По разным оценкам, морские микроорганизмы составляют около 70%. [4] или около 90%, [5] [6] биомассы в океане . Вместе они образуют морской микробиом . За миллиарды лет этот микробиом развил множество образов жизни и адаптаций и стал участвовать в глобальном круговороте почти всех химических элементов. [7] Микроорганизмы играют решающую роль в переработке питательных веществ в экосистемах, поскольку они действуют как разлагатели . Они также ответственны почти за весь фотосинтез , происходящий в океане, а также за круговорот углерода , азота , фосфора и других питательных веществ и микроэлементов. [8] Морские микроорганизмы улавливают большое количество углерода и производят большую часть мирового кислорода.
Небольшая часть морских микроорганизмов являются патогенными , вызывая болезни и даже смерть морских растений и животных. [9] Однако морские микроорганизмы перерабатывают основные химические элементы , производя и потребляя ежегодно около половины всего органического вещества, образующегося на планете. Будучи обитателями крупнейшей окружающей среды на Земле, морские микробные системы вызывают изменения во всех глобальных системах.
В июле 2016 года ученые сообщили об идентификации набора из 355 генов последнего универсального общего предка (LUCA) всей жизни на планете, включая морские микроорганизмы. [10] Несмотря на свое разнообразие, микроскопическая жизнь в океанах до сих пор плохо изучена. Например, роль вирусов в морских экосистемах практически не изучалась даже в начале XXI века. [11]
Микробные маты — самая ранняя форма жизни на Земле, существование которой подтверждено убедительными ископаемыми свидетельствами. На изображении изображен цианобактериально -водорослевой мат.
Строматолиты образуются из микробных матов, когда микробы медленно движутся вверх, чтобы не быть задушенными осадком.
Относительные размеры микроскопических образований
Микроорганизмы играют решающую роль в переработке питательных веществ в экосистемах, поскольку они действуют как разлагатели . Некоторые микроорганизмы являются патогенными , вызывая заболевания и даже смерть растений и животных. [9] Будучи обитателями крупнейшей окружающей среды на Земле, морские микробные системы вызывают изменения во всех глобальных системах. Микробы ответственны практически за весь фотосинтез , происходящий в океане, а также за круговорот углерода , азота , фосфора и других питательных веществ и микроэлементов. [8]
Хотя последние технологические разработки и научные открытия были значительными, нам все еще не хватает глубокого понимания на всех уровнях основных экологических вопросов, касающихся микроорганизмов в наших морях и океанах. Эти фундаментальные вопросы таковы: 1. Что там? Какие микроорганизмы присутствуют в наших морях и океанах и в каком количестве?они происходят? 2. Что они делают? Какие функции каждый из этих микроорганизмов выполняет в морской среде и какой вклад они вносят в глобальные круговороты энергии и вещества? 3. Какие факторы определяют наличие или отсутствие микроорганизма и как они влияют на биоразнообразие и функции и наоборот?
Диапазон размеров прокариот (бактерий и архей) и вирусов по сравнению с размерами других организмов и биомолекул. Под лупой брызги морской воды изобилуют микроскопической жизнью
Подводная микроскопическая жизнь разнообразна и до сих пор плохо изучена, например, роль вирусов в морских экосистемах. [13] Большинство морских вирусов представляют собой бактериофаги , которые безвредны для растений и животных, но необходимы для регулирования морских и пресноводных экосистем. [14] Они заражают и уничтожают бактерии в водных микробных сообществах и являются важнейшим механизмом переработки углерода в морской среде. Органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост новых бактерий и водорослей. [15] Вирусная активность также может способствовать биологическому насосу — процессу, посредством которого в глубинах углерод улавливается океана. [16]
Морские брызги, содержащие морские микроорганизмы, могут быть унесены высоко в атмосферу, где они станут аэропланктоном , и могут путешествовать по земному шару, прежде чем упасть обратно на Землю.
Поток переносимых по воздуху микроорганизмов кружит над планетой над погодными системами, но под коммерческими воздушными путями. [17] Некоторые странствующие микроорганизмы переносятся земными пыльными бурями, но большинство происходит из морских микроорганизмов, содержащихся в морских брызгах . В 2018 году ученые сообщили, что сотни миллионов вирусов и десятки миллионов бактерий ежедневно оседают на каждом квадратном метре планеты. [18] [19]
Микроскопические организмы обитают по всей биосфере . Масса прокариотных микроорганизмов, в которую входят бактерии и археи, но не ядросодержащие эукариотные микроорганизмы , может достигать 0,8 триллиона тонн углерода (от общей массы биосферы , оцениваемой от 1 до 4 триллионов тонн). [20] Одноклеточные барофильные морские микробы были обнаружены на глубине 10 900 м (35 800 футов) в Марианской впадине , самом глубоком месте мирового океана. [21] [22] Микроорганизмы живут внутри скал на глубине 580 м (1900 футов) ниже морского дна, на глубине 2590 м (8500 футов) океана у побережья северо-запада США . [21] [23] а также на глубине 2400 м (7900 футов; 1,5 мили) под морским дном у берегов Японии. [24] Самая высокая известная температура, при которой может существовать микробная жизнь, составляет 122 ° C (252 ° F) ( Methanopyrus kandleri ). [25] В 2014 году учёные подтвердили существование микроорганизмов, живущих на глубине 800 м (2600 футов) подо льдом Антарктиды . [26] [27] По словам одного исследователя: «Микробы можно найти повсюду — они чрезвычайно адаптируются к условиям и выживают, где бы они ни находились». [21] Морские микроорганизмы служат «основой всех морских пищевых сетей, перерабатывая основные элементы и производя и потребляя около половины органического вещества, образующегося на Земле каждый год». [28] [29]
Вирусы, находящиеся вне инфицированной клетки или в процессе заражения клетки, существуют в виде независимых частиц. Эти вирусные частицы, также известные как вирионы , состоят из двух или трех частей: (i) генетического материала (генома), состоящего из ДНК или РНК , длинных молекул , несущих генетическую информацию; (ii) белковая оболочка, называемая капсидом , которая окружает и защищает генетический материал; и в некоторых случаях (iii) оболочка из липидов , окружающая белковую оболочку, когда они находятся вне клетки. Формы этих вирусных частиц варьируются от простых спиральных и икосаэдрических форм для некоторых видов вирусов до более сложных структур для других. Большинство видов вирусов имеют вирионы, которые слишком малы, чтобы их можно было увидеть в оптический микроскоп . Средний вирион составляет примерно одну сотую размера средней бактерии .
Происхождение вирусов в эволюционной истории жизни неясно: некоторые, возможно, произошли от плазмид — фрагментов ДНК, которые могут перемещаться между клетками, — тогда как другие, возможно, произошли от бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , увеличивающего генетическое разнообразие . [32] Некоторые считают вирусы формой жизни, поскольку они несут генетический материал, размножаются и развиваются посредством естественного отбора . Однако им не хватает ключевых характеристик (таких как клеточная структура), которые обычно считаются необходимыми для того, чтобы считаться жизнью. Поскольку вирусы обладают некоторыми, но не всеми такими качествами, их называют «организмами на грани жизни». [33] и как репликаторы. [34]
Бактериофаги (фаги)
Множественные фаги, прикрепленные к клеточной стенке бактерий, при увеличении в 200 000 раз.
Вирусы встречаются везде, где есть жизнь, и, вероятно, существовали с момента появления живых клеток. [35] Происхождение вирусов неясно, поскольку они не образуют окаменелостей, поэтому молекулярные методы использовались для сравнения ДНК или РНК вирусов и являются полезным средством изучения того, как они возникли. [36]
Сегодня вирусы признаны древними и имеют происхождение, предшествовавшее расколу жизни на три домена . [37]
Мнения относительно того, являются ли вирусы формой жизни или органическими структурами, взаимодействующими с живыми организмами, расходятся. [34] Некоторые считают их формой жизни, поскольку они несут генетический материал, размножаются, создавая множество своих копий посредством самосборки, и развиваются посредством естественного отбора . Однако им не хватает ключевых характеристик, таких как клеточная структура, которая обычно считается необходимой для того, чтобы считаться жизнью. Поскольку вирусы обладают некоторыми, но не всеми такими качествами, их называют репликаторами. [34] и как «организмы на грани жизни». [33]
Бактериофаги , часто называемые просто фагами , представляют собой вирусы, паразитирующие на бактериях и археях. Морские фаги паразитируют на морских бактериях и археях, таких как цианобактерии . [38] Они представляют собой распространенную и разнообразную группу вирусов и являются наиболее распространенным биологическим объектом в морской среде, поскольку их хозяева, бактерии, обычно представляют собой численно доминирующую клеточную жизнь в море. Обычно в каждом мл морской воды содержится от 1 до 10 миллионов вирусов, или примерно в десять раз больше вирусов с двухцепочечной ДНК, чем клеточных организмов. [39] [40] хотя оценки численности вирусов в морской воде могут варьироваться в широком диапазоне. [41] [42] Долгое время хвостатые фаги отряда Caudovirales, казалось, доминировали в морских экосистемах по численности и разнообразию организмов. [38] Однако в результате более поздних исследований бесхвостые вирусы, по-видимому, доминируют на многих глубинах и в океанических регионах, за ними следуют семейства миовирусов Caudovirales , подовирусов и сифовирусов. [43] Фаги, принадлежащие к семействам: Кортиковирусы , [44] иновирусиды , [45] Микровирусы , [46] и автоликивирусиды [47] [48] [49] [50] Также известно, что они заражают разнообразные морские бактерии.
Существуют также архейные вирусы, которые реплицируются внутри архей : это вирусы с двухцепочечной ДНК необычной, а иногда и уникальной формы. [51] [52] Наиболее подробно эти вирусы изучены на термофильных археях, в частности на порядках Sulfolobales и Thermoproteales . [53]
Микроорганизмы составляют около 70% морской биомассы. [4] По оценкам, вирусы убивают 20% этой биомассы каждый день, а в океанах вирусов в 15 раз больше, чем бактерий и архей. Вирусы являются основными агентами, ответственными за быстрое уничтожение вредоносного цветения водорослей . [40] которые часто убивают других морских обитателей. [54] Число вирусов в океанах уменьшается по мере удаления от берега и глубже в воду, где меньше организмов-хозяев. [16]
Вирусы являются важным естественным средством передачи генов между различными видами, что увеличивает генетическое разнообразие и стимулирует эволюцию. [32] Считается, что вирусы играли центральную роль в ранней эволюции, до появления бактерий, архей и эукариот, во времена последнего универсального общего предка жизни на Земле. [55] Вирусы по-прежнему являются одним из крупнейших резервуаров неизведанного генетического разнообразия на Земле. [16]
Вирусы обычно имеют длину от 20 до 300 нанометров. Это можно сравнить с длиной бактерий, которая начинается примерно с 400 нанометров. Существуют также гигантские вирусы , часто называемые гирусами , обычно длиной около 1000 нанометров (один микрон). Все гигантские вирусы относятся к типу Nucleocytoviricota (NCLDV) вместе с поксвирусами .Самый крупный известный из них — Тупанвирус . Этот род гигантских вирусов был обнаружен в 2018 году в глубоком океане, а также в содовом озере, и его общая длина может достигать 2,3 микрона. [56]
Открытие и последующая характеристика гигантских вирусов вызвали споры относительно их эволюционного происхождения. [57] Две основные гипотезы их происхождения заключаются в том, что либо они произошли от небольших вирусов, перехвативших ДНК от организмов-хозяев, либо они эволюционировали из очень сложных организмов в современную форму, которая не является самодостаточной для размножения. [58] От какого сложного организма могли отделиться гигантские вирусы, также является предметом споров. Одно из предположений состоит в том, что точка происхождения на самом деле представляет собой четвертую область жизни, [59] [60] но это было в значительной степени обесценено. [61] [62]
Предками современных бактерий были одноклеточные микроорганизмы, которые были первыми формами жизни , появившимися на Земле около 4 миллиардов лет назад. На протяжении примерно 3 миллиардов лет большинство организмов были микроскопическими, а доминирующими формами жизни были бактерии и археи. [65] [66] Хотя бактериальные окаменелости существуют, такие как строматолиты , отсутствие у них отличительной морфологии не позволяет использовать их для изучения истории эволюции бактерий или для определения времени происхождения определенного вида бактерий. Однако последовательности генов могут быть использованы для реконструкции бактериальной филогении , и эти исследования показывают, что бактерии сначала отделились от архейной/эукариотической линии. [67] Бактерии также участвовали во втором великом эволюционном расхождении – археях и эукариотах. Здесь эукариоты возникли в результате вступления древних бактерий в эндосимбиотические ассоциации с предками эукариотических клеток, которые сами, возможно, были родственны археям . [68] [69] Это включало поглощение протоэукариотическими клетками альфапротеобактериальных симбионтов с образованием либо митохондрий , либо гидрогеносом , которые до сих пор встречаются у всех известных эукариев. Позже некоторые эукариоты, уже содержащие митохондрии, также поглотили цианобактериоподобные организмы. Это привело к образованию хлоропластов у водорослей и растений. Есть также некоторые водоросли, возникшие в результате еще более поздних эндосимбиотических событий. Здесь эукариоты поглотили эукариотические водоросли, которые развились в пластиду «второго поколения». [70] [71] Это известно как вторичный эндосимбиоз .
Бактерия Marinomonas arctica растет внутри арктического морского льда при минусовых температурах.
Pelagibacter ubique и его родственники, возможно, являются наиболее распространенными организмами в океане, и утверждается, что они, возможно, являются самыми распространенными бактериями в мире. Они составляют около 25% всех клеток микробного планктона , а летом они могут составлять примерно половину клеток, присутствующих в поверхностных водах океана умеренного пояса. Общая численность P. ubique и его родственников оценивается примерно в 2 × 10 особей. 28 микробы. [73] сообщил, журнал Nature Однако в феврале 2013 года что был обнаружен бактериофаг HTVC010P , атакующий P. ubique , и «вероятно, это действительно самый распространенный организм на планете». [74] [75]
Первоначально архей считали экстремофилами , живущими в суровых условиях, как, например, желтые археи, изображенные здесь в горячем источнике , но с тех пор они были обнаружены в гораздо более широком диапазоне сред обитания . [78]
Первоначально архей относили к бактериям , но эта классификация устарела. [80] Клетки архей обладают уникальными свойствами, отличающими их от двух других областей жизни: бактерий и эукариот . Археи далее делятся на несколько признанных типов . Классификация сложна, поскольку большинство из них не были изолированы в лаборатории и были обнаружены только путем анализа их нуклеиновых кислот в образцах из окружающей среды.
Археи особенно многочисленны в океанах, а археи в планктоне могут быть одной из самых многочисленных групп организмов на планете. Археи являются важной частью жизни Земли и могут играть роль как в углеродном, так и в азотном цикле . Термопротеоты (также известные как эоциты или кренархеоты) — это тип архей, который, как считается, очень распространен в морской среде и является одним из основных участников фиксации углерода. [82]
Эоциты могут быть самыми многочисленными среди морских архей.
