Экологическая преемственность

Экологическая сукцессия — это процесс изменения видов , составляющих экологическое сообщество, с течением времени.
Процесс сукцессии происходит либо после первоначальной колонизации вновь созданной среды обитания, либо после того, как нарушение существенно изменяет ранее существовавшую среду обитания. [1] Сукцессия, которая начинается в новых средах обитания, не подверженная влиянию ранее существовавших сообществ, называется первичной сукцессией , тогда как сукцессия, которая следует за разрушением ранее существовавшего сообщества, называется вторичной сукцессией . [1] Первичная сукцессия может произойти после потока лавы или появления нового острова из океана. Суртсей , вулканический остров у южного побережья Исландии , является важным примером места, где наблюдалась первичная сукцессия. [2] [3] С другой стороны, вторичная сукцессия происходит после нарушения сообщества, например, в результате пожара , сильного ветра или вырубки леса .
Преемственность была одной из первых теорий, выдвинутых в экологии . Экологическая преемственность была впервые задокументирована в дюнах Индианы на северо-западе Индианы и остается важной экологической темой исследований. [4] Со временем понимание преемственности изменилось с линейного прогресса к стабильному кульминационному состоянию на более сложную циклическую модель, которая не придает значения идее о том, что организмы имеют фиксированные роли или отношения. [5]
История
[ редактировать ]Предшественники идеи экологической преемственности восходят к началу XIX века. Еще в 1742 году французский натуралист Бюффон заметил, что в естественной эволюции леса тополя предшествуют дубам и букам. заставил Бюффона Позже богословский комитет Парижского университета отказаться от многих своих идей, поскольку они противоречили библейскому повествованию о сотворении мира. [6]
Швейцарский геолог Жан-Андре Делюк и позднее французский натуралист Адольф Дюро де ла Малль были первыми, кто использовал слово «преемственность» в отношении развития растительности после вырубки леса. [7] [8] В 1859 году Генри Дэвид Торо написал обращение под названием «Последовательность лесных деревьев». [9] в котором он описал сукцессию в дубово-сосновом лесу. «Наблюдателям давно известно, что белки зарывают орехи в землю, но я не знаю, чтобы кто-нибудь таким образом объяснил правильную смену лесов». [10] Австрийский ботаник Антон Кернер опубликовал исследование о сукцессии растений в бассейне реки Дунай в 1863 году. [11]
В исследовании Рагнара Хульта 1885 года об этапах развития лесов в Блекинге отмечалось, что луга становятся пустоши до того, как пустошь превращается в лес. береза На ранних стадиях развития леса доминировала , затем сосна (на сухой почве) и ель (на влажной почве). Если березу заменять дубом, она со временем превращается в бук . Болота переходят от мха к осоке, затем к болотной растительности, за которой следуют береза и, наконец, ель. [6]
ХК Коулз
[ редактировать ]
Между 1899 и 1910 годами Генри Чандлер Коулз из Чикагского университета разработал более формальную концепцию преемственности. Вдохновленный исследованиями датских дюн, проведенными Юджином Уормингом , Коулз изучал развитие растительности на песчаных дюнах на берегу озера Мичиган ( Дюны Индианы ). Он признал, что растительность на дюнах разного возраста можно интерпретировать как разные стадии общей тенденции развития растительности на дюнах (подход к изучению изменений растительности, позже названный заменой пространства-времени или исследованиями хронопоследовательности ). Впервые он опубликовал эту работу в виде статьи в «Ботанической газете» в 1899 году («Экологические связи растительности песчаных дюн озера Мичиган»). [12] В этой классической публикации и последующих статьях он сформулировал идею первичной сукцессии и понятие серии — повторяющейся последовательности изменений сообщества, специфичной для конкретных условий окружающей среды. [4] [13]
Глисон и Клементс
[ редактировать ]Однако примерно с 1900 по 1960 год в понимании сукцессии доминировали теории Фредерика Клементса , современника Коулза, который считал, что серии были в высшей степени предсказуемыми и детерминированными и сходились в климатически детерминированном стабильном климаксном сообществе независимо от стартовых условий. Клементс явно провел аналогию сукцессионного развития экологических сообществ с онтогенетическим развитием отдельных организмов, и его модель часто называют псевдоорганизмической теорией экологии сообществ. Клементс и его последователи разработали сложную таксономию сообществ и путей сукцессии.
Генри Глисон предложил противоположную концепцию еще в 1920-х годах. Модель Глисона была более сложной и гораздо менее детерминированной, чем модель Клементса. Наиболее фундаментально она отличается от точки зрения Клементса, предполагая гораздо большую роль случайных факторов и отрицая существование последовательных, резко ограниченных типов сообществ. Глисон утверждал, что распространение видов индивидуалистически реагировало на факторы окружающей среды, а сообщества лучше всего рассматривать как артефакты сопоставления распределения видов. Идеи Глисона, впервые опубликованные в 1926 году, по большей части игнорировались до конца 1950-х годов.
Две цитаты иллюстрируют противоположные взгляды Клементса и Глисона. Клементс писал в 1916 году:
Изучение развития растительности обязательно основывается на предположении, что единица или кульминационная формация представляет собой органическое образование. Как организм образование возникает, растет, созревает и умирает. При этом каждое климаксное образование способно воспроизводить себя, с необходимой точностью повторяя этапы своего развития.
— Фредерик Клементс [14]
в то время как Глисон в своей статье 1926 года сказал:
Ассоциация — это не организм и даже не растительная единица, а просто совпадение.
— Генри Глисон [15]
Идеи Глисона на самом деле больше соответствовали первоначальным представлениям Коулза о преемственности. О различии Клементса между первичной и вторичной преемственностью Коулз писал (1911):
Эта классификация, по-видимому, не имеет фундаментальной ценности, поскольку она разделяет такие тесно связанные явления, как эрозия и отложения, и объединяет такие непохожие вещи, как деятельность человека и оседание земли.
