приходят и моря уходят» . Ежегодный обзор наук о Земле и планетах . 49 (1): 679–728. Бибкод : 2021AREPS..49..679S . doi : 10.1146/annurev-earth-081320-064052 . ISSN 0084-6597 . S2CID 233708826 .
- ^ Перейти обратно: а б Фризон де Ламотт, Доминик; Фурдан, Брендан; Леле, Софи; Лепармантье, Франсуа; де Кларанс, Филипп (24 апреля 2015 г.). «Стиль рифтогенеза и этапы распада Пангеи» . Тектоника . 34 (5): 1009–1029. Бибкод : 2015Tecto..34.1009F . дои : 10.1002/2014TC003760 . S2CID 135409359 .
- ^ Хоссейнпур, Марал; Уильямс, Саймон; Сетон, Мария; Барнетт-Мур, Николас; Мюллер, Р. Дитмар (2 октября 2016 г.). «Тектоническая эволюция Западной Тетиса от юры до наших дней: сочетание геологических и геофизических данных с моделями сейсмической томографии» . Международное геологическое обозрение . 58 (13): 1616–1645. Бибкод : 2016IGRv...58.1616H . дои : 10.1080/00206814.2016.1183146 . hdl : 2123/20835 . ISSN 0020-6814 . S2CID 130537970 .
- ^ Барт, Г.; Франц, М.; Хойниш, К.; Эрнст, В.; Циммерманн, Дж.; Вольфграмм, М. (01 января 2018 г.). «Морская и наземная седиментация через переход T – J в Северо-Германском бассейне» . Палеогеография, Палеоклиматология, Палеоэкология . 489 : 74–94. Бибкод : 2018PPP...489...74B . дои : 10.1016/j.palaeo.2017.09.029 . ISSN 0031-0182 .
- ^ Корте, Кристоф; Хессельбо, Стивен П.; Ульманн, Клеменс В.; Дитль, Герд; Рул, Миха; Швайгерт, Гюнтер; Тибо, Николя (декабрь 2015 г.). «Юрский климатический режим, управляемый океанскими воротами» . Природные коммуникации . 6 (1): 10015. Бибкод : 2015NatCo...610015K . дои : 10.1038/ncomms10015 . ISSN 2041-1723 . ПМК 4682040 . ПМИД 26658694 .
- ^ Бьеррум, Кристиан Дж.; Сурлык, Финн; Калломон, Джон Х.; Слингерленд, Руди Л. (август 2001 г.). «Численное палеоокеанографическое исследование раннеюрского трансконтинентального Лавразийского морского пути» . Палеоокеанография и палеоклиматология . 16 (4): 390–404. Бибкод : 2001PalOc..16..390B . дои : 10.1029/2000PA000512 . S2CID 128465643 .
- ^ Митчелл, Эндрю Дж.; Эллисон, Питер А.; Горман, Джеральд Дж.; Пигготт, Мэтью Д.; Пейн, Кристофер К. (1 марта 2011 г.). «Приливная циркуляция в древнем эпиконтинентальном море: Лавразийский морской путь ранней юры» . Геология . 39 (3): 207–210. Бибкод : 2011Гео....39..207М . дои : 10.1130/G31496.1 . Проверено 21 апреля 2023 г.
- ^ Апчерч, Пол; Ханн, Крейг А.; Норман, Дэвид Б. (22 марта 2002 г.). «Анализ биогеографии динозавров: доказательства существования закономерностей викариантности и расселения, вызванных геологическими событиями» . Труды Королевского общества B: Биологические науки . 269 (1491): 613–621. дои : 10.1098/рспб.2001.1921 . ПМК 1690931 . ПМИД 11916478 .
- ^ Гейгер, Маркус; Кларк, Дэвид Норман; Метте, Вольфганг (март 2004 г.). «Переоценка времени распада Гондваны на основе седиментологических и сейсмических данных из бассейна Морондава, Мадагаскар» . Журнал африканских наук о Земле . 38 (4): 363–381. Бибкод : 2004JAfES..38..363G . doi : 10.1016/j.jafrearsci.2004.02.003 .
- ^ Нгуен, Луан С.; Холл, Стюарт А.; Берд, Дейл Э.; Болл, Филип Дж. (июнь 2016 г.). «Реконструкция континентальных окраин Восточной Африки и Антарктиды: АФРИКАНО-АНТАРКТИДСКАЯ РЕКОНСТРУКЦИЯ» . Журнал геофизических исследований: Solid Earth . 121 (6): 4156–4179. дои : 10.1002/2015JB012776 .
- ^ Итурральде-Винент, Мануэль А. (1 января 2003 г.). «Противоречивые палеонтологические и стратиграфические данные о формировании Карибского морского пути» . Мексиканский залив и Карибский бассейн: среда обитания углеводородов, формирование бассейна и тектоника плит . Том. 79. Американская ассоциация геологов-нефтяников . дои : 10.1306/M79877 . ISBN 978-1-62981-054-6 .
- ^ Блейки, Рональд К.; Рэнни, Уэйн Д. (2018), «Прибытие Врангелии и невадская складчатость: от позднего триаса до поздней юры: около 240–145 млн лет назад» , Древние ландшафты западной части Северной Америки , Чам: Springer International Publishing, стр. 89– 101, номер домена : 10.1007/978-3-319-59636-5_7 , ISBN 978-3-319-59634-1 , получено 10 апреля 2021 г.
- ^ Кленнетт, Эдвард Дж.; Сиглох, Карин; Михалинюк, Митчелл Г.; Сетон, Мария; Хендерсон, Марта А.; Хоссейни, Касра; Мохаммадзахери, Афсане; Джонстон, Стивен Т.; Мюллер, Р. Дитмар (август 2020 г.). «Количественная реконструкция томотектонических плит западной части Северной Америки и восточно-тихоокеанского бассейна» . Геохимия, геофизика, геосистемы . 21 (8): e09117. Бибкод : 2020GGG....2109117C . дои : 10.1029/2020GC009117 . ISSN 1525-2027 . S2CID 225443040 .
- ^ Йи, Чжию; Меерт, Джозеф Г. (16 августа 2020 г.). «Закрытие Монголо-Охотского океана средней юрой: сопоставление палеомагнитных и геологических данных» . Письма о геофизических исследованиях . 47 (15). Бибкод : 2020GeoRL..4788235Y . дои : 10.1029/2020GL088235 . ISSN 0094-8276 . S2CID 225430978 .
- ^ Бошман, Лидиан М.; ван Хинсберген, Доу Джей Джей (июль 2016 г.). «О загадочном рождении Тихоокеанской плиты в океане Панталасса» . Достижения науки . 2 (7): e1600022. Бибкод : 2016SciA....2E0022B . дои : 10.1126/sciadv.1600022 . ISSN 2375-2548 . ПМК 5919776 . ПМИД 29713683 .
- ^ Даниз, Сильвия; Холланд, Стивен М. (июль 2018 г.). «Стратиграфическая структура последовательностей для средней и поздней юры пролива Сандэнс, штат Вайоминг: последствия для корреляции, эволюции бассейна и изменения климата» . Журнал геологии . 126 (4): 371–405. Бибкод : 2018JG....126..371D . дои : 10.1086/697692 . ISSN 0022-1376 . S2CID 133707199 .
- ^ Хак, Билал У. (01 января 2018 г.). «Юрские изменения уровня моря: переоценка» . GSA Сегодня : 4–10. дои : 10.1130/GSATG359A.1 .
- ^ Вульпиус, Сара; Кисслинг, Вольфганг (январь 2018 г.). «Новые ограничения на последний морской переход арагонит-кальцит из раннеюрских ооидов» . Фации . 64 (1): 3. Бибкод : 2018Faci...64....3V . дои : 10.1007/s10347-017-0516-x . ISSN 0172-9179 . S2CID 135202813 .
- ^ Эйхензер, Килиан; Бальтазар, Уве; Смарт, Кристофер В.; Стандер, Джулиан; Хаага, Кристиан А.; Кисслинг, Вольфганг (август 2019 г.). «Юрский переход от абиотического к биотическому контролю над морским экологическим успехом» . Природа Геонауки . 12 (8): 638–642. дои : 10.1038/s41561-019-0392-9 . hdl : 10026.1/14472 . ISSN 1752-0894 . S2CID 197402218 .
- ^ Дай, Сяньдуо; Ду, Юаньшэн; Зиглер, Мартин; Ван, Чаовэн; Ма, Цяньли; Чай, Ронг; Го, Хуа (1 января 2022 г.). «Изменение климата в период от среднего триаса до поздней юры на северной окраине Южно-Китайской плиты: данные по индексам химического выветривания и минералогии глины» . Палеогеография, Палеоклиматология, Палеоэкология . 585 : 110744. Бибкод : 2022PPP...58510744D . дои : 10.1016/j.palaeo.2021.110744 . hdl : 1874/419504 . S2CID 243463781 . Проверено 9 января 2023 г.
- ^ Перейти обратно: а б Селлвуд, Брюс В.; Вальдес, Пол Дж. (2008). «Юрский климат» . Труды Ассоциации геологов . 119 (1): 5–17. Бибкод : 2008ПрГА..119....5С . дои : 10.1016/S0016-7878(59)80068-7 .
- ^ Перейти обратно: а б с д и ж г Скотезе, Кристофер Р.; Сун, Хайджун; Миллс, Бенджамин Дж.В.; ван дер Меер, Дауве Г. (апрель 2021 г.). «Палеотемпературы фанерозоя: изменение климата Земли за последние 540 миллионов лет» . Обзоры наук о Земле . 215 : 103503. Бибкод : 2021ESRv..21503503S . doi : 10.1016/j.earscirev.2021.103503 . ISSN 0012-8252 . S2CID 233579194 . Архивировано из оригинала 8 января 2021 года. Альтернативный URL.
- ^ Ландверс, Ян; Фёльнер, Георг; Петри, Стефан; Самес, Бенджамин; Вагрейх, Михаэль (20 мая 2021 г.). «Исследование тенденций и чувствительности мезозойского климата с помощью большого набора климатических моделей» . Палеоокеанография и палеоклиматология . 36 (6): e2020PA004134. Бибкод : 2021PaPa...36.4134L . дои : 10.1029/2020PA004134 . ПМЦ 8251552 . ПМИД 34240008 .
- ^ Чжан, Чжихуэй; Льв, Давэй; Лу, Человек; Ю, Цзычэн; Гао, Юань; Ван, Тяньтянь; Гао, Цзе; Ван, Чэншань (март 2023 г.). «Активность лесных пожаров, вызванная 405-тысячными орбитальными климатическими циклами в средней юре» . Глобальные и планетарные изменения . 222 : 104069. Бибкод : 2023GPC...22204069Z . дои : 10.1016/j.gloplacha.2023.104069 . S2CID 257059454 . Проверено 16 сентября 2023 г.
- ^ Олмон, Уоррен Д.; Мартин, Рональд Э. (весна 2014 г.). «Возвращение к морепродуктам во времени: фанерозойское увеличение морских трофических ресурсов и его макроэволюционные последствия» . Палеобиология . 40 (2): 256–287. дои : 10.1666/13065 . ISSN 0094-8373 . S2CID 86765146 . Проверено 16 сентября 2023 г.
- ^ Бужо, Седрик; Пелленард, Пьер; Деконинк, Жан-Франсуа; Хессельбо, Стивен П.; Доммерг, Жан-Луи; Брюно, Людовик; Кокерес, Теофиль; Лаффон, Реми; Гюре, Эмилия; Тибо, Николас (февраль 2017 г.). «Климатические и палеоокеанографические изменения в плинсбахском периоде (ранняя юра), выведенные на основе минералогии глин и геохимии стабильных изотопов (CO) (северо-запад Европы)» . Глобальные и планетарные изменения . 149 : 139–152. Бибкод : 2017GPC...149..139B . дои : 10.1016/j.gloplacha.2017.01.005 . hdl : 10871/25335 . Проверено 14 мая 2023 г.
- ^ Рюбзам, Вольфганг; Майер, Бернхард; Шварк, Лоренц (январь 2019 г.). «Динамика углерода в криосфере контролирует глобальное потепление в раннем тоаре и эволюцию уровня моря» . Глобальные и планетарные изменения . 172 : 440–453. Бибкод : 2019ГПЦ...172..440Р . дои : 10.1016/j.gloplacha.2018.11.003 . S2CID 133660136 .
- ^ Рюбзам, Вольфганг; Шварк, Лоренц (11 мая 2021 г.). «Влияние криосферы северного полушария на климат позднего плинсбаха – начала тоара и эволюцию окружающей среды» . Геологическое общество, Лондон, специальные публикации . 514 (1): СП514–2021–11. Бибкод : 2021GSLSP.514..359R . дои : 10.1144/SP514-2021-11 . ISSN 0305-8719 . S2CID 236600012 .
- ^ Икеда, Масаюки; Боле, Максимилиан; Баумгартнер, Питер О. (1 сентября 2016 г.). «Изменения в окислительно-восстановительном состоянии в орбитальном масштабе и потоки биогенного кремнезема/обломков среднеюрского радиолярита в Тефии (Согно, Ломбардия, Северная Италия): возможная связь с оледенением?» . Палеогеография, Палеоклиматология, Палеоэкология . 457 : 247–257. Бибкод : 2016PPP...457..247I . дои : 10.1016/j.palaeo.2016.06.009 . Проверено 10 апреля 2023 г.
- ^ Ли, Цзюнь; Хуан, Ченг-Мин; Ян, Го-Линь; Пан, Юань-Юань; Вэнь, Син-Юэ (январь 2022 г.). «Колебания климата средней юры по палеопочвенным записям бассейна Сычуань, юго-запад Китая» . Журнал палеогеографии . 11 (1): 97–122. Бибкод : 2022JPalG..11...97L . дои : 10.1016/j.jop.2022.01.003 . S2CID 252949484 .
- ^ Чжан, Чжихуэй; Ван, Чэншань; Льв, Давэй; Хэй, Уильям В.; Ван, Тяньтянь; Цао, Шуо (1 января 2020 г.). «Воздействие климата в масштабе прецессии на лесные пожары на торфяниках в парниковый период ранней и средней юры» . Глобальные и планетарные изменения . 184 : 103051. Бибкод : 2020GPC...18403051Z . дои : 10.1016/j.gloplacha.2019.103051 . ISSN 0921-8181 . Проверено 26 февраля 2024 г. - через Elsevier Science Direct.
- ^ Хессельбо, Стивен П.; Морганс-Белл, Хелен С.; МакЭлвейн, Дженнифер С.; Рис, П. Макаллистер; Робинсон, Стюарт А.; Росс, К. Элизабет (май 2003 г.). «Возмущение углеродного цикла в средней юре и сопутствующие изменения в земной палеосреде» . Журнал геологии . 111 (3): 259–276. Бибкод : 2003JG....111..259H . дои : 10.1086/373968 . ISSN 0022-1376 . Получено 4 мая 2024 г. - из журналов прессы Чикагского университета.
- ^ Вежбовский, Х.; Иоахимски, М. (22 октября 2007 г.). «Реконструкция морской палеосреды позднего байоса-бата с использованием соотношений изотопов углерода и кислорода известковых окаменелостей из польской цепи Юра (центральная Польша)» . Палеогеография, Палеоклиматология, Палеоэкология . 254 (3–4): 523–540. Бибкод : 2007PPP...254..523W . дои : 10.1016/j.palaeo.2007.07.010 . Проверено 10 апреля 2023 г.
- ^ Дженкинс, Хью К.; Схаутен-Хайберс, Л.; Схаутен, С.; Синнингхе Дамсте, Яап С. (2 февраля 2012 г.). «Теплые температуры поверхности моря в высоких широтах средней юры – раннего мела Южного океана» . Климат прошлого . 8 (1): 215–226. Бибкод : 2012CliPa...8..215J . дои : 10.5194/cp-8-215-2012 . S2CID 203095372 . Проверено 8 апреля 2023 г.