Галобактерии , обнаруженные в воде, почти насыщенной солью, теперь признаны архей.
Все живые организмы можно разделить на прокариотов и эукариотов . Жизнь зародилась как одноклеточные прокариоты, а затем развилась в более сложные эукариоты. В отличие от прокариотических клеток, эукариотические клетки высокоорганизованы. Прокариоты — это бактерии и археи, а эукариоты — другие формы жизни — протисты , растения, грибы и животные. Протисты обычно одноклеточные, тогда как растения, грибы и животные обычно многоклеточные .
Кажется очень правдоподобным, что корни эукариот лежат внутри архей; Ближайшими известными в настоящее время родственниками могут быть тип Heimdallarchaeota предполагаемого супертипа Асгарда . Эта теория представляет собой современную версию сценария, первоначально предложенного в 1984 году как гипотеза эоцитов , когда Thermoproteota были тогда ближайшими известными архейными родственниками эукариот.Возможную переходную форму микроорганизма между прокариотом и эукариотом обнаружили в 2012 году японские учёные. Parakaryon myojinensis — уникальный микроорганизм крупнее типичного прокариота, но с ядерным материалом, заключенным в мембрану, как у эукариот, и наличием эндосимбионтов . Считается, что это первая вероятная эволюционная форма микроорганизма, демонстрирующая стадию развития от прокариота к эукариоту. [83] [84]
Протисты — это эукариоты, которых нельзя отнести к растениям, грибам или животным. Обычно они одноклеточные и микроскопические. Жизнь зародилась как одноклеточные прокариоты (бактерии и археи), а затем развилась в более сложные эукариоты . Эукариоты — это более развитые формы жизни, известные как растения, животные, грибы и протисты. Термин «протист» исторически вошёл в употребление как удобный термин для обозначения эукариот, которых нельзя строго отнести к растениям, животным или грибам. Они не являются частью современной кладистики, поскольку являются парафилетическими (не имеют общего предка).
Простейших можно условно разделить на четыре группы в зависимости от того, какое у них питание: растительное, животное, грибковое и [85] или их смесь. [86]
Протисты — это очень разнообразные организмы, которые в настоящее время разделены на 18 типов, но их нелегко классифицировать. [89] [90] Исследования показали, что большое разнообразие протистов существует в океанах, глубоких морских жерлах и речных отложениях, что позволяет предположить, что большое количество эукариотических микробных сообществ еще не обнаружено. [91] [92] протистов мало исследовали Миксотрофных , но недавние исследования в морской среде показали, что миксотрофные протесты составляют значительную часть биомассы протистов . [87] Поскольку протисты являются эукариотами, они обладают внутри своей клетки по крайней мере одним ядром , а также такими органеллами, как митохондрии и тельца Гольджи . Протисты бесполы, но могут быстро размножаться посредством митоза или фрагментации .
Одноклеточные и микроскопические протисты
Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [93]
В отличие от клеток прокариот клетки эукариот высокоорганизованы. Растения, животные и грибы обычно многоклеточные и обычно макроскопические . Большинство протистов одноклеточные и микроскопические. Но есть исключения. Некоторые одноклеточные морские простейшие макроскопичны. Некоторые морские слизевики имеют уникальный жизненный цикл, включающий переключение между одноклеточными, колониальными и многоклеточными формами. [95] Другие морские протисты не являются ни одноклеточными, ни микроскопическими, например морские водоросли .
Другой способ классификации протистов — по способу их передвижения. Многие одноклеточные протисты, особенно простейшие, подвижны и могут совершать движения с помощью жгутиков , ресничек или ложноножек . Клетки, использующие жгутики для движения, обычно называются жгутиками , клетки, использующие реснички, обычно называются инфузориями , а клетки, использующие ложноножки, обычно называются амебами или амебоидами . Другие протисты не подвижны и, следовательно, не имеют механизма движения.
Жгутик хлыст (лат. « » ) — это похожий на плеть придаток, который выступает из тела клетки некоторых простейших (а также некоторых бактерий). Жгутиконосцы используют от одного до нескольких жгутиков для передвижения, а иногда и как органеллы питания и чувств .
Ресничка ресница (лат. ) — это крошечный жгутик. Инфузории используют несколько ресничек, число которых может исчисляться сотнями, чтобы перемещаться по воде.
К жгутиконосцам относятся бактерии, а также простейшие. Модель роторного двигателя, используемая бактериями, использует протоны электрохимического градиента для перемещения своих жгутиков. Крутящий момент в жгутиках бактерий создается частицами, которые проводят протоны вокруг основания жгутика. Направление вращения жгутиков у бактерий обусловлено заполненностью протонных каналов по периметру жгутикового мотора. [107]
Инфузории обычно имеют от сотен до тысяч ресничек, которые плотно упакованы в массивы. Во время движения отдельная ресничка деформируется с помощью рабочего хода с высоким коэффициентом трения, за которым следует восстановительный ход с низким коэффициентом трения. Поскольку в отдельном организме имеется множество ресничек, упакованных вместе, они демонстрируют коллективное поведение в метахрональном ритме . Это означает, что деформация одной реснички синфазна с деформацией соседней, вызывая волны деформации, распространяющиеся по поверхности организма. Эти распространяющиеся волны ресничек позволяют организму скоординировано использовать реснички для движения. Типичным примером реснитчатого микроорганизма является Paramecium , одноклеточное реснитчатое простейшее, покрытое тысячами ресничек. Сбивающиеся вместе реснички позволяют Paramecium двигаться в воде со скоростью 500 микрометров в секунду. [108]
более 1500 видов грибов . Из морской среды известно [109] Они паразитируют на морских водорослях или животных или представляют собой сапробы, питающиеся мертвым органическим веществом водорослей, кораллов, цист простейших, морских трав и других субстратов. [110] Споры многих видов имеют специальные придатки, облегчающие прикрепление к субстрату. [111] Морские грибы также можно найти в морской пене и вокруг гидротермальных зон океана. [112] разнообразный спектр необычных вторичных метаболитов . Морские грибы производят [113]
Типичный миллилитр морской воды содержит около 10 3 до 10 4 грибковые клетки. [119] Это число больше в прибрежных экосистемах и эстуариях из-за стока питательных веществ от наземных сообществ. Более высокое разнообразие микопланктона наблюдается вокруг побережий и в поверхностных водах на глубине до 1000 метров, вертикальный профиль которого зависит от обилия фитопланктона . [120] [121] Этот профиль меняется в зависимости от сезона из-за изменений в доступности питательных веществ. [122] Морские грибы выживают в среде с постоянным дефицитом кислорода и, следовательно, зависят от диффузии кислорода за счет турбулентности и кислорода, вырабатываемого фотосинтезирующими организмами . [123]
Лишайники — это мутуалистические ассоциации между грибом, обычно аскомицетом , и водорослью или цианобактерией . Некоторые лишайники встречаются в морской среде. [124] Еще больше их встречается в зоне всплеска , где они занимают разные вертикальные зоны в зависимости от того, насколько они устойчивы к погружению. [125] Некоторые лишайники живут долго; возраст одного вида составляет 8600 лет. [126] Однако продолжительность их жизни трудно измерить, поскольку определение одного и того же лишайника не является точным. [127] Лишайники растут путем вегетативного отрыва части, которая может быть определена, а может и не быть определена как один и тот же лишайник, и два лишайника разного возраста могут сливаться, что поднимает вопрос о том, является ли это одним и тем же лишайником. [127]
Морская улитка Littoraria irrorata повреждает растения Спартины в морских болотах, где она обитает, что позволяет спорам литоральных аскомицетовых грибов колонизировать растение. Затем улитка поедает грибковые наросты, а не саму траву. [128]
Согласно летописям окаменелостей, грибы относятся к позднему протерозою 900-570 миллионов лет назад. Ископаемые морские лишайники возрастом 600 миллионов лет были обнаружены в Китае. [129] Была выдвинута гипотеза, что микопланктон произошел от наземных грибов, вероятно, в палеозойскую эру (390 миллионов лет назад). [130]
Составное изображение, показывающее глобальное распространение фотосинтеза, включая океанический фитопланктон и наземную растительность . Темно-красный и сине-зеленый обозначают области высокой фотосинтетической активности в океане и на суше соответственно.
Первичные продуценты — это автотрофные организмы, которые производят себе пищу, а не поедают другие организмы. Это означает, что первичные продуценты становятся отправной точкой в пищевой цепи для гетеротрофных организмов, которые поедают другие организмы. Некоторые морские первичные продуценты представляют собой специализированные бактерии и археи, которые являются хемотрофами , добывающими себе пищу, собираясь вокруг гидротермальных источников и холодных выходов и используя хемосинтез . Однако большая часть первичной морской продукции происходит от организмов, которые используют фотосинтез на углекислом газе, растворенном в воде. Этот процесс использует энергию солнечного света для преобразования воды и углекислого газа. [133] : 186–187 в сахара, которые можно использовать как источник химической энергии, так и органических молекул, входящих в состав структурных компонентов клеток. [133] : 1242 Морские первичные производители важны, потому что они лежат в основе почти всей жизни морских животных, генерируя большую часть кислорода и пищи, которые обеспечивают другие организмы химической энергией, необходимой им для существования.
Цианобактерии были первыми организмами, развившими способность превращать солнечный свет в химическую энергию. Они образуют тип (подразделение) бактерий, который варьируется от одноклеточных до нитчатых и включает колониальные виды . Они встречаются практически повсюду на земле: во влажной почве, как в пресноводной, так и в морской среде и даже на антарктических скалах. [136] В частности, некоторые виды встречаются в виде дрейфующих клеток, плавающих в океане, и поэтому были одними из первых представителей фитопланктона .
Первыми первичными продуцентами, использовавшими фотосинтез, были океанические цианобактерии около 2,3 миллиарда лет назад. [137] [138] Выделение молекулярного кислорода цианобактериями как побочного продукта фотосинтеза вызвало глобальные изменения в окружающей среде Земли. Поскольку в то время кислород был токсичен для большей части жизни на Земле, это привело к почти исчезновению организмов, не переносящих кислород , — резкое изменение , которое изменило направление эволюции основных видов животных и растений. [139]
Крошечная (0,6 мкм ) морская цианобактерия Prochromococcus , открытая в 1986 году, сегодня составляет важную часть основы пищевой цепи океана и отвечает за большую часть фотосинтеза в открытом океане. [140] и примерно 20% кислорода в атмосфере Земли. [141] Возможно, это самый многочисленный род на Земле: один миллилитр поверхностной морской воды может содержать 100 000 и более клеток. [142]
Первоначально биологи считали цианобактерии водорослями и называли их «сине-зелеными водорослями». Более поздняя точка зрения состоит в том, что цианобактерии — это бактерии и, следовательно, даже не относятся к тому же царству , что и водоросли. Большинство авторов исключают всех прокариотов и, следовательно, цианобактерий из определения водорослей. [143] [144]
Динофлагелляты и диатомеи являются важными компонентами морских водорослей и имеют отдельные разделы ниже. Эвгленофиты — это тип одноклеточных жгутиконосцев, состоящий всего из нескольких морских представителей.
Не все водоросли микроскопические. Зеленые, красные и бурые водоросли имеют многоклеточные макроскопические формы, которые составляют знакомые морские водоросли . Зеленые водоросли , неофициальная группа, насчитывает около 8000 признанных видов. [145] Многие виды живут большую часть своей жизни в виде одиночных клеток или являются нитевидными, тогда как другие образуют колонии, состоящие из длинных цепочек клеток, или представляют собой высокодифференцированные макроскопические водоросли. Красные водоросли — (спорный) тип, насчитывающий около 7000 признанных видов. [146] в основном многоклеточные и включают множество известных морских водорослей. [146] [147] Бурые водоросли образуют класс, содержащий около 2000 признанных видов. [148] в основном многоклеточные и включают множество морских водорослей, таких как ламинария .В отличие от высших растений, у водорослей отсутствуют корни, стебли и листья. По размеру их можно разделить на микроводоросли и макроводоросли .
Микроводоросли — это микроскопические виды водорослей, невидимые невооруженным глазом. В основном это одноклеточные виды, существующие по отдельности, цепочками или группами, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских протистов, обсуждавшихся выше , а также фитопланктона, обсуждаемого ниже . Они очень разнообразны . По оценкам, существует 200 000–800 000 видов, из которых описано около 50 000 видов. [149] В зависимости от вида их размеры колеблются от нескольких микрометров (мкм) до нескольких сотен микрометров. Они специально адаптированы к среде, в которой преобладают силы вязкости.
Одноклеточные организмы обычно микроскопические. Есть исключения. Бокал Русалки , род субтропических зеленых водорослей , является одноклеточным, но удивительно большим и сложным по форме с одним большим ядром, что делает его модельным организмом для изучения клеточной биологии . [150] Другая одноклеточная водоросль, Caulerpa Taxifolia , имеет вид сосудистого растения с «листьями», аккуратно расположенными на стеблях, как у папоротника. Селективное разведение в аквариумах с целью получения более выносливых штаммов привело к случайному попаданию в Средиземное море, где они превратились в инвазивный вид, известный в просторечии как водоросли-убийцы . [151]
Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
Макроводоросли — это более крупные, многоклеточные и более заметные виды водорослей, обычно называемые морскими водорослями . Морские водоросли обычно растут на мелководье в прибрежных водах, где они прикреплены к морскому дну с помощью крепления . Как и микроводоросли, макроводоросли (водоросли) можно рассматривать как морские протисты, поскольку они не являются настоящими растениями. Но они не являются микроорганизмами, поэтому не входят в задачу данной статьи.
Планктон — это дрейфующие или плавающие организмы, которые не могут эффективно плавать против течения, и включают организмы из большинства областей жизни: бактерии , археи , водоросли , простейшие и животные . Красный, оранжевый, желтый и зеленый представляют районы, где наблюдается изобилие цветения водорослей. Синие области представляют собой зоны с низким содержанием питательных веществ, где фитопланктон существует в более низких концентрациях.
Планктон (от греч. странники ) — разнообразная группа организмов, обитающих в толще воды крупных водоемов, но не умеющих плыть против течения. В результате они блуждают или дрейфуют по течению. [153] Планктон определяется своей экологической нишей , а не какой-либо филогенетической или таксономической классификацией. Они являются важнейшим источником пищи для многих морских животных, от кормовой рыбы до китов . Планктон можно разделить на растительный компонент и животный компонент.
Фитопланктон, такой как эта колония Chaetocerossocialis , естественным образом излучает красный флуоресцентный свет, который рассеивает избыточную солнечную энергию, которую они не могут потребить посредством фотосинтеза. Это свечение может быть обнаружено спутниками как индикатор того, насколько эффективно фотосинтезирует фитопланктон океана. [154] [155]