— Генри Коулз [16]
Юджин Одум
[ редактировать ]В 1969 году Юджин Одум опубликовал «Стратегию развития экосистемы» , документ, оказавший большое влияние на сохранение и восстановление окружающей среды. Одум утверждал, что экологическая преемственность представляет собой упорядоченное движение к кульминационному состоянию, когда «на единицу потока энергии сохраняется максимальная биомасса и симбиотическая функция между организмами». [17] Одум подчеркнул, что сукцессия — это не просто изменение видового состава экосистемы, но также приводит к изменению более сложных атрибутов экосистемы, таких как структура и круговорот питательных веществ . [18]
современная эпоха
[ редактировать ]Более тщательное тестирование сукцессионных моделей и теории сообществ на основе данных обычно началось с работ Роберта Уиттакера и Джона Кертиса в 1950-х и 1960-х годах. Теория преемственности с тех пор стала менее монолитной и более сложной. Дж. Коннелл и Р. Слейер предприняли попытку систематизировать сукцессионные процессы по механизму. Среди британских и североамериканских экологов идея стабильной кульминационной растительности в значительной степени была отвергнута, а сукцессионные процессы стали рассматриваться как гораздо менее детерминированные, с важной ролью исторических случайностей и альтернативных путей в фактическом развитии сообществ. Продолжаются дебаты относительно общей предсказуемости сукцессионной динамики и относительной важности равновесных и неравновесных процессов. Бывший профессор Гарварда Фахри А. Баззаз ввел в дискуссию понятие масштаба , поскольку он считал, что на местном уровне или в масштабе небольшой территории процессы являются случайными и неоднородными, но, принимая во внимание более крупные региональные территории, нельзя отрицать определенные тенденции. [19]
Более поздние определения преемственности подчеркивают изменение как центральную характеристику. [17] Новые методы исследования значительно расширяют возможности современных учёных изучать последовательность, которая сейчас не считается ни полностью случайной, ни полностью предсказуемой. [18]
Факторы
[ редактировать ]Раньше экологическая последовательность рассматривалась как имеющая стабильную конечную стадию, называемую кульминацией , иногда называемую «потенциальной растительностью» участка и определяемую в первую очередь местным климатом. От этой идеи современные экологи в значительной степени отказались в пользу неравновесных представлений о динамике экосистем. Большинство природных экосистем подвергаются нарушениям со скоростью, которая делает «кульминацию» сообщества недостижимой. Изменение климата часто происходит со скоростью и частотой, достаточной для предотвращения достижения кульминационного состояния. Дополнения к доступным пулам видов посредством расширения ареала и интродукции также могут постоянно изменять сообщества. [20]
Развитие некоторых атрибутов экосистемы, таких как свойства почвы и циклы питательных веществ , находятся под влиянием свойств сообщества и, в свою очередь, влияют на дальнейшее сукцессионное развитие. Этот процесс обратной связи может происходить только в течение столетий или тысячелетий. В сочетании со стохастической природой возмущений и других долгосрочных (например, климатических) изменений такая динамика заставляет сомневаться в том, что концепция «кульминации» когда-либо применима или особенно полезна при рассмотрении фактической растительности. [21]
На траекторию сукцессионных изменений могут влиять начальные условия участка, тип нарушения, вызывающего сукцессию, взаимодействие присутствующих видов, а также более случайные факторы, такие как наличие колонистов или семян или погодные условия во время нарушения. . Некоторые аспекты преемственности в целом предсказуемы; другие могут протекать более непредсказуемо, чем при классической точке зрения на экологическую последовательность. Двумя важными факторами возмущения сегодня являются действия человека и изменение климата . [22]
Хотя идея фиксированного, предсказуемого процесса преемственности с одной четко определенной кульминацией является чрезмерно упрощенной моделью, некоторые предсказания, сделанные классической моделью, точны. Видовое разнообразие, общая биомасса растений, продолжительность жизни растений, важность организмов- разрушителей и общая стабильность увеличиваются по мере того, как сообщество приближается к кульминационному состоянию, в то время как скорость потребления почвенных питательных веществ, скорость биогеохимического круговорота и скорость чистой первичной продуктивности. все они уменьшаются по мере того, как сообщество приближается к кульминационному состоянию. [23]
В сообществах ранней сукцессии будут доминировать быстрорастущие, хорошо рассредоточенные виды ( оппортунистические , беглые или r-отобранные жизненные истории). Их еще называют видами-пионерами . По мере продолжения сукцессии эти виды будут иметь тенденцию заменяться более конкурентоспособными ( k-отобранными ) видами.
Некоторые из этих тенденций применимы не во всех случаях. Например, видовое разнообразие почти обязательно увеличивается во время ранней сукцессии по мере появления новых видов, но может снижаться в более поздней сукцессии, поскольку конкуренция устраняет оппортунистические виды и приводит к доминированию локально превосходящих конкурентов . Чистая первичная продуктивность , биомасса и трофические свойства демонстрируют переменные закономерности в зависимости от конкретной системы и участка.
Типы
[ редактировать ]Первичная преемственность
[ редактировать ]Сукцессионная динамика, начинающаяся с колонизации территории, которая ранее не была занята экологическим сообществом, называется первичной сукцессией. [1] Сюда входят недавно обнаженные поверхности камней или песка, потоки лавы и недавно обнаженные ледниковые отложения. [1] К стадиям первичной сукцессии относятся микроорганизмы-пионеры, [24] растения (лишайники и мхи), травянистый ярус, более мелкие кустарники и деревья. Животные начинают возвращаться, когда есть еда, которую они могут съесть. Когда это полностью функционирующая экосистема, она достигает кульминационной стадии сообщества. [25]
Вторичная преемственность
[ редактировать ]
- Стабильное сообщество лиственных лесов
- Возмущение, такое как лесной пожар, уничтожает лес.
- Огонь сжигает лес дотла
- Огонь оставляет после себя пустую, но не уничтоженную почву.
- Травы и другие травянистые растения отрастают первыми.