- ^ Перейти обратно: а б Ли, Гаоцзе; Ся, Гоцин; Йи, Хайшэн; Ву, Чихуа; Ваграйх, Михаэль (15 сентября 2022 г.). «Изменения климата, зафиксированные в стабильном изотопном составе углерода позднеюрской морской осадочной последовательности в бассейне Цянтан, Северный Тибет» . Журнал азиатских наук о Земле . 236 : 105317. Бибкод : 2022JAESc.23605317L . дои : 10.1016/j.jseaes.2022.105317 . S2CID 250103419 . Проверено 8 января 2023 г.
- ^ Дромарт, Г.; Гарсия, Ж.-П.; Пикард, С.; Атропс, Ф.; Лекюйер, К.; Шеппард, SMF (25 августа 2003 г.). «Ледниковый период на переходном этапе средней и поздней юры?» . Письма о Земле и планетологии . 213 (3–4): 205–220. Бибкод : 2003E&PSL.213..205D . дои : 10.1016/S0012-821X(03)00287-5 . Проверено 8 января 2023 г.
- ^ Филипп, Марк; Пухалон, Сара; Суан, Гийом; Муссе, Сильвен; Тевенар, Фредерик; Маттиоли, Эмануэла (15 января 2017 г.). «Палеоширотное распределение родов ископаемой древесины как показатель европейского юрского наземного климата» . Палеогеография, Палеоклиматология, Палеоэкология . 466 : 373–381. Бибкод : 2017PPP...466..373P . дои : 10.1016/j.palaeo.2016.11.029 . Проверено 10 июня 2023 г.
- ^ Их, ТР; Гилл, Британская Колумбия; Карутерс, А.Х.; Грёке, доктор медицинских наук; Тульский, ET; Мартиндейл, Колорадо; Поултон, ТП; Смит, Польша (февраль 2017 г.). «Записи изотопов углерода высокого разрешения тоарского океанического бескислородного события (ранняя юра) из Северной Америки и последствия для глобальных движущих сил тоарского углеродного цикла» . Письма о Земле и планетологии . 459 : 118–126. Бибкод : 2017E&PSL.459..118T . дои : 10.1016/j.epsl.2016.11.021 .
- ^ Роуз-Френч, Соня; Эчеваррия, Хавьер; Дамбореня, Сьюзен Э.; Мансенидо, Майкл О.; Дженкинс, Хью К.; Аль-Сувайди, Аиша; Хессельбо, Стивен П.; Риккарди, Альберт К. (1 июля 2019 г.). «Реакция населения во время океанического бескислородного явления: случай Posidonotis (Bivalvia) из нижней юры бассейна Неукен, Аргентина » Палеогеография, Палеоклиматология, Палеоэкология 525 : 57–67. Бибкод : 2019PPP...525... 57R дои : 10.1016/j.palaeo.2019.04.009 . hdl : 11336/128130 . S2CID 146525666 . Получено 23 ноября.
- ^ Перейти обратно: а б с Виналл, Пол Б.; Бонд, Дэвид П.Г. (2008). «Записи о массовом вымирании в конце триаса и ранней юре на Британских островах» . Труды Ассоциации геологов . 119 (1): 73–84. Бибкод : 2008PrGA..119...73W . дои : 10.1016/S0016-7878(08)80259-3 . Проверено 23 ноября 2022 г.
- ^ Перейти обратно: а б Реолид, Матиас; Маттиоли, Эмануэла; Дуарте, Луис В.; Рюбзам, Вольфганг (22 сентября 2021 г.). «Тоарское океаническое бескислородное событие: где мы находимся?» . Геологическое общество, Лондон, специальные публикации . 514 (1): 1–11. Бибкод : 2021GSLSP.514....1R . дои : 10.1144/SP514-2021-74 . ISSN 0305-8719 . S2CID 238683028 .
- ^ РОДРИГЕС, Бруно; Дуарте, Луис В.; СИЛЬВА, Рикардо Л.; Мендонса Филью, Жоау Грасиано (15 сентября 2020 г.). «Осадочное органическое вещество и ранние тоарские изменения окружающей среды в Лузитанском бассейне (Португалия)» . Палеогеография, Палеоклиматология, Палеоэкология . 554 : 109781. Бибкод : 2020PPP...55409781R . дои : 10.1016/j.palaeo.2020.109781 . S2CID 219059687 . Проверено 27 сентября 2022 г.
- ^ Дера, Гийом; Неж, Паскаль; Доммерг, Жан-Луи; Фара, Эммануэль; Лаффон, Реми; Пелленард, Пьер (январь 2010 г.). «Динамика раннего юрского морского вымирания с высоким разрешением: на примере плинсбахско-тоарских аммонитов (Cephalopoda)» . Журнал Геологического общества . 167 (1): 21–33. Бибкод : 2010JGSoc.167...21D . дои : 10.1144/0016-76492009-068 . ISSN 0016-7649 . S2CID 128908746 .
- ^ Ариас, Кармен (1 октября 2013 г.). «События вымирания остракод в раннем тоаре (ранней юре) на Пиренейском хребте: влияние изменений температуры и длительного воздействия низких концентраций растворенного кислорода» . Палеогеография, Палеоклиматология, Палеоэкология . 387 : 40–55. Бибкод : 2013PPP...387...40A . дои : 10.1016/j.palaeo.2013.07.004 . Проверено 23 ноября 2022 г.
- ^ Хесс, Сильвия; Надь, Йено; Лаурсен, Гитте Вестергор (28 января 2014 г.). «Бентосные фораминиферы из нижнеюрских трансгрессивных аргиллитов юго-западной части Баренцева моря — возможное высокоширотное выражение глобального плинсбахско-тоарского круговорота?» . Полярные исследования . 33 (1): 20206. doi : 10.3402/polar.v33.20206 . S2CID 128492520 .
- ^ Реолид, Матиас; Копстейк, Филип; Джонсон, Бен (15 октября 2019 г.). «Сообщества, вымирания и появления фораминифер, связанные с раннетоарским океаническим бескислородным событием в скважине Лланбедр (ферма Мохрас), бассейн залива Кардиган, Соединенное Королевство» . Палеогеография, Палеоклиматология, Палеоэкология . 532 : 109277. Бибкод : 2019PPP...53209277R . дои : 10.1016/j.palaeo.2019.109277 . S2CID 200072488 . Проверено 23 ноября 2022 г.
- ^ Даниз, Сильвия; Клеманс, Мари-Эмили; Прайс, Грегори Д.; Мерфи, Дэниел П.; Гомес, Хуан Дж.; Твитчетт, Ричард Дж. (15 июня 2019 г.). «Стратиграфический и экологический контроль изменения морского донного сообщества в результате раннего тоарского вымирания (Иберийский хребет, Испания)» . Палеогеография, Палеоклиматология, Палеоэкология . 524 : 183–200. Бибкод : 2019PPP...524..183D . дои : 10.1016/j.palaeo.2019.03.039 . hdl : 10026.1/13668 . S2CID 134835736 . Проверено 23 ноября 2022 г.
- ^ Карутерс, Эндрю Х.; Смит, Пол Л.; Грёке, Даррен Р. (сентябрь 2013 г.). «Плинсбахско-тоарское (раннеюрское) вымирание — глобальное многоэтапное событие» . Палеогеография, Палеоклиматология, Палеоэкология . 386 : 104–118. Бибкод : 2013PPP...386..104C . дои : 10.1016/j.palaeo.2013.05.010 .
- ^ Вёрёш, Аттила; Кочиш, Адам; Палфи, Йожеф (1 сентября 2016 г.). «Гибель последних двух отрядов брахиопод со шпилями (Spiriferinida и Athyridida) во время тоарского (раннеюрского) вымирания» . Палеогеография, Палеоклиматология, Палеоэкология . 457 : 233–241. Бибкод : 2016PPP...457..233В . дои : 10.1016/j.palaeo.2016.06.022 . Проверено 29 октября 2022 г.
- ^ Хоран, Фернандо Гарсия; Баэса-Карратала, Хосе Франсиско; Гой, Антонио (1 октября 2018 г.). «Изменения размера тела брахиопод до массового вымирания в раннем тоаре (юрском периоде)» . Палеогеография, Палеоклиматология, Палеоэкология . 506 : 242–249. Бибкод : 2018PPP...506..242G . дои : 10.1016/j.palaeo.2018.06.045 . hdl : 10045/77781 . S2CID 135368506 . Проверено 29 октября 2022 г.
- ^ Максвелл, Эрин Э.; Винсент, Пегги (6 ноября 2015 г.). «Влияние раннего тоарского океанического бескислородного события на размер тела ихтиозавра и состав фауны в юго-западном немецком бассейне» . Палеобиология . 42 (1): 117–126. дои : 10.1017/pab.2015.34 . ISSN 0094-8373 . S2CID 131623205 .
- ^ Сюй, Вэйму; Рул, Миха; Дженкинс, Хью К.; Хессельбо, Стивен П.; Райдинг, Джеймс Б.; Селби, Дэвид; Наафс, Б. Дэвид А.; Вейерс, Йохан WH; Панкост, Ричард Д.; Тегелаар, Эрик В.; Идиз, Эрдем Ф. (февраль 2017 г.). «Связывание углерода в расширенной озерной системе во время тоарского океанического бескислородного явления» . Природа Геонауки . 10 (2): 129–134. Бибкод : 2017NatGe..10..129X . дои : 10.1038/ngeo2871 . hdl : 10871/24965 . ISSN 1752-0894 .
- ^ Мюллер, Тамаш; Юрикова, Хана; Гутжар, Маркус; Томашович, Адам; Шлёгль, Ян; Либетрау, Волкер; Дуарте, Луис против; Миловский, Растислав; Суан, Гийом; Маттиоли, Эмануэла; Питтет, Бернард (01 декабря 2020 г.). «Закисление океана во время раннего тоарского вымирания: данные по изотопам бора в брахиоподах» . Геология . 48 (12): 1184–1188. Бибкод : 2020Geo....48.1184M . дои : 10.1130/G47781.1 . hdl : 10023/20595 . ISSN 0091-7613 .
- ^ Трекалли, Альберто; Спангенберг, Хорхе; Адатте, Тьерри; Фёлльми, Карл Б.; Паренте, Мариано (декабрь 2012 г.). «Карбонатная платформа свидетельствует о закислении океана в начале тоарского океанического бескислородного события» . Письма о Земле и планетологии . 357–358: 214–225. Бибкод : 2012E&PSL.357..214T . дои : 10.1016/j.epsl.2012.09.043 .
- ^ Эттингер, Николас П.; Ларсон, Тоти Э.; Керанс, Чарльз; Тибодо, Элисон М.; Хаттори, Келли Э.; Качур, Шон М.; Мартиндейл, Роуэн К. (23 сентября 2020 г.). Эберли, Грегор (ред.). «Закисление океана и аноксия фототической зоны в тоарском океаническом бескислородном событии: идеи Адриатической карбонатной платформы» . Седиментология . 68 : 63–107. дои : 10.1111/сед.12786 . ISSN 0037-0746 . S2CID 224870464 .
- ^ Пападоманолаки, Нина М.; Ленстра, Витце К.; Уолтерс, Мариетт; Сломп, Кэролайн П. (1 июля 2022 г.). «Усиленная рециркуляция фосфора во время прошлой океанической аноксии усиливалась низкими темпами аутигенеза апатита» . Достижения науки . 8 (26): eabn2370. Бибкод : 2022SciA....8N2370P . дои : 10.1126/sciadv.abn2370 . hdl : 1874/421467 . ПМЦ 10883373 . ПМИД 35776794 . S2CID 250218660 .
- ^ Теннант, Джонатан П.; Мэннион, Филип Д.; Апчерч, Пол (02 сентября 2016 г.). «Динамика разнообразия четвероногих, регулируемая уровнем моря, в юрско-меловом периоде» . Природные коммуникации . 7 (1): 12737. Бибкод : 2016NatCo...712737T . дои : 10.1038/ncomms12737 . ISSN 2041-1723 . ПМК 5025807 . ПМИД 27587285 .
- ^ Перейти обратно: а б Лукас, Спенсер Г.; Таннер, Лоуренс Х. (октябрь 2015 г.). «Неморские биотические события в конце триаса» . Журнал палеогеографии . 4 (4): 331–348. Бибкод : 2015JPalG...4..331L . дои : 10.1016/j.jop.2015.08.010 .
- ^ Мандер, Люк; Кюршнер, Вольфрам М.; МакЭлвейн, Дженнифер С. (31 августа 2010 г.). «Объяснение противоречивых данных о разнообразии растений триаса и юры» . Труды Национальной академии наук . 107 (35): 15351–15356. Бибкод : 2010PNAS..10715351M . дои : 10.1073/pnas.1004207107 . ISSN 0027-8424 . ПМЦ 2932585 . ПМИД 20713737 .
- ^ Барбака, Мэри; Пацина, Грегори; Кочиш, Адам Т.; Ярзинка, Агата; Зиая, Ядвига; Бодор, Эмезе (август 2017 г.). «Изменения наземной флоры на рубеже триаса и юры в Европе» . Палеогеография, Палеоклиматология, Палеоэкология . 480 : 80–93. Бибкод : 2017PPP...480...80B . дои : 10.1016/j.palaeo.2017.05.024 .
- ^ Эльгорриага, Андрес; Эскапа, Игнасио Х.; Кунео, Н. Рубен (июль 2019 г.). «Реликтовые чешуекрылые (Peltaspermales) из асфальтовой формации Раннеюрского периода Каньядон, Патагония, Аргентина» . Международный журнал наук о растениях . 180 (6): 578–596. дои : 10.1086/703461 . ISSN 1058-5893 . S2CID 195435840 .
- ^ Бомфлер, Бенджамин; Бломенкемпер, Патрик; Керп, Ганс; Маклафлин, Стивен (2018), «Полярные регионы мезозойско-палеогенового тепличного мира как убежища для групп реликтовых растений» , Трансформационная палеоботаника , Elsevier, стр. 593–611, doi : 10.1016/b978-0-12-813012-4.00024- 3 , ISBN 978-0-12-813012-4 , получено 12 ноября 2020 г.
- ^ Аткинсон, Брайан А.; Сербет, Рудольф; Хигер, Тимоти Дж.; Тейлор, Эдит Л. (октябрь 2018 г.). «Дополнительные доказательства мезозойской диверсификации хвойных пород: пыльцевая шишка Chimaerostrobus minutus gen. et sp. nov. (Coniferales) из нижней юры Антарктиды» . Обзор палеоботаники и палинологии . 257 : 77–84. Бибкод : 2018RPaPa.257...77A . дои : 10.1016/j.revpalbo.2018.06.013 . S2CID 133732087 .
- ^ Перейти обратно: а б с Лесли, Эндрю Б.; Болье, Джереми; Холман, Гарт; Кэмпбелл, Кристофер С.; Мэй, Вэньбинь; Раубсон, Линда Р.; Мэтьюз, Сара (сентябрь 2018 г.). «Обзор эволюции современных хвойных деревьев с точки зрения летописи окаменелостей» . Американский журнал ботаники . 105 (9): 1531–1544. дои : 10.1002/ajb2.1143 . ПМИД 30157290 . S2CID 52120430 .
- ^ Стоки, Рут А.; Ротвелл, Гар В. (июль 2020 г.). «Диверсификация кроновой группы араукарий: роль Araucaria famii sp. nov. в позднемеловой (кампанской) радиации араукариевых в северном полушарии» . Американский журнал ботаники . 107 (7): 1072–1093. дои : 10.1002/ajb2.1505 . ISSN 0002-9122 . ПМИД 32705687 . S2CID 225568264 .
- ^ Эскапа, Игнатий Х.; Каталано, Сантьяго А. (октябрь 2013 г.). «Филогенетический анализ араукариевых: интеграция молекул, морфология и окаменелости» . Международный журнал наук о растениях . 174 (8): 1153–1170. дои : 10.1086/672369 . hdl : 11336/3583 . ISSN 1058-5893 . S2CID 56238574 .