Фитопланктон — это растительные компоненты планктонного сообщества («фито» происходит от греческого слова « растение »). Они автотрофны (самопитаются), то есть производят собственную пищу и им не нужно потреблять другие организмы.
Фитопланктон выполняет три важнейшие функции: он генерирует почти половину мирового атмосферного кислорода, регулирует уровень углекислого газа в океане и атмосфере и составляет основу морской пищевой сети . При подходящих условиях в поверхностных водах может наблюдаться цветение водорослей фитопланктона. Фитопланктон — это r-стратег , который быстро растет и может удваивать свою популяцию каждый день. Цветки могут стать токсичными и лишить воду кислорода. Однако численность фитопланктона обычно контролируется за счет того, что фитопланктон истощает доступные питательные вещества и выедает зоопланктон. [156]
Рисунки Геккеля 1904 г. (нажмите, чтобы узнать подробности)
Диатомеи образуют (спорный) тип, содержащий около 100 000 признанных видов, в основном одноклеточных водорослей. Диатомовые водоросли производят около 20 процентов кислорода, производимого на планете каждый год. [93] поглощают более 6,7 миллиардов тонн кремния из вод, в которых они обитают, ежегодно [159] и составляют почти половину органического материала, обнаруженного в океанах.
Диатомеи – один из наиболее распространенных видов фитопланктона.
Их защитные оболочки (фрустлы) изготовлены из кремния.
Диатомовые водоросли имеют кремнеземную оболочку ( панцирь ) с радиальной (центрической) или двусторонней (пеннатной) симметрией.
Диатомовые водоросли заключены в защитные кремнеземные (стеклянные) оболочки, называемые панцирями . Каждая панцирь состоит из двух взаимосвязанных частей, покрытых крошечными отверстиями, через которые диатомовые водоросли обмениваются питательными веществами и отходами. [156] Панцири мертвых диатомей дрейфуют на дно океана, где за миллионы лет они могут накапливаться на глубине до полумили . [160]
Кварцованная панцирь пеннатной диатомеи с двумя перекрывающимися половинками.
Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [161]
Ископаемая диатомовая водоросль
Существует более 100 000 видов диатомовых водорослей , на долю которых приходится 50% первичной продукции океана.
Кокколитофоры — это мелкие одноклеточные фотосинтезирующие протисты с двумя жгутиками для передвижения. Большинство из них защищено панцирем, покрытым декоративными круглыми пластинками или чешуйками, называемыми кокколитами . Кокколиты состоят из карбоната кальция. Кальцитовые раковины важны для морского углеродного цикла. [163] Термин кокколитофор происходит от греческого слова « камень, несущий семя» , что связано с их небольшим размером и камнями кокколита, которые они несут. При правильных условиях они цветут, как и другой фитопланктон, и могут сделать океан молочно-белым. [164]
Модель механизма генерации энергии у морских бактерий
(1) Когда солнечный свет падает на молекулу родопсина (2) он меняет свою конфигурацию, поэтому протон вылетает из клетки (3) химический потенциал заставляет протон возвращаться в клетку (4) таким образом генерируя энергию (5) в форме аденозинтрифосфата . [165]
Фототрофический метаболизм основан на использовании одного из трех пигментов, преобразующих энергию: хлорофилла , бактериохлорофилла и ретиналя . Ретиналь – это хромофор, содержащийся в родопсинах . О значении хлорофилла в преобразовании световой энергии пишут уже несколько десятилетий, однако фототрофия на основе пигментов сетчатки только начинает изучаться. [166]
В 2000 году группа микробиологов под руководством Эдварда Делонга сделала важнейшее открытие в понимании морских углеродных и энергетических циклов. Они обнаружили ген у нескольких видов бактерий [168] [169] отвечает за выработку белка родопсина , ранее не встречавшегося у бактерий. Эти белки, обнаруженные в клеточных мембранах, способны преобразовывать энергию света в биохимическую энергию за счет изменения конфигурации молекулы родопсина при попадании на нее солнечного света, вызывая перекачку протона изнутри наружу и последующий приток, генерирующий энергию. [170] Археоподобные родопсины впоследствии были обнаружены среди различных таксонов, простейших, а также у бактерий и архей, хотя они редки в сложных многоклеточных организмах . [171] [172] [173]
Исследования 2019 года показывают, что эти «похищающие солнце бактерии» более распространены, чем считалось ранее, и могут изменить влияние глобального потепления на океаны. «Результаты расходятся с традиционной интерпретацией морской экологии, изложенной в учебниках, согласно которой почти весь солнечный свет в океане улавливается хлорофиллом в водорослях. Вместо этого бактерии, оснащенные родопсином, действуют как гибридные автомобили, приводимые в движение органическими веществами, когда они доступны, большинство бактерий — и под действием солнечного света, когда питательных веществ не хватает». [174] [166]
Существует астробиологическая гипотеза, называемая гипотезой пурпурной Земли , которая предполагает, что первоначальные формы жизни на Земле были основаны на сетчатке глаза, а не на хлорофилле, из-за чего Земля выглядела фиолетовой, а не зеленой. [175] [176]
В 1930-е годы Альфред Редфилд обнаружил сходство между составом элементов фитопланктона и основными растворенными питательными веществами в глубинах океана. [177] Редфилд предположил, что соотношение углерода, азота и фосфора (106:16:1) в океане контролируется потребностями фитопланктона, поскольку фитопланктон впоследствии выделяет азот и фосфор по мере реминерализации. Это соотношение стало известно как соотношение Редфилда и используется в качестве фундаментального принципа при описании стехиометрии морской воды и эволюции фитопланктона. [178]
Однако коэффициент Редфилда не является универсальной величиной и может меняться в зависимости от таких факторов, как географическая широта. [179] В зависимости от распределения ресурсов фитопланктон можно разделить на три различные стратегии роста: выживающую, цветущую и универсальную. Фитопланктон-выживальщик имеет высокое соотношение N:P (>30) и содержит множество механизмов по приобретению ресурсов для поддержания роста в условиях ограниченных ресурсов. Фитопланктон Bloomer имеет низкое соотношение N:P (<10), содержит высокую долю механизмов роста и адаптирован к экспоненциальному росту. Фитопланктон общего назначения имеет соотношение N:P, аналогичное соотношению Редфилда, и содержит относительно равные механизмы приобретения ресурсов и роста. [178]
Зоопланктон — животный компонент планктонного сообщества («зоопарк» происходит от греческого « животное »). Они гетеротрофны (питаются другими), что означает, что они не могут производить собственную пищу и вместо этого должны потреблять в пищу другие растения или животных. В частности, это означает, что они едят фитопланктон.
Рисунки Геккеля 1904 г. (нажмите, чтобы узнать подробности)
Радиолярии — это одноклеточные хищные протисты , заключенные в сложные шаровидные раковины, обычно сделанные из кремнезема и пронизанные отверстиями. Их название происходит от латинского слова «радиус». Они ловят добычу, вытягивая части своего тела через отверстия. Как и в случае с кремнеземными панцирями диатомей, раковины радиолярий могут опускаться на дно океана, когда радиолярии умирают, и сохраняются как часть океанских отложений . Эти останки в виде микрокаменелостей предоставляют ценную информацию о прошлых условиях океана. [185]
Как и диатомеи, радиолярии бывают разных форм.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Рисунки Геккеля 1904 г. (нажмите, чтобы узнать подробности)
Как и радиолярии, фораминиферы ( сокращенно форамы ) представляют собой одноклеточные хищные протисты, также защищенные раковинами с отверстиями. Их название происходит от латинского слова «носители дыр». Их раковины, часто называемые панцирями , имеют камеры (по мере роста у них появляются дополнительные камеры). Раковины обычно состоят из кальцита, но иногда из склеенных частиц осадка или хитона и (редко) кремнезема. Большинство форамов донные, но около 40 видов — планктонные. [187] Они широко исследуются с использованием хорошо известных летописей окаменелостей, которые позволяют ученым делать много выводов о окружающей среде и климате прошлого. [185]
Фораминиферы
...может иметь более одного ядра
...и защитные шипы
Фораминиферы — важные одноклеточные протисты зоопланктона , имеющие тесты на кальций.
Миксотроф — это организм, который может использовать смесь различных источников энергии и углерода вместо одного трофического режима в континууме от полной автотрофии на одном конце до гетеротрофии на другом. Подсчитано, что миксотрофы составляют более половины всего микроскопического планктона. [192] Существует два типа эукариотических миксотрофов: те, которые имеют собственные хлоропласты , и те, которые имеют эндосимбионты , а также другие, которые приобретают их посредством клептопластики или путем порабощения всей фототрофной клетки. [193]
Недавние исследования морского микрозоопланктона показали, что 30–45% численности инфузорий являются миксотрофными, а до 65% биомассы амебоидов, форамов и радиолярий - миксотрофными. [87]
Phaeocystis — важный род водорослей, встречающийся в составе морского фитопланктона по всему миру. Он имеет полиморфный жизненный цикл: от свободноживущих клеток до крупных колоний. [196] Он обладает способностью образовывать плавающие колонии, где сотни клеток встроены в гелевую матрицу, которая может значительно увеличиваться в размерах во время цветения . [197] В результате Phaeocystis вносит важный вклад в содержание морского углерода. [198] и циклы серы . [199] Виды Phaeocystis являются эндосимбионтами акантарианских радиолярий. [200] [201]
Миксотрофный планктон, сочетающий фототрофию и гетеротрофию – таблица на основе Stoecker et al., 2017. [202]
назвал их конститутивными миксотрофами . Mitra et al., 2016 [203] Питающийся фитопланктон: фотосинтезирующие протисты с наследственными пластидами и способностью поедать добычу.
назвал их неконститутивными миксотрофами . Mitra et al., 2016 [203] Зоопланктон, обладающий фотосинтезом: микрозоопланктон или многоклеточный зоопланктон, приобретающий фототрофию за счет удержания хлоропластов. а или поддержание эндосимбионтов водорослей.
Генералисты
Протисты, сохраняющие хлоропласты и редко другие органеллы многих таксонов водорослей.
Большинство инфузорий - олиготрихов, сохраняющих пластиды. а
Специалисты
1. Протисты, сохраняющие хлоропласты, а иногда и другие органеллы одного вида водорослей или очень близких видов водорослей.
а Удержание хлоропластов (или пластид) = секвестрация = порабощение. Некоторые виды, сохраняющие пластиды, также сохраняют другие органеллы и цитоплазму добычи.
Динофлагелляты являются частью группы водорослей и образуют тип одноклеточных жгутиконосцев, насчитывающий около 2000 морских видов. [204] Название происходит от греческого «динос», что означает вращение , и латинского «жгутик», означающего кнут или плеть . Это относится к двум хлыстообразным приспособлениям (жгутикам), используемым для движения вперед. Большинство динофлагеллят защищены красно-коричневой целлюлозной броней. Как и другой фитопланктон, динофлагелляты являются r-стратегами , которые при правильных условиях могут цвести и вызывать красные приливы . Раскопки могут быть самой базальной линией жгутиковых. [102]
По трофической ориентации динофлагелляты нельзя разделить на единые категории. Известно, что некоторые динофлагелляты фотосинтезируют , но большая часть из них на самом деле миксотрофны , сочетая фотосинтез с поеданием добычи ( фаготрофия ). [205] Некоторые виды являются эндосимбионтами морских животных и других протистов и играют важную роль в биологии коралловых рифов . Другие предшествуют другим простейшим, а некоторые формы являются паразитическими. Многие динофлагелляты миксотрофны и их также можно отнести к фитопланктону.
Токсичная динофлагеллята Dinophys acuta приобретает хлоропласты от своей добычи. «Он не может поймать криптофиты сам по себе и вместо этого полагается на заглатывание инфузорий, таких как красный Myrionecta Rubra , которые изолируют свои хлоропласты от определенной клады криптофитов (Geminigera/Plagioselmis/Teleaulax)». [202]
Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря.
Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.
Насселлярии радиолярии могут находиться в симбиозе с динофлагеллятами.
Некоторые динофлагелляты биолюминесцентны . Ночью океанская вода может светиться изнутри и сверкать синим светом из-за этих динофлагеллят. [210] [211] Биолюминесцентные динофлагелляты обладают сцинтиллонами , отдельными цитоплазматическими тельцами, содержащими динофлагеллятлюциферазу , основной фермент, участвующий в люминесценции. Люминесценция, иногда называемая фосфоресценцией моря , возникает в виде коротких (0,1 секунды) синих вспышек или искр при стимуляции отдельных сцинтиллонов, обычно в результате механических воздействий, например, от лодки, пловца или прибоя. [212]
Отложения на дне океана имеют два основных происхождения: терригенное и биогенное.
Терригенные отложения составляют около 45% от общего количества морских отложений и возникают в результате эрозии горных пород на суше, переноса реками и земельным стоком, переносимой ветром пыли, вулканов или измельчения ледниками.
Биогенные отложения составляют остальные 55% от общего количества осадков и происходят из скелетных останков морских протистов (одноклеточных планктонных и бентосных микроорганизмов). Также могут присутствовать гораздо меньшие количества осажденных минералов и метеорной пыли. В контексте морских отложений «ил» относится не к консистенции осадка, а к его биологическому происхождению. Термин «ил» первоначально был использован Джоном Мюрреем , «отцом современной океанографии», который предложил термин «радиолярийный ил» для обозначения кремнеземных отложений раковин радиолярий, вынесенных на поверхность во время экспедиции «Челленджер» . [217] Биогенный ил представляет собой пелагический осадок, содержащий не менее 30 процентов скелетных останков морских организмов.
Кокколитофоры являются крупнейшим глобальным источником биогенного карбоната кальция и вносят значительный вклад в глобальный углеродный цикл. [221] Они являются основной составляющей меловых отложений, таких как белые скалы Дувра .
Распределение типов отложений на морском дне В каждой цветной области преобладает показанный тип материала, хотя, вероятно, будут присутствовать и другие материалы. Дополнительную информацию см. здесь.
Морской микробентос — это микроорганизмы, обитающие в донной зоне океана, то есть вблизи морского дна или на нем, а также внутри или на поверхностных отложениях морского дна. Слово бентос происходит от греческого языка и означает «глубина моря». Микробентос встречается повсюду на морском дне континентальных шельфов или вблизи него, а также в более глубоких водах, с большим разнообразием в донных отложениях или на них. На мелководье луга с морской травой , коралловые рифы и леса из водорослей представляют собой особенно богатую среду обитания. В фотических зонах как фотосинтезирующие организмы доминируют бентосные диатомеи. В приливных зонах изменение приливов сильно ограничивает возможности микробентоса.
Elphidium - широко распространенный многочисленный род донных форамов.
И фораминиферы, и диатомеи имеют планктонную и донную формы, то есть могут дрейфовать в толще воды или обитать на отложениях на дне океана. В любом случае, после смерти их раковины оказываются на морском дне. Эти оболочки широко используются в качестве индикаторов климата . Химический состав раковин является следствием химического состава океана в момент образования раковин. О температуре воды в прошлом также можно судить по соотношению стабильных изотопов кислорода в раковинах, поскольку более легкие изотопы легче испаряются в более теплой воде, оставляя более тяжелые изотопы в раковинах. Информацию о климате прошлого можно получить на основе обилия форов и диатомовых водорослей, поскольку они, как правило, более распространены в теплой воде. [223]