- Небольшие кусты и деревья начинают заселять территорию.
- Быстрорастущие вечнозеленые деревья развиваются в полной мере, тогда как теневыносливые деревья развиваются в подлеске.
- Недолговечные и теневыносливые вечнозеленые деревья умирают, когда над ними возвышаются более крупные лиственные деревья. Экосистема сейчас вернулась в то же состояние, в котором она началась.
Вторичная сукцессия следует за серьезным нарушением или удалением ранее существовавшего сообщества, в котором есть остатки предыдущей экосистемы. [1] На вторичную сукцессию сильно влияют условия, предшествовавшие нарушению, такие как развитие почвы, семенные банки , оставшееся органическое вещество и остаточные живые организмы. [1] Из-за остаточной плодовитости и ранее существовавших организмов изменения сообщества на ранних стадиях вторичной сукцессии могут быть относительно быстрыми. [1]
Вторичная сукцессия наблюдается и изучается гораздо чаще, чем первичная сукцессия. Особенно распространенные типы вторичной сукцессии включают реакцию на природные нарушения, такие как пожар, наводнение и сильные ветры, а также на антропогенные нарушения, такие как лесозаготовки и сельское хозяйство. При вторичной сукцессии почву и организмы необходимо оставить нетронутыми, чтобы у нового материала была возможность восстановиться. [9]
Например, во фрагментированной старой полевой среде обитания, созданной в восточном Канзасе, древесные растения «более быстро (на единицу площади) колонизировались на больших и близлежащих участках ». [26]

Вторичная сукцессия может быстро изменить ландшафт. В 1900-х годах в национальном парке Акадия случился лесной пожар, уничтоживший большую часть ландшафта. Первоначально на территории росли вечнозеленые деревья. После пожара на этой территории понадобился как минимум год, чтобы вырастить кустарник. Со временем вместо вечнозеленых растений начали расти лиственные деревья. [25]
Вторичная сукцессия произошла в национальном парке Шенандоа после наводнения рек Мурман и Рапидан в 1995 году, которое уничтожило растительный и животный мир. [27]
Сезонная и циклическая динамика
[ редактировать ]В отличие от вторичной сукцессии, эти типы изменений растительности не зависят от нарушений , а представляют собой периодические изменения, возникающие в результате нестабильного взаимодействия видов или повторяющихся событий. Эти модели модифицируют концепцию кульминации в сторону одного из динамических состояний.
Причины сукцессии растений
[ редактировать ]Аутогенная сукцессия может быть вызвана изменениями в почве, вызванными находящимися там организмами. Эти изменения включают накопление органических веществ в подстилке или гуминовом слое, изменение питательных веществ в почве или изменение pH почвы из-за произрастающих там растений. Структура самих растений также может изменить сообщество. [28] Например, когда более крупные виды, такие как деревья, созревают, они создают тень на развивающейся лесной подстилке , которая, как правило, исключает виды, требующие света. Теневыносливые виды проникнут на территорию.
Аллогенная сукцессия обусловлена внешними воздействиями окружающей среды, а не растительностью. Например, изменения почвы из-за эрозии, выщелачивания или отложения ила и глины могут изменить содержание питательных веществ и водные отношения в экосистемах. Животные также играют важную роль в аллогенных изменениях, поскольку они являются опылителями, распространителями семян и травоядными животными. Они также могут увеличивать содержание питательных веществ в почве в определенных областях или перемещать почву (как это делают термиты, муравьи и кроты), создавая участки в среде обитания. Это может создать места регенерации, благоприятствующие определенным видам.
Климатические факторы могут быть очень важными, но в гораздо более длительном временном масштабе, чем любые другие. Изменения температуры и характера осадков будут способствовать изменениям в сообществах. По мере потепления климата в конце каждого ледникового периода происходили большие последовательные изменения. Растительность тундры и голые ледниковые отложения сменились смешанным лиственным лесом. , Парниковый эффект приводящий к повышению температуры, вероятно, приведет к глубоким аллогенным изменениям в следующем столетии. Геологические и климатические катастрофы, такие как извержения вулканов, землетрясения, лавины, метеориты, наводнения, пожары и сильный ветер, также приносят аллогенные изменения.
Механизмы
[ редактировать ]В 1916 году Фредерик Клементс опубликовал описательную теорию сукцессии и выдвинул ее как общую экологическую концепцию. [14] Его теория преемственности оказала мощное влияние на экологическую мысль. Концепцию Клементса обычно называют классической экологической теорией . [ нужна ссылка ] По мнению Клементса, преемственность — это процесс, включающий несколько этапов: [14] [ нужна страница ]
- Нудация: Сукцессия начинается с развития голого участка, называемого нудацией (нарушение). [14]
- Миграция: относится к прибытию пропагул . [14]
- Экесис: включает в себя зарождение и первоначальный рост растительности. [14]
- Конкуренция: по мере того, как растительность укореняется, растет и распространяется, различные виды начинают конкурировать за пространство, свет и питательные вещества. [14]
- Реакция: на этом этапе аутогенные изменения, такие как накопление гумуса, влияют на среду обитания, и одно растительное сообщество заменяет другое. [14]
- Стабилизация: формируется предположительно стабильное климаксное сообщество. [14]
Серальные сообщества
[ редактировать ]

Серальное сообщество — это промежуточная стадия экосистемы, приближающейся к своему кульминационному сообществу . Во многих случаях развивается более одной последовательной стадии, пока не достигаются условия кульминации. [29] Призере . - это совокупность серий, составляющих застройку территории от поверхностей, не покрытых растительностью, до кульминационного сообщества В зависимости от субстрата и климата встречаются разные серии.