- ^ Стоки, Рут А.; Ротвелл, Гар В. (март 2013 г.). «Pararaucaria carrii sp. nov., Анатомически сохранившиеся свидетельства существования семейства хвойных Cheirolepidiaceae в Северном полушарии» . Международный журнал наук о растениях . 174 (3): 445–457. дои : 10.1086/668614 . ISSN 1058-5893 . S2CID 59269291 .
- ^ Эскапа, Игнасио; Кунео, Рубен; Аксмит, Брайан (сентябрь 2008 г.). «Новый род Cupressaceae (sensu lato) из юры Патагонии: значение для гомологий мегаспорангиатных шишек хвойных деревьев» . Обзор палеоботаники и палинологии . 151 (3–4): 110–122. Бибкод : 2008RPaPa.151..110E . дои : 10.1016/j.revpalbo.2008.03.002 .
- ^ Контрерас, Дори Л.; Эскапа, Игнатий Х.; Ирибаррен, Росио Дж.; Кунео, Н. Рубен (октябрь 2019 г.). «Реконструкция ранней эволюции Cupressaceae: описание всего растения нового вида Austrohamia из формации Каньядон Асфальто (ранняя юра), Аргентина» . Международный журнал наук о растениях . 180 (8): 834–868. дои : 10.1086/704831 . ISSN 1058-5893 . S2CID 202862782 .
- ^ Ма, Цин-Вэнь; К. Фергюсон, Дэвид; Лю, Хай-Мин; Сюй, Цзин-Сянь (2020). «Сжатия секвойи (Cupressaceae sensu lato) из средней юры Даохугоу, Нинчэн, Внутренняя Монголия, Китай» . Палеобиоразнообразие и палеосреда . 1 (9): 1. дои : 10.1007/s12549-020-00454-z . S2CID 227180592 . Проверено 9 марта 2021 г.
- ^ Домогацкая Ксения Владимировна; Герман, Алексей Борисович (май 2019 г.). «Новые виды рода Schizolepidopsis (хвойные) из альба Высокой Арктики России и геологическая история рода» . Меловые исследования . 97 : 73–93. Бибкод : 2019CrRes..97...73D . дои : 10.1016/j.cretres.2019.01.012 . S2CID 134849082 .
- ^ Мацунага, Келли К.С.; Херендин, Патрик С.; Кузнец, Фабиан; Ичинноров, Нииден; Крейн, Питер Р.; Ши, Гунл (10 мая 2021 г.). «Яйцеклеточные шишки Schizolepidopsis ediae sp. nov. Дают представление об эволюции сосновых» . Международный журнал наук о растениях . 182 (6): 490–507. дои : 10.1086/714281 . ISSN 1058-5893 . S2CID 235426888 .
- ^ Ротвелл, Гар В.; Мейпс, Джин; Стоки, Рут А.; Хилтон, Джейсон (апрель 2012 г.). «Семенная шишка Eathiestrobus gen. nov.: Ископаемые свидетельства юрского происхождения сосновых». Американский журнал ботаники . 99 (4): 708–720. дои : 10.3732/ajb.1100595 . ПМИД 22491001 .
- ^ Смит, Селена Ю.; Стоки, Рут А.; Ротвелл, Гар В.; Литтл, Стефан А. (2 января 2017 г.). «Новый вид Pityostrobus (Pinaceae) из мелового периода Калифорнии: движение к пониманию меловой радиации сосновых» . Журнал систематической палеонтологии . 15 (1): 69–81. Бибкод : 2017JSPal..15...69S . дои : 10.1080/14772019.2016.1143885 . ISSN 1477-2019 . S2CID 88292891 .
- ^ Донг, Чонг; Ши, Гунгл; Эррера, Фабиани; Ван, Юндон; Херендин, Патрик С; Крейн, Питер Р. (18 июня 2020 г.). «Окаменелости средней и поздней юры из северо-восточного Китая обнаруживают морфологический застой серёжки» . Национальный научный обзор . 7 (11): 1765–1767. дои : 10.1093/nsr/nwaa138 . ISSN 2095-5138 . ПМЦ 8288717 . ПМИД 34691509 .
- ^ Андрюхов-Коломбо, Ана; Эскапа, Игнатий Х; Аагесен, Одинокий; Мацунага, Келли К.С. (4 августа 2023 г.). «В поисках утраченного времени: прослеживание разнообразия ископаемых Podocarpaceae на протяжении веков» . Ботанический журнал Линнеевского общества . 203 (4): 315–336. doi : 10.1093/botlinnean/boad027 . hdl : 11336/227952 . ISSN 0024-4074 .
- ^ Поул, Майк; Ван, Юндон; Бугдаева Евгения Владимировна; Донг, Чонг; Тянь, Нин; Ли, Лицинь; Чжоу, Нин (15 декабря 2016 г.). «Взлет и упадок подозамитов в Восточной Азии — образ жизни вымерших хвойных» . Палеогеография, Палеоклиматология, Палеоэкология . Мезозойские экосистемы – Климат и биота. 464 : 97–109. Бибкод : 2016PPP...464...97P . дои : 10.1016/j.palaeo.2016.02.037 . ISSN 0031-0182 .
- ^ Перейти обратно: а б с Чжоу, Чжи-Янь (март 2009 г.). «Обзор ископаемых Ginkgoales» . Палеомир . 18 (1): 1–22. дои : 10.1016/j.palwor.2009.01.001 .
- ^ Носова, Наталья (октябрь 2013 г.). «Ревизия рода Grenana Samylina из средней юры Ангрена, Узбекистан» . Обзор палеоботаники и палинологии . 197 : 226–252. Бибкод : 2013RPaPa.197..226N . дои : 10.1016/j.revpalbo.2013.06.005 .
- ^ Донг, Чонг; Ши, Гунгл; Чжан, Сяоцин; Ван, Цзыси; Ван, Юндон (ноябрь 2022 г.). «Окаменелости средней-поздней юры из Северо-Восточного Китая подтверждают принадлежность семенных структур Umaltolepis и листьев Pseudotorellia» . Обзор палеоботаники и палинологии . 306 : 104763. Бибкод : 2022RPaPa.30604763D . дои : 10.1016/j.revpalbo.2022.104763 . S2CID 251917169 .
- ^ Перейти обратно: а б Попа, Михай Э. (июнь 2014 г.). «Раннеюрские беннеттитальные репродуктивные структуры Румынии» . Палеобиоразнообразие и палеосреда . 94 (2): 327–362. Бибкод : 2014PdPe...94..327P . дои : 10.1007/s12549-014-0165-9 . ISSN 1867-1594 . S2CID 128411467 .
- ^ Тейлор, Т. (2009), «Цикадофиты» , Биология и эволюция ископаемых растений , Elsevier, стр. 703–741, doi : 10.1016/b978-0-12-373972-8.00017-6 , ISBN 978-0-12-373972-8 , получено 12 декабря 2020 г.
- ^ Потт, Кристиан; Маклафлин, Стивен (01 июня 2014 г.). «Разнообразный характер роста Williamsoniaceae (Bennettitales): раскрытие экологии ключевой группы мезозойских растений» . Палеобиоразнообразие и палеосреда . 94 (2): 307–325. Бибкод : 2014ПдПе...94..307П . дои : 10.1007/s12549-014-0157-9 . ISSN 1867-1608 . S2CID 84440045 .
- ^ Фармер, Конрад К.; Ян, Цян; Джеймс-Блей, Джордж А.; Хоттон, Кэрол Л.; Монтейро, Энтони; Ван, Юн-Цзе; Горева, Юлия; Ши, Чункун; Сильестрем, Сандра; Роуз, Тим Р.; Дилчер, Дэвид Л. (10 февраля 2016 г.). «Эволюционное сближение среднемезозойских златоглазок и кайнозойских бабочек» . Труды Королевского общества B: Биологические науки . 283 (1824): 20152893. doi : 10.1098/rspb.2015.2893 . ISSN 0962-8452 . ПМК 4760178 . ПМИД 26842570 .
- ^ Храмов Александр Владимирович ; Лукашевич, Елена Дмитриевна (июль 2019 г.). «Юрский двукрылый опылитель с чрезвычайно длинным хоботком» . Исследования Гондваны . 71 : 210–215. Бибкод : 2019GondR..71..210K . дои : 10.1016/j.gr.2019.02.004 . S2CID 134847380 .
- ^ Перейти обратно: а б Цай, Чэньян; Эскалона, Гермес Э.; Ли, Лицинь; Инь, Цивэй; Хуан, Диин; Энгель, Майкл С. (сентябрь 2018 г.). «Опыление саговников жуками в мезозое» . Современная биология . 28 (17): 2806–2812.e1. Бибкод : 2018CBio...28E2806C . дои : 10.1016/j.cub.2018.06.036 . ПМИД 30122529 . S2CID 52038878 .
- ^ Перейти обратно: а б Койро, Марио; Потт, Кристиан (декабрь 2017 г.). «Eobowenia gen. nov. из раннего мела Патагонии: признак раннего расхождения Bowenia?» . Эволюционная биология BMC . 17 (1): 97. Бибкод : 2017BMCEE..17...97C . дои : 10.1186/s12862-017-0943-x . ISSN 1471-2148 . ПМК 5383990 . ПМИД 28388891 .
- ^ Вайда, Виви; Пучетайте, Милда; Маклафлин, Стивен; Энгдаль, Андерс; Хеймдал, Джимми; Увдал, Пер (август 2017 г.). «Молекулярные признаки ископаемых листьев предоставляют неожиданные новые доказательства взаимоотношений вымерших растений» . Экология и эволюция природы . 1 (8): 1093–1099. Бибкод : 2017NatEE...1.1093V . дои : 10.1038/s41559-017-0224-5 . ISSN 2397-334X . ПМИД 29046567 . S2CID 3604369 .
- ^ Койро, Марио; Аллио, Реми; Мазет, Натан; Сейфулла, Лейла Дж.; Кондамин, Фабьен Л. (11 июня 2023 г.). «Согласование окаменелостей с филогениями раскрывает происхождение и макроэволюционные процессы, объясняющие глобальное биоразнообразие саговников» . Новый фитолог . 240 (4): 1616–1635. дои : 10.1111/nph.19010 . ISSN 0028-646X . ПМЦ 10953041 . ПМИД 37302411 . S2CID 259137975 .
- ^ Бейтман, Ричард М. (01 января 2020 г.). Орт, Дональд (ред.). «Охота на Снарка: ошибочные поиски мифических покрытосеменных юрского периода» . Журнал экспериментальной ботаники . 71 (1): 22–35. дои : 10.1093/jxb/erz411 . ISSN 0022-0957 . ПМИД 31538196 .
- ^ Койро, Марио; Робертс, Эмили А.; Хофманн, Криста-Ч.; Сейфулла, Лейла Дж. (14 декабря 2022 г.). «Обрезка длинных ветвей: согласие как путь к раскрытию эволюционной истории Gnetales» . Границы экологии и эволюции . 10 : 1082639. дои : 10.3389/fevo.2022.1082639 . ISSN 2296-701X .
- ^ Эльгорриага, Андрес; Эскапа, Игнатий Х.; Кунео, Н. Рубен (2 сентября 2019 г.). «Caytoniales Южного полушария: растительные и репродуктивные остатки из трапиальной формации Лонко (нижняя юра), Патагония» . Журнал систематической палеонтологии . 17 (17): 1477–1495. Бибкод : 2019JSPal..17.1477E . дои : 10.1080/14772019.2018.1535456 . ISSN 1477-2019 . S2CID 92287804 .
- ^ Слодоник, Мириам; Хилл, Роберт С.; Маклафлин, Стивен (октябрь 2023 г.). «Komlopteris: устойчивая линия посттриасовых користоспермов в Гондване» . Обзор палеоботаники и палинологии . 317 : 104950. Бибкод : 2023RPaPa.31704950S . дои : 10.1016/j.revpalbo.2023.104950 . S2CID 260015702 .
- ^ Перейти обратно: а б Кустачер, Эвелин; Вишер, Хэнк; ван Конийненбург-ван Циттерт, Йоханна Х.А. (01 сентября 2019 г.). «Существовали ли чекановские уже в поздней перми?» . ПалЗ . 93 (3): 465–477. Бибкод : 2019PalZ... 93..465K дои : 10.1007/ s12542-019-00468-9 ISSN 1867-6812 . S2CID 199473893 .
- ^ Тейлор, Т. (2009), «Гимноспермы с неясным родством» , «Биология и эволюция ископаемых растений » , Elsevier, стр. 757–785, doi : 10.1016/b978-0-12-373972-8.00019-x , ISBN 978-0-12-373972-8 , получено 13 декабря 2020 г.
- ^ Сунь, Чуньлинь; Ли, Юньфэн; Дилчер, Дэвид Л.; Ван, Хуншань; Ли, Тао; На, Юлин; Ван, Аньпин (ноябрь 2015 г.). «Вводный отчет о биоразнообразии среднеюрского Phoenicopsis (Czekanowskiales) из бассейна Ордос, Китай» . Научный вестник . 60 (21): 1858–1865. Бибкод : 2015SciBu..60.1858S . дои : 10.1007/s11434-015-0904-y . S2CID 140617907 .
- ^ Паттемор, Дж., Ригби, Дж. Ф. и Плейфорд, Г., 2015. Триасово-юрские птеридоспермы Австралазии: видообразование, разнообразие и упадок. Boletín Geológico y Minero, 126 (4): 689–722.
- ^ Ског, Джудит Э. (апрель 2001 г.). «Биогеография мезозойских лептоспорангиатных папоротников, родственных современным папоротникам» . Бриттония . 53 (2): 236–269. Бибкод : 2001Britt..53..236S . дои : 10.1007/bf02812701 . ISSN 0007-196X . S2CID 42781830 .
- ^ Тянь, Нин; Ван, Ён-Донг; Чжан, Ву; Чжэн, Шао-Линь; Чжу, Чжи-Пэн; Лю, Чжун-Цзянь (01 марта 2018 г.). «Перминерализованные осмундовые и глейхеновые папоротники из юры Внутренней Монголии, Северо-Восточный Китай» . Палеобиоразнообразие и палеосреда . 98 (1): 165–176. Бибкод : 2018ПдПе...98..165Т . дои : 10.1007/s12549-017-0313-0 . ISSN 1867-1608 . S2CID 134149095 .
- ^ Регаладо, Ледис; Шмидт, Александр Р.; Мюллер, Патрик; Нидермайер, Лиза; Крингс, Майкл; Шнайдер, Харальд (июль 2019 г.). «Heinrichsia cheilanthoides gen. et sp. nov., ископаемый папоротник семейства Pteridaceae (Polypodiales) из меловых янтарных лесов Мьянмы» . Журнал систематики и эволюции . 57 (4): 329–338. дои : 10.1111/jse.12514 . ISSN 1674-4918 . S2CID 182754946 .
- ^ Ли, Чуньсян; Мяо, Синьюань; Чжан, Ли-Бин; Ма, Джунье; Хао, Цзяшэн (январь 2020 г.). «Переоценка систематического положения юрско-раннемелового рода папоротников Coniopteris» . Меловые исследования . 105 : 104136. Бибкод : 2020CrRes.10504136L . дои : 10.1016/j.cretres.2019.04.007 . S2CID 146355798 .
- ^ Коралл, Петра; Прайер, Кэтлин М. (февраль 2014 г.). Пармакелис, Аристейдис (ред.). «Глобальная биогеография чешуйчатых древовидных папоротников (Cyatheaceae): свидетельства викариантности Гондваны и ограниченного трансокеанического распространения» . Журнал биогеографии . 41 (2): 402–413. Бибкод : 2014JBiog..41..402K . дои : 10.1111/jbi.12222 . ISSN 0305-0270 . ПМЦ 4238398 . ПМИД 25435648 .
- ^ Аксмит, Брайан Дж.; Крингс, Майкл; Тейлор, Томас Н. (сентябрь 2001 г.). «Кленчатый папоротник из верхнего триаса Северной Каролины (США)» . Американский журнал ботаники . 88 (9): 1558–1567. дои : 10.2307/3558399 . ISSN 0002-9122 . JSTOR 3558399 . ПМИД 21669688 .