Внезапное вымирание , убившее динозавров 66 миллионов лет назад, также привело к вымиранию трех четвертей всех других видов животных и растений. Однако впоследствии глубоководные бентосные форумы процветали. В 2020 году сообщалось, что исследователи изучили химический состав тысяч образцов этих донных образований и использовали свои результаты для создания самых подробных климатических записей Земли за всю историю. [224] [225]
Некоторые эндолиты живут чрезвычайно долго. В 2013 году исследователи сообщили о наличии на дне океана эндолитов возрастом, возможно, миллионы лет, со временем образования 10 000 лет. [226] Они медленно метаболизируются и не находятся в спящем состоянии. Возраст некоторых Actinomycetota , обнаруженных в Сибири, оценивается в полмиллиона лет. [227] [228] [229]
(А) Микробные взаимодействия варьируются от взаимовыгодных до вредных для одного или нескольких партнеров. Синие двунаправленные стрелки подчеркивают, что отношения между классификациями могут меняться, часто под влиянием условий окружающей среды. (B) Симбиозы хозяина и микроба следует рассматривать в контексте микробных сообществ, где хозяин участвует во множественных и часто различных симбиотических отношениях. (C) Микробные сообщества находятся под влиянием множества микробно-микробных симбиозов, начиная от сотрудничества (например, синтрофии или кометаболизма) до конкуренции. Стрелки обозначают в целом благоприятные (синие) и вредные (красные) результаты для одного (одинарный наконечник) или обоих (двойной наконечник) участников. Обратите внимание, что, как и в случае симбиоза хозяин-микроб, эти отношения можно рассматривать как изменчивые и находящиеся под влиянием условий окружающей среды. [230]
Концепция холобионта была первоначально определена доктором Линн Маргулис в ее книге 1991 года «Симбиоз как источник эволюционных инноваций» как совокупность хозяина и многих других видов, живущих внутри него или вокруг него, которые вместе образуют дискретную экологическую единицу . [231] Компонентами холобионта являются отдельные виды или бионты , а совокупный геном всех бионтов — хологеном . [232]
Эта концепция впоследствии развивалась с момента появления этого первоначального определения, [233] при этом фокус перемещается на виды микробов, связанные с хозяином. Таким образом, холобионт включает в себя хозяина, вирома , микробиома и других членов, каждый из которых каким-то образом способствует функционированию целого. [234] [235] Голобионт обычно включает в себя эукариот- хозяина и все симбиотические вирусы , бактерии , грибы и т. д., живущие на нем или внутри него. [236]
Однако существуют разногласия по поводу того, можно ли рассматривать холобионтов как отдельные эволюционные единицы. [237]
Микробные сообщества морского льда (SIMCO) относятся к группам микроорганизмов, живущих внутри и на границе раздела морского льда на полюсах. Ледяная матрица, в которой они обитают, имеет сильные вертикальные градиенты солености, освещенности, температуры и питательных веществ. На химический состав морского льда больше всего влияет соленость рассола, которая влияет на pH и концентрацию растворенных питательных веществ и газов. Рассол , образующийся при таянии морского льда, создает в морском льду поры и каналы, в которых могут жить эти микробы. В результате этих градиентов и динамических условий более высокая численность микробов обнаруживается в нижнем слое льда, хотя некоторые из них обнаруживаются в среднем и верхнем слоях. [251]
Гидротермальные источники расположены там, где тектонические плиты расходятся и расширяются. Это позволяет воде из океана проникать в земную кору, где она нагревается магмой. Растущее давление и температура вынуждают воду выходить из этих отверстий, на выходе вода накапливает растворенные минералы и химические вещества из пород, с которыми она сталкивается. Вентиляционные отверстия можно охарактеризовать по температуре и химическому составу как диффузные отверстия , из которых выделяется чистая, относительно прохладная вода, обычно ниже 30 °C, как белые курильщики , которые выделяют воду молочного цвета при более высоких температурах, около 200–330 °C, и как черные курильщики , которые выделяют воду. затемняются накопившимися осадками сульфида при высоких температурах, около 300-400°С. [252]
Микробные сообщества гидротермальных жерл представляют собой микроскопические одноклеточные организмы, которые живут и размножаются в химически отличной области вокруг гидротермальных жерл. К ним относятся организмы в микробных матах , свободно плавающие клетки и бактерии, находящиеся в эндосимбиотических отношениях с животными. Поскольку на этих глубинах нет солнечного света, энергия обеспечивается хемосинтезом , при котором симбиотические бактерии и археи образуют нижнюю часть пищевой цепи и способны поддерживать различные организмы, такие как гигантские трубчатые черви и черви Помпеи . Эти организмы используют эти симбиотические отношения, чтобы использовать и получать химическую энергию, которая высвобождается в этих гидротермальных жерлах. [253] Хемолитоавтотрофные бактерии могут получать питательные вещества и энергию в результате геологической активности гидротермальных источников для фиксации углерода в органических формах. [254]
Вирусы также являются частью микробного сообщества гидротермальных источников, и их влияние на микробную экологию в этих экосистемах является развивающейся областью исследований. [255] Вирусы — самая распространенная форма жизни в океане, таящая в себе величайший резервуар генетического разнообразия. [256] Поскольку их инфекции часто приводят к летальному исходу, они представляют собой значительный источник смертности и, таким образом, оказывают широкое влияние на биологические океанографические процессы, эволюцию и биогеохимический цикл в океане. [257] Однако были обнаружены доказательства того, что вирусы, обнаруженные в жерловых средах обитания, приняли более мутуалистическую , чем паразитическую эволюционную стратегию, чтобы выжить в экстремальной и нестабильной среде, в которой они существуют. [258] Было обнаружено, что глубоководные гидротермальные источники содержат большое количество вирусов, что указывает на высокую вирусную продукцию. [259] Как и в других морских средах, глубоководные гидротермальные вирусы влияют на численность и разнообразие прокариот и, следовательно, влияют на микробный биогеохимический цикл, лизис своих хозяев для репликации. [260] Однако, в отличие от своей роли источника смертности и контроля над популяцией, предполагается, что вирусы также повышают выживаемость прокариот в экстремальных условиях, выступая в качестве резервуаров генетической информации. Таким образом, считается, что взаимодействие виросферы с микроорганизмами в условиях стресса окружающей среды способствует выживанию микроорганизмов за счет распространения генов хозяина посредством горизонтального переноса генов . [261]
Флуоресцентная микроскопия различного пикопланктона в океане, большая часть которого не может быть эффективно изучена, поскольку они сопротивляются попыткам лабораторного культивирования.
— Глубинная биосфера это та часть биосферы , которая находится ниже первых нескольких метров поверхности. Он простирается как минимум на 5 километров ниже поверхности материка и на 10,5 километров ниже поверхности моря, а температура может превышать 100 °C.
Над поверхностью живые организмы потребляют органические вещества и кислород. Ниже они недоступны, поэтому они используют «съедобные продукты» ( доноры электронов ), такие как водород, выделяемый из горных пород в результате различных химических процессов, метан, восстановленные соединения серы и аммоний. Они «дышат» акцепторами электронов , такими как нитраты и нитриты, оксиды марганца и железа, окисленные соединения серы и углекислый газ.
На больших глубинах очень мало энергии, а метаболизм может быть в миллион раз медленнее, чем на поверхности. Клетки могут жить тысячи лет, прежде чем делиться, и их возраст не ограничен. На долю недр приходится около 90% биомассы бактерий и архей и 15% общей биомассы биосферы. Встречаются также эукариоты, в основном микроскопические, но включающие и многоклеточную жизнь. Вирусы также присутствуют и заражают микробы.
Подземная среда жизни
В 2018 году исследователи из Обсерватории Deep Carbon объявили, что формы жизни , в том числе 70% бактерий и архей на Земле, общая биомасса которых составляет 23 миллиарда тонн углерода до 4,8 км (3,0 мили) , живут на глубине , в том числе на глубине до 2,5 км ( 3,0 мили). 1,6 мили) ниже морского дна. [262] [263] [264] В 2019 году были обнаружены микробные организмы, живущие на глубине 7900 футов (2400 м) под поверхностью земли, дышащие серой и питающиеся такими камнями, как пирит, в качестве постоянного источника пищи. [265] [266] [267] Это открытие произошло в древнейшей из известных вод на Земле. [268]
Эти аэробные микроорганизмы, обнаруженные глубоко в бедных органикой отложениях, находились в квази-взвешенном состоянии около 100 миллионов лет.
Неопознанный сферический водорослевый микроорганизм, часть микробной темной материи.
На сегодняшний день биологам не удалось культивировать в лаборатории подавляющее большинство микроорганизмов. Это особенно относится к бактериям и археям и связано с недостатком знаний или способности обеспечить необходимые условия для роста. [271] [272] Термин «микробная темная материя» стал использоваться для описания микроорганизмов, о существовании которых ученые знали, но которые не смогли культивировать, и поэтому свойства которых остаются неуловимыми. [271] Микробная темная материя не связана с темной материей физики и космологии, но получила свое название из-за сложности ее эффективного изучения. Трудно оценить его относительную величину, но общепринятая общая оценка состоит в том, что менее одного процента видов микробов в данной экологической нише пригодны для культивирования. В последние годы предпринимаются усилия по расшифровке темной материи микробов посредством изучения их генома последовательности ДНК по образцам окружающей среды. [273] а затем, получив представление об их метаболизме из секвенированного генома, продвигая знания, необходимые для их выращивания.
Оценки количества видов микробов в трех сферах жизни
Бактерии — самая старая и наиболее биоразнообразная группа, за ней следуют археи и грибы (самые новые группы). В 1998 году, еще до того, как стало известно о масштабах микробной жизни, Роберт М. Мэй [274] По оценкам, на планете существовало 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [275] По оценкам, количество видов микроорганизмов может достигать 1 триллиона. [276]
Микробное разнообразие
Сравнительное представление известного и оцененного (маленькая рамка) и еще неизвестного (большая рамка) микробного разнообразия, которое применимо как к морским, так и к наземным микроорганизмам. Текстовые поля относятся к факторам, которые отрицательно влияют на знания о микробном разнообразии, существующем на планете. [276]
Стратегии отбора проб планктона по размерным классам и численности Синий фон указывает отфильтрованный объем, необходимый для получения достаточного количества микроорганизмов для анализа. Фактические объемы, из которых были отобраны образцы организмов, всегда регистрируются. [277]
Традиционно филогения микроорганизмов и их систематика устанавливались на основе исследований морфологии . Однако достижения в области молекулярной филогенетики позволили установить эволюционное родство видов путем анализа более глубоких характеристик, таких как их ДНК и последовательности белков , например рибосомальной ДНК . [278] Отсутствие легкодоступных морфологических особенностей, таких как те, которые присутствуют у животных и растений , особенно затрудняло ранние попытки классификации бактерий и архей. Это привело к ошибочной, искаженной и запутанной классификации, примером которой, как отметил Карл Вёзе , является Pseudomonas , этимология которой иронично соответствовала ее таксономии, а именно «ложная единица». [279] Многие бактериальные таксоны были реклассифицированы или переопределены с использованием молекулярной филогенетики.
Marinomonas arctica — бактерия, которая растет внутри арктического морского льда при минусовых температурах.
Было бы сложно последовательно разделить эти два микроба, используя только изображения. Однако, если их штрих-коды выровнены друг по другу, а их основания окрашены в цвет, чтобы их было более четко видно, становится легко увидеть, какие основания различаются у этих двух микробов. Таким образом можно различить миллионы различных видов микробов. [280]
Три различных способа обработки пробы: использование необработанных проб, фиксация или консервация и фильтрация. Для микроскопии и проточной цитометрии необработанные образцы либо измеряются сразу, либо их необходимо фиксировать для последующих измерений. Поскольку молекулярные методы, анализ пигментов и обнаружение молекулярных индикаторов обычно требуют концентрированных клеток, остатки фильтров служат для измерений фитопланктона. Молекулярная характеристика и количественная оценка следовых молекул выполняется с использованием хроматографии , масс-спектрометрии и ядерного магнитного резонанса . спектроскопии [281]
Новые технологии секвенирования и накопление данных о последовательностях привели к смене парадигмы, подчеркнув как повсеместное распространение микробных сообществ в ассоциациях внутри высших организмов, так и критическую роль микробов в здоровье экосистемы. [283] Эти новые возможности произвели революцию в микробной экологии , поскольку высокопроизводительный анализ геномов и метагеномов обеспечивает эффективные методы раскрытия функционального потенциала отдельных микроорганизмов, а также целых сообществ в их естественной среде обитания. [284] [285] [286]
Технологии секвенирования ДНК, используемые в морской метагеномике
Процесс открытия включает в себя отбор проб морской среды, секвенирование ДНК и генерацию контигов . Открываются ранее неизвестные гены, пути и даже целые геномы. Эти технологии редактирования генома используются для извлечения и модификации ценных микроорганизмов для производства, особенно в морской метагеномике. Организмы могут быть культивируемыми и некультивируемыми. Метагеномика предоставляет особенно ценную информацию о некультивируемых образцах. [287]
Омики — термин, неофициально используемый для обозначения разделов биологии , названия которых заканчиваются суффиксом —омика , таких как геномика , протеомика , метаболомика и гликомика . Морская омика недавно стала отдельной областью исследований. [288] Омика направлена на коллективную характеристику и количественную оценку пулов биологических молекул, которые отражаются на структуре, функциях и динамике организма или организмов. Например, функциональная геномика направлена на выявление функций как можно большего числа генов данного организма. Он сочетает в себе разные -омные методы, такие как транскриптомика и протеомика с насыщенными коллекциями мутантов . [289] [290]
Многие омы, выходящие за пределы исходного генома, стали полезными и в последние годы были широко приняты учеными-исследователями. Суффикс -omics может служить простым сокращением для инкапсуляции поля; например, исследование интерактомики достаточно известно как относящееся к крупномасштабному анализу взаимодействий ген-ген, белок-белок или белок-лиганд, в то время как протеомика стала общепринятым термином для изучения белков в больших масштабах.
Биогеохимическое моделирование на основе данных метаомики [293]
Схематическая концептуальная основа морского биогеохимического моделирования на основе данных окружающей среды, изображений и метаомики. [294] Схематизирован полуавтоматический вычислительный конвейер для объединения биомаркеров с биогеохимическими данными. [295] которые могут быть включены в классические биогеохимические модели [296] для создания следующего поколения моделей метаомики, основанных на биогеохимических признаках, путем рассмотрения их соответствующих характеристик. Такие новые подходы, основанные на метаомике, направлены на улучшение мониторинга и прогнозирования океанических процессов, учитывая при этом сложность планктонной системы. [297] [298]