Изменения в жизни животных
[ редактировать ]Теория сукцессии была разработана преимущественно ботаниками. Изучение сукцессии применительно к целым экосистемам было начато в трудах Рамона Маргалефа , а Юджина Одума публикация «Стратегии развития экосистем» считается его формальной отправной точкой. [30]
В животной жизни также происходят изменения с изменением сообществ. На лишайниковой стадии фауна скудна. В его состав входят несколько клещей, муравьев и пауков, живущих в трещинах и щелях. На разнотравной стадии фауна качественно увеличивается. Среди животных, встречающихся на этом этапе, — нематоды, личинки насекомых, муравьи, пауки, клещи и др. Популяция животных увеличивается и диверсифицируется по мере развития лесного климаксного сообщества. Фауна состоит из беспозвоночных, таких как слизни, улитки, черви, многоножки, многоножки, муравьи, клопы; и позвоночные животные, такие как белки, лисы, мыши, кроты, змеи, различные птицы, саламандры и лягушки.
Обзор исследований преемственности, проведенный Ходкинсоном и др. (2002) [31] задокументировал то, что, вероятно, впервые заметил Дарвин во время своего путешествия на корабле HMS Beagle:
Часто повторяющееся описание величественной пальмы и других благородных растений, затем птиц и, наконец, человека, овладевшего коралловыми островками, как только они образовались в Тихом океане, вероятно, не совсем верно; Боюсь, это разрушает поэзию этой истории, что насекомые и пауки, питающиеся перьями и грязью, а также паразитические насекомые и пауки должны быть первыми обитателями недавно образовавшейся океанической суши.
- К. Дарвин [32]
Эти натуралисты отмечают, что до появления автотрофов существовала пищевая сеть, образованная гетеротрофами, построенная на аллохтонных примесях мертвого органического вещества (некромассы). Работа Сайкса и Словика над вулканическими системами, такими как вулкан Касаточи на Алеутских островах (2010 г.) [33] поддерживает эту идею.
Микросуцессия
[ редактировать ]Последовательность микроорганизмов , включая грибы и бактерии, встречающиеся в микросреде обитания, известна как микросукцессия или серула. В искусственных бактериальных метасообществах подвижных штаммов на чипе было показано, что экологическая преемственность основана на компромиссе между колонизирующими и конкурентными способностями. Эксплуатировать локации или исследовать ландшафт? Escherichia coli — беглый вид, тогда как Pseudomonas aeruginosa — более медленный колонизатор, но более сильный конкурент. [7] Как и у растений, микробная сукцессия может происходить во вновь доступных средах обитания ( первичная сукцессия ), таких как поверхность листьев растений, недавно обнаженные поверхности камней (т. е. ледниковый тилль) или кишки детенышей животных. [24] а также на нарушенных сообществах ( вторичная сукцессия ), таких как те, которые растут на недавно погибших деревьях, гниющих плодах, [34] или помет животных. Микробные сообщества также могут меняться из-за продуктов, выделяемых присутствующими бактериями. Изменения pH в среде обитания могут создать идеальные условия для обитания нового вида. В некоторых случаях новые виды могут превзойти существующие виды за питательные вещества, что приведет к гибели основных видов. Изменения также могут происходить в результате микробной последовательности при изменении доступности воды и температуры. Теории макроэкологии лишь недавно стали применяться к микробиологии , и еще многое предстоит понять в этой развивающейся области. Недавнее исследование микробной последовательности оценило баланс между стохастическими солончаков и детерминированными процессами при бактериальной колонизации хронопоследовательности . Результаты этого исследования показывают, что, как и в случае макросукцессии, ранняя колонизация ( первичная сукцессия ) в основном находится под влиянием стохастичности, тогда как вторичная сукцессия этих бактериальных сообществ в большей степени находилась под влиянием детерминистических факторов. [35]

Концепция кульминации
[ редактировать ]Согласно классической экологической теории , сукцессия прекращается, когда сера достигает равновесия или устойчивого состояния с физической и биотической средой. Если не произойдет серьезных потрясений, оно будет сохраняться неопределенно долго. [1] Эта конечная точка последовательности называется кульминацией.
Климакс сообщество
[ редактировать ]Окончательное или стабильное сообщество в серии — это климаксное сообщество или климатическая растительность . Он самовоспроизводится и находится в равновесии с физической средой обитания. [1] В климаксном сообществе нет чистого годового накопления органического вещества. Годовое производство и использование энергии в таком сообществе сбалансированы.
Характеристики
[ редактировать ]- Растительность устойчива к условиям окружающей среды.
- Он имеет большое разнообразие видов, хорошо дренированную пространственную структуру и сложные пищевые цепи.
- Климаксная экосистема сбалансирована. Существует равновесие между валовой первичной продукцией и общим дыханием, между энергией, используемой от солнечного света, и энергией, выделяемой при разложении, между поглощением питательных веществ из почвы и возвратом питательных веществ с опадом в почву.
- Особи на стадии кульминации заменяются другими такими же. Таким образом, видовой состав сохраняет равновесие.
- Это показатель климата данной местности. Формы жизни или роста указывают на климатический тип.
Виды климакса
[ редактировать ]- Климатическая кульминация
- Если имеется только один кульминационный момент и развитие климаксного сообщества контролируется климатом региона, его называют климатическим кульминацией. Например, развитие кленово-букового климаксового сообщества на влажной почве. Климатический кульминационный момент является теоретическим и развивается там, где физические условия субстрата не настолько экстремальны, чтобы изменять воздействие преобладающего регионального климата.
- Эдафический кульминационный момент
- Когда в регионе существует более одного климаксного сообщества, измененного местными условиями субстрата, такими как влажность почвы, питательные вещества почвы, топография, экспозиция склонов, пожар и деятельность животных, это называется эдафическим климаксом . Сукцессия заканчивается эдафической кульминацией, когда топография, почва, вода, огонь или другие нарушения таковы, что климатическая кульминация не может развиться.
- Катастрофическая кульминация
- Кульминационная растительность уязвима для таких катастрофических событий, как лесной пожар. Например, в является последней Калифорнии растительность чапараля растительностью. Лесной пожар уничтожает зрелую растительность и разрушители. Далее следует быстрое развитие травянистой растительности, пока не восстановится преобладание кустарников. Это известно как катастрофическая кульминация.