- ^ Эльгорриага, Андрес; Эскапа, Игнасио Х.; Бомфлер, Бенджамин; Кунео, Рубен; Оттоне, Эдуардо Г. (февраль 2015 г.). «Реконструкция и филогенетическое значение нового вида Equisetum Linnaeus из нижней юры Серро-Байо (провинция Чубут, Аргентина)» . Амегинян . 52 (1): 135–152. дои : 10.5710/AMGH.15.09.2014.2758 . hdl : 11336/66623 . ISSN 0002-7014 . S2CID 6134534 .
- ^ Gould, RE 1968. Морфология Equisetum Laterale Phillips, 1829 и E. bryanii sp. ноябрь из мезозоя юго-восточного Квинсленда. Австралийский журнал ботаники 16: 153–176.
- ^ Перейти обратно: а б Эльгорриага, Андрес; Эскапа, Игнатий Х.; Ротвелл, Гар В.; Томеску, Александр М.Ф.; Рубен Кунео, Н. (август 2018 г.). «Происхождение Equisetum: эволюция хвощей (Equisetales) внутри основной клады эуфиллофитов Sphenopsida» . Американский журнал ботаники . 105 (8): 1286–1303. дои : 10.1002/ajb2.1125 . ПМИД 30025163 .
- ^ Ченнинг, Алан; Замунер, Альба; Эдвардс, Дайанна; Гвидо, Диего (2011). «Equisetum Thermale sp. nov. (Equisetales) из юрского месторождения горячих источников Сан-Агустин, Патагония: анатомия, палеоэкология и предполагаемая палеоэкофизиология» . Американский журнал ботаники . 98 (4): 680–697. дои : 10.3732/ajb.1000211 . hdl : 11336/95234 . ISSN 1537-2197 . ПМИД 21613167 .
- ^ Вуд, Дэниел; Беснар, Гийом; Бирлинг, Дэвид Дж.; Осборн, Колин П.; Кристин, Паскаль-Антуан (18 июня 2020 г.). «Филогеномика указывает на «живое ископаемое» Isoetes, разнообразное в кайнозое» . ПЛОС ОДИН . 15 (6): e0227525. Бибкод : 2020PLoSO..1527525W . дои : 10.1371/journal.pone.0227525 . ISSN 1932-6203 . ПМЦ 7302493 . ПМИД 32555586 .
- ^ Перейти обратно: а б Мамонтов Юрий С.; Игнатов, Михаил С. (июль 2019 г.). «Как полагаться на недостоверное: Примеры из мезозойских мохообразных Забайкалья» . Журнал систематики и эволюции . 57 (4): 339–360. дои : 10.1111/jse.12483 . ISSN 1674-4918 . S2CID 92268163 .
- ^ Биппус, Александр К.; Саворетти, Адольфина; Эскапа, Игнасио Х.; Гарсиа-Массини, Хуан; Гвидо, Диего (октябрь 2019 г.). «Heinrichsiella patagonica gen. et sp. nov.: перминерализованный акрокарпный мох из юры Патагонии» . Международный журнал наук о растениях . 180 (8): 882–891. дои : 10.1086/704832 . ISSN 1058-5893 . S2CID 202859471 .
- ^ Ли, Жуюнь; Ли, Сяоцян; Дэн, Шэнхуэй; Вс, Байниан (август 2020 г.). «Морфология и микроструктура Pellites hamiensis nov. sp., среднеюрского печеночника из северо-западного Китая и его эволюционное значение» . Геобиос . 62 : 23–29. Бибкод : 2020Geobi..62...23L . doi : 10.1016/j.geobios.2020.07.003 . S2CID 225500594 .
- ^ Ли, Жуй-Юнь; Ван, Сюэ-лянь; Чен, Цзин-Вэй; Дэн, Шэн-Хуэй; Ван, Цзы-Си; Донг, Цзюнь-Линг; Сунь, Бай-Нянь (июнь 2016 г.). «Новый слоевищ печеночника: Pallaviciniites sandaolingensis sp. nov. из средней юры бассейна Турфан-Хами, северо-запад Китая» . ПалЗ . 90 (2): 389–397. Бибкод : 2016PalZ...90..389L . дои : 10.1007/s12542-016-0299-3 . ISSN 0031-0220 . S2CID 131295547 .
- ^ Ли, Жуюнь; Ли, Сяоцян; Ван, Хуншань; Солнце, Байниан (2019). «Ricciopsis sandaolingensis sp. nov., новый ископаемый мохообразный из среднеюрской формации Сишаньяо в бассейне Турфан-Хами, Синьцзян, Северо-Западный Китай» . Электронная палеонтология . 22 (2). дои : 10.26879/917 . ISSN 1094-8074 .
- ^ Аллен, Бетани Дж.; Стаббс, Томас Л.; Бентон, Майкл Дж.; Путтик, Марк Н. (март 2019 г.). Маннион, Филип (ред.). «Селективность вымирания архозавроморфов во время триасово-юрского массового вымирания» . Палеонтология . 62 (2): 211–224. Бибкод : 2019Palgy..62..211A . дои : 10.1111/пала.12399 . hdl : 1983/e3fc2e40-c849-42ed-99fe-ea17fc26b2ec . S2CID 55009185 .
- ^ Толягич, Оля; Батлер, Ричард Дж. (23 июня 2013 г.). «Триасово-юрское массовое вымирание как триггер мезозойской радиации крокодиломорфов» . Письма по биологии . 9 (3): 20130095. doi : 10.1098/rsbl.2013.0095 . ISSN 1744-9561 . ПМК 3645043 . ПМИД 23536443 .
- ^ Мелстром, Киган М.; Ирмис, Рэндалл Б. (июль 2019 г.). «Повторяющаяся эволюция травоядных крокодилоформ в эпоху динозавров» . Современная биология . 29 (14): 2389–2395.e3. Бибкод : 2019CBio...29E2389M . дои : 10.1016/j.cub.2019.05.076 . ПМИД 31257139 . S2CID 195699188 .
- ^ Перейти обратно: а б Стаббс, Томас Л.; Пирс, Стефани Э.; Эльслер, Армин; Андерсон, Филип С.Л.; Рэйфилд, Эмили Дж.; Бентон, Майкл Дж. (31 марта 2021 г.). «Экологические возможности, взлет и падение эволюционных инноваций крокодиломорфов» . Труды Королевского общества B: Биологические науки . 288 (1947): 20210069. doi : 10.1098/rspb.2021.0069 . ПМК 8059953 . ПМИД 33757349 . S2CID 232326789 .
- ^ Шпиндлер, Фредерик; Лауэр, Рене; Тишлингер, Гельмут; Маузер, Матиас (5 июля 2021 г.). «Покровы пелагических крокодиломорфов (Thalattosuria: Metriorhynchidae)» . Электронная палеонтология . 24 (2): 1–41. дои : 10.26879/1099 . ISSN 1094-8074 .
- ^ Ирмис, Рэндалл Б.; Несбитт, Стерлинг Дж.; Сьюс, Ханс-Дитер (2013). «Ранние крокодиломорфы» . Геологическое общество, Лондон, специальные публикации . 379 (1): 275–302. Бибкод : 2013GSLSP.379..275I . дои : 10.1144/SP379.24 . ISSN 0305-8719 . S2CID 219190410 .
- ^ Перейти обратно: а б Уилберг, Эрик В.; Тернер, Алан Х.; Брошу, Кристофер А. (24 января 2019 г.). «Эволюционная структура и время основных изменений среды обитания Crocodylomorpha» . Научные отчеты . 9 (1): 514. Бибкод : 2019NatSR...9..514W . дои : 10.1038/s41598-018-36795-1 . ISSN 2045-2322 . ПМК 6346023 . ПМИД 30679529 .
- ^ От Сассо, К.; Пасини, Г.; Флери, Г.; Магануко, С. (2017). « Razanandrongobe sakalavae , гигантский мезоэвкрокодил из средней юры Мадагаскара, является старейшим из известных нотозухов » ПерДж . 5 : е3 дои : 10.7717/peerj.3481 . ПМК 5499610 . ПМИД 28690926 .
- ^ Перейти обратно: а б Джойс, Уолтер Г. (апрель 2017 г.). «Обзор ископаемых летописей базальных мезозойских черепах». Бюллетень Музея естественной истории Пибоди . 58 (1): 65–113. дои : 10.3374/014.058.0105 . ISSN 0079-032X . S2CID 54982901 .
- ^ Стерли, Джулиана; де ла Фуэнте, Марсело С.; Ружье, Гильермо В. (04 июля 2018 г.). «Новые останки Condorchelys antiqua (Testudinata) из ранней-средней юры Патагонии: анатомия, филогения и педоморфоз в ранней эволюции черепах» . Журнал палеонтологии позвоночных . 38 (4): (1)–(17). Бибкод : 2018JVPal..38....1S . дои : 10.1080/02724634.2018.1480112 . hdl : 11336/99525 . ISSN 0272-4634 . S2CID 109556104 .
- ^ Салливан, Патрик М.; Джойс, Уолтер Г. (август 2017 г.). «Анатомия панциря и таза позднеюрской черепахи Platychelys oberndorferi на основе материалов из Золотурна, Швейцария» . Швейцарский журнал палеонтологии . 136 (2): 323–343. Бибкод : 2017SwJP..136..323S . дои : 10.1007/s13358-017-0136-7 . ISSN 1664-2376 . S2CID 90587841 .
- ^ Эверс, Сережа В.; Бенсон, Роджер Б.Дж. (январь 2019 г.). Смит, Эндрю (ред.). «Новая филогенетическая гипотеза черепах, имеющая значение для времени и количества эволюционных переходов к морскому образу жизни в группе» . Палеонтология . 62 (1): 93–134. Бибкод : 2019Palgy..62...93E . дои : 10.1111/пала.12384 . S2CID 134736808 .
- ^ Анкетен, Жереми; Пюнтенер, Кристиан; Джойс, Уолтер Г. (октябрь 2017 г.). «Обзор ископаемых записей черепах клады Thalassochelydia» . Бюллетень Музея естественной истории Пибоди . 58 (2): 317–369. дои : 10.3374/014.058.0205 . ISSN 0079-032X . S2CID 31091127 .
- ^ Перейти обратно: а б Эванс, Сьюзен Э.; Джонс, Марк Э.Х. (2010), «Происхождение, ранняя история и диверсификация лепидозавроморфных рептилий» , Новые аспекты мезозойского биоразнообразия , Конспекты лекций по наукам о Земле, том. 132, Берлин, Гейдельберг: Springer Berlin Heidelberg, стр. 27–44, Bibcode : 2010LNES..132...27E , doi : 10.1007/978-3-642-10311-7_2 , ISBN 978-3-642-10310-0 , получено 7 января 2021 г.
- ^ Браунштейн, Чейз Д.; Мейер, Далтон Л.; Фаббри, Мэтью; Бхуллар, Бхарт-Анджан С.; Готье, Джеймс А. (29 ноября 2022 г.). «Эволюционное происхождение длительно существующей чешуйчатой радиации» . Природные коммуникации . 13 (1): 7087. Бибкод : 2022NatCo..13.7087B . дои : 10.1038/ s41467-022-34217-5 ISSN 2041-1723 . ПМЦ 9708687 . ПМИД 36446761 .
- ^ Эррера-Флорес, Хорхе А.; Стаббс, Томас Л.; Бентон, Майкл Дж. (2017). «Макроэволюционные закономерности ринхоцефалии: является ли туатара (Sphenodon punctatus) живым ископаемым?» . Палеонтология . 60 (3): 319–328. Бибкод : 2017Palgy..60..319H . дои : 10.1111/пала.12284 . ISSN 1475-4983 .
- ^ Бербринк, Фрэнк Т; Грацциотин, Фелипе Дж; Пайрон, Р. Александр; Кандалл, Дэвид; Доннеллан, Стив; Ирландка, Фрэнсис; Кио, Дж. Скотт; Краус, Фред; Мерфи, Роберт В.; Нунан, Брайс; Раксуорси, Кристофер Дж (01 мая 2020 г.). Томсон, Роберт (ред.). «Опрос данных геномного масштаба чешуйчатых (ящериц, змей и амфисбенов) не подтверждает наличие ключевых традиционных морфологических отношений» . Систематическая биология . 69 (3): 502–520. дои : 10.1093/sysbio/syz062 . ISSN 1063-5157 . ПМИД 31550008 .
- ^ Клири, Терри Дж.; Бенсон, Роджер Б.Дж.; Эванс, Сьюзен Э.; Барретт, Пол М. (21 марта 2018 г.). «Разнообразие лепидозавров в мезозое – палеогене: потенциальная роль ошибок выборки и экологических факторов» . Королевское общество открытой науки . 5 (3): 171830. Бибкод : 2018RSOS....571830C . дои : 10.1098/rsos.171830 . ПМЦ 5882712 . ПМИД 29657788 .
- ^ Эванс, SE (1998). «Кроновая группа ящериц (Reptilia, Squamata) из средней юры Британских островов» Палеонтография, Abteilung A 250 (4–6): 123–154. Бибкод : 1998PalAA.250..123E . дои : 10.1127/пала/250/1998/123 . S2CID 246932992 .
- ^ Донг, Липин; Ван, Юань; Моу, Лицзе; Чжан, Гуозе; Эванс, Сьюзен Э. (13 сентября 2019 г.). «Новая юрская ящерица из Китая» . Геодиверситас . 41 (16): 623. doi : 10.5252/geodiversitas2019v41a16 . ISSN 1280-9659 . S2CID 204256127 .
- ^ Симойнс, Тьяго Р.; Колдуэлл, Майкл В.; Нидам, Рэндалл Л.; Хименес-Уидобро, Паулина (сентябрь 2016 г.). «Остеология, филогения и функциональная морфология двух юрских видов ящериц и ранняя эволюция скансориальности у гекконов». Зоологический журнал Линнеевского общества . дои : 10.1111/zoj.12487 .
- ^ Даза, доктор медицинских наук; Бауэр, AM; Стэнли, Эл.; Болет, А.; Диксон, Б.; Лосос, Дж. Б. (01 ноября 2018 г.). «Загадочная миниатюрная и ослабленная целая ящерица из среднемелового янтаря Мьянмы» . Бревиора . 563 (1): 1. дои : 10.3099/MCZ49.1 . hdl : 1983/0955fcf4-a32a-4498-b920-1421dcea67de . ISSN 0006-9698 . S2CID 91589111 .
- ^ Гриффитс, Элизабет Ф.; Форд, Дэвид П.; Бенсон, Роджер Б.Дж.; Эванс, Сьюзен Э. (сентябрь 2021 г.). Рута, Марчелло (ред.). «Новые сведения о юрском лепидозавроморфе Marmoretta oxoniensis» . Статьи по палеонтологии . 7 (4): 2255–2278. Бибкод : 2021PPal....7.2255G . дои : 10.1002/spp2.1400 . ISSN 2056-2799 . S2CID 239140732 .
- ^ Мацумото, Р.; Эванс, SE (2010). «Хористодеры и пресноводные комплексы Лавразии» . Журнал иберийской геологии . 36 (2): 253–274. Бибкод : 2010JIbG...36..253M . дои : 10.5209/rev_JIGE.2010.v36.n2.11 . ISSN 1698-6180 .
- ^ Мацумото, Рёко; Донг, Липин; Ван, Юань; Эванс, Сьюзен Э. (18 июня 2019 г.). «Первая запись почти полной хористодеры (Reptilia: Diapsida) из верхней юры провинции Хэбэй, Китайская Народная Республика» . Журнал систематической палеонтологии . 17 (12): 1031–1048. Бибкод : 2019JSPal..17.1031M . дои : 10.1080/14772019.2018.1494220 . ISSN 1477-2019 . S2CID 92421503 .
- ^ Перейти обратно: а б Торн, премьер-министр; Рута, М.; Бентон, MJ (17 мая 2011 г.). «Перезагрузка эволюции морских рептилий на границе триаса и юры» . Труды Национальной академии наук . 108 (20): 8339–8344. Бибкод : 2011PNAS..108.8339T . дои : 10.1073/pnas.1018959108 . ISSN 0027-8424 . ПМК 3100925 . ПМИД 21536898 .