Использование данных омики с морским фитопланктоном [299]
В качестве примера того, как данные омики могут использоваться с морским фитопланктоном для информирования науки о системе Земли , были использованы метатранскриптомные последовательности из природных сообществ фитопланктона, чтобы помочь идентифицировать физиологические особенности (клеточную концентрацию рибосом и их рРНК ), лежащие в основе адаптации к условиям окружающей среды (температуре). Механистическая клеточная модель фитопланктона была использована для проверки значимости выявленного физиологического признака для клеточной стехиометрии . Экологический отбор в модели глобальной морской экосистемы, основанной на признаках, затем связывал возникающие стратегии роста и распределения клеток с крупномасштабными закономерностями света, питательных веществ и температуры в поверхностной морской среде. Глобальные прогнозы распределения клеточных ресурсов и стехиометрии (соотношение N:P) соответствовали закономерностям в метатранскриптомных данных. [300] и широтные закономерности в соотношениях элементов морского планктона и органического вещества. [301] Трехмерное изображение рибосомы показывает рРНК темно-синим и темно-красным цветом. Более светлые цвета представляют рибосомальные белки . Полосы выше показывают температурно-зависимое содержание эукариотического рибосомального белка S14. [300]
Микроорганизмы и изменение климата в морских и наземных биомах [6]
В морской среде первичная микробная продукция вносит существенный вклад в CO 2 секвестрацию . Морские микроорганизмы также перерабатывают питательные вещества для использования в морской пищевой сети и при этом выделяют CO 2 в атмосферу. Микробная биомасса и другие органические вещества (остатки растений и животных) превращаются в ископаемое топливо в течение миллионов лет. Напротив, сжигание ископаемого топлива приводит к высвобождению парниковых газов за небольшую долю этого времени. В результате углеродный цикл разбалансирован, и уровень CO 2 в атмосфере будет продолжать расти до тех пор, пока будет продолжаться сжигание ископаемого топлива. [6]
Микроорганизмы играют ключевую роль в круговороте углерода и питательных веществ, здоровье животных (включая человека) и растений, сельском хозяйстве и глобальной пищевой сети. Микроорганизмы живут во всех средах на Земле, населенных макроскопическими организмами, и являются единственными формами жизни в других средах, таких как глубокие недра и «экстремальные» среды. Микроорганизмы восходят к зарождению жизни на Земле, по крайней мере, 3,8 миллиарда лет назад, и они, вероятно, будут существовать намного дольше, чем любые будущие события вымирания... Если мы не осознаем важность микробных процессов, мы фундаментально ограничиваем наше понимание биосферы Земли и ее реакций. к изменению климата и тем самым поставить под угрозу усилия по созданию экологически устойчивого будущего. [6]
Морские микроорганизмы, известные как цианобактерии, впервые появились в океанах в докембрийскую эпоху примерно 2 миллиарда лет назад. На протяжении тысячелетий фотосинтез морских микроорганизмов, вырабатываемый кислородом, помогал формировать химическую среду в эволюции растений, животных и многих других форм жизни. Морские микроорганизмы впервые наблюдал в 1675 году голландский мастер линз Антони ван Левенгук .
^ Jump up to: а б Рыбицкий Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–6. ISSN 0038-2353 .
^ Болуис, Х. и Кретуу, М.С. (2016) «Что такого особенного в морских микроорганизмах?». В: Ж. Дж. Сталь и М. С. Кретойу (ред.) Морской микробиом , стр. 3–20, Springer. ISBN 9783319330006
^ Jump up to: а б Рыбицкий Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–186.
^ Мэхи, WJ; Ван Регенмортель, MHV, ред. (2009). Настольная энциклопедия общей вирусологии . Оксфорд: Академическая пресса. п. 28. ISBN 978-0-12-375146-1 .
^ «Кандидат Пелагибактер Убик» . Геномы бактерий . Европейский институт биоинформатики. 2011. Архивировано из оригинала 1 декабря 2008 года . Проверено 8 января 2012 г.
^ Ямагучи, Масаси; и др. (1 декабря 2012 г.). «Прокариот или эукариот? Уникальный микроорганизм из морских глубин». Журнал электронной микроскопии . 61 (6): 423–431. doi : 10.1093/jmicro/dfs062 . ПМИД 23024290 .
^ Добрый день, Эй Джей; Аранда да Силва, А.; Павловский, Дж. (1 декабря 2011 г.). «Ксенофиофоры (Rhizaria, Foraminifera) из каньона Назаре (окраина Португалии, северо-восток Атлантики)». Глубоководные исследования. Часть II: Актуальные исследования в океанографии . Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Бибкод : 2011DSRII..58.2401G . дои : 10.1016/j.dsr2.2011.04.005 .
^ Синглтон, Пол (2006). Словарь по микробиологии и молекулярной биологии, 3-е издание, переработанное . Чичестер, Великобритания: John Wiley & Sons. п. 32. ISBN 978-0-470-03545-0 .
^ Гутьеррес, Марсело Х; Пантоха, Сильвио; Хиноны, Ренато а и Гонсалес, Родриго Р. Первое упоминание о палистых грибах в прибрежной апвеллинговой экосистеме у центрального Чили. Гаяна (Concepc.) [онлайн]. 2010, т.74, №1, стр. 66-73. ISSN 0717-6538 .
^ Jump up to: а б Шридхар, КР (2009). «10. Водные грибы – они планктонные?». Планктонная динамика вод Индии . Джайпур, Индия: Публикации Пратикши. стр. 133–148.
^ Jump up to: а б Кэмпбелл, Нил А.; Рис, Джейн Б.; Урри, Лиза Андреа; Каин, Майкл Л.; Вассерман, Стивен Александер; Минорский, Петр В.; Джексон, Роберт Брэдли (2008). Биология (8-е изд.). Сан-Франциско: Пирсон – Бенджамин Каммингс. ISBN 978-0-321-54325-7 .
^ Старкс, Сенн (31 октября 2012 г.). «Место под солнцем» . Фландрия сегодня . Архивировано из оригинала 4 марта 2016 года . Проверено 8 декабря 2012 г. По мнению исследователей из Гиля, водоросли — это урожай будущего.
^ Мандоли Д.Ф. (1998). «Разработка плана тела и фазового перехода во время развития вертлужной впадины: как строится сложная архитектура гигантской одноклеточной клетки?». Ежегодный обзор физиологии растений и молекулярной биологии растений . 49 : 173–198. дои : 10.1146/annurev.arplant.49.1.173 . ПМИД 15012232 . S2CID 6241264 .
^ Рост, Б. и Рибеселл, У. (2004) «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». В: Кокколитофоры: от молекулярных процессов к глобальному воздействию , страницы 99–125, Springer. ISBN 9783662062784 .
^ Васильев, Мэгги (2006) «Кокколитофор» , Те Ара - Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
^ Спаркс, Уильям Б.; ДасСарма, С.; Рид, Индиана (декабрь 2006 г.). «Эволюционная конкуренция между примитивными фотосинтетическими системами: существование ранней фиолетовой Земли?». Тезисы докладов о заседании Американского астрономического общества . 38 : 901. Бибкод : 2006AAS...209.0605S .
^ Редфилд, Альфред К. (1934). «О пропорциях органических производных в морской воде и их связи с составом планктона». В Джонстоне, Джеймс; Дэниел, Ричард Джеллико (ред.). Мемориальный том Джеймса Джонстона . Ливерпуль: Университетское издательство Ливерпуля. стр. 176–92. OCLC 13993674 .
^ Шеманн, Вероника; Беквор, Сильви; Стефельс, Жаклин; Руссо, Вероника; Ланселот, Кристиана (1 января 2005 г.). «Цветение Phaeocystis в мировом океане и механизмы его контроля: обзор». Журнал морских исследований . Ресурсы железа и океанические питательные вещества – развитие глобального экологического моделирования. 53 (1–2): 43–66. Бибкод : 2005JSR....53...43S . CiteSeerX 10.1.1.319.9563 . дои : 10.1016/j.seares.2004.01.008 .
^ Баски, Э.Дж. (1995). «Рост и биолюминесценция Noctiluca scintillans на различных водорослевых диетах». Журнал исследований планктона . 17 (1): 29–40. дои : 10.1093/планкт/17.1.29 .
^ Грета Р. Хасл; Эрик Э. Сиверцен; Карен А. Штайдингер; Карл Танген (25 января 1996 г.). «Морские диатомеи» . В Кармело Р. Томасе (ред.). Определение морских диатомей и динофлагеллят . Академическая пресса. стр. 5–385. ISBN 978-0-08-053441-1 . Проверено 13 ноября 2013 г.
^ Мохеймани, Северная Каролина; Уэбб, JP; Боровицка, Массачусетс (2012), «Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор. Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор», Algal Research , 1 (2): 120–133, doi : 10.1016/ j.algal.2012.06.002
^ Тайс, КР, Дейли, Н.М., Классен, Дж. Л., Брукер, РМ, Бейнс, Дж. Ф., Босх, Т. К., Крайан, Дж. Ф., Гилберт, С. Ф., Гуднайт, Си Джей, Ллойд, Э. А. и Сапп, Дж. (2016) «Получение концепция гологенома справа: экоэволюционная основа хозяев и их микробиомов». Мсистемс , 1 (2). дои : 10.1128/mSystems.00028-16 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Булгарелли Д., Шлеппи К., Спепен С., Ван Темаат ЭВЛ и Шульце-Леферт П. (2013) «Структура и функции бактериальной микробиоты растений». Ежегодный обзор биологии растений , 64 : 807–838. doi : 10.1146/annurev-arplant-050312-120106 .
^ Санчес-Канисарес, К., Хоррин, Б., Пул, П.С. и Ткач, А. (2017) «Понимание холобионта: взаимозависимость растений и их микробиома». Текущее мнение по микробиологии , 38 : 188–196. дои : 10.1016/j.mib.2017.07.001 .
^ Дуглас, А.Э. и Веррен, Дж.Х. (2016) «Дыры в гологеноме: почему симбиозы хозяина и микроба не являются холобионтами». МБио , 7 (2). дои : 10.1128/mBio.02099-15 .
^ Ноултон, Н. и Ровер, Ф. (2003) «Многовидовой микробный мутуализм на коралловых рифах: хозяин как среда обитания». Американский натуралист , 162 (S4): S51-S62. дои : 10.1086/378684 .
^ Томпсон, Дж.Р., Ривера, Х.Э., Клосек, С.Дж. и Медина, М. (2015) «Микробы в коралловом холобионте: партнеры в эволюции, развитии и экологических взаимодействиях». Границы клеточной и инфекционной микробиологии , 4 : 176. два : 10.3389/fcimb.2014.00176 .
^ Кавальканти, Г.С., Шукла, П., Моррис, М., Рибейро, Б., Фоли, М., Доан, М.П., Томпсон, К.С., Эдвардс, М.С., Динсдейл, Э.А. и Томпсон, Флорида (2018) «Родолиты-холобионты в меняющемся океане: взаимодействие хозяина и микробов обеспечивает устойчивость коралловых водорослей в условиях закисления океана». BMC Genomics , 19 (1): 1–13. два : 10.1186/s12864-018-5064-4 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Робинсон, Кэрол и Нагаппа Рамайя. «Скорость микробного гетеротрофного метаболизма ограничивает микробный углеродный насос». Американская ассоциация содействия развитию науки, 2011 г.
^ Монрой, П., Эрнандес-Гарсия, Э., Росси, В. и Лопес, К. (2017) «Моделирование динамического опускания биогенных частиц в океаническом потоке». Нелинейные процессы в геофизике , 24 (2): 293–305. дои : 10.5194/npg-24-293-2017 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
^ Саймон М., Гроссарт Х., Швейцер Б. и Плауг Х. (2002) «Микробная экология органических агрегатов в водных экосистемах». Водная микробная экология , 28 : 175–211. дои : 10.3354/ame028175 .
^ Боэций, Антье; Анесио, Александр М.; Деминг, Джоди В.; Микуки, Джилл А.; Рапп, Жозефина З. (1 ноября 2015 г.). «Микробная экология криосферы: морской лед и ледниковая среда обитания». Обзоры природы Микробиология . 13 (11): 677–690. дои : 10.1038/nrmicro3522 . ПМИД 26344407 . S2CID 20798336 .
^ Кадар, Энико; Коста, Валентина; Сантос, Рикардо С.; Пауэлл, Джонатан Дж. (июль 2006 г.). «Тканевое распределение микроэссенциальных металлов в жерловых двустворчатых моллюсках Bathymodiolus azoricus и связанных с ними организмах (эндосимбионтных бактериях и паразитических полихетах) из геохимически различных жерл Срединно-Атлантического хребта». Журнал морских исследований . 56 (1): 45–52. Бибкод : 2006JSR....56...45K . doi : 10.1016/j.seares.2006.01.002 . ISSN 1385-1101 .
^ Ортманн, Алиса К.; Саттл, Кертис А. (август 2005 г.). «Высокое содержание вирусов в глубоководной гидротермальной системе указывает на вирусно-опосредованную смертность микробов». Глубоководные исследования. Часть I: Статьи океанографических исследований . 52 (8): 1515–1527. Бибкод : 2005DSRI...52.1515O . дои : 10.1016/j.dsr.2005.04.002 . ISSN 0967-0637 .
^ Лоллар, Гранат С.; Уорр, Оливер; Рассказываю, Джон; Осберн, Магдалена Р.; Шервуд Лоллар, Барбара (2019). « Следуй за водой: гидрогеохимические ограничения на микробные исследования на глубине 2,4 км под поверхностью в Обсерватории глубоких жидкостей и глубокой жизни Кидд-Крик». Геомикробиологический журнал . 36 (10): 859–872. Бибкод : 2019GmbJ...36..859L . дои : 10.1080/01490451.2019.1641770 . S2CID 199636268 .
^ Лю, Л., Ли, Ю., Ли, С., Ху, Н., Хэ, Ю., Понг, Р., Лин, Д., Лу, Л. и Лоу, М. (2012) «Сравнение систем секвенирования нового поколения». BioMed Research International , 2012 : 251364. дои : 10.1155/2012/251364 .
^ Берг, Габриэле ; Дарья Рыбакова, Дорин Фишер, Томислав Чернава, Мари-Кристин Шампомье Верже, Тревор Чарльз, Сяоюлонг Чен, Лука Коколин, Келли Эверсол, Джема Эрреро Коррал, Мария Казу, Линда Кинкель, Лене Ланге, Нельсон Лима, Александр Лой, Джеймс А. Маклин , Эммануэль Маген, Тим Моклин, Райан МакКлюр, Биргит Миттер, Мэттью Райан, Инга Саранд, Хауке Смидт, Беттина Шелкле, Хьюго Рум, Дж. Сегал Киран, Джозеф Селвин, Рафаэль Соарес Корреа де Соуза, Лео ван Овербек, Браджеш К. Сингх , Михаэль Вагнер, Аарон Уолш, Анджела Сессич и Майкл Шлотер (2020) «Возврат к определению микробиома: старые концепции и новые проблемы». Микробиом , 8 (103): 1–22. два : 10.1186/s40168-020-00875-0 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Чжан В., Ли Ф. и Не Л. (2010) «Интеграция множественного анализа омики для микробной биологии: применение и методологии». Микробиология , 156 (2): 287–301. два : 10.1099/mic.0.034793-0 .
^ Капотонди, А., Джакокс, М., Боулер, К., Кавано, М., Леходи, П., Барри, Д., Броди, С., Чаффрон, С., Ченг, В., Диас, Д.Ф., Эвейяр, Д. и еще 20 человек (2019 г.) «Потребности в наблюдениях, поддерживающие моделирование и прогнозирование морских экосистем: от глобального океана до региональных и прибрежных систем». Границы морской науки , 6 : 623. два : 10.3389/fmars.2019.00623 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Сунагава С., Коэльо Л.П., Чаффрон С., Култима Дж.Р., Лабади К., Салазар Г. и др. (2015). «Океанический планктон. Структура и функции глобального микробиома океана». Наука , 348 : 1261359. дои : 10.1126/science.1261359 .
^ Гуиди Л., Шаффрон С., Биттнер Л., Эвейяр Д., Лархлими А., Ру С. и др. (2016). «Планктонные сети способствуют экспорту углерода в олиготрофный океан». Природа , 532 : 465–470. дои : 10.1038/nature16942 .
^ Гайеннон, А., Баклоути, М., Диас, Ф., Палмьери, Дж., Бёвье, Дж., Лебопен-Броссье, К. и др. (2015). «Новый взгляд на экспорт органического углерода в Средиземном море на основе 3D-моделирования». Биогеонауки , 11 : 425–442. дои : 10.5194/bg-12-7025-2015 .
^ Лука, С., Парфри, Л.В., и Добели, М. (2016). «Функция разделения и таксономия в микробиоме глобального океана». Наука , 353 : 1272–1277. doi : 10.1126/science.aaf4507 .
^ Коулз, В.Дж., Стукель, М.Р., Брукс, М.Т., Берд, А., Крамп, BC, Моран, Массачусетс и др. (2017). «Биогеохимия океана, смоделированная с помощью новой геномики, основанной на признаках». Наука , 358 : 1149–1154. дои : 10.1126/science.aan5712 .
^ Мок, Т., Дэйнс, С.Дж., Гейдер, Р., Коллинз, С., Методиев, М., Миллар, А.Дж., Моултон, В. и Лентон, Т.М. (2016) «Преодоление разрыва между омикой и наукой о системе Земли». чтобы лучше понять, как изменение окружающей среды влияет на морские микробы». Биология глобальных изменений , 22 (1): 61–75. дои : 10.1111/gcb.12983 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Jump up to: а б Тоузленд А., Дэйнс С.Дж. и Кларк Дж.Р. и др. (2013) «Влияние температуры на распределение ресурсов и метаболизм морского фитопланктона». Изменение климата природы , 3 : 979–984. дои : 10.1038/nclimate1989 .
^ Мартини AC, Фам CTA, Примо Ф.В., Вругт Дж.А., Мур Дж.К., Левин С.А. и Ломас М.В. (2013) «Сильные широтные закономерности в соотношениях элементов морского планктона и органического вещества». Nature Geoscience , 6 : 279–283. два : 10.1038/ngeo1757 .
Arc.Ask3.Ru Номер скриншота №: 2e4642a82f16af222bf9bf002e6d70fc__1717309860 URL1:https://arc.ask3.ru/arc/aa/2e/fc/2e4642a82f16af222bf9bf002e6d70fc.html Заголовок, (Title) документа по адресу, URL1: Marine microorganisms - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)
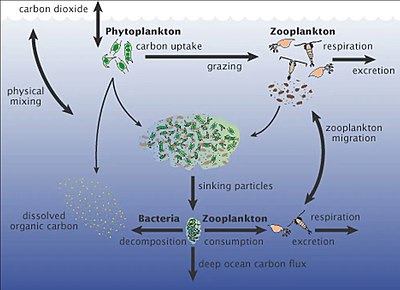