- Дисклимакс
- Когда устойчивое сообщество, не являющееся климатической или эдафической кульминацией для данного участка, поддерживается человеком или его домашними животными, оно обозначается как дисклимакс (климакс возмущения) или антропогенный субклимакс (антропогенный). Например, чрезмерный выпас пастбищ может привести к образованию пустынного сообщества кустарников и кактусов, где местный климат фактически позволит пастбищам поддерживать себя.
- Субклимакс
- Длительная стадия, предшествующая кульминации, называется субклимаксом .
- Преклимакс и постклимакс
- На отдельных территориях в сходных климатических условиях развиваются разные климаксные сообщества. Если в сообществе есть формы жизни ниже, чем те, которые находятся в ожидаемой климатической кульминации, это называется преклимаксом ; Сообщество, в котором формы жизни выше, чем те, которые находятся в ожидаемом климатическом кульминации, является постклимаксом . Полоски преклимакса развиваются в менее влажных и более жарких районах, тогда как нити постклимакса развиваются в более влажных и прохладных местах, чем в окружающем климате.
Теории
[ редактировать ]Существует три школы интерпретаций, объясняющих концепцию кульминации:
- Теория моноклимакса или климатической кульминации была выдвинута Клементсом (1916) и признает только одну кульминацию, характеристики которой определяются исключительно климатом (климатическая кульминация). Процессы сукцессии и изменения среды преодолевают влияние различий топографии, почвенного материала и других факторов. Вся территория будет покрыта единым растительным сообществом. Сообщества, отличные от кульминации, связаны с ним и признаются субклимаксом, посткульминацией и дисклимаксом.
- Теория поликлимакса была предложена Тэнсли (1935). Он предполагает, что кульминационная растительность региона состоит из более чем одной кульминационной растительности, контролируемой влажностью почвы, питательными веществами почвы, топографией, экспозицией склонов, пожарами и деятельностью животных.
- Теория паттернов кульминации была предложена Уиттакером (1953). Теория закономерностей кульминации признает множество кульминаций, определяемых реакцией популяций видов на биотические и абиотические условия. Согласно этой теории, общая среда экосистемы определяет состав, видовую структуру и баланс климаксного сообщества. Окружающая среда включает в себя реакцию видов на влажность, температуру и питательные вещества, их биотические взаимоотношения, наличие флоры и фауны для колонизации территории, случайное распространение семян и животных, почвы, климат и такие нарушения, как огонь и ветер. Характер климаксной растительности будет меняться по мере изменения окружающей среды. Климаксное сообщество представляет собой структуру популяций, которая соответствует структуре окружающей среды и меняется вместе с ней. Центральным и наиболее распространенным сообществом является климатическая кульминация.
Теория альтернативных стабильных состояний предполагает, что существует не одна конечная точка, а множество, которые переходят друг от друга в течение экологического времени.
Сукцессия по типу среды обитания
[ редактировать ]Сукцессия леса
[ редактировать ]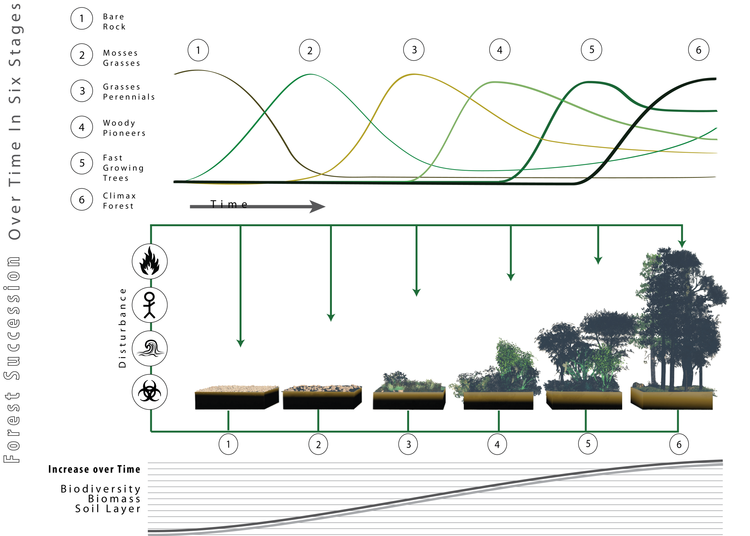
Леса, являясь экологической системой, подвержены процессу сукцессии видов. [36] Существуют «оппортунистические» или «первопроходческие» виды, которые производят большое количество семян, которые разносятся ветром и, следовательно, могут колонизировать большие пустые территории. [37] Они способны прорастать и расти под прямыми солнечными лучами. После того, как они образовали закрытый навес , отсутствие прямого солнечного излучения в почве затрудняет развитие их собственных сеянцев. Тогда у теневыносливых видов появится возможность обосноваться под защитой первопроходцев. Когда пионеры погибают, их заменяют теневыносливые виды. Эти виды способны расти под пологом, а значит, при отсутствии нарушений, сохранятся. По этой причине тогда говорят, что позиция достигла своего апогея. Когда происходит нарушение, возможности для пионеров снова открываются, при условии, что они присутствуют или находятся в разумных пределах.
Примером видов-первопроходцев в лесах северо-востока Северной Америки являются Betula papyrifera ( Белая береза ) и Prunus serotina ( Черная вишня ), которые особенно хорошо приспособлены к использованию больших промежутков в лесных пологах, но не переносят тени и в конечном итоге вытесняются. другими теневыносливыми видами при отсутствии нарушений, создающих такие просветы. В тропиках хорошо известные пионерные лесные виды можно найти среди родов Cecropia , Ochroma и Trema . [37]
В природе вещи не черно-белые, есть промежуточные стадии. Поэтому вполне нормально, что между двумя крайностями света и тени существует градиент, и существуют виды, которые могут действовать как первопроходцы или толерантные, в зависимости от обстоятельств. Чрезвычайно важно знать толерантность видов, чтобы практиковать эффективное лесоводство .