- ^ Перейти обратно: а б Мун, Бенджамин К.; Стаббс, Томас Л. (13 февраля 2020 г.). «Ранние высокие темпы и неравномерность эволюции ихтиозавров» . Коммуникационная биология . 3 (1): 68. дои : 10.1038/s42003-020-0779-6 . ISSN 2399-3642 . ПМК 7018711 . ПМИД 32054967 .
- ^ Перейти обратно: а б с д и Фишер, Валентин; Вайс, Роберт; Туи, Бен (22 февраля 2021 г.). «Уточнение круговорота морских рептилий на этапе перехода ранней и средней юры» . ПерДж . 9 : е10647. дои : 10.7717/peerj.10647 . ISSN 2167-8359 . ПМК 7906043 . ПМИД 33665003 .
- ^ Винтрих, Таня; Хаяси, Сёдзи; Уссей, Александра; Накадзима, Ясухиса; Сандер, П. Мартин (01 декабря 2017 г.). «Триасовый скелет плезиозавра и гистология костей свидетельствуют об эволюции уникального строения тела» . Достижения науки . 3 (12): e1701144. Бибкод : 2017SciA....3E1144W . дои : 10.1126/sciadv.1701144 . ISSN 2375-2548 . ПМК 5729018 . ПМИД 29242826 .
- ^ Бенсон, Роджер Б.Дж.; Эванс, Марк; Дракенмиллер, Патрик С. (16 марта 2012 г.). «Высокое разнообразие, низкое неравенство и небольшой размер тела плезиозавров (Reptilia, Sauropterygia) на границе триаса и юры» . ПЛОС ОДИН . 7 (3): e31838. Бибкод : 2012PLoSO...731838B . дои : 10.1371/journal.pone.0031838 . ISSN 1932-6203 . ПМК 3306369 . ПМИД 22438869 .
- ^ О'Киф, Ф. Робин (2002). «Эволюция морфотипов плезиозавров и плиозавров у Plesiosauria (Reptilia: Sauropterygia)» . Палеобиология . 28 (1): 101–112. Бибкод : 2002Pbio...28..101O . doi : 10.1666/0094-8373(2002)028<0101:TEOPAP>2.0.CO;2 . ISSN 0094-8373 . S2CID 85753943 .
- ^ Бенсон, Роджер Б.Дж.; Эванс, Марк; Смит, Адам С.; Сассун, Джудит; Мур-Фэй, Скотт; Кетчам, Хилари Ф.; Форрест, Ричард (31 мая 2013 г.). «Гигантский череп плиозавра из поздней юры Англии» . ПЛОС ОДИН . 8 (5): e65989. Бибкод : 2013PLoSO...865989B . дои : 10.1371/journal.pone.0065989 . ISSN 1932-6203 . ПМК 3669260 . ПМИД 23741520 .
- ^ Гао, Тин; Ли, Да-Цин; Ли, Лун-Фэн; Ян, Цзин-Тао (13 августа 2019 г.). «Первая запись пресноводного плезиозавра из средней юры Ганьсу, северо-запад Китая, с ее значением для местной палеобиогеографии» . Журнал палеогеографии . 8 (1): 27. Бибкод : 2019JPalG...8...27G . дои : 10.1186/s42501-019-0043-5 . ISSN 2524-4507 . S2CID 199547716 .
- ^ Кир, Бенджамин П. (2 августа 2012 г.). «Пересмотр юрских плезиозавров Австралии» . Палеонтология . 55 (5): 1125–1138. Бибкод : 2012Palgy..55.1125K . дои : 10.1111/j.1475-4983.2012.01183.x .
- ^ О'Салливан, Майкл; Мартилл, Дэвид М. (17 ноября 2017 г.). «Таксономия и систематика Parapsicephalus purdoni (Reptilia: Pterosauria) из нижнеюрской аргиллитовой формации Уитби, Уитби, Великобритания» . Историческая биология . 29 (8): 1009–1018. Бибкод : 2017HBio...29.1009O . дои : 10.1080/08912963.2017.1281919 . ISSN 0891-2963 . S2CID 132532024 .
- ^ Перейти обратно: а б с д Бествик, Иордания; Анвин, Дэвид М.; Батлер, Ричард Дж.; Хендерсон, Дональд М.; Пурнелл, Марк А. (ноябрь 2018 г.). «Диетические гипотезы птерозавров: обзор идей и подходов: Диетические гипотезы птерозавров» . Биологические обзоры . 93 (4): 2021–2048. дои : 10.1111/brv.12431 . ПМК 6849529 . ПМИД 29877021 .
- ^ Брусатте, Стивен Л; Бентон, Майкл Дж; Рута, Марчелло; Ллойд, Грэм Т. (23 декабря 2008 г.). «Первые 50 миллионов лет эволюции динозавров: макроэволюционная закономерность и морфологическое неравенство» . Письма по биологии . 4 (6): 733–736. дои : 10.1098/rsbl.2008.0441 . ПМК 2614175 . ПМИД 18812311 .
- ^ Брусатте, СЛ; Бентон, MJ; Рута, М.; Ллойд, GT (12 сентября 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционном излучении динозавров» (PDF) . Наука . 321 (5895): 1485–88. Бибкод : 2008Sci...321.1485B . дои : 10.1126/science.1161833 . hdl : 20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b . ПМИД 18787166 . S2CID 13393888 . Архивировано из оригинала (PDF) 24 июня 2014 г. Проверено 14 января 2012 г.
- ^ Темп Мюллер, Родриго; Аугусто Претто, Флавио; Кербер, Леонардо; Сильва-Невес, Эдуардо; Диаш-да-Сильва, Сержио (28 марта 2018 г.). «Комментарий к статье «Недостающее звено динозавра? Чилозавр и ранняя эволюция птицетазовых динозавров» » . Письма по биологии . 14 (3): 20170581. doi : 10.1098/rsbl.2017.0581 . ISSN 1744-9561 . ПМЦ 5897605 . ПМИД 29593074 .
- ^ Занер, Мэрион; Бринкманн, Винанд (август 2019 г.). «Триасовый теропод аверострановой линии из Швейцарии и ранняя эволюция динозавров» . Экология и эволюция природы . 3 (8): 1146–1152. Бибкод : 2019NatEE...3.1146Z . дои : 10.1038/s41559-019-0941-z . ISSN 2397-334X . ПМК 6669044 . ПМИД 31285577 .
- ^ Сассо, Криштиану Даль; Магануко, Симоне; Кау, Андреа (19 декабря 2018 г.). «Старейший цератозавр (Dinosauria: Theropoda) из нижней юры Италии проливает свет на эволюцию трехпалой руки птиц» . ПерДж . 6 : е5976. дои : 10.7717/peerj.5976 . ISSN 2167-8359 . ПМК 6304160 . ПМИД 30588396 .
- ^ Ван, Шуо; Штиглер, Йозеф; Амио, Ромен; Ван, Сюй; Ду, Го-хао; Кларк, Джеймс М.; Сюй, Син (январь 2017 г.). «Чрезвычайные онтогенетические изменения у цератозаврового теропода» . Современная биология . 27 (1): 144–148. Бибкод : 2017CBio...27..144W . дои : 10.1016/j.cub.2016.10.043 . ПМИД 28017609 . S2CID 441498 .
- ^ Занно, Линдси Э.; Маковицкий, Питер Дж. (4 января 2011 г.). «Экоморфология травоядных и закономерности специализации в эволюции тероподовых динозавров» . Труды Национальной академии наук . 108 (1): 232–237. Бибкод : 2011PNAS..108..232Z . дои : 10.1073/pnas.1011924108 . ISSN 0027-8424 . ПМК 3017133 . ПМИД 21173263 .
- ^ Перейти обратно: а б Раухут, Оливер В.М.; Пол, Диего (11 декабря 2019 г.). «Вероятный базальный аллозавроид из ранней среднеюрской формации Каньядон Асфальто в Аргентине подчеркивает филогенетическую неопределенность столбнячных тероподных динозавров» . Научные отчеты . 9 (1): 18826. Бибкод : 2019NatSR...918826R . дои : 10.1038/s41598-019-53672-7 . ISSN 2045-2322 . ПМК 6906444 . ПМИД 31827108 .
- ^ Бенсон, RBJ (2010). «Описание Megalosaurus bucklandii (Dinosauria: Theropoda) из бата Великобритании и взаимоотношения теропод средней юры» . Зоологический журнал Линнеевского общества . 158 (4): 882–935. дои : 10.1111/j.1096-3642.2009.00569.x .
- ^ Раухут, Оливер В.М.; Милнер, Анджела С.; Мур-Фэй, Скотт (2010). «Краниальная остеология и филогенетическое положение динозавра-теропода Proceratosaurus bradleyi (Woodward, 1910) из средней юры Англии» . Зоологический журнал Линнеевского общества . 158 : 155–195. дои : 10.1111/j.1096-3642.2009.00591.x .
- ^ Цинь З., Кларк Дж., Шуаньер Дж. и Сюй X. (2019). Новый альваресзавровый теропод из верхнеюрской формации Шишугоу на западе Китая. Научные отчеты, 9: 11727. два : 10.1038/s41598-019-48148-7
- ^ Аньолин, Федерико Л.; Лу, Джун-Чанг; Кундрат, Мартин; Сюй, Ли (02 июня 2021 г.). «Остеология альваресзаврид: новые данные по краниальной анатомии» . Историческая биология . 34 (3): 443–452. дои : 10.1080/08912963.2021.1929203 . ISSN 0891-2963 . S2CID 236221732 .
- ^ Ван, Мин; О'Коннор, Цзинмай К.; Сюй, Син; Чжоу, Чжунхэ (май 2019 г.). «Новый юрский скансориоптеригид и потеря перепончатых крыльев у тероподовых динозавров» . Природа . 569 (7755): 256–259. Бибкод : 2019Natur.569..256W . дои : 10.1038/s41586-019-1137-z . ISSN 1476-4687 . ПМИД 31068719 . S2CID 148571099 .
- ^ Хартман, Скотт; Мортимер, Микки; Уол, Уильям Р.; Ломакс, Дин Р.; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ISSN 2167-8359 . ПМК 6626525 . ПМИД 31333906 .
- ^ Раухут, Оливер В.М.; Фот, Кристиан (2020), Фот, Кристиан; Раухут, Оливер В.М. (ред.), «Происхождение птиц: современный консенсус, противоречия и появление перьев» , «Эволюция перьев: от их происхождения до наших дней» , «Увлекательные науки о жизни», Cham: Springer International Publishing, стр. . 27–45, номер домена : 10.1007/978-3-030-27223-4_3 , ISBN. 978-3-030-27223-4 , S2CID 216372010 , получено 5 января 2021 г.
- ^ Норман, Дэвид Б. (01 января 2021 г.). «Scelidosaurus harrisonii (Dinosauria: Ornithischia) из ранней юры Дорсета, Англия: биология и филогенетические связи» . Зоологический журнал Линнеевского общества . 191 (1): 1–86. doi : 10.1093/zoolinnean/zlaa061 . ISSN 0024-4082 .
- ^ Годфруа, Паскаль; Синица, София М.; Чинкотта, Од; Макнамара, Мария Э.; Решетова Светлана А.; Дуайи, Даниэль (2020), Фот, Кристиан; Раухут, Оливер В.М. (ред.), «Покровные структуры Kulindadromeus zabaikalicus, базального неорнитишского динозавра из юрского периода Сибири» , «Эволюция перьев: от их происхождения до наших дней» , «Увлекательные науки о жизни», Cham: Springer International Publishing, стр. . 47–65, номер домена : 10.1007/978-3-030-27223-4_4 , ISBN. 978-3-030-27223-4 , S2CID 216261986 , получено 5 января 2021 г.
- ^ Макдональд, Эндрю Т. (22 мая 2012 г.). Фарке, Эндрю А. (ред.). «Филогения базальных игуанодонтов (Dinosauria: Ornithischia): обновленная информация» . ПЛОС ОДИН . 7 (5): e36745. Бибкод : 2012PLoSO...736745M . дои : 10.1371/journal.pone.0036745 . ISSN 1932-6203 . ПМЦ 3358318 . ПМИД 22629328 .
- ^ Хан, Фэнлу; Форстер, Кэтрин А.; Кларк, Джеймс М.; Сюй, Син (09 декабря 2015 г.). «Новый таксон базальных цератопсов из Китая и ранняя эволюция цератопсов» . ПЛОС ОДИН . 10 (12): e0143369. Бибкод : 2015PLoSO..1043369H . дои : 10.1371/journal.pone.0143369 . ISSN 1932-6203 . ПМК 4674058 . ПМИД 26649770 .
- ^ Перейти обратно: а б с Пол, Д.; Рамезани Дж.; Гомес, К.; Карбаллидо, JL; Карабахал, А. Паулина; Раухут, OWM; Эскапа, Айдахо; Кунео, Северная Каролина (25 ноября 2020 г.). «Вымирание травоядных динозавров связано с глобальным потеплением в начале юрского периода» . Труды Королевского общества B: Биологические науки . 287 (1939): 20202310. doi : 10.1098/rspb.2020.2310 . ISSN 0962-8452 . ПМЦ 7739499 . ПМИД 33203331 .
- ^ Сандер, П. Мартин; Кристиан, Андреас; Клаусс, Маркус; Фехнер, Регина; Ну и дела, Кэрол Т.; Грибелер, Ева-Мария; Гунга, Ханнс-Кристиан; Хуммель, Юрген; Мэллисон, Генрих; Перри, Стивен Ф.; Пройшофт, Хольгер (февраль 2011 г.). «Биология динозавров-зауроподов: эволюция гигантизма» . Биологические обзоры . 86 (1): 117–155. дои : 10.1111/j.1469-185X.2010.00137.x . ПМК 3045712 . ПМИД 21251189 .
- ^ Макфи, Блэр В.; Бенсон, Роджер Б.Дж.; Бота-Бринк, Дженнифер; Борди, Эмесе М.; Шуаньер, Иона Н. (8 октября 2018 г.). «Гигантский динозавр из древнейшей юры Южной Африки и переход к четвероногости у ранних зауроподоморфов» . Современная биология . 28 (19): 3143–3151.e7. Бибкод : 2018CBio...28E3143M . дои : 10.1016/j.cub.2018.07.063 . ПМИД 30270189 . S2CID 52890502 .
- ^ Вильетти, Пиа А.; Барретт, Пол М.; Бродерик, Тим Дж.; Муньиква, Дарлингтон; МакНивен, Роуэн; Бродерик, Люси; Чапель, Кимберли; Глинн, Дэйв; Эдвардс, Стив; Зондо, Мишель; Бродерик, Патрисия (январь 2018 г.). «Стратиграфия местности типа вулканодона и ее значение для региональных корреляций внутри супергруппы Кару» . Журнал африканских наук о Земле . 137 : 149–156. Бибкод : 2018JAfES.137..149В . дои : 10.1016/j.jafrearsci.2017.10.015 .
- ^ Ройо-Торрес, Рафаэль; Кобос, Альберто; Мочо, Питер; Алькала, Луис (01 января 2021 г.). «Происхождение и эволюция динозавров-туриазавров установлено с помощью нового экземпляра «розетты» из Испании» . Зоологический журнал Линнеевского общества . 191 (1): 201–227. doi : 10.1093/zoolinnean/zlaa091 . ISSN 0024-4082 .
- ^ Рен, Синь-Синь; Хуан, Цзянь-Донг; Ты, Хай-Лу (27 мая 2020 г.). «Второй маменчизавридный динозавр из средней юры Восточного Китая» . Историческая биология . 32 (5): 602–610. Бибкод : 2020HBio...32..602R . дои : 10.1080/08912963.2018.1515935 . ISSN 0891-2963 . S2CID 91927243 .