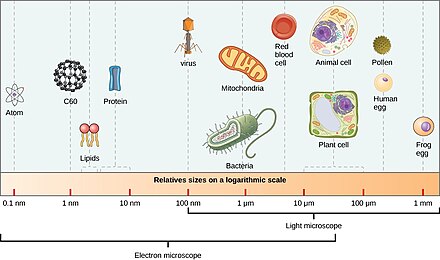




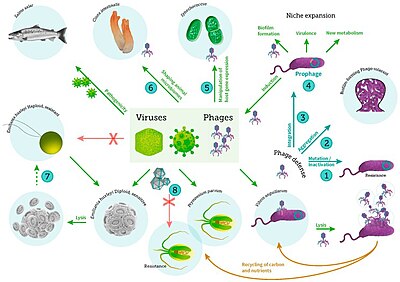
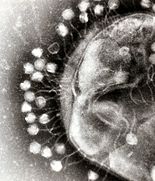
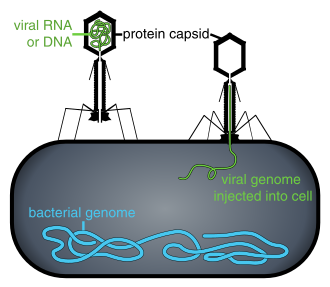
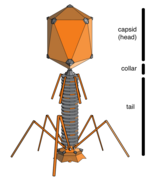


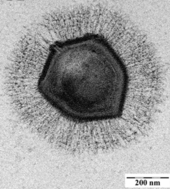



 Морская Thiomargarita namibiensis , крупнейшая известная бактерия.
Морская Thiomargarita namibiensis , крупнейшая известная бактерия.
 Хлоропласты эндосимбиотическом глаукофитов происхождении имеют слой пептидогликана что свидетельствует об их цианобактерий от , . [72]
Хлоропласты эндосимбиотическом глаукофитов происхождении имеют слой пептидогликана что свидетельствует об их цианобактерий от , . [72] Бактерия Marinomonas arctica растет внутри арктического морского льда при минусовых температурах.
Бактерия Marinomonas arctica растет внутри арктического морского льда при минусовых температурах.