Последовательность водно-болотных угодий
[ редактировать ]Поскольку существует множество типов водно-болотных угодий, сукцессия может следовать по широкому спектру траекторий и моделей на водно-болотных угодьях. Согласно классической модели, процесс вторичной сукцессии предполагает, что водно-болотные угодья со временем развиваются от исходного состояния открытой воды с небольшим количеством растений до кульминационного состояния, покрытого лесом , когда разложившееся органическое вещество накапливается с течением времени, образуя торф . Однако многие водно-болотные угодья поддерживаются в равновесном состоянии за счет регулярных нарушений или естественных процессов, которые не напоминают предсказанный лесной «кульминационный момент». [38] Идея о том, что пруды и водно-болотные угодья постепенно превращаются в сушу, подверглась критике и была поставлена под сомнение из-за отсутствия доказательств. [5]
Сукцессия водно-болотных угодий — это уникально сложный, нелинейный процесс, определяемый гидрологией . [39] Гидрологические факторы часто действуют против линейных процессов, которые предсказывают наступление «кульминационного» состояния. Энергия, переносимая движущейся водой, может стать постоянным источником беспокойства. Например, на прибрежных водно-болотных угодьях приливы и отливы постоянно воздействуют на экологическое сообщество. Пожар также может поддерживать состояние равновесия на водно-болотных угодьях, сжигая растительность и тем самым прерывая накопление торфа. [38] Вода, поступающая на водно-болотные угодья и покидающая их, подчиняется закономерностям, которые в целом цикличны, но неустойчивы. Например, сезонные наводнения и высыхания могут происходить с ежегодными изменениями количества осадков, вызывая сезонные изменения в сообществе водно-болотных угодий, которые поддерживают его в стабильном состоянии. [5] Однако необычно сильный дождь или необычно сильная засуха могут привести к тому, что водно-болотные угодья войдут в петлю положительной обратной связи, где она начнет меняться в линейном направлении. [39] Поскольку водно-болотные угодья чувствительны к изменениям естественных процессов, которые их поддерживают, деятельность человека, инвазивные виды и изменение климата могут инициировать долгосрочные изменения в экосистемах водно-болотных угодий. [38]
Последовательность пастбищ
[ редактировать ]Долгое время считалось, что пастбища представляют собой раннюю стадию сукцессии, где преобладают сорные виды и имеют небольшую природоохранную ценность. Однако сравнение лугов, образовавшихся после восстановления после долгосрочных нарушений, таких как сельскохозяйственная обработка почвы , с древними или «старовозрастными» лугами, показало, что луга по своей сути не являются раннесукцессионными сообществами. Скорее, пастбища подвергаются многовековому процессу сукцессии, и, по оценкам, пастбищам, которые распахиваются для сельского хозяйства или уничтожаются иным образом, потребуется минимум 100 лет, а потенциально в среднем 1400 лет, чтобы восстановиться до прежнего уровня биоразнообразия. [40] Однако посадка большого разнообразия видов лугов поздней сукцессии в нарушенной окружающей среде может ускорить восстановление способности почвы улавливать углерод, что приведет к накоплению вдвое большего количества углерода, чем на естественно восстанавливающихся пастбищах за тот же период времени. [41]
Многие луговые экосистемы поддерживаются за счет нарушений, таких как пожары и выпас скота крупными животными, или в противном случае процесс сукцессии превратит их в леса или кустарники. Фактически, ведутся споры о том, следует ли вообще считать пожар нарушением экосистем прерий Северной Америки , поскольку он поддерживает, а не нарушает состояние равновесия. [42] Многие виды пастбищ поздней сукцессии имеют приспособления, которые позволяют им хранить питательные вещества под землей и быстро прорастать после «надземных» нарушений, таких как пожар или выпас скота. События, вызывающие беспокойство, которые серьезно разрушают или разрушают почву, такие как обработка почвы, уничтожают эти виды поздней сукцессии, возвращая пастбища на раннюю сукцессионную стадию, в которой доминируют пионеры , тогда как пожар и выпас приносят пользу видам поздней сукцессии. [40] Как слишком сильное, так и слишком малое вмешательство может нанести ущерб биоразнообразию зависимых от возмущений экосистем, таких как луга. [43]
На полузасушливых лугах Северной Америки введение животноводства и отсутствие пожаров привело к переходу от трав к древесной растительности, особенно мескитовой . [44] Однако способы, с помощью которых экологическая сукцессия при частых нарушениях приводит к образованию экосистем, подобных тем, которые наблюдаются в сохранившихся прериях, плохо изучены. [45] [43]
См. также
[ редактировать ]- Модель экологической сукцессии Коннелла – Слейера
- Циклическая последовательность
- Экологическая стабильность
- Гипотеза промежуточного возмущения
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Фишер М.Р. (2018). Экологическая биология . Открытые образовательные ресурсы штата Орегон.
- ^ «Суртсей» . Центр всемирного наследия ЮНЕСКО.
- ^ Фредриксен, Хелле Б.; Краглунд, Ханс-Оле; Экелунд, Флемминг (2016). «Первичная последовательность микрофауны на вулканическом острове Суртсей, Исландия» . Полярные исследования . 20 (1): 61–73. дои : 10.3402/polar.v20i1.6500 . S2CID 82682454 .
- ^ Jump up to: а б Смит С., Марк С. (январь 2009 г.). «Исторические корни охраны природы в регионе Северо-Западной Индианы и Чикаго: от науки к сохранению» . Журнал Южного Берега . 3 : 1–10.
- ^ Jump up to: а б с Миддлтон, Бет А. (2016). Сукцессия на водно-болотных угодьях . Спрингер. ISBN 978-94-007-6172-8 .
- ^ Jump up to: а б Ларсен Дж. А. (22 октября 2013 г.). Экология северных равнинных болот и хвойных лесов . Эльзевир . ISBN 9781483269863 .