- ^ Рен, Синь-Синь; Цзян, Шан; Ван, Сюй-Ри; Пэн, Гуан-Чжао; Йе, Ён; Цзя, Лей; Ты, Хай-Лу (14 ноября 2022 г.). «Повторное исследование Dashanpusaurus dongi (Sauropoda: Macronaria) подтверждает глобальное распространение динозавров-неозауропод в начале средней юры» . Палеогеография, Палеоклиматология, Палеоэкология . 610 : 111318. doi : 10.1016/j.palaeo.2022.111318 . ISSN 0031-0182 .
- ^ Лукас, Спенсер Г. (2018), Таннер, Лоуренс Х. (редактор), «Наземные четвероногие позднего триаса: биостратиграфия, биохронология и биотические события» , Мир позднего триаса , Темы геобиологии, том. 46, Чам: Springer International Publishing, стр. 351–405, номер документа : 10.1007/978-3-319-68009-5_10 , ISBN. 978-3-319-68008-8 , получено 25 апреля 2021 г.
- ^ Аверьянов Александр О.; Мартин, Томас; Скутщас, Павел П.; Резвый, Антон С.; Бакиров, Изек А. (март 2008 г.). «Земноводные из среднеюрской Балабансай Свиты в Ферганской впадине, Киргизия (Центральная Азия) » Палеонтология . 51 (2): 471–485. Бибкод : 2008Palgy..51..471A . дои : 10.1111/j.1475-4983.2007.00748.x .
- ^ Уоррен, А.А.; Хатчинсон, Миннесота; Хилл, Дороти (13 сентября 1983 г.). «Последний лабиринтодонт? Новый брахиопоид (Amphibia, Temnospondyli) из ранней юрской вечнозеленой формации Квинсленда, Австралия» . Философские труды Лондонского королевского общества. Б. Биологические науки . 303 (1113): 1–62. Бибкод : 1983RSPTB.303....1W . дои : 10.1098/rstb.1983.0080 .
- ^ Рейли, Стивен М.; Йоргенсен, Майкл Э. (февраль 2011 г.). «Эволюция прыжков у лягушек: морфологические доказательства базального двигательного состояния бесхвостых и радиации опорно-двигательных систем у бесхвостых кроновой группы». Журнал морфологии . 272 (2): 149–168. дои : 10.1002/jmor.10902 . ПМИД 21210487 . S2CID 14217777 .
- ^ Баэс, Ана Мария; Николи, Лаура (март 2008 г.). «Новый вид Notobatrachus (Amphibia, Salientia) из средней юры северо-западной Патагонии» . Журнал палеонтологии . 82 (2): 372–376. Бибкод : 2008JPal...82..372B . дои : 10.1666/06-117.1 . hdl : 11336/135748 . ISSN 0022-3360 . S2CID 130032431 .
- ^ Марьянович, Давид; Лорен, Мишель (04 июля 2014 г.). «Обновленное палеонтологическое древо лисамфибий с комментариями по анатомии саламандр юрской кроновой группы (Urodela)» . Историческая биология . 26 (4): 535–550. Бибкод : 2014HBio...26..535M . дои : 10.1080/08912963.2013.797972 . ISSN 0891-2963 . S2CID 84581331 .
- ^ Шох, Райнер Р.; Вернебург, Ральф; Фойгт, Себастьян (26 мая 2020 г.). «Триасовый ствол-саламандра из Кыргызстана и происхождение саламандр» . Труды Национальной академии наук . 117 (21): 11584–11588. Бибкод : 2020PNAS..11711584S . дои : 10.1073/pnas.2001424117 . ISSN 0027-8424 . ПМК 7261083 . ПМИД 32393623 .
- ^ Скутщас, Павел; Штейн, Коэн (апрель 2015 г.). «Гистология длинных костей стеблевой саламандры Kokartus Honorarius (Amphibia: Caudata) из средней юры Кыргызстана» . Журнал анатомии . 226 (4): 334–347. дои : 10.1111/joa.12281 . ПМЦ 4386933 . ПМИД 25682890 .
- ^ Цзя, Цзя; Гао, Кэ-Цинь (04 марта 2019 г.). «Новая стеблевая саламандра-гинобиид (Urodela, Cryptobranchoidea) из верхней юры (оксфорд) провинции Ляонин, Китай» . Журнал палеонтологии позвоночных . 39 (2): e1588285. Бибкод : 2019JVPal..39E8285J . дои : 10.1080/02724634.2019.1588285 . ISSN 0272-4634 . S2CID 164310171 .
- ^ Гао, К.-К.; Шубин, Нью-Хэмпшир (10 апреля 2012 г.). «Позднеюрский саламандроид из западного Ляонина, Китай» . Труды Национальной академии наук . 109 (15): 5767–5772. Бибкод : 2012PNAS..109.5767G . дои : 10.1073/pnas.1009828109 . ISSN 0027-8424 . ПМЦ 3326464 . ПМИД 22411790 .
- ^ Цзя, Цзя; Гао, Кэ-Цинь (4 мая 2016 г.). «Новый базальный саламандроид (амфибия, уродела) из поздней юры Цинлуна, провинция Хэбэй, Китай» . ПЛОС ОДИН . 11 (5): e0153834. Бибкод : 2016PLoSO..1153834J . дои : 10.1371/journal.pone.0153834 . ISSN 1932-6203 . ПМЦ 4856324 . ПМИД 27144770 .
- ^ Эванс, SE; Лалли, К.; Чуре, округ Колумбия; Старший, А.; Майзано, JA (2005). «Позднеюрская саламандра (Amphibia: Caudata) из формации Моррисон в Северной Америке» . Зоологический журнал Линнеевского общества . 143 (4): 599–616. дои : 10.1111/j.1096-3642.2005.00159.x .
- ^ Сантос, Родольфо Отавио; Лорен, Мишель; Захер, Хусам (3 ноября 2020 г.). «Обзор окаменелостей червяг (Lissamphibia: Gymnophionomorpha) с комментариями по их использованию для калибровки молекулярных деревьев времени» . Биологический журнал Линнеевского общества . 131 (4): 737–755. doi : 10.1093/biolinnean/blaa148 . ISSN 0024-4066 .
- ^ Хаддуми, Хамид; Аллен, Ронан; Меслоух, Саид; Метаис, Грегори; Монбарон, Мишель; Понс, Дениз; Ярость, Жан-Клод; Вулло, Ромен; Зухри, Самир (январь 2016 г.). «Гуэльб-эль-Ахмар (бат, годовая синклиналь, восточное Марокко): первая континентальная флора и фауна, включая млекопитающих из средней юры Африки» (PDF) . Гондваны Исследования 29 (1): 290–319. Бибкод : 2016GondR..29..290H . дои : 10.1016/j.gr.2014.12.004 . ISSN 1342-937X .
- ^ Перейти обратно: а б Ли, Майкл С.Ю.; Бек, Робин, доктор медицины (31 августа 2015 г.). «Эволюция млекопитающих: юрская искра» . Современная биология . 25 (17): Р759–Р761. Бибкод : 2015CBio...25.R759L . дои : 10.1016/j.cub.2015.07.008 . ПМИД 26325137 . S2CID 11088107 .
- ^ Джи, К.; Луо, З.-Х.; Юань, C.-X.; Табрум, Арканзас (2006). «Плавающая форма млекопитающих из средней юры и экоморфологическое разнообразие ранних млекопитающих» (PDF) . Наука . 311 (5, 764): 1, 123–1, 127. Бибкод : 2006Sci...311.1123J . дои : 10.1126/science.1123026 . ПМИД 16497926 . S2CID 46067702 .
- ^ Мэн, Цин-Цзинь; Гроссникл, Дэвид М.; Лю, Ди; Чжан, Юй-Гуан; Неандер, I апреля; Цзи, Цян; Ло, Чжэ-Си (август 2017 г.). «Новые планирующие млекопитающие юрского периода» . Природа . 548 (7667): 291–296. Бибкод : 2017Natur.548..291M . дои : 10.1038/nature23476 . ISSN 1476-4687 . ПМИД 28792929 . S2CID 205259206 .
- ^ Мэн, Дж.; Ху, Ю.-М.; Ван, Ю.-К.; Ван, X.-L.; Лизать. (2007). «Исправление: мезозойское планирующее млекопитающее из северо-восточного Китая». Nature 446 (7131): 102. Бибкод : 2007Natur.446Q.102M . дои : 10.1038/nature05639 .
- ^ Луо, З.-Х.; Вибл, младший (2005). «Позднеюрские роющие млекопитающие и диверсификация ранних млекопитающих». Наука . 308 (5718): 103–107. Бибкод : 2005Sci...308..103L . дои : 10.1126/science.1108875 . ISSN 0036-8075 . ПМИД 15802602 . S2CID 7031381 .
- ^ Ло, Чжэ-Си; Чифелли, Ричард Л.; Килан-Яворовска, Зофия (январь 2001 г.). «Двойное происхождение трибосфенических млекопитающих» . Природа . 409 (6816): 53–57. Бибкод : 2001Natur.409...53L . дои : 10.1038/35051023 . ISSN 0028-0836 . ПМИД 11343108 . S2CID 4342585 .
- ^ Мао, Фанъюань; Брюэр, Филиппа; Хукер, Джерри Дж.; Мэн, Джин (31 декабря 2022 г.). «Новые аллотериальные образцы из среднеюрского карьера Вудитон (Оксфордшир) и их значение для разнообразия и филогении харамииданов» . Журнал систематической палеонтологии . 20 (1): 1–37. дои : 10.1080/14772019.2022.2097021 . ISSN 1477-2019 .
- ^ Чжэ-Си Ло; Чонг-Си Юань; Цин-Цзинь Мэн; Цян Цзи (25 августа 2011 г.). «Юрское плацентарное млекопитающее и дивергенция сумчатых и плацентарных» (PDF) . Природа . 476 (7361): 442–445. Бибкод : 2011Natur.476..442L . дои : 10.1038/nature10291 . ПМИД 21866158 . S2CID 205225806 . Архивировано из оригинала (PDF) 10 ноября 2013 г. Электронный дополнительный материал.
- ^ Король, Бенедикт; Бек, Робин, доктор медицины (10 июня 2020 г.). «Тип датирования поддерживает новые решения спорных отношений между ранними млекопитающими» . Труды Королевского общества B: Биологические науки . 287 (1928): 20200943. doi : 10.1098/rspb.2020.0943 . ПМК 7341916 . ПМИД 32517606 .
- ^ Рута, Марчелло; Бота-Бринк, Дженнифер; Митчелл, Стивен А.; Бентон, Майкл Дж. (22 октября 2013 г.). «Радиация цинодонтов и план морфологического разнообразия млекопитающих» . Труды Королевского общества B: Биологические науки . 280 (1769): 20131865. doi : 10.1098/rspb.2013.1865 . ISSN 0962-8452 . ПМЦ 3768321 . ПМИД 23986112 .
- ^ Абдала, Фернандо; Гаэтано, Леандро К. (2018), Таннер, Лоуренс Х. (редактор), «Записи цинодонтов позднего триаса: время инноваций в линии млекопитающих» , Мир позднего триаса , Темы геобиологии, том. 46, Чам: Springer International Publishing, стр. 407–445, номер документа : 10.1007/978-3-319-68009-5_11 , ISBN. 978-3-319-68008-8 , получено 24 мая 2021 г.
- ^ Ду, Исин; Кьяри, Марко; Каради, Виктор; Никора, Альда; Оноуэ, Тецудзи; Палфи, Джозеф; Роги, Гвидо; Томимацу, Юки; Риго, Мануэль (апрель 2020 г.). «Асинхронное исчезновение конодонтов: новые ограничения на пограничных участках триаса и юры в Тетисе и Панталассе» . Обзоры наук о Земле . 203 : 103176. Бибкод : 2020ESRv..20303176D . doi : 10.1016/j.earscirev.2020.103176 . hdl : 11577/3338908 . S2CID 216173452 .
- ^ Гино, Самуэль; Гудеманд, Николя (декабрь 2020 г.). «Глобальные изменения климата являются причиной основных тенденций разнообразия конодонтов, но не их окончательной гибели» . Глобальные и планетарные изменения . 195 : 103325. Бибкод : 2020GPC...19503325G . дои : 10.1016/j.gloplacha.2020.103325 . S2CID 225005180 .
- ^ У, Фэйсян; Жанвье, Филипп; Чжан, Чи (31 октября 2023 г.). «Рост хищничества юрских миног» . Природные коммуникации . 14 (1): 6652. Бибкод : 2023NatCo..14.6652W . дои : 10.1038/s41467-023-42251-0 . ISSN 2041-1723 . ПМЦ 10618186 . ПМИД 37907522 .
- ^ Кемп, Энн; Кавин, Лайонел; Гино, Гийом (апрель 2017 г.). «Эволюционная история двоякодышащих рыб с новой филогенией постдевонских родов» . Палеогеография, Палеоклиматология, Палеоэкология . 471 : 209–219. Бибкод : 2017PPP...471..209K . дои : 10.1016/j.palaeo.2016.12.051 .
- ^ Браунштейн, Чейз Доран; Харрингтон, Ричард С; Рядом, Томас Дж. (июль 2023 г.). «Биогеография современных двоякодышащих рыб прослеживает распад Гондваны» . Журнал биогеографии . 50 (7): 1191–1198. Бибкод : 2023JBiog..50.1191B . дои : 10.1111/jbi.14609 . ISSN 0305-0270 .
- ^ Кавин, Лайонел; Купелло, Камила; Ябумото, Ёситака; Лео, Фрагосо; Дирси, Утумпорн; Брито, Пол М. (2019). «Филогения и история эволюции мавсониид целакантов» (PDF) . Бюллетень Музея естественной истории и истории человечества Китакюсю, серия А. 17 : 3–13.
- ^ Клемент, Гаэль (30 сентября 2005 г.). «Новый целакант (Actinistia, Sarcopterygii) из юры Франции и вопрос о ближайшем родственнике ископаемого Латимерии» . Журнал палеонтологии позвоночных . 25 (3): 481–491. doi : 10.1671/0272-4634(2005)025[0481:ANCASF]2.0.CO;2 . ISSN 0272-4634 . S2CID 86338307 .
- ^ Скшицкая, Роксана (3 июля 2014 г.). «Ревизия двух реликтовых актиноптеригий из средне- или верхнеюрской карабастауской свиты, хребет Каратау, Казахстан» . Алчеринга: Австралазийский журнал палеонтологии . 38 (3): 364–390. Бибкод : 2014Alch...38..364S . дои : 10.1080/03115518.2014.880267 . ISSN 0311-5518 . S2CID 129308632 .
- ^ Хилтон, Эрик Дж.; Гранде, Лэнс; Джин, Фан (январь 2021 г.). «Переописание † Yanosteus longidorsalis Jin et al. (Chondrostei, Acipenseriformes, †Peipiaosteidae) из раннего мела Китая» . Журнал палеонтологии . 95 (1): 170–183. Бибкод : 2021JPal...95..170H . дои : 10.1017/jpa.2020.80 . ISSN 0022-3360 . S2CID 225158727 .
- ^ Перейти обратно: а б Пойато-Ариса, Франсиско Хосе; Мартин-Абад, Хьюго (19 июля 2020 г.). «История двух линий: сравнительный анализ окаменелостей Amiiformes и Pycnodontiformes (Osteischtyes, Actinopterygii)» . Испанский журнал палеонтологии . 28 (1): 79. doi : 10.7203/sjp.28.1.17833 . HDL : 10486/710030 . ISSN 2255-0550 .
- ^ Дисри, Утумпорн; Наксри, Вилайлак; Джинтасакул, Пратуенг; Нода, Ёсиказу; Юкава, Хирокадзу; Хоссни, Тамара Эл; Кэвин, Лайонел (27 марта 2023 г.). «Новая рыба синамиин (Actinopterygii) из раннего мела Таиланда: значение для эволюционной истории линии амиидов» . Разнообразие . 15 (4): 491. дои : 10.3390/d15040491 . ISSN 1424-2818 .