 Эоциты могут быть самыми многочисленными среди морских архей.
Эоциты могут быть самыми многочисленными среди морских архей. Галобактерии , обнаруженные в воде, почти насыщенной солью, теперь признаны архей.
Галобактерии , обнаруженные в воде, почти насыщенной солью, теперь признаны архей. Плоские квадратные клетки архей Haloquadratum walsbyi.
Плоские квадратные клетки архей Haloquadratum walsbyi. Methanosarcina barkeri — морская архея, производящая метан.
Methanosarcina barkeri — морская архея, производящая метан. Термофилы , такие как Pyrolobus fumarii , выживают при температуре выше 100 °C.
Термофилы , такие как Pyrolobus fumarii , выживают при температуре выше 100 °C.









![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[93]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [93]
Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [93]![У диатомовых водорослей есть стеклянные клеточные стенки, состоящие из кремнезема и называемые панцирями.[94]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg) У диатомовых водорослей стеклянные клеточные стенки состоят из кремнезема и называются панцирями . [94]
У диатомовых водорослей стеклянные клеточные стенки состоят из кремнезема и называются панцирями . [94] Ископаемые панцири диатомей от 32 до 40 млн лет назад
Ископаемые панцири диатомей от 32 до 40 млн лет назад
 Одноклеточная водоросль Gephyrocapsa Oceanica.
Одноклеточная водоросль Gephyrocapsa Oceanica. Две динофлагелляты
Две динофлагелляты
 Зооксантеллы — фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы.
Зооксантеллы — фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы.
 Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа
Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[93]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
![У диатомовых водорослей есть стеклянные клеточные стенки, состоящие из кремнезема и называемые панцирями.[94]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)








 Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм.
Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм.![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр до 3,8 см.[96]](//upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр до 3,8 см. [96]
Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр до 3,8 см. [96] Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину.
Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину.![Ксенофиофор, еще один одноклеточный фораминифер, обитает в абиссальных зонах. Имеет гигантскую раковину диаметром до 20 см.[97]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg) Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . Имеет гигантскую раковину диаметром до 20 см. [97]
Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . Имеет гигантскую раковину диаметром до 20 см. [97] Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может вырасти до 50 м в высоту.
Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может вырасти до 50 м в высоту.
![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр до 3,8 см.[96]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png)

![Ксенофиофор, еще один одноклеточный фораминифер, обитает в абиссальных зонах. Имеет гигантскую раковину диаметром до 20 см.[97]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg)








 Бактериальный жгутик, вращаемый молекулярным мотором у его основания.
Бактериальный жгутик, вращаемый молекулярным мотором у его основания. лосося Сперматозоиды
лосося Сперматозоиды
 Инфузория Oxytricha trifallax с хорошо заметными ресничками.
Инфузория Oxytricha trifallax с хорошо заметными ресничками. Амеба с проглоченными диатомовыми водорослями
Амеба с проглоченными диатомовыми водорослями






 Более 10 000 морских видов относятся к веслоногим ракообразным , маленьким, часто микроскопическим ракообразным.
Более 10 000 морских видов относятся к веслоногим ракообразным , маленьким, часто микроскопическим ракообразным. Темнопольная фотография гастротриха . длиной 0,06–3,0 мм, червеобразного животного, живущего между частицами осадка
Темнопольная фотография гастротриха . длиной 0,06–3,0 мм, червеобразного животного, живущего между частицами осадка Панцирный Pliciloricus enigmaticus длиной около 0,2 мм обитает в пространствах между морским гравием.
Панцирный Pliciloricus enigmaticus длиной около 0,2 мм обитает в пространствах между морским гравием.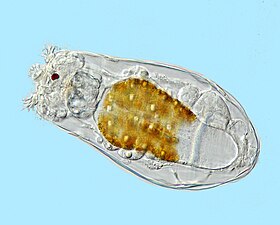 Коловушки , обычно длиной 0,1–0,5 мм, могут выглядеть как протисты, но являются многоклеточными и принадлежат к отряду Animalia.
Коловушки , обычно длиной 0,1–0,5 мм, могут выглядеть как протисты, но являются многоклеточными и принадлежат к отряду Animalia. Тихоходки (водяные медведи) длиной около 0,5 мм являются одними из самых выносливых известных животных.
Тихоходки (водяные медведи) длиной около 0,5 мм являются одними из самых выносливых известных животных.





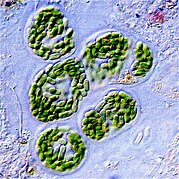
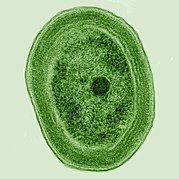
 Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
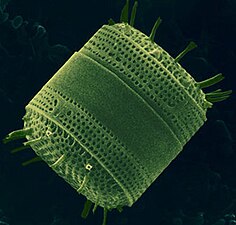 Центрическая диатомовая водоросль
Центрическая диатомовая водоросль Динофлагелляты
Динофлагелляты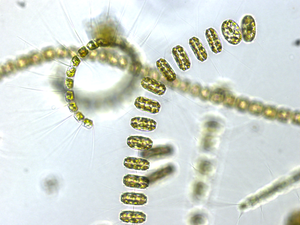 Колониальные цепи водорослей
Колониальные цепи водорослей






 Фитопланктон — основа пищевой цепи океана.
Фитопланктон — основа пищевой цепи океана. Они бывают разных форм и размеров.
Они бывают разных форм и размеров. Колониальный фитопланктон
Колониальный фитопланктон Цианобактерия Prochromococcus обеспечивает большую часть первичной продукции океана.
Цианобактерия Prochromococcus обеспечивает большую часть первичной продукции океана. Зеленые цианобактерии выброшены на скалу в Калифорнии.
Зеленые цианобактерии выброшены на скалу в Калифорнии.






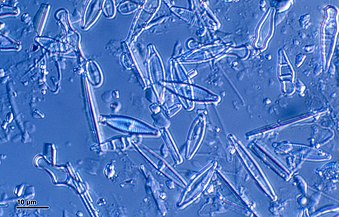 Диатомеи – один из наиболее распространенных видов фитопланктона.
Диатомеи – один из наиболее распространенных видов фитопланктона. Их защитные оболочки (фрустлы) изготовлены из кремния.
Их защитные оболочки (фрустлы) изготовлены из кремния.
 Они бывают разных форм и размеров
Они бывают разных форм и размеров



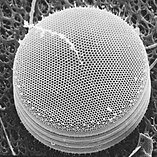
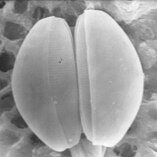
 Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [161]
Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [161] Ископаемая диатомовая водоросль
Ископаемая диатомовая водоросль Существует более 100 000 видов диатомовых водорослей , на долю которых приходится 50% первичной продукции океана.
Существует более 100 000 видов диатомовых водорослей , на долю которых приходится 50% первичной продукции океана.


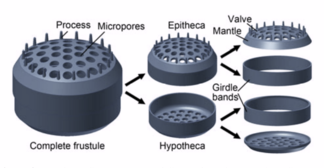
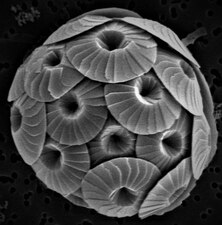 У кокколитофоров есть пластинки, называемые кокколитами.
У кокколитофоров есть пластинки, называемые кокколитами. Coccolithus pelagicus подвид. браарудии
Coccolithus pelagicus подвид. браарудии
 Кокколитофор . Emiliania huxleyi
Кокколитофор . Emiliania huxleyi Цветение водорослей Emiliania huxleyi у южного побережья Англии.
Цветение водорослей Emiliania huxleyi у южного побережья Англии.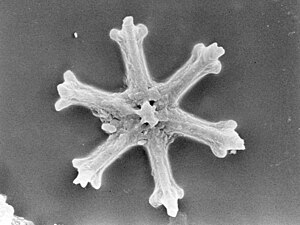 Вымершее ископаемое
Вымершее ископаемое





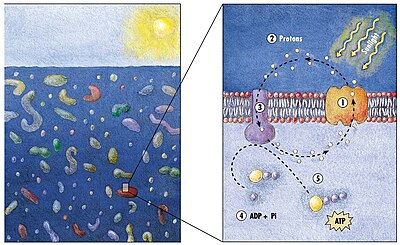

 Копеподы питаются фитопланктоном. Этот несет яйца.
Копеподы питаются фитопланктоном. Этот несет яйца. Тинтинниды инфузории Фавелла
Тинтинниды инфузории Фавелла

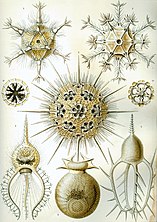
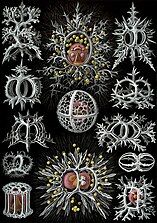
 Как и диатомеи, радиолярии бывают разных форм.
Как и диатомеи, радиолярии бывают разных форм. Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката. Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов
Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов Схематическая диаграмма сферической раковины радиолярий в разрезе
Схематическая диаграмма сферической раковины радиолярий в разрезе Кладококк абиетинус
Кладококк абиетинус




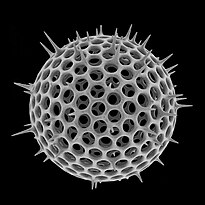




 разрез, показывающий камеры спирального отверстия
разрез, показывающий камеры спирального отверстия Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи
Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи Группа планктонных форм.
Группа планктонных форм. Ископаемые образования нуммулитид разных размеров из эоцена.
Ископаемые образования нуммулитид разных размеров из эоцена.![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[188]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)




![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[188]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)
 Раковинная амеба , Cyphoderia sp.
Раковинная амеба , Cyphoderia sp.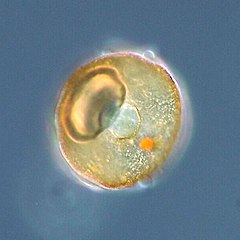
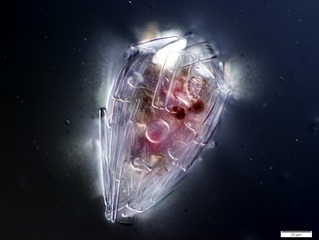 Ксеногенная раковинная амеба, покрытая диатомовыми водорослями (из коллекции амеб Пенарда )
Ксеногенная раковинная амеба, покрытая диатомовыми водорослями (из коллекции амеб Пенарда ) Голая амеба, Chaos sp.
Голая амеба, Chaos sp. Обнаженная амеба с пищевыми вакуолями и проглоченной диатомовой водорослью.
Обнаженная амеба с пищевыми вакуолями и проглоченной диатомовой водорослью.




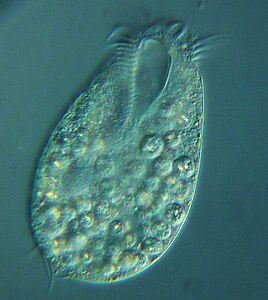

 голофиры Яйцо
голофиры Яйцо![Mesodinium rubrum производит темно-красные цветы, используя порабощенные хлоропласты из водорослей-жертвы [191]](//upload.wikimedia.org/wikipedia/commons/thumb/d/df/Mesodinium_rubrum.jpg/300px-Mesodinium_rubrum.jpg) Mesodinium Rubrum дает темно-красные цветы, используя порабощенные хлоропласты из водорослей-жертв. [191]
Mesodinium Rubrum дает темно-красные цветы, используя порабощенные хлоропласты из водорослей-жертв. [191]


![Mesodinium rubrum производит темно-красные цветы, используя порабощенные хлоропласты из водорослей-жертвы [191]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/df/Mesodinium_rubrum.jpg/300px-Mesodinium_rubrum.jpg)

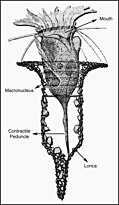
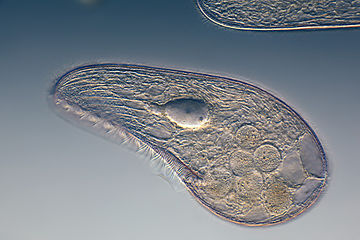







 Euglena mutabilis — фотосинтезирующий жгутиконосец .
Euglena mutabilis — фотосинтезирующий жгутиконосец . Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .
Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .

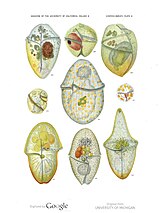

 Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря.
Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря. Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.
Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи. Насселлярии радиолярии могут находиться в симбиозе с динофлагеллятами.
Насселлярии радиолярии могут находиться в симбиозе с динофлагеллятами. Динофлагеллята Dinophys acuta.
Динофлагеллята Dinophys acuta.




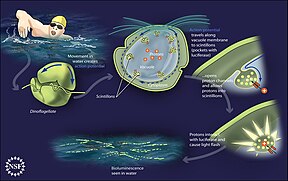


 Tripos muelleri можно узнать по рогам U-образной формы.
Tripos muelleri можно узнать по рогам U-образной формы.
![Karenia brevis вызывает красные приливы, крайне токсичные для человека[214]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/214px-Karenia_brevis.jpg) Karenia brevis вызывает красные приливы, очень токсичные для человека [214]
Karenia brevis вызывает красные приливы, очень токсичные для человека [214]
 Noctiluca scintillans — биолюминесцентная динофлагеллята. [215]
Noctiluca scintillans — биолюминесцентная динофлагеллята. [215]

![Karenia brevis вызывает красные приливы, крайне токсичные для человека[214]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/214px-Karenia_brevis.jpg)










 Кизельгур — это мягкая кремнистая состоящая осадочная порода, из микрофоссилий в форме панцирей ( раковин) одноклеточных диатомей (нажмите, чтобы увеличить).
Кизельгур — это мягкая кремнистая состоящая осадочная порода, из микрофоссилий в форме панцирей ( раковин) одноклеточных диатомей (нажмите, чтобы увеличить). Известковые микрофоссилии из морских отложений, состоящие в основном из звездчатых дисков с примесью кокколитов.
Известковые микрофоссилии из морских отложений, состоящие в основном из звездчатых дисков с примесью кокколитов. Раковины ( раковины ), обычно состоящие из карбоната кальция, из ила фораминифер на глубоком дне океана.
Раковины ( раковины ), обычно состоящие из карбоната кальция, из ила фораминифер на глубоком дне океана. Иллюстрация Глобигерины ила
Иллюстрация Глобигерины ила![Мрамор может содержать протистские микрофоссилии фораминифер, кокколитофоров, известковый наннопланктон и водоросли, остракоды, птероподы, кальпионеллиды и мшанки[222].](//upload.wikimedia.org/wikipedia/commons/thumb/6/61/MarmoCipollino_FustoBasMassenzioRoma.jpg/320px-MarmoCipollino_FustoBasMassenzioRoma.jpg) Мрамор может содержать протистские микрофоссилии фораминифер, кокколитофоров, известковый наннопланктон и водоросли, остракоды , птероподы , кальпионеллиды и мшанки. [222]
Мрамор может содержать протистские микрофоссилии фораминифер, кокколитофоров, известковый наннопланктон и водоросли, остракоды , птероподы , кальпионеллиды и мшанки. [222] Опал может содержать протистские микрофоссилии диатомей, радиолярий, силикофлагеллят и эбридианов. [222]
Опал может содержать протистские микрофоссилии диатомей, радиолярий, силикофлагеллят и эбридианов. [222]




![Мрамор может содержать протистские микрофоссилии фораминифер, кокколитофоров, известковый наннопланктон и водоросли, остракоды, птероподы, кальпионеллиды и мшанки[222].](http://upload.wikimedia.org/wikipedia/commons/thumb/6/61/MarmoCipollino_FustoBasMassenzioRoma.jpg/320px-MarmoCipollino_FustoBasMassenzioRoma.jpg)




 Распределение типов отложений на морском дне
Распределение типов отложений на морском дне



 Elphidium - широко распространенный многочисленный род донных форамов.
Elphidium - широко распространенный многочисленный род донных форамов. Heterohelix — вымерший род донных форамов.
Heterohelix — вымерший род донных форамов.

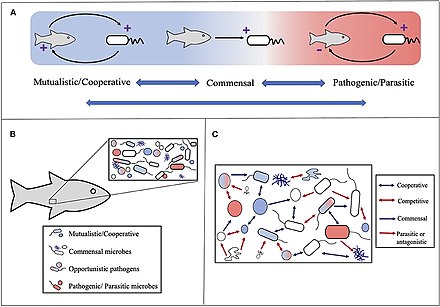
![Коралловый холобионт[240]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/26/Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg/237px-Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg)
![Губка холобионт[241]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6b/The_sponge_holobiont.webp/304px-The_sponge_holobiont.webp.png)
![Голобионт морских водорослей[242]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Processes_within_the_seagrass_holobiont.webp/197px-Processes_within_the_seagrass_holobiont.webp.png)
![Изменение климата и родолитовый голобионт[243]](http://upload.wikimedia.org/wikipedia/commons/thumb/f/f7/Climate_change_stressors_and_rhodolith_holobiont_fitness.webp/415px-Climate_change_stressors_and_rhodolith_holobiont_fitness.webp.png)
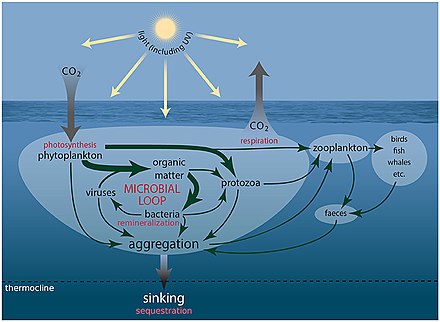

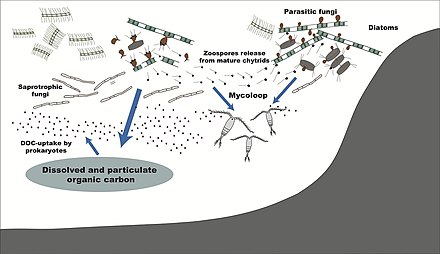

 Pelagibacter ubique , самая распространенная бактерия в океане, играет важную роль в глобальном углеродном цикле .
Pelagibacter ubique , самая распространенная бактерия в океане, играет важную роль в глобальном углеродном цикле .![Морской снег – это дождь органических частиц, который падает из верхних вод в глубины океана.[248] Он является крупным экспортером углерода.](//upload.wikimedia.org/wikipedia/commons/thumb/5/59/Marine_snow.jpg/270px-Marine_snow.jpg) Морской снег — это дождь органических частиц, падающий из верхних вод в глубины океана. [248] Это крупный экспортер углерода.
Морской снег — это дождь органических частиц, падающий из верхних вод в глубины океана. [248] Это крупный экспортер углерода.
![Морской снег – это дождь органических частиц, который падает из верхних вод в глубины океана.[248] Он является крупным экспортером углерода.](http://upload.wikimedia.org/wikipedia/commons/thumb/5/59/Marine_snow.jpg/270px-Marine_snow.jpg)




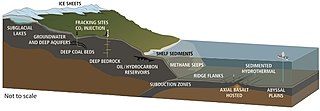
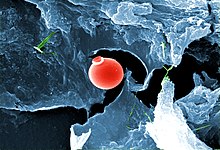
![Оценки количества видов микробов в трех сферах жизни. Бактерии являются самой старой и наиболее биоразнообразной группой, за ней следуют археи и грибы (самые последние группы). В 1998 году, еще до того, как стало известно о масштабах микробной жизни, Роберт М. Мэй[274] подсчитал, что на планете существует 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон[275] подсчитали, что количество видов микроорганизмов может достигать 1 триллиона.[276]](//upload.wikimedia.org/wikipedia/commons/thumb/b/bc/Microbial_species_present_in_the_three_domains_of_life.png/889px-Microbial_species_present_in_the_three_domains_of_life.png) Оценки количества видов микробов в трех сферах жизниБактерии — самая старая и наиболее биоразнообразная группа, за ней следуют археи и грибы (самые новые группы). В 1998 году, еще до того, как стало известно о масштабах микробной жизни, Роберт М. Мэй [274] По оценкам, на планете существовало 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [275] По оценкам, количество видов микроорганизмов может достигать 1 триллиона. [276]
Оценки количества видов микробов в трех сферах жизниБактерии — самая старая и наиболее биоразнообразная группа, за ней следуют археи и грибы (самые новые группы). В 1998 году, еще до того, как стало известно о масштабах микробной жизни, Роберт М. Мэй [274] По оценкам, на планете существовало 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [275] По оценкам, количество видов микроорганизмов может достигать 1 триллиона. [276]![Микробное разнообразие Сравнительное представление известного и оцененного (маленькая рамка) и еще неизвестного (большая рамка) микробного разнообразия, которое применимо как к морским, так и к наземным микроорганизмам. Текстовые поля относятся к факторам, которые отрицательно влияют на знания о микробном разнообразии, существующем на планете.[276]](//upload.wikimedia.org/wikipedia/commons/thumb/1/16/Known%2C_estimated_and_unknown_microbial_diversity.webp/661px-Known%2C_estimated_and_unknown_microbial_diversity.webp.png) Микробное разнообразиеСравнительное представление известного и оцененного (маленькая рамка) и еще неизвестного (большая рамка) микробного разнообразия, которое применимо как к морским, так и к наземным микроорганизмам. Текстовые поля относятся к факторам, которые отрицательно влияют на знания о микробном разнообразии, существующем на планете. [276]
Микробное разнообразиеСравнительное представление известного и оцененного (маленькая рамка) и еще неизвестного (большая рамка) микробного разнообразия, которое применимо как к морским, так и к наземным микроорганизмам. Текстовые поля относятся к факторам, которые отрицательно влияют на знания о микробном разнообразии, существующем на планете. [276]![Оценки количества видов микробов в трех сферах жизни. Бактерии являются самой старой и наиболее биоразнообразной группой, за ней следуют археи и грибы (самые последние группы). В 1998 году, еще до того, как стало известно о масштабах микробной жизни, Роберт М. Мэй[274] подсчитал, что на планете существует 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон[275] подсчитали, что количество видов микроорганизмов может достигать 1 триллиона.[276]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bc/Microbial_species_present_in_the_three_domains_of_life.png/889px-Microbial_species_present_in_the_three_domains_of_life.png)
![Микробное разнообразие Сравнительное представление известного и оцененного (маленькая рамка) и еще неизвестного (большая рамка) микробного разнообразия, которое применимо как к морским, так и к наземным микроорганизмам. Текстовые поля относятся к факторам, которые отрицательно влияют на знания о микробном разнообразии, существующем на планете.[276]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/16/Known%2C_estimated_and_unknown_microbial_diversity.webp/661px-Known%2C_estimated_and_unknown_microbial_diversity.webp.png)

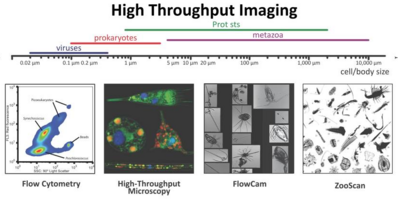
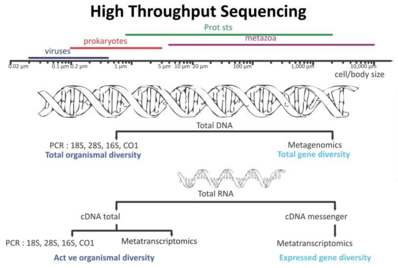


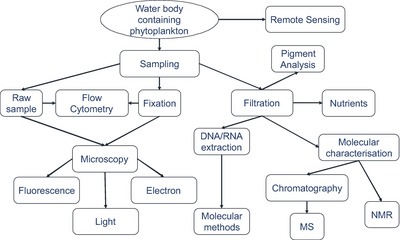
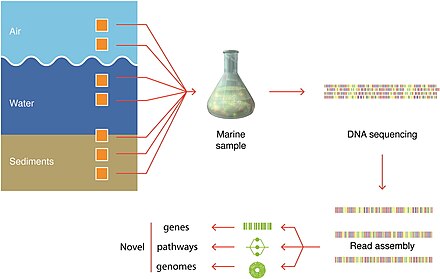
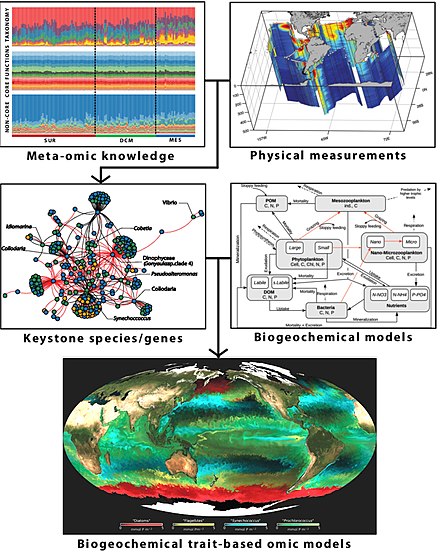

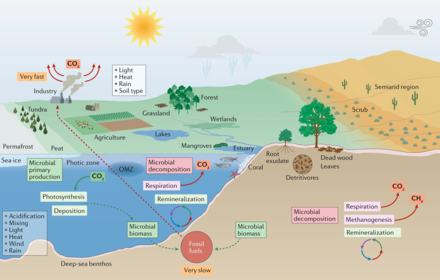
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .  В данную статью включен текст из этого источника, находящегося в свободном доступе .
В данную статью включен текст из этого источника, находящегося в свободном доступе . 