- ^ Jump up to: а б с Уэтерингтон М.Т., Надь К., Дер Л., Абрахам А., Нурлаг Дж., Галайда П., Кеймер Дж.Э. (ноябрь 2022 г.). «Экологическая преемственность и компромисс между конкуренцией и колонизацией в микробных сообществах» . БМК Биология . 20 (1): 262. дои : 10.1186/s12915-022-01462-5 . ПМК 9710175 . ПМИД 36447225 .
- ^ Делюк Ж.А. (1813). Геологические путешествия по некоторым частям Франции, Швейцарии и Германии . Публичная библиотека Лиона: ФК и Дж. Ривингтон.
- ^ Jump up to: а б Торо HD, Эмерсон Р.В. (1887). Чередование лесных деревьев: и диких яблонь . Хоутон, Миффлин . Получено 12 апреля 2014 г. - через Archive.org.
- ^ Торо HD (2013). Крамер Дж.С. (ред.). Очерки: полностью аннотированное издание . Нью-Хейвен, Коннектикут: Издательство Йельского университета.
- ^ Баззаз Ф.А. (1996). Растения в изменяющихся условиях . Великобритания: Издательство Кембриджского университета. п. 3. ISBN 9-780521-398435 .
- ^ Коулз ЕС (1899). «Экологические связи растительности песчаных дюн озера Мичиган. Часть I. Географические связи дюнных флор» . Ботанический вестник . 27 (2). Издательство Чикагского университета: 95–117. дои : 10.1086/327796 . S2CID 84315469 .
- ^ Шонс М. «Генри Чендлер Коулз» . Нэшнл Географик. Архивировано из оригинала 21 февраля 2013 года . Проверено 25 июня 2014 г.
- ^ Jump up to: а б с д и ж г час я Клементс Ф.Е. (1916). Сукцессия растений: анализ развития растительности . Институт Карнеги в Вашингтоне.
- ^ Глисон Х.А. (январь 1926 г.). «Индивидуалистическая концепция растительного объединения». Бюллетень Ботанического клуба Торри . 53 (1): 7–26. дои : 10.2307/2479933 . JSTOR 2479933 .
- ^ Коулз ХК (1911). «Причины вегетационных циклов» . Анналы Ассоциации американских географов . 1 (1): 3–20. дои : 10.2307/2560843 . JSTOR 2560843 .
- ^ Jump up to: а б Кристенсен, Норман Л. (2014). «Исторический взгляд на сукцессию лесов и ее значение для практики восстановления и сохранения экосистем в Северной Америке». Лесная экология и управление . 330 : 312–322. дои : 10.1016/j.foreco.2014.07.026 .
- ^ Jump up to: а б Аноми, Кеннет А.; Нири, Брэд; Чен, Цзясинь; Мэр Стивен Дж. (2022 г.). «Критический обзор сукцессионной динамики в бореальных лесах Северной Америки» . Экологические обзоры . 30 (4): 563–594. дои : 10.1139/er-2021-0106 . S2CID 247965093 .
- ^ Баззаз Ф (1996). Растения в изменяющихся условиях . Великобритания: Издательство Кембриджского университета. стр. 4–5. ISBN 9-780521-398435 .
- ^ Карлтон, Джеймс Т. (1 октября 1996 г.). «Схема, процесс и прогноз в экологии морских инвазий» . Биологическая консервация . Биология вторжения. 78 (1): 97–106. дои : 10.1016/0006-3207(96)00020-1 . ISSN 0006-3207 .
- ^ Фроуз, Ян (01 февраля 2024 г.). «Обратная связь между растениями и почвой в пространственно-временных масштабах: от немедленных эффектов до наследия» . Биология и биохимия почвы . 189 : 109289. doi : 10.1016/j.soilbio.2023.109289 . ISSN 0038-0717 .
- ^ Баззаз Ф.А. (1996). Растения в изменяющихся условиях . Великобритания: Издательство Кембриджского университета. п. 1. ISBN 9-780521-398435 .
- ^ Эмери, С. (2010). «Преемственность: более пристальный взгляд» . Природа .
- ^ Jump up to: а б Ортис-Альварес Р., Фьерер Н., де Лос Риос А., Касамайор Э.О., Барберан А. (июнь 2018 г.). «Последовательные изменения таксономической структуры и функциональных признаков бактериальных сообществ в ходе первичной сукцессии» . Журнал ISME . 12 (7): 1658–1667. дои : 10.1038/s41396-018-0076-2 . ПМК 6018800 . ПМИД 29463893 .
- ^ Jump up to: а б Редакторы биологического словаря (31 января 2017 г.). «Экологическая сукцессия – определение, виды и примеры» . Биологический словарь . Проверено 8 мая 2019 г.
- ^ Кук В.М., Яо Дж., Фостер Б.Л., Холт Р.Д., Патрик Л.Б. «Вторичная преемственность в экспериментально фрагментированном ландшафте: модели сообщества в пространстве и времени» . Министерство сельского хозяйства США . Проверено 30 сентября 2013 г.
- ^ Баниски С. (3 июля 1995 г.). «Наводнение меняет облик парка Шенандоа» . Балтимор Сан . Проверено 5 июля 2019 г.
- ^ Ван дер Путтен, Вашингтон; Мортимер, СР; Хедлунд, К.; Ван Дейк, К.; Браун, В.К.; Лепа, Дж.; Родригес-Барруэко, К.; Рой, Дж.; Диас Лен, штат Техас; Гормсен, Д.; Кортальс, ГВ; Лаворель, С.; Регина, И. Санта; Смилауэр, П. (1 июля 2000 г.). «Разнообразие видов растений как фактор ранней сукцессии на заброшенных полях: подход с участием нескольких участков» . Экология . 124 (1): 91–99. Бибкод : 2000Экол.124...91В . дои : 10.1007/s004420050028 . ISSN 1432-1939 . ПМИД 28308417 . S2CID 38703575 .