- ^ Брито, Пауло М.; Альварадо-Ортега, Хесус; Менье, Франсуа Ж. (декабрь 2017 г.). «Самый ранний известный лепизостеоид расширяет ареал анатомически современных щук до поздней юры» . Научные отчеты . 7 (1): 17830. Бибкод : 2017NatSR...717830B . doi : 10.1038/s41598-017-17984-w . ISSN 2045-2322 . ПМК 5736718 . ПМИД 29259200 .
- ^ Арратия Г. Мезозойские галекостомы и ранняя радиация костистых рыб. В: Арратиа Дж., Тинтори А., редакторы. Мезозойские рыбы 3 – Систематика, палеосреда и биоразнообразие. Мюнхен: Верлаг д-р Фридрих Пфайль; 2004. с. 279–315.
- ^ Це, Цзе-Кей; Питтман, Майкл; Чанг, Ми-манн (26 марта 2015 г.). «Образец Paralycoptera Chang & Chou 1977 (Teleostei: Osteoglossoidei) из Гонконга (Китай) с потенциальным позднеюрским возрастом, который расширяет временной и географический ареал этого рода» . ПерДж . 3 : е865. дои : 10.7717/peerj.865 . ISSN 2167-8359 . ПМК 4380157 . ПМИД 25834774 .
- ^ Листон Дж., Ньюбри М., Чалландс Т. и Адамс К., 2013, «Рост, возраст и размер юрской пахикормиды Leedsichthyspromaticus (Osteichthyes: Actinopterygii) в: Arratia, G., Schultze, Х. и Уилсон, М. (ред.) Мезозойские рыбы 5 – Глобальное разнообразие и эволюция . Верлаг, д-р Фридрих Пфайль, Мюнхен, Германия, стр. 145–175.
- ^ Рис, Ян; Андервуд, Чарли Дж. (17 января 2008 г.). «Гибодонтные акулы английского бата и келловея (средней юры)» . Палеонтология . 51 (1): 117–147. Бибкод : 2008Palgy..51..117R . дои : 10.1111/j.1475-4983.2007.00737.x .
- ^ Корсо, Якопо Даль; Бернарди, Массимо; Сунь, Ядонг; Сун, Хайджун; Сейфулла, Лейла Дж.; Прето, Нерей; Джанолла, Пьеро; Раффелл, Аластер; Кустачер, Эвелин; Роги, Гвидо; Мерико, Агостино (сентябрь 2020 г.). «Вымирание и рассвет современного мира в Карнии (поздний триас)» . Достижения науки . 6 (38). Бибкод : 2020SciA....6...99D . дои : 10.1126/sciadv.aba0099 . ПМЦ 7494334 . ПМИД 32938682 .
- ^ Андервуд, Чарли Дж. (март 2006 г.). «Диверсификация Neoselachii (Chondrichthyes) в юрский и меловой период» . Палеобиология . 32 (2): 215–235. Бибкод : 2006Pbio...32..215U . дои : 10.1666/04069.1 . ISSN 0094-8373 . S2CID 86232401 .
- ^ Штумпф, Себастьян; Кривет, Юрген (01 декабря 2019 г.). «Новый комплекс плинсбахских пластиножаберных (Vertebrata, Chondrichthyes) из Европы и его вклад в понимание разнообразия и закономерностей распространения пластиножаберных поздней ранней юры» . ПалЗ . 93 (4): 637–658. Бибкод : 2019PalZ...93..637S . дои : 10.1007/s12542-019-00451-4 . ISSN 1867-6812 . S2CID 181782998 .
- ^ Андервуд, Чарли Дж.; Класон, Керин М. (июнь 2019 г.). «Позднеюрский скат Kimmerobatis etchesi gen. et sp. nov. и юрская радиация Batoidea » Труды Ассоциации геологов . 130 (3–4): 345–354. Бибкод : 2019ПрГА..130..345У дои : 10.1016/j.pgeola.2017.06.009 . S2CID 90691006 .
- ^ Кривет, Юрген; Клуг, Стефани (декабрь 2011 г.). «Новая юрская коровья акула (Chondrichthyes, Hexanchiformes) с комментариями по юрской гексанхиформной систематике» . Швейцарский журнал геонаук . 104 (С1): 107–114. дои : 10.1007/s00015-011-0075-z . ISSN 1661-8726 . S2CID 84405176 .
- ^ Срдич, Алекс; Даффин, Кристофер Дж.; Мартилл, Дэвид М. (август 2016 г.). «Первое появление оректолобиформной акулы Акаимиа в формации Оксфордской глины (юрский период, келловей) Англии» . Труды Ассоциации геологов . 127 (4): 506–513. Бибкод : 2016ПрГА..127..506С . дои : 10.1016/j.pgeola.2016.07.002 .
- ^ Слейтер, Тиффани С.; Эшбрук, Кейт; Кривет, Юрген (август 2020 г.). Кэвин, Лайонел (ред.). «Эволюционные взаимоотношения бычьих акул (Chondrichthyes, Heterodontiformes)» . Статьи по палеонтологии . 6 (3): 425–437. Бибкод : 2020PPal....6..425S . дои : 10.1002/spp2.1299 . hdl : 10468/10339 . ISSN 2056-2802 . S2CID 214133104 .
- ^ Джамбура, Патрик Л.; Киндлиманн, Рене; Лопес-Ромеро, Фавиэль; Маррама, Джузеппе; Пфафф, Катрин; Штумпф, Себастьян; Тюрчер, Юлия; Андервуд, Чарли Дж.; Уорд, Дэвид Дж.; Кривет, Юрген (декабрь 2019 г.). «Микрокомпьютерная томография показывает развитие уникальной структуры минерализации зубов у макрельевых акул (Chondrichthyes; Lamniformes) в глубоком прошлом» . Научные отчеты . 9 (1): 9652. Бибкод : 2019NatSR...9.9652J . дои : 10.1038/s41598-019-46081-3 . ISSN 2045-2322 . ПМК 6609643 . ПМИД 31273249 .
- ^ Лопес-Ромеро, Фавиэль А.; Штумпф, Себастьян; Пфафф, Катрин; Маррама, Джузеппе; Йохансон, Зерина; Кривет, Юрген (28 июля 2020 г.). «Эволюционные тенденции консервативной формы нейрокраниума у акул-ангелов (Squatiniformes, Elasmobranchii)» . Научные отчеты . 10 (1): 12582. Бибкод : 2020NatSR..1012582L . дои : 10.1038/s41598-020-69525-7 . ISSN 2045-2322 . ПМЦ 7387474 . ПМИД 32724124 .
- ^ Штумпф, Себастьян; Шеер, Удо; Кривет, Юрген (04 марта 2019 г.). «Новый род и вид вымерших наземных акул, †Diprosopovenator hilperti, gen. et sp. nov. (Carcharhiniformes, †Pseudoscyliorhinidae, fam. nov.), из верхнего мела Германии» . Журнал палеонтологии позвоночных . 39 (2): e1593185. Бибкод : 2019JVPal..39E3185S . дои : 10.1080/02724634.2019.1593185 . ISSN 0272-4634 . S2CID 155785248 .
- ^ Клюг, Стефани; Тюткен, Томас; Крылья, Оливер; Пфретчнер, Ганс-Ульрих; Мартин, Томас (сентябрь 2010 г.). «Собрание пресноводных акул позднеюрского периода (Chondrichthyes, Hybodontiformes) из южной части Джунгарского бассейна, Синьцзян, Северо-Западный Китай» . Палеобиоразнообразие и палеосреда . 90 (3): 241–257. Бибкод : 2010ПдПе...90..241К . дои : 10.1007/s12549-010-0032-2 . ISSN 1867-1594 . S2CID 129236098 .
- ^ Попов Евгений Владимирович; Дельсат, Доминик; Фельтен, Роланд (2 июля 2019 г.). «Новый род Callorhinchid (Holocephali, Chimaeroidei) из раннего байоса Оттанжа-Рюмеланжа, на границе Люксембурга и Франции» . Палеонтологические исследования . 23 (3): 220. дои : 10.2517/2018PR021 . ISSN 1342-8144 . S2CID 198423356 .
- ^ Даффин, Кристофер Дж.; Милан, Йеспер (14 ноября 2017 г.). «Новый голоцефал мириакантид из ранней юры Дании» . Бюллетень Геологического общества Дании . 65 : 161–170. дои : 10.37570/bgsd-2017-65-10 . ISSN 2245-7070 .
- ^ Лабандейра, Конрад К. (23 мая 2018 г.), «Ископаемая история разнообразия насекомых», Биоразнообразие насекомых , Чичестер, Великобритания: John Wiley & Sons, Ltd, стр. 723–788, doi : 10.1002/9781118945582.ch24 , ISBN 978-1-118-94558-2
- ^ Нел, Патрисия; Бертран, Сильвен; Нел, Андре (декабрь 2018 г.). «Диверсификация насекомых начиная с девона: новый подход, основанный на морфологическом несоответствии ротового аппарата» . Научные отчеты . 8 (1): 3516. Бибкод : 2018НатСР...8.3516Н . дои : 10.1038/s41598-018-21938-1 . ISSN 2045-2322 . ПМЦ 5824790 . ПМИД 29476087 .
- ^ Маккенна, Дуэйн Д.; Шин, Сынгван; Аренс, Дирк; Балке, Майкл; Беза-Беза, Кристиан; Кларк, Дэйв Дж.; Донат, Александр; Эскалона, Гермес Э.; Фридрих, Франк; Летч, Харальд; Лю, Шанлинь (03 декабря 2019 г.). «Эволюция и геномная основа разнообразия жуков» . Труды Национальной академии наук . 116 (49): 24729–24737. Бибкод : 2019PNAS..11624729M . дои : 10.1073/pnas.1909655116 . ISSN 0027-8424 . ПМК 6900523 . ПМИД 31740605 .
- ^ Шин, Сынгван; Кларк, Дэйв Дж; Леммон, Алан Р.; Мориарти Леммон, Эмили; Эйткен, Александр Л; Хаддад, Стефани; Фаррелл, Брайан Д; Марвальди, Адриана Э; Оберприелер, Рольф Г; Маккенна, Дуэйн Д. (01 апреля 2018 г.). «Филогеномные данные дают новое и надежное представление о филогении и эволюции долгоносиков» . Молекулярная биология и эволюция . 35 (4): 823–836. дои : 10.1093/molbev/msx324 . hdl : 11336/57287 . ISSN 0737-4038 . ПМИД 29294021 . S2CID 4366092 .
- ^ Сюй, Чуньпэн; Фанг, Янан; Фанг, Ян; Ван, Хэ; Чжоу, Цянь; Цзян, Сюэин; Чжан, Хайчунь (14 февраля 2024 г.). «Раннеюрские прямокрылые насекомые из южной части Джунгарского бассейна, северо-запад Китая, с обсуждением изменений биоразнообразия прямокрылых на границе триаса и юры» . Геологическое общество, Лондон, специальные публикации . 538 (1): 147–154. дои : 10.1144/SP538-2021-184 . ISSN 0305-8719 . Проверено 22 июня 2024 г. - из публикаций геологического общества коллекции Лайелла.
- ^ ван Элдейк, Тимо Дж.Б.; Вапплер, Торстен; Стротер, Пол К.; ван дер Вейст, Кэролин М.Х.; Раджаи, Хосейн; Вишер, Хенк; ван де Шутбрюгге, Бас (январь 2018 г.). «Триасово-юрское окно в эволюцию чешуекрылых» . Достижения науки . 4 (1): e1701568. Стартовый код : 2018SciA....4.1568V . дои : 10.1126/sciadv.1701568 . ISSN 2375-2548 . ПМК 5770165 . ПМИД 29349295 .
- ^ Кохли, Манприт Каур; Уэр, Джессика Л.; Бехли, Гюнтер (2016). «Как датировать стрекозу: окаменелости для стрекоз» . Электронная палеонтология . 19 (1). дои : 10.26879/576 .
- ^ Хуан, Ди Ин; Энгель, Майкл С.; Цай, Чэньян; Нел, Андре (май 2013 г.). «Мезозойские гигантские блохи из северо-восточного Китая (Siphonaptera): таксономия и значение для палеоразнообразия» . Китайский научный бюллетень . 58 (14): 1682–1690. Бибкод : 2013ЧСБу..58.1682Х . дои : 10.1007/s11434-013-5769-3 . hdl : 1808/14426 . ISSN 1001-6538 . S2CID 53578959 .
- ^ Гао, Тайпин; Ши, Чунгкун; Расницын Александр П.; Сюй, Син; Ван, Шуо; Рен, Донг (июль 2013 г.). «Новые переходные блохи из Китая, подчеркивающие разнообразие эктопаразитических насекомых раннего мела» . Современная биология . 23 (13): 1261–1266. Бибкод : 2013CBio...23.1261G . дои : 10.1016/j.cub.2013.05.040 . ПМИД 23810530 . S2CID 9646168 .
- ^ Лабандейра, Конрад К.; Ли, Лунфэн (2021), Де Баетс, Кеннет; Хантли, Джон Уоррен (ред.), «История паразитизма насекомых и среднемезозойская паразитоидная революция» , «Эволюция и ископаемые летописи паразитизма» , «Темы геобиологии», том. 49, Чам: Springer International Publishing, стр. 377–533, номер документа : 10.1007/978-3-030-42484-8_11 , ISBN. 978-3-030-42483-1 , S2CID 236738176 , получено 2 декабря 2021 г.
- ^ Ян, Хунру; Ши, Чаофан; Энгель, Майкл С; Чжао, Чжипенг; Рен, Донг; Гао, Тайпин (02 апреля 2020 г.). «Ранние специализации юрских палочников по мимикрии и защите» . Национальный научный обзор . 8 (1): nwaa056. дои : 10.1093/nsr/nwaa056 . ISSN 2095-5138 . ПМЦ 8288419 . ПМИД 34691548 .
- ^ Хуан, Ди-ин; Нел, Андре; Зомпро, Оливер; Уоллер, Ален (11 июня 2008 г.). «Мантофазматодеи сейчас в юрском периоде» . естественные науки . 95 (10): 947–952. Бибкод : 2008NW.....95..947H . дои : 10.1007/s00114-008-0412-x . ISSN 0028-1042 . ПМИД 18545982 . S2CID 35408984 .
- ^ Хуан, Ди-Ин; Нел, Андре (август 2009 г.). «Старейшие паутины из средней юры Внутренней Монголии, Китай (Insecta: Embiodea)» . Зоологический журнал Линнеевского общества . 156 (4): 889–895. дои : 10.1111/j.1096-3642.2008.00499.x .
- ^ Энгель, Майкл С.; Винтертон, Шон Л.; Брейткройц, Лаура CV (07.01.2018). «Филогения и эволюция Neuropterida: куда нас занесли кружевные крылья?» . Ежегодный обзор энтомологии . 63 (1): 531–551. doi : 10.1146/annurev-ento-020117-043127 . ISSN 0066-4170 . ПМИД 29324039 .
- ^ Селден, Пол А.; Бейкер, Энн С.; Фиппс, Кеннет Дж. (2008). «Пурьбатидный клещ (паукообразный: Acari) из оксфордской глины (юрский период: верхний келловей) карьера South Cave Station, Йоркшир, Великобритания» . Палеонтология . 51 (3): 623–633. Бибкод : 2008Palgy..51..623S . дои : 10.1111/j.1475-4983.2008.00769.x . hdl : 1808/8353 . ISSN 1475-4983 . S2CID 54046836 .
- ^ Сивхед, Ульф; Уоллворк, Джон А. (март 1978 г.). «Раннеюрский панцирный клещ из южной Швеции» . Переговоры Геологического общества Стокгольма . 100 (1): 65–70. дои : 10.1080/11035897809448562 . ISSN 0016-786X .
- ^ Перейти обратно: а б Магальяс, Иван Л.Ф.; Азеведо, Гильерме ХФ; Михалик, Питер; Рамирес, Мартин Х. (февраль 2020 г.). «Пересмотр окаменелостей пауков: значение для калибровки деревьев и свидетельства крупного круговорота фауны со времен мезозоя» . Биологические обзоры . 95 (1): 184–217. дои : 10.1111/brv.12559 . ISSN 1464-7931 . ПМИД 31713947 . S2CID 207937170 .