- ^ Майкл Г. Барбур и Уильям Дуайт Биллингс (2000) Наземная растительность Северной Америки , Cambridge University Press, 708 страниц. ISBN 0-521-55986-3 , ISBN 978-0-521-55986-7
- ^ Баззаз Ф.А. (1996). Растения в меняющейся среде . Издательство Кембриджского университета. п. 4. ISBN 9-780521-398435 .
- ^ Ходкинсон, ID; Уэбб, Северная Каролина; Коулсон, С.Дж. (2002). «Первичная сборка сообщества на суше – недостающие стадии: почему гетеротрофные организмы всегда появляются первыми?» . Журнал экологии . 90 (3): 569-577 . Проверено 2 августа 2024 г.
- ^ Дарвин, Чарльз (1845). Журнал исследований естествознания и геологии стран, посещенных во время кругосветного путешествия HMS Beagle под командованием капитана Фитца Роя, RN 2-е издание (2-е изд.). Лондон: Джон Мюррей . Проверено 2 августа 2024 г.
- ^ Сайкс, Дерек С.; Словик, Йозеф (2010). «Наземные членистоногие острова Касаточи до и после извержения, Аляска, 2008–2009 годы: переход от растительной пищевой сети к некромассе» . Арктические, антарктические и альпийские исследования . 42 (3): 297–305 . Проверено 2 августа 2024 г.
- ^ Мартин П.Л., Кинг В., Белл Т.Х., Питер К. (2021). «Гниль и грибковая сукцессия яблок с горькой гнилью по градиенту разнообразия растительности» . Журнал «Фитобиомы» . 6 : PBIOMES-06-21-0039-R. doi : 10.1094/PBIOMES-06-21-0039-R . ISSN 2471-2906 . S2CID 239658496 .
- ^ Дини-Андреоте Ф., Стеген Х.К., ван Эльсас Дж.Д., Саллес Х.Ф. (март 2015 г.). «Распутывание механизмов, которые обеспечивают баланс между стохастическими и детерминированными процессами в микробной последовательности» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (11): Е1326–Е1332. Бибкод : 2015PNAS..112E1326D . дои : 10.1073/pnas.1414261112 . ПМК 4371938 . ПМИД 25733885 .
- ^ МакЭвой Т. (2004). «Положительное воздействие лесного хозяйства». Сукцессия видов и толерантность . Остров Пресс. п. 32.
- ^ Jump up to: а б Будовский Г (1965). «Распространение видов тропических лесов Америки в свете сукцессионных процессов». Турриальба . 15 (1): 40–42.
- ^ Jump up to: а б с Мозли, Кендра. «Экология водно-болотных угодий. Основные принципы» (PDF) . Министерство сельского хозяйства США.
- ^ Jump up to: а б Цвейг, CL; Кухни, WM (2009). «Смена нескольких штатов на водно-болотных угодьях: новое использование моделей состояний и переходов» . Экология . 90 (7): 1900–1909. дои : 10.1890/08-1392.1 . hdl : 1834/22259 . ПМИД 19694138 .
- ^ Jump up to: а б Нерлекар, Ашиш Н.; Вельдман, Джозеф В. (2020). «Высокое разнообразие растений и медленное зарастание старовозрастных лугов» . ПНАС . 117 (31): 18550–18556. дои : 10.1073/pnas.1922266117 . ПМЦ 7414179 . ПМИД 32675246 .
- ^ Ян, И; Тилман, Дэвид; Фьюри, Джордж; Леман, Кларенс (2019). «Связывание углерода почвой ускоряется за счет восстановления биоразнообразия пастбищ» . Природные коммуникации . 10 (718): 718. doi : 10.1038/s41467-019-08636-w . ПМК 6372642 . ПМИД 30755614 .
- ^ Эванс, EW; Бриггс, Дж. М.; Финк, Э.Дж.; Гибсон, диджей; Джеймс, Юго-Запад; Кауфман, Д.В.; Систедт, Т.Р. «Является ли пожар помехой для лугов?» (PDF) .
- ^ Jump up to: а б Шнор, Тим; Брюун, Ханс Хенрик; Олссон, Пал Аксель (2015). «Нарушение почвы как мера восстановления пастбищ - влияние на видовой состав растений и функциональные особенности растений» . ПЛОС ОДИН . 10 (4): e0123698. Бибкод : 2015PLoSO..1023698S . дои : 10.1371/journal.pone.0123698 . ПМЦ 4395216 . ПМИД 25875745 .
- ^ Ван Аукен, Огайо (2000). «Вторжение кустарников на полузасушливые луга Северной Америки». Ежегодный обзор экологии и систематики . 31 : 197–215. doi : 10.1146/annurev.ecolsys.31.1.197 .
- ^ Бомбергер, Мэри Л.; Шилдс, Шелли; Харрисон, Л. Тайрон; Киллер, Кэтлин. «Сравнение старой полевой последовательности в прерии высокой травы и прерии песчаных холмов Небраски» .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь )
Дальнейшее чтение
[ редактировать ]- Коннелл Дж. Х. , Слейер Р. О. (1977). «Механизмы сукцессии в природных сообществах и их роль в стабильности и организации сообществ». Американский натуралист . 111 (982): 1119–44. дои : 10.1086/283241 . S2CID 3587878 .
- Фроуз Дж., Прах К., Пижл В., Ханель Л., Старый Дж., Таёвский К. и др. (2008). «Взаимодействие между развитием почвы, растительностью и почвенной фауной во время спонтанной сукцессии на участках после добычи». Европейский журнал почвенной биологии . 44 : 109–121. дои : 10.1016/j.ejsobi.2007.09.002 .
Внешние ссылки
[ редактировать ]- Научное пособие: Преемственность. Объяснение преемственности для старшеклассников.
- Биографический очерк Генри Чендлера Коулза.
- Роберт Мерфи видит в значительной степени идеологическую, а не научную основу для неприятия преемственности со стороны нынешней экологической ортодоксальности и стремится восстановить преемственность с помощью целостных и телеологических аргументов.