- ^ Селден, Пол А.; Данлоп, Джейсон А. (2014). «Первый ископаемый паук (Araneae: Palpimanoidea) из нижней юры (Гриммен, Германия)». Зоотакса . 3894 (1): 161–168. дои : 10.11646/zootaxa.3894.1.13 . ПМИД 25544628 .
- ^ Селден, Пенсильвания; Ши, СК; Рен, Д. (2013). «Гигантский паук из юрского периода Китая демонстрирует большее разнообразие круговой стеблевой группы» . Naturwissenschaften . 100 (12): 1171–1181. Бибкод : 2013NW....100.1171S . дои : 10.1007/s00114-013-1121-7 . ПМЦ 3889289 . ПМИД 24317464 .
- ^ Данлоп, Джейсон А.; Каменц, Карстен; Шольц, Герхард (июнь 2007 г.). «Переосмысление морфологии юрского скорпиона Liassoscorpionides» . Строение и развитие членистоногих . 36 (2): 245–252. Бибкод : 2007ArtSD..36..245D . дои : 10.1016/j.asd.2006.09.003 . ПМИД 18089103 .
- ^ Хуан, Диин; Селден, Пол А.; Данлоп, Джейсон А. (август 2009 г.). «Жнецы (Arachnida: Opiliones) из средней юры Китая» . Naturwissenschaften . 96 (8): 955–962. Бибкод : 2009NW.....96..955H . дои : 10.1007/s00114-009-0556-3 . ISSN 0028-1042 . ПМИД 19495718 . S2CID 9570512 .
- ^ Гирибет, Гонсало; Туриньо, Ана Люсия; Ши, Чункун; Рен, Донг (март 2012 г.). «Прекрасно сохранившийся сенокосец (Arthropoda, Arachnida, Opiliones) из средней юры Китая» . Разнообразие и эволюция организмов . 12 (1): 51–56. дои : 10.1007/s13127-011-0067-x . ISSN 1439-6092 . S2CID 15658216 .
- ^ Данхилл, Александр М.; Фостер, Уильям Дж.; Скиберрас, Джеймс; Твитчетт, Ричард Дж. (январь 2018 г.). Хаутманн, Майкл (ред.). «Влияние массового вымирания позднего триаса на функциональное разнообразие и состав морских экосистем» . Палеонтология . 61 (1): 133–148. Бибкод : 2018Palgy..61..133D . дои : 10.1111/пала.12332 .
- ^ Кисслинг, Вольфганг (декабрь 2009 г.). «Геологический и биологический контроль эволюции рифов» . Ежегодный обзор экологии, эволюции и систематики . 40 (1): 173–192. doi : 10.1146/annurev.ecolsys.110308.120251 . ISSN 1543-592X .
- ^ Перейти обратно: а б с Кломпмейкер, А.А.; Швейцер, CE; Фельдманн, Р.М.; Ковалевски, М. (01 ноября 2013 г.). «Влияние рифов на возникновение мезозойских морских ракообразных» . Геология . 41 (11): 1179–1182. Бибкод : 2013Geo....41.1179K . дои : 10.1130/G34768.1 . ISSN 0091-7613 .
- ^ Хадсон, Венди; Харт, Малкольм Б.; Смарт, Кристофер В. (1 января 2009 г.). «Палеобиогеография ранних планктонных фораминифер» . Бюллетень геологического общества Франции . 180 (1): 27–38. дои : 10.2113/gssgfbull.180.1.27 . ISSN 1777-5817 .
- ^ Вигган, Николас Дж.; Райдинг, Джеймс Б.; Фенсом, Роберт А.; Маттиоли, Эмануэла (01 мая 2018 г.). «Байос (средняя юра): ключевой интервал ранней мезозойской радиации фитопланктона» . Обзоры наук о Земле . 180 : 126–146. Бибкод : 2018ESRv..180..126W . doi : 10.1016/j.earscirev.2018.03.009 . ISSN 0012-8252 .
- ^ Затонь, М.; Тейлор, PD (31 декабря 2009 г.). «Микроконхиды (Tentaculita) из средней юры Польши» . Бюллетень геонаук : 653–660. дои : 10.3140/bull.geosci.1167 . ISSN 1802-8225 .
- ^ Жирар, Винсент; Сен-Мартен, Симона; Баффето, Эрик; Сен-Мартен, Жан-Поль; Неродо, Дидье; Пейро, Даниэль; Роги, Гвидо; Мальчики, Юджин; Сутитхорн, Варавуд (2020). Сен-Мартен, Ж.-П.; Сен-Мартен, С. (ред.). «Тайский янтарь: понимание ранней истории диатомовых водорослей?» . BSGF – Бюллетень наук о Земле . 191 : 23. дои : 10.1051/bsgf/2020028 . ISSN 1777-5817 .
- ^ Горжелак, Пшемыслав; Саламон, Мариуш А.; Тшесиок, Давид; Лах, Рафал; Баумиллер, Томаш К. (апрель 2016 г.). «Динамика разнообразия постпалеозойских криноидей – в поисках факторов, влияющих на макроэволюцию криноидей» . Летайя . 49 (2): 231–244. Бибкод : 2016Лета..49..231Г . дои : 10.1111/let.12141 .
- ^ Хопкинс, Мелани Дж.; Смит, Эндрю Б. (24 марта 2015 г.). «Динамические эволюционные изменения постпалеозойских ежей и важность масштаба при интерпретации изменений в темпах эволюции» . Труды Национальной академии наук . 112 (12): 3758–3763. Бибкод : 2015PNAS..112.3758H . дои : 10.1073/pnas.1418153112 . ISSN 0027-8424 . ПМЦ 4378421 . ПМИД 25713369 .
- ^ Шольц, Герхард (ноябрь 2020 г.). «Предшественник Eocarcinus Withers, 1932 (Malacostraca, Decapoda, Meiura) представляет собой стволовую группу брахиуранов » Строение и развитие членистоногих . 59 : Бибкод : 2020ArtSD..5900991S 100991. дои : 10.1016/j.asd.2020.100991 . ПМИД 32891896 .
- ^ Швейцер, Кэрри Э.; Фельдманн, Родни М. (1 мая 2010 г.). «Самая древняя брахюра (Decapoda: Homolodromioidea: Glaessneropsoidea), известная на сегодняшний день (юрский период)» . Журнал биологии ракообразных . 30 (2): 251–256. дои : 10.1651/09-3231.1 . ISSN 0278-0372 . S2CID 84707572 .
- ^ Перейти обратно: а б Гино, Даниэль (14 ноября 2019 г.). «Новые гипотезы о древнейших брахюрах (Crustacea, Decapoda, Brachyura)» . Геодиверситас . 41 (1): 747. doi : 10.5252/geodiversitas2019v41a22 . ISSN 1280-9659 . S2CID 214220075 .
- ^ Фраайе, Рене; Швайгерт, Гюнтер; Нютцель, Александр; Гавлик, Филипп (1 января 2013 г.). «Новые раннеюрские раки-отшельники из Германии и Франции» . Журнал биологии ракообразных . 33 (6): 802–817. дои : 10.1163/1937240X-00002191 . ISSN 0278-0372 .
- ^ Мироненко, Александр (январь 2020 г.). «Рак-отшельник, сохранившийся внутри раковины аммонита из верхней юры центральной России: значение для палеоэкологии аммоноидей» . Палеогеография, Палеоклиматология, Палеоэкология . 537 : 109397. Бибкод : 2020PPP...53709397M . дои : 10.1016/j.palaeo.2019.109397 .
- ^ Бракен-Гриссом, Хизер Д.; Ахён, Шейн Т.; Уилкинсон, Ричард Д.; Фельдманн, Родни М.; Швейцер, Кэрри Э.; Брейнхольт, Джесси В.; Бендалл, Мэтью; Палеро, Ферран; Чан, Тин-Ям; Фелдер, Дэррил Л.; Роблес, Рафаэль (01 июля 2014 г.). «Появление омаров: филогенетические отношения, морфологическая эволюция и сравнение времени дивергенции древней группы (Decapoda: Achelata, Astacidea, Glypheidea, Polychelida)» . Систематическая биология . 63 (4): 457–479. дои : 10.1093/sysbio/syu008 . ISSN 1063-5157 . ПМИД 24562813 .
- ^ Чан, Бенни К.К.; Дрейер, Никлас; Гейл, Энди С; Гленнер, Хенрик; Эверс-Сауседо, Кристина; Перес-Лосада, Маркос; Колбасов Григорий А; Крэндалл, Кейт А; Хёг, Йенс Т (25 февраля 2021 г.). «Эволюционное разнообразие ракушек с обновленной классификацией ископаемых и живых форм» . Зоологический журнал Линнеевского общества . 193 (3): 789–846. doi : 10.1093/zoolinnean/zlaa160 . HDL : 11250/2990967 . ISSN 0024-4082 .
- ^ Гейл, Энди; Швайгерт, Гюнтер (январь 2016 г.). Хаутманн, Майкл (ред.). «Новый усоногий с фосфатным панцирем (Crustacea, Thoracica) из нижней юры (тоарка) Германии – древнейшая эпипланктонная ракушка» . Палеонтология . 59 (1): 59–70. Бибкод : 2016Palgy..59...59G . дои : 10.1111/пала.12207 . S2CID 128383968 .
- ^ Вёрёш, Аттила; Кочиш, Адам Т.; Палфи, Йожеф (сентябрь 2016 г.). «Гибель последних двух отрядов брахиопод со шпилями (Spiriferinida и Athyridida) во время тоарского (раннеюрского) вымирания» . Палеогеография, Палеоклиматология, Палеоэкология . 457 : 233–241. Бибкод : 2016PPP...457..233В . дои : 10.1016/j.palaeo.2016.06.022 .
- ^ Вёрёш, Аттила; Кочиш, Адам Т.; Палфи, Йожеф (2019). «Массовые вымирания и вымирания клад в истории брахиопод: краткий обзор и постпалеозойское тематическое исследование» . Rivista Italiana di Paleontologia e Stratigrafia . 125 (3). дои : 10.13130/2039-4942/12184 . ISSN 2039-4942 . Архивировано из оригинала 01 сентября 2020 г. Проверено 25 декабря 2020 г.
- ^ Манойлович, Марко; Клэпхэм, Мэтью Э. (23 ноября 2020 г.). «Роль нарушения субстрата, вызванного биотурбацией, в упадке мезозойских брахиопод» . Палеобиология . 47 : 86–100. дои : 10.1017/pab.2020.50 . ISSN 0094-8373 .
- ^ Тейлор, Пол Д.; Эрнст, Андрей (июнь 2008 г.). «Мшанки в переходном периоде: обедневшая и неоднородная юрская биота» . Палеогеография, Палеоклиматология, Палеоэкология . 263 (1–2): 9–23. Бибкод : 2008PPP...263....9T . дои : 10.1016/j.palaeo.2008.01.028 .
- ^ Роуз, Соня; Де Ренци, Майкл; Дамбореня, Сусана Э.; Маркес-Алиага, Ана (ноябрь 2011 г.). «Преодоление кризисов: динамика разнообразия двустворчатых моллюсков в раннем триасе и ранней юре» . Палеогеография, Палеоклиматология, Палеоэкология . 311 (3–4): 184–199. Бибкод : 2011PPP...311..184R . дои : 10.1016/j.palaeo.2011.08.020 . hdl : 11336/81358 .
- ^ Мондал, Субхронил; Харрис, Питер Дж. (февраль 2016 г.). «Влияние таксономических поправок на тенденции фанерозойского родового богатства морских двустворчатых моллюсков с обсуждением общей истории клады» . Палеобиология . 42 (1): 157–171. Бибкод : 2016Pbio...42..157M . дои : 10.1017/pab.2015.35 . ISSN 0094-8373 . S2CID 87260961 .
- ^ Ша, Дж.; Честари, Р.; Фабби, С. (апрель 2020 г.). «Палеобиогеографическое распространение рудистских двустворчатых моллюсков (Hippuritida) в оксфорде – раннем апте (поздней юре – раннем меле)» . Меловые исследования . 108 : 104289. Бибкод : 2020CrRes.10804289S . дои : 10.1016/j.cretres.2019.104289 . S2CID 210248232 .
- ^ Пейдж, Кевин Н. (январь 2008 г.). «Эволюция и география юрских аммоноидей» . Труды Ассоциации геологов . 119 (1): 35–57. Бибкод : 2008ПрГА..119...35П . дои : 10.1016/S0016-7878(08)80257-X .
- ^ Сандовал, Джозеф; О'Догерти, Джин; Гекс, Жан (1 августа 2001 г.). «Скорость эволюции юрских аммонитов в зависимости от колебаний уровня моря» . ПАЛЕОС 16 (4): 311–335. Бибкод : 2001Представитель..16..311S . doi : 10.1669/0883-1351(2001)016<0311:EROJAI>2.0.CO;2 . S2CID 129982065 . Получено 26 августа.
- ^ Иба, Ясухиро; Сано, Синъити; Муттерлозе, Йорг (2 мая 2014 г.). Самондс, Карен Э. (ред.). «Ранняя эволюционная история белемнитов: новые данные из Японии» . ПЛОС ОДИН . 9 (5): е95632. Бибкод : 2014PLoSO...995632I . дои : 10.1371/journal.pone.0095632 . ISSN 1932-6203 . ПМЦ 4008418 . ПМИД 24788872 .
- ^ Хоффманн, Рене; Стивенс, Кевин (февраль 2020 г.). «Палеобиология белемнитов – основа интерпретации геохимии трибун» . Биологические обзоры . 95 (1): 94–123. дои : 10.1111/brv.12557 . ISSN 1464-7931 . ПМИД 31729839 . S2CID 208036104 .
- ^ Фокс, Дирк; Вайс, Роберт (11 июля 2008 г.). «Таксономия, морфология и филогения нижнеюрских лолигосепиидных колеоидов (Cephalopoda)» . Новогодний ежегодник по геологии и палеонтологии-трактаты . 249 (1): 93–112. дои : 10.1127/0077-7749/2008/0249-0093 . ISSN 0077-7749 .
- ^ Перейти обратно: а б с Фукс, Дирк; Иба, Ясухиро; Хейнг, Александр; Иидзима, Масая; Клюг, Кристиан; Ларсон, Нил Л.; Швайгерт, Гюнтер (февраль 2020 г.). Брайард, Арно (ред.). «Muensterelloidea: филогения и эволюция характера мезозойских стволовых осьминогов» . Статьи по палеонтологии . 6 (1): 31–92. Бибкод : 2020PPal....6...31F . дои : 10.1002/spp2.1254 . ISSN 2056-2802 . S2CID 198256507 .
- ^ Фукс, Дирк; Швайгерт, Гюнтер (июнь 2018 г.). «Первые остатки гладиусов средней и поздней юры предоставляют новые свидетельства подробного происхождения перисто- и перисторассеченных осьминогов (Coleoidea)» . ПалЗ . 92 (2): 203–217. Бибкод : 2018PalZ...92..203F . дои : 10.1007/s12542-017-0399-8 . ISSN 0031-0220 . S2CID 135245479 .
- ^ Крута, Изабель; Руже, Изабель; Шарбонье, Сильвен; Барден, Жереми; Фернандес, Винсент; Жермен, Дэмиен; Брайард, Арно; Ландман, Нил (2016). «Proteroctopus Ribeti в эволюции колеоидов» . Палеонтология . 59 (6): 767–773. Бибкод : 2016Palgy..59..767K . дои : 10.1111/пала.12265 . ISSN 1475-4983 . S2CID 132420410 .
Внешние ссылки
[ редактировать ]- Примеры юрских окаменелостей
- Юрский период (шкала хроностратиграфии)
- Юрские окаменелости в Харбери, Уорикшир
- Юрские микроископаемые: более 65 изображений фораминифер
- . Британская энциклопедия . Том. 15 (11-е изд.). 1911 г. С картой и таблицей.